Die Apicomplexa is 'n subfilum van die eensellige eukariote. Hulle is amper geheel parasitêr en die meeste van hulle is bekender onder die naam sporosoë.
Hulle het 'n kenmerkende struktuur wat mikroskopies waargeneem kan word. Hulle apikale kompleks aan een einde van die organisme het organelle wat mikroneme, roptrië en poolringe genoem word. Die poolringe bestaan uit mikrotubule en sommige spesie het 'n konoïed daartussen. Hierdie strukture speel 'n belangrike rol wanneer die organisme sy gasheer se selle binnedring.
Die lewensiklus van die apicomplexa bestaan uit drie fases: sporogonie, merogonie en gametogonie.[1]
Die Apicomplexa is 'n subfilum van die eensellige eukariote. Hulle is amper geheel parasitêr en die meeste van hulle is bekender onder die naam sporosoë.

Hulle het 'n kenmerkende struktuur wat mikroskopies waargeneem kan word. Hulle apikale kompleks aan een einde van die organisme het organelle wat mikroneme, roptrië en poolringe genoem word. Die poolringe bestaan uit mikrotubule en sommige spesie het 'n konoïed daartussen. Hierdie strukture speel 'n belangrike rol wanneer die organisme sy gasheer se selle binnedring.
Die lewensiklus van die apicomplexa bestaan uit drie fases: sporogonie, merogonie en gametogonie.
in die sporogoniefasie vorm die sigote die sporosoïete in die merogoniefase vorm die sporosoïete die merosoïete en vermeerder julle on die gemetogoniefase vorm die merosoïete die geslagtelike gamete wat versmelt om sigote te vormSporlular və ya Apikomplekslər (lat. Apicomplexa) — İbtidailərin (lat. Alveolata) qrupuna daxil olan tipi.
Bu tipin əsas əlaməti qeyri-cinsi çoxalma, cinsi çoxalma və spor əmələgətirmənin növbələşməsidir. Bu tipə daxil olan ibtidailərin hamısı parazit həyat keçirirlər. Əksər halda insan və heyvanlarda ağır xəstəliklər törədir. Tip 2 sinfə bölünür: koksidikimilər və qreqarinlər.
Sporlular tipinə daxil olan bütün nümayəndələr parazit həyat tərzi keçirir. Bunların mürəkkəb həyat dövriyyəsində xüsusi rüşeym mərhələsi - spor vardır. Spor daxilində bir və ya bir neçə rüşeym ola bilər. İbtidailərin digər parazit qruplarından fərqli olaraq, sporlularda həzm orqanellaları - ağız və həzm vakuolu olmur. Osmotik yolla qidalanırlar. Çoxalmaları bəzilərində yalnız cinsiyyətli yolla (qreqarinlərdə), ya da cinsiyyətli və qeyri-cinsiyyətli nəslin bir-birilə növbələşməsi yolu ilə (koksidilərdə və qan sporlularında) olur. Sporlular hüceyrədaxili parazit olub, çoxhüceyrəlilərin bütün siniflərində, xüsusilə həlqəvi qurdlarda, ilbizlərdə, buğumayaqlılarda və onurğalılarda daha çox təsadüf edilərək, özlərinin həyat fəaliyyəti zamanı ağır xəstəliklər (koksidioz, insanda malyariya, iribuynuzlu heyvanlarda piroplazmoz) törədir. Parazit ibtidailərin həyat dövriyyəsində 3 mərhələ ayırd edilir: I - vegetativ forma; 2 - qeyri-cinsiyyətli çoxalma; 3 - cinsiyyətli çoxalma. Bunlarda vegetativ forma - trofozoit, şizont isə şizoqoniya adlanan qeyri-cinsiyyətli çoxalmaya hazırlıq mərhələsidir. Şizoqoniya nəticəsində qız hüceyrə formaları əmələ gəlir və bunlar merozoit adlanır. Cinsiyyətli çoxalma (qametoqoniya) zamanı isə əvvəlcə tam yetişməyən cinsiyyət hüceyrələri (qametositlər) əmələ gəlir, sonra daşıdıqları funksiyadan asılı olaraq onlar arasında diferensiasiya gedir və mikroqametosit (erkək qamet) və makroqametosit (dişi qamet) formalaşır. Parazit sonrakı inkişafını əsas sahibin bədənində davam etdirir. Erkək və dişi qametlər birləşir və ziqot əmələ gəlir.
Sporlular və ya Apikomplekslər (lat. Apicomplexa) — İbtidailərin (lat. Alveolata) qrupuna daxil olan tipi.
Bu tipin əsas əlaməti qeyri-cinsi çoxalma, cinsi çoxalma və spor əmələgətirmənin növbələşməsidir. Bu tipə daxil olan ibtidailərin hamısı parazit həyat keçirirlər. Əksər halda insan və heyvanlarda ağır xəstəliklər törədir. Tip 2 sinfə bölünür: koksidikimilər və qreqarinlər.
Sporlular tipinə daxil olan bütün nümayəndələr parazit həyat tərzi keçirir. Bunların mürəkkəb həyat dövriyyəsində xüsusi rüşeym mərhələsi - spor vardır. Spor daxilində bir və ya bir neçə rüşeym ola bilər. İbtidailərin digər parazit qruplarından fərqli olaraq, sporlularda həzm orqanellaları - ağız və həzm vakuolu olmur. Osmotik yolla qidalanırlar. Çoxalmaları bəzilərində yalnız cinsiyyətli yolla (qreqarinlərdə), ya da cinsiyyətli və qeyri-cinsiyyətli nəslin bir-birilə növbələşməsi yolu ilə (koksidilərdə və qan sporlularında) olur. Sporlular hüceyrədaxili parazit olub, çoxhüceyrəlilərin bütün siniflərində, xüsusilə həlqəvi qurdlarda, ilbizlərdə, buğumayaqlılarda və onurğalılarda daha çox təsadüf edilərək, özlərinin həyat fəaliyyəti zamanı ağır xəstəliklər (koksidioz, insanda malyariya, iribuynuzlu heyvanlarda piroplazmoz) törədir. Parazit ibtidailərin həyat dövriyyəsində 3 mərhələ ayırd edilir: I - vegetativ forma; 2 - qeyri-cinsiyyətli çoxalma; 3 - cinsiyyətli çoxalma. Bunlarda vegetativ forma - trofozoit, şizont isə şizoqoniya adlanan qeyri-cinsiyyətli çoxalmaya hazırlıq mərhələsidir. Şizoqoniya nəticəsində qız hüceyrə formaları əmələ gəlir və bunlar merozoit adlanır. Cinsiyyətli çoxalma (qametoqoniya) zamanı isə əvvəlcə tam yetişməyən cinsiyyət hüceyrələri (qametositlər) əmələ gəlir, sonra daşıdıqları funksiyadan asılı olaraq onlar arasında diferensiasiya gedir və mikroqametosit (erkək qamet) və makroqametosit (dişi qamet) formalaşır. Parazit sonrakı inkişafını əsas sahibin bədənində davam etdirir. Erkək və dişi qametlər birləşir və ziqot əmələ gəlir.
Els apicomplexos (Apicomplexa) són un extens grup de cromalveolats caracteritzat per la presència d'un orgànul únic anomenat complex apical. Són unicel·lulars, formen espores i són exclusivament paràsits d'animals. Les estructures mòbils tals com els flagels o pseudòpodes estan absents excepte en certes etapes dels gàmetes. És un grup divers incloent a organismes com Coccidiasina, Gregarinasina, Piroplasmida, Haemogregarina i Plasmodiidae. Algunes malalties causades per aquests organismes són:
La majoria dels seus membres té un cicle vital complex, implicant reproducció asexual i sexual. Normalment un hostatger es contamina ingerint quists, que es divideixen per produir els esporozous que entren en les cèl·lules. Les cèl·lules rebenten, alliberant merozoits que infecten noves cèl·lules. Això pot ocórrer diverses vegades, fins que produeixen gametocits, que donen lloc a gàmetes que es fusionen per crear quists nous. Hi ha moltes variacions en aquest patró bàsic, però, i molts apicomplexos tenen més d'un hoste.
El complex apical és un orgànul situat en una punta de la cèl·lula que inclou les vesícules anomenades roptries i micronemes, que s'obren en la part anterior de la cèl·lula. Aquestes secreten els enzims que permeten al paràsit entrar en altres cèl·lules. L'extremitat està envoltada per una banda de microtúbuls, anomenada anillo polar, i entre els Conoidasida hi ha també con truncat (embut) de fibril·les anomenat conoide. Sobre la resta de la cèl·lula, a excepció d'una petita boca anomenada microporus, la membrana es recolza en les vesícules anomenades alvèols, formant una estructura semirígida.
La presència d'alvèols i d'altres característiques col·loquen els apicomplexos en el grup dels alveolats. Diversos flagel·lats relacionats com els Perkinsus i Colpodella tenen estructures similars a l'anell polar i varen ser inclosos abans en aquest grup, però ara sembla que són parents més propers als dinoflagel·lats. Són probablement similars a l'avantpassat comú d'ambdós grups.
Una altra semblança amb els dinoflagel·lats, és que les cèl·lules dels apicomplexos contenen un sol plast, anomenat apicoplast, envoltat per tres o quatre membranes. Es pensa que la seva funció inclou la síntesi de lípids, que sembla que és necessària per a la seva supervivència. Es considera generalment que comparteixen un origen comú amb els cloroplasts dels dinoflagel·lats, encara que alguns estudis suggereixen que es deriven en última instància d'algues verdes en comptes de les algues vermelles.
Els apicomplexos abasten un grup d'organismes que abans eren classificats com a esporozous, paràsits sense flagels, pseudòpodes ni cilis. La major part dels apicomplexos són mòbils, però.
El grup dels apicomplexos inclou molts patògens importants per als éssers humans i animals domèstics. En contraste amb els bacteris patògens, aquests paràsits són eucarionts i comparteixen moltes rutes metabòliques amb els seus hostatgers. Això fa que el tractament terapèutic pugui arribar a ser extremadament difícil (una droga que faci mal al paràsit probablement també farà mal a l'hostatger). No hi ha actualment vacuna o tractament eficaços o disponibles per a la majoria de les malalties causades per aquests paràsits. La investigació biomèdica sobre els paràsits és difícil, ja que a vegades és impossible mantenir cultius vius del paràsit en el laboratori i manipular aquests organismes. Recentment, diverses espècies s'han seleccionat per seqüenciar el seu genoma. La disponibilitat d'aquestes seqüències genòmiques proporciona una nova oportunitat per aprendre més sobre l'evolució i la capacitat bioquímica d'aquests paràsits.
Els apicomplexos (Apicomplexa) són un extens grup de cromalveolats caracteritzat per la presència d'un orgànul únic anomenat complex apical. Són unicel·lulars, formen espores i són exclusivament paràsits d'animals. Les estructures mòbils tals com els flagels o pseudòpodes estan absents excepte en certes etapes dels gàmetes. És un grup divers incloent a organismes com Coccidiasina, Gregarinasina, Piroplasmida, Haemogregarina i Plasmodiidae. Algunes malalties causades per aquests organismes són:
Malària (Plasmodium) Babesiosi (Babesia) Coccidiosi, incloent: Criptosporidiosi (Cryptosporidium parvum) Ciclosporosi (Cyclospora cayetanensis) Toxoplasmosi (Toxoplasma gondii) Isosporiasi (Isospora belli)La majoria dels seus membres té un cicle vital complex, implicant reproducció asexual i sexual. Normalment un hostatger es contamina ingerint quists, que es divideixen per produir els esporozous que entren en les cèl·lules. Les cèl·lules rebenten, alliberant merozoits que infecten noves cèl·lules. Això pot ocórrer diverses vegades, fins que produeixen gametocits, que donen lloc a gàmetes que es fusionen per crear quists nous. Hi ha moltes variacions en aquest patró bàsic, però, i molts apicomplexos tenen més d'un hoste.
El complex apical és un orgànul situat en una punta de la cèl·lula que inclou les vesícules anomenades roptries i micronemes, que s'obren en la part anterior de la cèl·lula. Aquestes secreten els enzims que permeten al paràsit entrar en altres cèl·lules. L'extremitat està envoltada per una banda de microtúbuls, anomenada anillo polar, i entre els Conoidasida hi ha també con truncat (embut) de fibril·les anomenat conoide. Sobre la resta de la cèl·lula, a excepció d'una petita boca anomenada microporus, la membrana es recolza en les vesícules anomenades alvèols, formant una estructura semirígida.
La presència d'alvèols i d'altres característiques col·loquen els apicomplexos en el grup dels alveolats. Diversos flagel·lats relacionats com els Perkinsus i Colpodella tenen estructures similars a l'anell polar i varen ser inclosos abans en aquest grup, però ara sembla que són parents més propers als dinoflagel·lats. Són probablement similars a l'avantpassat comú d'ambdós grups.
Una altra semblança amb els dinoflagel·lats, és que les cèl·lules dels apicomplexos contenen un sol plast, anomenat apicoplast, envoltat per tres o quatre membranes. Es pensa que la seva funció inclou la síntesi de lípids, que sembla que és necessària per a la seva supervivència. Es considera generalment que comparteixen un origen comú amb els cloroplasts dels dinoflagel·lats, encara que alguns estudis suggereixen que es deriven en última instància d'algues verdes en comptes de les algues vermelles.
Els apicomplexos abasten un grup d'organismes que abans eren classificats com a esporozous, paràsits sense flagels, pseudòpodes ni cilis. La major part dels apicomplexos són mòbils, però.
El grup dels apicomplexos inclou molts patògens importants per als éssers humans i animals domèstics. En contraste amb els bacteris patògens, aquests paràsits són eucarionts i comparteixen moltes rutes metabòliques amb els seus hostatgers. Això fa que el tractament terapèutic pugui arribar a ser extremadament difícil (una droga que faci mal al paràsit probablement també farà mal a l'hostatger). No hi ha actualment vacuna o tractament eficaços o disponibles per a la majoria de les malalties causades per aquests paràsits. La investigació biomèdica sobre els paràsits és difícil, ja que a vegades és impossible mantenir cultius vius del paràsit en el laboratori i manipular aquests organismes. Recentment, diverses espècies s'han seleccionat per seqüenciar el seu genoma. La disponibilitat d'aquestes seqüències genòmiques proporciona una nova oportunitat per aprendre més sobre l'evolució i la capacitat bioquímica d'aquests paràsits.
Výtrusovci (Apicomplexa) je skupina jednobuněčných alveolátních organismů. Dříve byla řazena do říše prvoků (Protozoa), dnes (podle použitého systému) mezi Chromalveolata resp. Chromista. Výtrusovci dosahují velikosti 0,004–0,04mm. Jsou to výhradně nitrobuněční (intracelulární) parazité, nebo parazité mimobuněční (extracelulární). Tělo je tvořeno jedinou buňkou se systémem organel apikálního komplexu, kryje ho pelikula, která se skládá z vnější a vnitřní membrány. Potrava je přijímána pasivně - pomocí osmózy, nebo pinocytózou. Vylučování probíhá povrchem těla, dýchání má formu anaerobní. Pohyb obstarává buď posuv podélných lišt na povrchu těla, nebo soustava myofibril umožňující kontrakci buňky a její klouzavý pohyb.
Výtrusovci mají složité vývojové cykly, střídá se u nich pohlavní a nepohlavní rozmnožování. Nejprve u nich dochází k rozdělení jedince - sporozoita, na množství dceřiných jedinců (tento jev se nazývá schizogonie, nebo také merogonie). Toto se opakuje 2–3krát, následuje zrací dělení buněk (meióza), tvorba gamet, jejich splynutím vznikne oocysta (tento jev se nazývá gametogonie), cyklus uzavírá nepohlavní namnožení uvnitř oocysty (sporogonie). U třídy výtrusovců - hromadinek (Gregarinia) sporozoit roste v tzv. trofozoit.
Výtrusovci (Apicomplexa) je skupina jednobuněčných alveolátních organismů. Dříve byla řazena do říše prvoků (Protozoa), dnes (podle použitého systému) mezi Chromalveolata resp. Chromista. Výtrusovci dosahují velikosti 0,004–0,04mm. Jsou to výhradně nitrobuněční (intracelulární) parazité, nebo parazité mimobuněční (extracelulární). Tělo je tvořeno jedinou buňkou se systémem organel apikálního komplexu, kryje ho pelikula, která se skládá z vnější a vnitřní membrány. Potrava je přijímána pasivně - pomocí osmózy, nebo pinocytózou. Vylučování probíhá povrchem těla, dýchání má formu anaerobní. Pohyb obstarává buď posuv podélných lišt na povrchu těla, nebo soustava myofibril umožňující kontrakci buňky a její klouzavý pohyb.
Die Apicomplexa (von lat. apex ‚Spitze‘) sind einzellige, zu den Eukaryoten gehörige Parasiten, die infektiöse Sporozysten oder Oozysten produzieren und einen charakteristischen Generationswechsel durchlaufen, bei dem sich geschlechtliche und ungeschlechtlich vermehrende Zellen abwechseln.
Bekannte Vertreter sind die Malaria-Erreger der Gattung Plasmodium sowie Toxoplasma gondii, der Erreger der Toxoplasmose.
Charakteristische Merkmale der Apicomplexa sind:
Die Apicomplexa besitzen einen spezialisierten Plastid, den Apicoplasten. Im Gegensatz zu den Plastiden der höheren Pflanzen und Grünalgen besitzt das Organell zwei zusätzliche Hüllmembranen, womit sich eine Gesamtzahl von vier Hüllmembranen ergibt. Es wird angenommen, dass sich der Plastid aus einer Rotalge entwickelt hat, die im Zuge einer sekundären Endosymbiose durch eine andere Zelle aufgenommen wurde. So stammen die beiden inneren Hüllmembranen vom ursprünglichen Plastiden der Rotalge, und jeweils eine Membran ist aus den Zellmembranen der Rotalge und des Wirtes hervorgegangen.
Proteine, die im Apicoplast lokalisiert sind, besitzen eigene zweigeteilte Signalpeptide, die den Transport über die Zellmembran sichern. Der Apicoplast besitzt ein eigenes Plastiden-Genom, welches dem pflanzlicher Plastiden ähnelt. Unter anderem wurden Enzyme gefunden, die üblicherweise in Pflanzen Teil des Photosynthese-Apparates sind, in den Apicoplasten aber andere Aufgaben besitzen, denn die Apicomplexa betreiben als obligate Endoparasiten keine Photosynthese. Ein Beispiel dafür ist die Ferredoxin-NADPH-Reduktase, die in Pflanzen Teil des Photosystems I ist.
Obwohl die Apicomplexa die Fähigkeit zur Photosynthese verloren haben, ist der rudimentäre Plastid, der Apicoplast, zum Überleben notwendig.[2]
Die meisten Apicomplexa sind haploid. Oft wird als ungeschlechtliche Vermehrungsphase zunächst eine Schizogonie (Syn. Merogonie, Zerfallsteilung) durchlaufen, bei der ein mehrkerniges Stadium (der Schizont/Meront) sich in unterschiedlich viele Zellen (die Merozoiten) teilt. Aus einigen Merozoiten können Gamonten und später Gameten entstehen, die während der Gamogonie (Gametogamie) zu einer Zygote verschmelzen. Aus der Zygote entstehen, nachdem die Meiose und oftmals eine weitere ungeschlechtliche Vermehrung (Sporogonie) vollzogen wurde, infektiöse Sporozoiten, die noch in Zystenhüllen stecken können.
Es können bei den Apicomplexa ein oder mehrere Wirte auftreten, die sich auf unterschiedliche Weise infizieren können. So werden z. B. die Oozysten oft mit dem Kot ausgeschieden und können durch eine Schmutz- und Schmierinfektion in einen neuen Wirt gelangen. Viele Sporozoen bilden in Zwischenwirten auch Dauerstadien in einer Cystenhülle aus, die von einem fleischfressenden Endwirt beim Verzehr von rohem Fleisch aufgenommen werden können. Beim Malaria-Erreger werden die Sporozoiten von Mücken beim Stechakt übertragen.
Adl et al. (2012) unterteilen die Apicomplexa in folgende Gruppen:[3]
Die Apicomplexa (von lat. apex ‚Spitze‘) sind einzellige, zu den Eukaryoten gehörige Parasiten, die infektiöse Sporozysten oder Oozysten produzieren und einen charakteristischen Generationswechsel durchlaufen, bei dem sich geschlechtliche und ungeschlechtlich vermehrende Zellen abwechseln.
Bekannte Vertreter sind die Malaria-Erreger der Gattung Plasmodium sowie Toxoplasma gondii, der Erreger der Toxoplasmose.
Apicomplexa ili Apicomplexia (lat. apex = gore + complexus = kompleks – odnosi se na skup organela u sporozoitu) je veliko koljeno parazitskih Alveolata. Većina njih ima jedinstven oblik organela sa plastidima zvanim apikoplasti i apikalnu složenu strukturu. Organela je adaptacija koju apikoplekse koriste pri penetraciji u ćeliju domaćina.
Apicomplexa su jednoćelijska i sporne. Sve su vrste obligatne endoparazitske životinje, osim u rodu Nephromyces, simbionti morskih životinja, prvobitno klasificiran kao gljive. Pokretne strukture, kao što su bičevi ili pseudopodje prisutne su samo u određenim fazama gameta.[1] except Nephromyces, a symbiont in marine animals, originally classified as a chytrid fungus.[2] Apicomplexa su raznolika grupa koja uključuje organizme poput kokcidija, gregarina, piroplazme, hemogregarina i plazmoji.
Apicomplexa izazivaju bolesti, kao što su:
Apicomplexa obuhvata većinu grupe koja se nekada zvala Sporozoa, grupu parazitskih "protozoa" ", uglavnom bez bičeva, treplji ili pseudopodija. Međutim, većina je pokretna upotrebom klizajućih mehanizama, koji počiva na statičkoj adheziji malih motornih miozina. Ostale glavne linije bile su Ascetosporea (sada u Rhizaria), Myxozoa (sada je poznato da su visoko izvedene knidarijske životinje), i Microsporidia (za koji se sada zna da potiču od gljiva). Naziv Sporozoa uzima se ponekad kao sinonim za Apicomplexa, ili povremeno kao podskup.[4]
Koljeno Apicomplexa sadrži sve eukariote sa grupom struktura i organela koje se zajedno nazivaju apikalni kompleks. Ovaj kompleks sastoji se od strukturnih komponenti i sekrecijske organele koje su potrebne za invaziju na domaćinske ćelije tokom parazitskih faza životnog ciklusa. Apicomplexa imaju složene životne cikluse koji uključuju nekoliko faza i obično prolaze i aseksualne jedinke i spolnu replikaciju. Sve Apicomplexa su, u nekom dijelu životnog ciklusa obligatni paraziti, pri čemu neki parazitiraju na dva odvojena domaćina u aseksualnim i seksualnim fazama.
Pored sačuvanog apikalnog kompleksa, Apicomplexa su i morfološki raznoliki. Različiti organizmi unutar Apicomplexa, kao i različiti životni stadiji određenih apikompleksa, mogu značajno varirati u veličini, obliku i subćelijskoj strukturi. Kao i drugi eukarioti, Apicomplexa imaju jedro , endoplazmatski retikulum i Golgijev kompleks. Obično imaju jednu mitohondriju, kao i drugi izvedeni endosimbionati organel zvanu apikoplast koja ima zasebni 35-kilobazni kružni genom (s izuzetkom vrsta roda Cryptosporidium i Gregarina niphandrodes, kojima nedostaje apikoplast). Svi članovi ovog tipa imaju infektivni stadij – sporozoit – koji u apikalnom kompleksu ima tri različite strukture. Apikalni kompleks sastoji se od skupa spiralno raspoređenih mikrotubula s ( konoidom), sekretorna tijela i jedan ili više polarni prstenova. Mogu imati i dodatna vitka sekretorna tijela koja su gusta od elektrona (mikroneme), okružena sa jednim ili dva polarna prstena. Ova struktura daje naziv taksonu. Daljnja skupina sfernih organela raspoređena je u cijeloj ćeliji, a nije lokalizirana u apikalnom kompleksu i poznate su kao guste granule. Obično imaju srednji promjer oko 0,7 µm. Izlučivanje sadržaja gustih granul događa se nakon invazije parazita i lokalizacije unutar parazitoforne vakuole i traje nekoliko minuta.
Replikacija:
Mobilnost: Apikompleksa imaju jedinstvenu sposobnost klizanja koja im omogućava prolazak kroz tkiva i ulaz u i napuštanje ćelija domaćina. Ova sposobnost omogućena je primjenom adhezija i malih statičkih motornih miozina. Ostale karakteristike koje su zajedničke ovom tipu su nedostatak cilija, seksualna reprodukcija, upotreba mikropora za uzimanje hrane i proizvodnja oocista koje sadrže sporozoite kao zarazni oblik. Čini se da su u ovom tipu transpozoni rijetki, ali su identificirani u rodovima Ascogregarina i Eimeria.
Većina predstavnika apikompleks ima složen životni ciklus, koji uključuje i aseksualnu i seksualnu reprodukciju. Obično se domaćin inficira aktivnom invazijom parazita (slično [[entoza|entozi]), koji se dijele i proizvode sporozoite koji ulaze u njegove ćelije. Na kraju se ćelije rasprsnu, oslobađajući merozoite, koji inficiraju nove ćelije. To se može dogoditi nekoliko puta, sve dok se ne naprave gamonti, koji formiraju gamete koje se spajaju kako bi stvorili nove ciste. Međutim, u ovom osnovnom obrascu pojavljuju se mnoge varijacije, a mnogi Apicomplexa imaju više od jednog domaćina.
Apikalni kompleks uključuje vezikule nazvane roptrije i mikroneme, koji se otvaraju u prednjem dijelu ćelije. Sekretini enzimi omogućavaju parazitu da uđe u druge ćelije. Vrh je okružen trakom mikrotubula, nazvanom polarnim prstenom, a među konoidima je i lijevak tubulinskih proteina koji se nazivaju konoidi. Preko ostatka ćelije, osim umanjenih usta zvanih mikropore, membrana se oslanja na vezikule zvane alveole, tvoreći polukrutu pelikulu.
Prisustvo alveola i drugih osobina svrstava Apicomplexa u grupu koja se zove alveolata. Nekoliko srodnih alveola, poput Perkinsus i Colpodella, imaju strukture slične polarnom prstenu i ranije su bile ovdje uključene, ali čini se da su većina bliži srodnici dinoflagelata . Vjerovatno su slične zajedničkom pretku dviju skupina.
Još jedna sličnost je da mnoge ćelije apikopleksa sadrže jedan plastid, zvani apikoplast, okružen s tri ili četiri membrane. Smatra se da njegove funkcije uključuju zadatke poput biosinteze lipida i hema, a čini se da im je to potrebno za opstanak. Općenito, smatra se da plastidi imaju zajedničko porijeklo s hloroplastima dinoflagelata, a dokazi upućuju na porijeklo iz crvenih, a ne iz zelenih algi. [5][6]
Apikompleksa imaju vrste tri grupe – kokcidijska, gregarinska i hemosporidna. Izgleda da su kokcidi i gregarini srodnički povezani. Perkinsus, iako se nekad smatrao članom Apicomplexa, premješten je u novi takson – Perkinsozoa. [7]
Gregarine su obično paraziti anelida, artropoda i školjki. Često se nalaze u crijevima svojih domaćina, ali mogu upasti i u ostala tkiva. U tipskom životnom ciklusu gregarina, unutar ćelije domaćina, trofozoit se razvija u shizont. Zatim se shizogonijom dijeli na merozoite. Merozoiti se oslobađaju razgradnjom ćelije domaćina i upada u ostale ćelije. U nekom dijelu životnog ciklusa apikompleksa formiraju se gametociti. Oni se oslobađaju razgradnjom ćelija domaćina, koje se grupiraju. Svaki gametocit formira više gameta. Gume se stapaju s drugom i tvore oociste. Oociste preuzimaju domaćini da ga preuzmu novi domaćini.
Kokcidije su općenito paraziti kičmenjaka s. Kao i gregarine, oni su obično paraziti epitelnih ćelija crijeva, ali mogu zaraziti i druga tkiva.
Životni ciklus kokcija uključuje meronije, gametogonije i sporogonije. Iako je sličan onom gregarinama, razlikuje se u formiranju zigota. Neki trofozoiti se povećavaju i postaju makrogameti, dok se drugi opetovano dijele i tvore mikrogamete (anizogamija). Mikrogameti su pokretni i mgu doći do makrogamatima da ih oplode. Oplođeni makrogamet formira zigot koji se pretvara u oocistu koja se normalno oslobađa iz tijela. Shizigija, kada se pojavi, uključuje izrazito neskladne gamete. Životni ciklus je obično sa haploidima, s jedinom diploidnom fazom u zigotu, a koja je obično kratkotrajna.
Glavna razlika između kokcidija i gregarina je u gamantima. Kod kokcidija su to male, unutarćelijske i bez epimera ili mikrokorona. U gregarinima su to velike, vanćelijske, sa epimeritima ili mikronima. Druga razlika između kokcidija i gregarina također je u gamontima. U kokcidijama pojedinačni gamont postaje makroametocit, dok u gregarina gamonti stvaraju višestruke gametonite.
Hemosporidije imaju složenije životne cikluse, koji se razlikuju između člankonožaca i domaćina kičmenjaka. Trofozoit parazitira u eritrocitima ili drugim tkivima kičmenjaka domaćina. Mikrogameti i makrogameti se uvijek nalaze u krvi. Gamete preuzima vektorski insekt tokom sisanja krvi. Mikrogamete migriraju u crijevima insekata i spajaju se s makrogametima. Oplođeni makrogamat tada postaje ookineta koja prodire u tijelo vektora. Ookinete se zatim pretvaraju u oociste i dijele se u početku mejotski, a zatim mitozom (haplontni životni cikluso) da bi se stvorio sporozoit. Sporozoiti bježe iz oociste i migriraju unutar tijela vektora do pljuvačnih žlijezda gdje se ubrizgavaju u organizam novog kičmenjaka domaćina kada se vektor insekata ponovno hrani.
Mnogi paraziti apikompleksa važni su uzročnici bolesti ljudi i domaćih životinja. Za razliku od patogenih bakterija, apikoplekski paraziti su eukariotski i dijele mnoge metaboličke puteve sa svojim životinjskim domaćinima. Zbog toga je terapijski ciljni tretman izuzetno težak - lijek koji šteti apikopleksnom parazitu vjerovatno će štetiti i čovjeku domaćinu. Dosad nije bila dostupna efikasna vakcina za većinu bolesti izazvanih tim parazitima. Biomedicinska istraživanja ovih parazita izazovna su jer je često teško, ako ne i nemoguće, održavati žive kulture parazita u laboratoriji i genetički manipulirati tim organizmima. Posljednjih godina nekoliko vrsta apikopleksa odabrano je za sekvenciranje genoma. Dostupnost genomskih sekvenci pruža novu priliku naučnicima da saznaju više o evoluciji i biohemijskim osobenostima ovih parazita. Pretežni izvor tih genskih informacija je baza EuPathDB Plasmodium. Baza podataka koja uključuje eksperimentalne i računarske podatke za kokcidije (ToxoDB), piroplazme (PiroplasmaDB) i vrsta roda Cryptosporidium (CryptoDB). Jedan od mogućih ciljeva za pripravljanje lijekove je da djeluje na plastid, a čini se da postojeći lijekovi poput tetraciklina, koji su efikasni protiv apikopleksa, djeluju i protiv plastida.
Mnogi kokcidiomorfi imaju srednjeg domaćina, kao i primarnog domaćina, a evolucija izbora domaćina odvijala se na različite načine i u različitim vremenima u tim skupinama. Za neke kokcidiomorfe izvorni domaćin postao je posredni domaćin, dok je za druge postao konačni. U rodovima Aggregata, Atoxoplasma, Cystoisospora, Schellackia i Toxoplasma, original je sada definitivan, dok je u rodovima Akiba , Babesiosoma, Babesia , [ [Haemogregarina]], Haemoproteus, Hepatozoon, Kariolysus, Leucocytozoon , Plasmodium, Sarcocystis i Theileria, izvorni domaćini su sada posredni.
Slične strategije za povećanje vjerovatnoće prenošenja razvijale su se u više rodova. Ciste polienergidnih oocista i tkiva nalaze se u predstavnicima redova Protococcidiorida i Eimeriida. Hipnozoit se nalaze u Karyolysus lacerata i u većini vrsta plazmodijum; transovarijalni prijenos parazita događa se u životnim ciklusima rodova Karyolyzus i Babesia.
Pretpostavka je da se horizontalni transfer gena u evoluciji ovog koljena dogodio relativno rano, prenošenjem histona H4 u lizinu 20 (H4K20), kao modifikator KMT5A ( Set8), od životinje domaćina do pretka apikopleksana. Drugi gen – H3K36 metiltransferaza (Ashr3) – može se prenijeti i horizontalno.
Unutar Apicomplexa nalaze se paraziti:
Unutar grupe Adelorine nalaze se vrste koje inficiraju beskičmenjake i druge koje inficiraju kičmenjake. Eimeriorina – najveći podred u ovom tipu – životni ciklus uključuje i seksualnu i aseksualnu fazu. Aseksualne faze reproduciraju se shizogonijom. Muški gametocit stvara veliki broj gameta, a zigot stvara oocist, što je infektivni stadij. Većina je monoksena (inficira samo jednog domaćina), ali nekoliko ih je heteroksenih (životni ciklus uključuje dva ili više domaćina). Još uvijek raspravlja se o broju porodica u ovom podredu, pri čemu je broj porodica između jedne i 20, ovisno o autorima, a broj rodova između 19 i 25.
Svi članovi ovog koljena parazitiraju i razvijaju se od slobodnoživećeg pretka . Pretpostavlja se da je ovaj način života evoluirao u vrijeme divergencije dinoflagelata i apikompleksa..[8][9] Procjenjuje se da je njihova dalja evolucija trajala oko 800 miliona godina.[10] Smatra se da su, od postojećih kladusa, najstariji arhigregarini.[11] Ti filogenetski odnosi rijetko su proučavani na nivou podrazreda. Haemosporidije su povezane sa gregarinama, a piroplazme i kokcidije su sestrinske grupe. Čini se da su Haemosporidia i piroplazma sestrinski kladusi i više su vezani za kokcidije nego za gregarine.[12]
ApicomplexaGregarines
CoccidiaJednodomaćinske kokcidije (Eimeria, Isospora, Cyclospora)
Incistirajuće kokcidije (Toxoplasma, Sarcocystis, Frenkellia)
HematozoaPiroplazme (Babesia, Theileria)
Hemosporidia (Plasmodium, Leucocytozoon)
Prve Apicomplexa vidio je Antonie van Leeuwenhoek, koji je 1674. godine u žučnom mjehuru zeca ugledao vjerovatno oociste "Eimeria stiedae". Dufour je 1828. godine imenovao prvu vrstu koljena, "Gregarina ovata" "u crijevima uholadže. On je mislio da su oni posebna grupa povezana sa trematodama, u to vrijeme uključenim u Vermes. Od tada ih je mnogo više identificirano i imenovano. Tokom 1826-1850. imenovane su 41 vrste i šest rodova Apicomplexa. U periodu 1951–1975 dodane su 1873 nove vrste i 83 nova roda.
Stariji takson Sporozoa, uključen u Protozoa, stvorio je Leuckart 1879. Kroz historiju su sa sadašnjim Apicomplexa grupirale mnoge nepovezane grupe. Naprimjer, Kudo (1954) u sporozoa uključuje vrste grupe Ascetosporea (Rhizaria), Microsporidia (Fungi), Myxozoa (Animalia) i Helicosporidium (Chlorophyta), dok je Zierdt (1978.) uključio rod Blastocystis (Stramenopiles). Dermocystidium se također smatran sporozoom. Nisu sve ove grupe imale spore, ali sve su bile parazitske. Međutim, i drugi parazitski ili simbiotski jednoćelijski organizmi,ako su imali bičeve, također su bili uključeni u protozojske skupine izvan Sporozoa (Flagellata, Ciliophora i Sarcodina) (npr., mnoge Kinetoplastida, Retortamonadida, Diplomonadida, Trichomonadida, Hypermastigida), cilija (npr. Balantidium) ili pseudopodije (npr. Entamoeba, Acanthamoeba,Naegleria). Ako imaju ćelijske zidove, takođe bi mogli biti uključene u biljno carstvo između bakterija ili kvasca.
Sporozoa se više ne smatra biološki valjanim taksonom i upotreba tog termina je obeshrabrena, iako ga neki autori još uvijek koriste kao sinonim za Apicomplexa. U novije vreme su druge grupe bile isključene iz Apicomplexa, npr. Perkinsus i Colpodella (sada u Protalveolati). Polje razvrstavanja Apicomplexa je u toku i klasifikacija se mijenjala tokom svih godina otkako je 1970. formalno imenovani takson.
Do 1987. godine dovršeno je sveobuhvatno istraživanje ovog taksona: ukupno je imenovano 4516 vrsta i 339 rodova. Sastojali su se od:
Iako je izvršena znatna revizija ovog taksona (pored Haemosporidia sada ima 17 rodova, a ne 9), ovi brojevi su verovatno još uvek približno tačni.
Razni plazmodiji
Izumirli: Paleohaimatus calabresi
Cista Eimeria maxima (Coccidiasina)
Cista Sarcocystis hominis (Coccidiasina)
Apicomplexa atawa sakapeung sok disebut Sporozoa (basa Latin, spore = siki, zoa = sato) nyaéta kelompok protista unisélulér nu siklus hirupna miboga wangun siga spora. Apicomplexa teu mibanda alat gerak. Sakabéh apicomplexa hirup minangka parasit dina awak sato jeung manusa.[3]
Cara réproduksi apicomplexa nyaéta sacara aséksual jeung séksual. Gantina sistem aséksual jeung séksualna kompléks, kalawan robahna bisa sababaraha kali ganti bentuk sarta kudu boga dua atau leuwih indung inang. Réproduksi aséksual dilakonan ku cara meulah binér. Sedengkeun réproduksi séksual dilakonan ku ngawangun gamet tuluy diteruskeun ku ngahijina gamet jalu jeung bikang. Conto apicomplexa nyaéta Toxoplasma gondii anu ngalantarankeun panyakit toksoplasmosis jeung Plasmodium anu ngalantarankeun panyakit malaria di manusa.[4]
Toxoplasma gondii asup ka awak manusa ngaliwatan kadaharan, misalna daging nu katepaan kista Toxoplasma ti féses ucing. Inféksi Toxoplasma utama ngabahayakeun pikeun indung nu keur reuneuh sabab bisa maéhan émbrio atawa orok nu dibabarkeun bisa cacat. Plasmodium asup ka awak manusa ngaliwatan reungit Anopheles bikang. Di jero awak manusa, plasmodium nyerang sél-sél haté jeung sél-sél getih (éritrosit). Aya opat rupa plasmodium anu bisa ngalantarankeun panyakit malaria nyaéta Plasmodium vivax, Plasmodium ovale, Plasmodium malariae, jeung Plasmodium falciparum. Plasmodium vivax jeung Plasmodium ovale ngalantarankeun panyakit malaria tertiana, Plasmodium malariae ngalantarankeun panyakit malaria kuartana, jeung Plasmodium falciparum ngalantarankeun panyakit malaria anu pangbahayana nyaéta malaria tropikana[3]
Plasmodium vivax jeung Plasmodium ovale bisa hirup sanajan teu aktif di jero sél haté anu kajangkit malaria mangbulan-bulan nepi ka taunan. Balukarna, di hiji mangsa panyakit malaria bisa kambuh deui. Cara megatkeun siklus hirupna nyaéta ku cara nyempitkeun daérah hirupna nyaéta cai. Boh nguras cai atawa nutup penampungan cai.[3]
De Apicomplexa sünd ene groote Grupp vun Protozoa, wat teö't Riek vun de Protisten höörn. Dat sünd keene Deeren oder Planten. Dat Toxoplasma gondii höört to de Apicomplexa. Malaria is ok vun de Apicomplexa. Dat meeste vun de Parasiten, de lang Sporozoa nöömt warrn, ward vandage unner de Apicomplexa rekent.
Apicomplexa es un phylo de Alveolata, Protista.
Apicomplexa ili Apicomplexia (lat. apex = gore + complexus = kompleks – odnosi se na skup organela u sporozoitu) je veliko koljeno parazitskih Alveolata. Većina njih ima jedinstven oblik organela sa plastidima zvanim apikoplasti i apikalnu složenu strukturu. Organela je adaptacija koju apikoplekse koriste pri penetraciji u ćeliju domaćina.
Apicomplexa su jednoćelijska i sporne. Sve su vrste obligatne endoparazitske životinje, osim u rodu Nephromyces, simbionti morskih životinja, prvobitno klasificiran kao gljive. Pokretne strukture, kao što su bičevi ili pseudopodje prisutne su samo u određenim fazama gameta. except Nephromyces, a symbiont in marine animals, originally classified as a chytrid fungus. Apicomplexa su raznolika grupa koja uključuje organizme poput kokcidija, gregarina, piroplazme, hemogregarina i plazmoji.
Apicomplexa izazivaju bolesti, kao što su:
Babesioza (Babesia) Malarija (Plasmodium) Kriptosporidioza (Cryptosporidium parvum) Ciklosporijaza (Cyclospora cayetanensis) Cistoizosporijaza (Cystoisospora belli (bivša "Isospora Belli")) Toksoplasmoza (Toxoplasma gondii).Apicomplexa obuhvata većinu grupe koja se nekada zvala Sporozoa, grupu parazitskih "protozoa" ", uglavnom bez bičeva, treplji ili pseudopodija. Međutim, većina je pokretna upotrebom klizajućih mehanizama, koji počiva na statičkoj adheziji malih motornih miozina. Ostale glavne linije bile su Ascetosporea (sada u Rhizaria), Myxozoa (sada je poznato da su visoko izvedene knidarijske životinje), i Microsporidia (za koji se sada zna da potiču od gljiva). Naziv Sporozoa uzima se ponekad kao sinonim za Apicomplexa, ili povremeno kao podskup.
Apicomplexa atawa sakapeung sok disebut Sporozoa (basa Latin, spore = siki, zoa = sato) nyaéta kelompok protista unisélulér nu siklus hirupna miboga wangun siga spora. Apicomplexa teu mibanda alat gerak. Sakabéh apicomplexa hirup minangka parasit dina awak sato jeung manusa.
Sporozoa ya iku filum kalima saka Krajan Protista, kang uga dikenal kanthi jeneng Apicomplexa, ngumpulaké spésies-spésies parasit protozoa intraseluler obligat kang diklasifikasiaké minangka Sporozoa utawa Sporozoans, amarga mbentuk sel-sel reproduksi kang dikenal kanthi jeneng spora.[1]
Sporozoa minangka siji-sijiné anggota Protozoa kang ora nduwéni piranti obah. Sporozoa urip dadi parasit. Cara éntuk pangané kanthi njupuk nutrisi inangé,contoné Plasmodium kang dadi anggota sporozoa kang paling dikenal. Sajeroning awak manungsa, Plasmodium bisa njalari lelara malaria. Nularé saka cokotan lemut Anopheles wédok.[2]
Reproduksi sporozoa kanthi cara aseksual ya iku schizogoni (nyigar awak ing njero awak inang) utawa sporogoni (mbentuk spora ing njero awak inang) lan kanthi cara sèksual ya iku peleburan loro gamet ing njero awak lemut(inang).[3]
Sporozoa ya iku filum kalima saka Krajan Protista, kang uga dikenal kanthi jeneng Apicomplexa, ngumpulaké spésies-spésies parasit protozoa intraseluler obligat kang diklasifikasiaké minangka Sporozoa utawa Sporozoans, amarga mbentuk sel-sel reproduksi kang dikenal kanthi jeneng spora.
Sporozoa minangka siji-sijiné anggota Protozoa kang ora nduwéni piranti obah. Sporozoa urip dadi parasit. Cara éntuk pangané kanthi njupuk nutrisi inangé,contoné Plasmodium kang dadi anggota sporozoa kang paling dikenal. Sajeroning awak manungsa, Plasmodium bisa njalari lelara malaria. Nularé saka cokotan lemut Anopheles wédok.
Reproduksi sporozoa kanthi cara aseksual ya iku schizogoni (nyigar awak ing njero awak inang) utawa sporogoni (mbentuk spora ing njero awak inang) lan kanthi cara sèksual ya iku peleburan loro gamet ing njero awak lemut(inang).
La sporozones (ance clamada Apicomplexa) es un filo grande de protistas parasital. La plu posese un forma unica de organeta cual composa un tipo de plasto nomida un apicoplasto, e un strutur apical complicada. La organeta es un adata cual la sporozon aplica en la penetra de un selula ospitante.
Los es uniselulal e forma sporas. Tota spesies es endoparasitos obligada de animales, estra Nefromises, un simbiosica en animales maral. Strutures de move, como flajelas o pseudopodes, esiste sola en alga pasos de la gametas.
The Apicomplexa are a diverse group that includes organisms such as the coccidia, gregarines, piroplasms, haemogregarines, and plasmodia. Diseases caused by Apicomplexa include:
La sporozones es un grupo diversa cual inclui organismes como la cocsidias, gregarines, piroplasmas, hemogregarines, e plasmodio. Maladias causada par estas inclui:
Апикомплекслар, яки Споровиклар (лат. Apicomplexa) — Protozoa тибы Альвеолят төркеменнән (лат. Alveolata).
Споровиклар паразитлар булып кына яшиләр. Паразитлык итүгә җайлашу аларда бик көчле һәм нык камилләшкән. Аларның күзәнәк төзелеше шул дәрәҗәгә җитеп гадиләшкән, хәтта организм әйләнә-тирәдә ирекле яши алмый: хуҗа организмыннан читтә булганда, споровиклар тышча белән яхшы сакланган тынлык халәтендәге споралар рәвешен алалар. Моннан тыш, споровикларга хуҗаларын алмаштыруга бәйле катлаулы яшәү циклы хас. Аларның хуҗалары — төрле умырткасыз һәм умырткалы хайваннар. Споровикларның берничә төре кешедә паразитлык итә. Мәсәлән, бизгәк плазмодие үсешенең билгеле бер стадияләрендә кеше канының кызыл тәнчекләрендә — эритроцитларда паразитлык итә һәм Урта диңгез буенда, тропикларда таралган җитди авыру — өч көнлек бизгәккә сәбәпче була.
Апикомплекслар, яки Споровиклар (лат. Apicomplexa) — Protozoa тибы Альвеолят төркеменнән (лат. Alveolata).
Споровиклар паразитлар булып кына яшиләр. Паразитлык итүгә җайлашу аларда бик көчле һәм нык камилләшкән. Аларның күзәнәк төзелеше шул дәрәҗәгә җитеп гадиләшкән, хәтта организм әйләнә-тирәдә ирекле яши алмый: хуҗа организмыннан читтә булганда, споровиклар тышча белән яхшы сакланган тынлык халәтендәге споралар рәвешен алалар. Моннан тыш, споровикларга хуҗаларын алмаштыруга бәйле катлаулы яшәү циклы хас. Аларның хуҗалары — төрле умырткасыз һәм умырткалы хайваннар. Споровикларның берничә төре кешедә паразитлык итә. Мәсәлән, бизгәк плазмодие үсешенең билгеле бер стадияләрендә кеше канының кызыл тәнчекләрендә — эритроцитларда паразитлык итә һәм Урта диңгез буенда, тропикларда таралган җитди авыру — өч көнлек бизгәккә сәбәпче була.
The Apicomplexa (also called Apicomplexia) are a large phylum of mainly parasitic alveolates. Most of them possess a unique form of organelle that comprises a type of non-photosynthetic plastid called an apicoplast, and an apical complex structure. The organelle is an adaptation that the apicomplexan applies in penetration of a host cell.
The Apicomplexa are unicellular and spore-forming. Most are obligate endoparasites of animals,[3] except Nephromyces, a symbiont in marine animals, originally classified as a chytrid fungus,[4] and the Chromerida, some of which are photosynthetic partners of corals. Motile structures such as flagella or pseudopods are present only in certain gamete stages.
The Apicomplexa are a diverse group that includes organisms such as the coccidia, gregarines, piroplasms, haemogregarines, and plasmodia. Diseases caused by Apicomplexa include:
The name Apicomplexa derives from two Latin words—apex (top) and complexus (infolds)—for the set of organelles in the sporozoite. The Apicomplexa comprise the bulk of what used to be called the Sporozoa, a group of parasitic protozoans, in general without flagella, cilia, or pseudopods. Most of the Apicomplexa are motile, however, with a gliding mechanism[5] that uses adhesions and small static myosin motors.[6] The other main lines were the Ascetosporea (a group of Rhizaria), the Myxozoa (highly derived cnidarian animals), and the Microsporidia (derived from fungi). Sometimes, the name Sporozoa is taken as a synonym for the Apicomplexa, or occasionally as a subset.
The phylum Apicomplexa contains all eukaryotes with a group of structures and organelles collectively termed the apical complex.[7] This complex consists of structural components and secretory organelles required for invasion of host cells during the parasitic stages of the Apicomplexan life cycle.[7] Apicomplexa have complex life cycles, involving several stages and typically undergoing both asexual and sexual replication.[7] All Apicomplexa are obligate parasites for some portion of their life cycle, with some parasitizing two separate hosts for their asexual and sexual stages.[7]
Besides the conserved apical complex, Apicomplexa are morphologically diverse. Different organisms within Apicomplexa, as well as different life stages for a given apicomplexan, can vary substantially in size, shape, and subcellular structure.[7] Like other eukaryotes, Apicomplexa have a nucleus, endoplasmic reticulum and Golgi complex.[7] Apicomplexa generally have a single mitochondrion, as well as another endosymbiont-derived organelle called the apicoplast which maintains a separate 35 kilobase circular genome (with the exception of Cryptosporidium species and Gregarina niphandrodes which lack an apicoplast).[7]
All members of this phylum have an infectious stage—the sporozoite—which possesses three distinct structures in an apical complex. The apical complex consists of a set of spirally arranged microtubules (the conoid), a secretory body (the rhoptry) and one or more polar rings. Additional slender electron-dense secretory bodies (micronemes) surrounded by one or two polar rings may also be present. This structure gives the phylum its name. A further group of spherical organelles is distributed throughout the cell rather than being localized at the apical complex and are known as the dense granules. These typically have a mean diameter around 0.7 μm. Secretion of the dense-granule content takes place after parasite invasion and localization within the parasitophorous vacuole and persists for several minutes.
Replication:
Mobility:
Apicomplexans have a unique gliding capability which enables them to cross through tissues and enter and leave their host cells. This gliding ability is made possible by the use of adhesions and small static myosin motors.[8]
Other features common to this phylum are a lack of cilia, sexual reproduction, use of micropores for feeding, and the production of oocysts containing sporozoites as the infective form.
Transposons appear to be rare in this phylum, but have been identified in the genera Ascogregarina and Eimeria.[9]

Most members have a complex lifecycle, involving both asexual and sexual reproduction. Typically, a host is infected via an active invasion by the parasites (similar to entosis), which divide to produce sporozoites that enter its cells. Eventually, the cells burst, releasing merozoites, which infect new cells. This may occur several times, until gamonts are produced, forming gametes that fuse to create new cysts. Many variations occur on this basic pattern, however, and many Apicomplexa have more than one host.
The apical complex includes vesicles called rhoptries and micronemes, which open at the anterior of the cell. These secrete enzymes that allow the parasite to enter other cells. The tip is surrounded by a band of microtubules, called the polar ring, and among the Conoidasida is also a funnel of tubulin proteins called the conoid.[10] Over the rest of the cell, except for a diminished mouth called the micropore, the membrane is supported by vesicles called alveoli, forming a semirigid pellicle.[11]
The presence of alveoli and other traits place the Apicomplexa among a group called the alveolates. Several related flagellates, such as Perkinsus and Colpodella, have structures similar to the polar ring and were formerly included here, but most appear to be closer relatives of the dinoflagellates. They are probably similar to the common ancestor of the two groups.[12]
Another similarity is that many apicomplexan cells contain a single plastid, called the apicoplast, surrounded by either three or four membranes. Its functions are thought to include tasks such as lipid and heme biosynthesis, and it appears to be necessary for survival. In general, plastids are considered to have a common origin with the chloroplasts of dinoflagellates, and evidence points to an origin from red algae rather than green.[13][14]
Within this phylum are four groups — coccidians, gregarines, haemosporidians (or haematozoans, including in addition piroplasms), and marosporidians. The coccidians and haematozoans appear to be relatively closely related.[15]
Perkinsus , while once considered a member of the Apicomplexa, has been moved to a new phylum — Perkinsozoa.[16]
The gregarines are generally parasites of annelids, arthropods, and molluscs. They are often found in the guts of their hosts, but may invade the other tissues. In the typical gregarine lifecycle, a trophozoite develops within a host cell into a schizont. This then divides into a number of merozoites by schizogony. The merozoites are released by lysing the host cell, which in turn invade other cells. At some point in the apicomplexan lifecycle, gametocytes are formed. These are released by lysis of the host cells, which group together. Each gametocyte forms multiple gametes. The gametes fuse with another to form oocysts. The oocysts leave the host to be taken up by a new host.[17]

In general, coccidians are parasites of vertebrates. Like gregarines, they are commonly parasites of the epithelial cells of the gut, but may infect other tissues.
The coccidian lifecycle involves merogony, gametogony, and sporogony. While similar to that of the gregarines it differs in zygote formation. Some trophozoites enlarge and become macrogamete, whereas others divide repeatedly to form microgametes (anisogamy). The microgametes are motile and must reach the macrogamete to fertilize it. The fertilized macrogamete forms a zygote that in its turn forms an oocyst that is normally released from the body. Syzygy, when it occurs, involves markedly anisogamous gametes. The lifecycle is typically haploid, with the only diploid stage occurring in the zygote, which is normally short-lived.[18]
The main difference between the coccidians and the gregarines is in the gamonts. In the coccidia, these are small, intracellular, and without epimerites or mucrons. In the gregarines, these are large, extracellular, and possess epimerites or mucrons. A second difference between the coccidia and the gregarines also lies in the gamonts. In the coccidians, a single gamont becomes a macrogametocyte, whereas in the gregarines, the gamonts give rise to multiple gametocytes.[19]

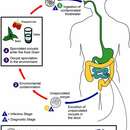
The Haemosporidia have more complex lifecycles that alternate between an arthropod and a vertebrate host. The trophozoite parasitises erythrocytes or other tissues in the vertebrate host. Microgametes and macrogametes are always found in the blood. The gametes are taken up by the insect vector during a blood meal. The microgametes migrate within the gut of the insect vector and fuse with the macrogametes. The fertilized macrogamete now becomes an ookinete, which penetrates the body of the vector. The ookinete then transforms into an oocyst and divides initially by meiosis and then by mitosis (haplontic lifecycle) to give rise to the sporozoites. The sporozoites escape from the oocyst and migrate within the body of the vector to the salivary glands where they are injected into the new vertebrate host when the insect vector feeds again.[20]
The class Marosporida Mathur, Kristmundsson, Gestal, Freeman, and Keeling 2020 is a newly recognized lineage of apicomplexans that is sister to the Coccidia and Hematozoa. It is defined as a phylogenetic clade containing Aggregata octopiana Frenzel 1885, Merocystis kathae Dakin, 1911 (both Aggregatidae, originally coccidians), Rhytidocystis sp. 1 and Rhytidocystis sp. 2 Janouškovec et al. 2019 (Rhytidocystidae Levine, 1979, originally coccidians, Agamococcidiorida), and Margolisiella islandica Kristmundsson et al. 2011 (closely related to Rhytidocystidae). Marosporida infect marine invertebrates. Members of this clade retain plastid genomes and the canonical apicomplexan plastid metabolism. However, marosporidians have the most reduced apicoplast genomes sequenced to date, lack canonical plastidial RNA polymerase and so provide new insights into reductive organelle evolution.[15]

Many of the apicomplexan parasites are important pathogens of human and domestic animals. In contrast to bacterial pathogens, these apicomplexan parasites are eukaryotic and share many metabolic pathways with their animal hosts. This makes therapeutic target development extremely difficult – a drug that harms an apicomplexan parasite is also likely to harm its human host. At present, no effective vaccines are available for most diseases caused by these parasites. Biomedical research on these parasites is challenging because it is often difficult, if not impossible, to maintain live parasite cultures in the laboratory and to genetically manipulate these organisms. In recent years, several of the apicomplexan species have been selected for genome sequencing. The availability of genome sequences provides a new opportunity for scientists to learn more about the evolution and biochemical capacity of these parasites. The predominant source of this genomic information is the EuPathDB[21] family of websites, which currently provides specialised services for Plasmodium species (PlasmoDB),[22][23] coccidians (ToxoDB),[24][25] piroplasms (PiroplasmaDB),[26] and Cryptosporidium species (CryptoDB).[27][28] One possible target for drugs is the plastid, and in fact existing drugs such as tetracyclines, which are effective against apicomplexans, seem to operate against the plastid.[29]
Many Coccidiomorpha have an intermediate host, as well as a primary host, and the evolution of hosts proceeded in different ways and at different times in these groups. For some coccidiomorphs, the original host has become the intermediate host, whereas in others it has become the definitive host. In the genera Aggregata, Atoxoplasma, Cystoisospora, Schellackia, and Toxoplasma, the original is now definitive, whereas in Akiba, Babesiosoma, Babesia, Haemogregarina, Haemoproteus, Hepatozoon, Karyolysus, Leucocytozoon, Plasmodium, Sarcocystis, and Theileria, the original hosts are now intermediate.
Similar strategies to increase the likelihood of transmission have evolved in multiple genera. Polyenergid oocysts and tissue cysts are found in representatives of the orders Protococcidiorida and Eimeriida. Hypnozoites are found in Karyolysus lacerate and most species of Plasmodium; transovarial transmission of parasites occurs in lifecycles of Karyolysus and Babesia.
Horizontal gene transfer appears to have occurred early on in this phylum's evolution with the transfer of a histone H4 lysine 20 (H4K20) modifier, KMT5A (Set8), from an animal host to the ancestor of apicomplexans.[30] A second gene—H3K36 methyltransferase (Ashr3 in plants)—may have also been horizontally transferred.[31]
Within the Apicomplexa are three suborders of parasites: [32]
Within the Adelorina are species that infect invertebrates and others that infect vertebrates. The Eimeriorina—the largest suborder in this phylum—the lifecycle involves both sexual and asexual stages. The asexual stages reproduce by schizogony. The male gametocyte produces a large number of gametes and the zygote gives rise to an oocyst, which is the infective stage. The majority are monoxenous (infect one host only), but a few are heteroxenous (lifecycle involves two or more hosts).
The number of families in this later suborder is debated, with the number of families being between one and 20 depending on the authority and the number of genera being between 19 and 25.
The first Apicomplexa protozoan was seen by Antonie van Leeuwenhoek, who in 1674 saw probably oocysts of Eimeria stiedae in the gall bladder of a rabbit. The first species of the phylum to be described, Gregarina ovata in earwigs intestines, was named by Dufour in 1828. He thought that they were a peculiar group related to the trematodes, at that time included in Vermes.[33] Since then, many more have been identified and named. During 1826–1850, 41 species and six genera of Apicomplexa were named. In 1951–1975, 1873 new species and 83 new genera were added.[33]
The older taxon Sporozoa, included in Protozoa, was created by Leuckart in 1879[34] and adopted by Bütschli in 1880.[35] Through history, it grouped with the current Apicomplexa many unrelated groups. For example, Kudo (1954) included in the Sporozoa species of the Ascetosporea (Rhizaria), Microsporidia (Fungi), Myxozoa (Animalia), and Helicosporidium (Chlorophyta), while Zierdt (1978) included the genus Blastocystis (Stramenopiles).[36] Dermocystidium was also thought to be sporozoan. Not all of these groups had spores, but all were parasitic.[33] However, other parasitic or symbiotic unicellular organisms were included too in protozoan groups outside Sporozoa (Flagellata, Ciliophora and Sarcodina), if they had flagella (e.g., many Kinetoplastida, Retortamonadida, Diplomonadida, Trichomonadida, Hypermastigida), cilia (e.g., Balantidium) or pseudopods (e.g., Entamoeba, Acanthamoeba, Naegleria). If they had cell walls, they also could be included in plant kingdom between bacteria or yeasts.
Sporozoa is no longer regarded as biologically valid and its use is discouraged,[37] although some authors still use it as a synonym for the Apicomplexa. More recently, other groups were excluded from Apicomplexa, e.g., Perkinsus and Colpodella (now in Protalveolata).
The field of classifying Apicomplexa is in flux and classification has changed throughout the years since it was formally named in 1970.[1]
By 1987, a comprehensive survey of the phylum was completed: in all, 4516 species and 339 genera had been named. They consisted of:[38][33]
Although considerable revision of this phylum has been done (the order Haemosporidia now has 17 genera rather than 9), these numbers are probably still approximately correct.[39]
Jacques Euzéby in 1988[40] created a new class Haemosporidiasina by merging subclass Piroplasmasina and suborder Haemospororina.
The division into Achromatorida and Chromatorida, although proposed on morphological grounds, may have a biological basis, as the ability to store haemozoin appears to have evolved only once.[41]
Roberts and Janovy in 1996 divided the phylum into the following subclasses and suborders (omitting classes and orders):[42]
These form the following five taxonomic groups:
Perkins et al. proposed the following scheme.[43] It is outdated as the Perkinsidae have since been recognised as a sister group to the dinoflagellates rather that the Apicomplexia:
The name Protospiromonadida has been proposed for the common ancestor of the Gregarinomorpha and Coccidiomorpha.[44]
Another group of organisms that belong in this taxon are the corallicolids.[45] These are found in coral reef gastric cavities. Their relationship to the others in this phylum has yet to be established.
Another genus has been identified - Nephromyces - which appears to be a sister taxon to the Hematozoa.[46] This genus is found in the renal sac of molgulid ascidian tunicates.
Members of this phylum, except for the photosynthetic chromerids,[47] are parasitic and evolved from a free-living ancestor. This lifestyle is presumed to have evolved at the time of the divergence of dinoflagellates and apicomplexans.[48][49] Further evolution of this phylum has been estimated to have occurred about 800 million years ago.[50] The oldest extant clade is thought to be the archigregarines.[48]
These phylogenetic relations have rarely been studied at the subclass level. The Haemosporidia are related to the gregarines, and the piroplasms and coccidians are sister groups.[51] The Haemosporidia and the Piroplasma appear to be sister clades, and are more closely related to the coccidians than to the gregarines.[9] Marosporida is a sister group to Coccidiomorphea.[15]
Myzozoa Apicomplexa s.l.Squirmida (Digyalum, Filipodium, Platyproteum)
Chrompodellids/ApicomonadeaChromerida (Chromera, Vitrella, Piridium)
Voromonadida (Alphamonas, Voromonas)
Apicomplexa s.s. Gregarines s.l.Gregarines s.s.
MarosporidaAggregatidae (Aggregata, Merocystis)
Rhytidocystidae (Rhytidocystis)
Coccidiomorphea CoccidiaCoccidia with a single host (Eimeria, Isospora, Cyclospora)
Cyst-forming coccidia (Toxoplasma, Sarcocystis, Frenkellia)
HematozoaPiroplasms (Babesia, Theileria)
Hemosporidia (Plasmodium, Leucocytozoon)
Janouškovec et al 2015 presents a somewhat different phylogeny, supporting the work of others showing multiple events of plastids losing photosynthesis. More importantly this work provides the first phylogenetic evidence that there have also been multiple events of plastids becoming genome-free.[52]
The Apicomplexa (also called Apicomplexia) are a large phylum of mainly parasitic alveolates. Most of them possess a unique form of organelle that comprises a type of non-photosynthetic plastid called an apicoplast, and an apical complex structure. The organelle is an adaptation that the apicomplexan applies in penetration of a host cell.
The Apicomplexa are unicellular and spore-forming. Most are obligate endoparasites of animals, except Nephromyces, a symbiont in marine animals, originally classified as a chytrid fungus, and the Chromerida, some of which are photosynthetic partners of corals. Motile structures such as flagella or pseudopods are present only in certain gamete stages.
The Apicomplexa are a diverse group that includes organisms such as the coccidia, gregarines, piroplasms, haemogregarines, and plasmodia. Diseases caused by Apicomplexa include:
Babesiosis (Babesia) Malaria (Plasmodium) Cryptosporidiosis (Cryptosporidium parvum) Cyclosporiasis (Cyclospora cayetanensis) Cystoisosporiasis (Cystoisospora belli) Toxoplasmosis (Toxoplasma gondii)The name Apicomplexa derives from two Latin words—apex (top) and complexus (infolds)—for the set of organelles in the sporozoite. The Apicomplexa comprise the bulk of what used to be called the Sporozoa, a group of parasitic protozoans, in general without flagella, cilia, or pseudopods. Most of the Apicomplexa are motile, however, with a gliding mechanism that uses adhesions and small static myosin motors. The other main lines were the Ascetosporea (a group of Rhizaria), the Myxozoa (highly derived cnidarian animals), and the Microsporidia (derived from fungi). Sometimes, the name Sporozoa is taken as a synonym for the Apicomplexa, or occasionally as a subset.
Los apicomplejos (Apicomplexa) son un extenso grupo de protistas exclusivamente endoparásitos de animales,[2] con unas 5000 especies descritas.[3] Se caracterizan por la presencia de un orgánulo único denominado complejo apical, una adaptación del organismo que le permite la entrada en la célula hospedadora. Son unicelulares, forman esporas y carecen de estructuras móviles tales como flagelos o pseudópodos, excepto en ciertas etapas de los gametos. Es un grupo diverso incluyendo a organismos como gregarinas, hemogregarinas, coccidios, hemosporidios (Plasmodium) y piroplasmas (Babesia).[4] Entre las enfermedades causadas por estos organismos están la coccidiosis, malaria y babesiosis.
A excepción de Nephromyces, que es un simbionte de animales marinos,[5] todas las demás especies de apicomplejos son endoparásitos obligados de animales.[6] Entre las enfermedades que causan a los seres humanos destacan:
Todas las especies de este grupo tienen una etapa infecciosa (el esporozoito) que posee un complejo apical, que da nombre al grupo. El complejo apical es un orgánulo situado en una punta de la célula que posee tres estructuras distintivas: un conjunto de microtúbulos dispuestos en espiral (el conoide), un cuerpo secretor (las roptrias) y una o más bandas de microtúbulos (el anillo polar). Puede incluir también vesículas secretoras (las micronemas) rodeadas por uno o dos anillos polares.[4] El complejo apical le permite al organismo la entrada en la célula huésped combinando la acción perforadora del conoide con las secreciones químicas de las roptrias. Además del complejo apical, existen unos orgánulos esféricos secretores situados alrededor de la célula (denominados gránulos densos), que actúan después de la penetración en la célula huésped, persistiendo durante varios minutos. Sobre el resto de la célula, a excepción de una pequeña boca llamada microporo, la membrana se apoya en unas vesículas denominadas alvéolos corticales, formando una estructura semirrígida.
Los flagelos se encuentran sólo en los gametos móviles. Están dirigidos posteriormente y varían en número, por lo general se presentan de uno a tres. Los apicomplejos tienen una capacidad de deslizamiento única que les permite atravesar tejidos y entrar y salir de las células huésped. Esta capacidad de deslizamiento se debe al uso de adherencias y por pequeños motores estáticos de miosina. La mayoría de los apicomplejos tienen un apicoplasto (plástido no fotosintético). Otras características comunes a este filo son la reproducción sexual, el uso de microporos para la alimentación y la producción de ooquistes que contienen esporozoitos.
Más en detalle, la célula de los apicomplejos presenta las siguientes características: el núcleo es haploide, presenta aparato de Golgi, presenta cuerpos basales (aunque hemosporidios y piroplásmidos presentan en sus cuerpos basales tripletes de microtúbulos normales, coccidios y gregarinas tienen nueve singletes), las mitocondrias tienen crestas tubulares, carece de centriolos, cloroplastos, eyectosomas o inclusiones, en algunas especies presenta plástidos incoloros, y la célula está rodeada por una película de tres capas (la estructura alveolar) penetrada por microporos. En cuanto a la replicación: la mitosis es normalmente cerrada, con huso intranuclear, aunque en algunas especies es abierta en los polos, la división celular es generalmente por esquizogonia, y la meiosis se produce en el cigoto.
La mayoría de los apicomplejos tiene un ciclo vital complejo, implicando reproducción asexual y sexual. Típicamente, un huésped se contamina ingiriendo quistes, los cuales se dividen para producir los esporozoitos (fase de esporulación) que entran en sus células. Eventualmente las células revientan, liberando merozoitos (fase de esquizogonia) que infectan nuevas células. Esto puede ocurrir varias veces, hasta que se producen gametocitos (fase de gamogonia), los cuales dan lugar a gametos que se fusionan para crear quistes nuevos y completar el ciclo. Hay muchas variaciones en este patrón básico, sin embargo, y muchos apicomplejos tiene más de un huésped.
Los apicomplejos se dividen en cuatro grupos principales: gregarinas, coccidios, hemosporidios y piroplasmas.[7] Gregarinas y coccidios parecen estar estrechamente relacionados (se agrupan en la clase Conoidasida), lo mismo que hemosporidios y piroplasmas (clase Aconoidasida).
Las gregarinas generalmente son parásitos de los anélidos, artrópodos y moluscos. A menudo se encuentran en los intestinos de sus anfitriones, pero pueden invadir otros tejidos. En el ciclo de vida típico de la gregarina, un trofozoito se desarrolla dentro de la célula huésped produciendo un esquizonte. Este luego se divide por esquizogonia dando lugar a numerosos merozoitos. Los merozoitos se liberan de la célula huésped mediante lisis e invaden nuevas células, proceso que se repite varias veces. En algún momento del ciclo de vida de la gregarina se forman gametocitos. Estos se liberan por lisis de las células huésped que las contienen. Cada gametocito forma múltiples gametos y estos se fusionan por parejas para formar ooquistes. Los ooquistes abandonan el antiguo huésped para buscar uno nuevo.
Los coccidios son, en general, parásitos de vertebrados. Como las gregarinas, normalmente son parásitos de las células epiteliales del intestino, aunque pueden infectar otros tejidos, y además incluyen algunos parásitos sanguíneos. El ciclo de vida de los coccidios implica merogonia, gametogonia y esporogonia. Aunque el ciclo es similar al de las gregarinas, se diferencia en la formación de cigoto. Algunos trofozoítos aumentan de tamaño y se convierten en macrogametos, mientras que otros se dividen varias veces para formar microgametos (anisogamia). Los microgametos son móviles y buscan al macrogameto para fertilizarlo. El macrogameto fertilizado se convierte en cigoto que a su vez forma un ooquiste, que normalmente abandona el huésped. La meiosis, cuando se produce, implica gametos marcadamente anisógamos. El ciclo de vida es típicamente haploide, con una única etapa diploide que ocurre en el cigoto, que normalmente es de corta duración.
Las principales diferencias entre coccidios y gregarinas se dan en los gamontes. En los coccidios los gamontes son pequeños, intracelulares y carecen epimeritos o mucrones, mientras que en las gregarinas son grandes, extracelulares y presentan epimeritos o mucrones. Además, en los coccidios un gamonte se convierte en un único macrogametocito, mientras que en las gregarinas da lugar a múltiples gametocitos.
Los hemosporidios tienen ciclos de vida más complejos que alternan entre un huésped artrópodo y uno vertebrado. Los trofozoitos parasitan eritrocitos u otros tejidos del hospedador vertebrado. Los microgametos y macrogametos siempre se encuentran en la sangre. Los gametos son absorbidos por el insecto vector durante la ingestión de sangre. Los microgametos migran al intestino del insecto y se fusionan con los macrogametos. El macrogameto fertilizado se convierte ahora en un oocineto, que penetra en el cuerpo del vector. El oocineto se transforma a continuación en un ooquiste y se divide inicialmente por meiosis y luego por mitosis (ciclo de vida haploide) para dar lugar a los esporozoitos. Los esporozoitos se liberan del ooquiste y migran a las glándulas salivales del vector, desde donde se inyectan al nuevo huésped vertebrado cuando el insecto lo pica.
Los piroplasmas tienen ciclos de vida muy similares a los hemosporidios, de los que se distinguen por no formar ooquistes ni esporas, además de que en la fase de trofozoito se separan del eritrocito por una sola membrana, al contrario que otros parásitos sanguíneos, que por lo general tienen al menos dos membranas.
Trofozoito de Plasmodium dentro de un eritrocito (Haemosporida)
Dos trofozoitos de Babesia vogeli dentro de un eritrocito (Piroplasmida)
La presencia de alvéolos corticales y otros rasgos colocan a Apicomplexa en el grupo Alveolata. Varios flagelados relacionados, tales como Perkinsus y Colpodella, tienen estructuras similares al anillo polar y fueron incluidos antes en este grupo, pero ahora parece que son parientes más cercanos de Dinoflagellata. Estos organismos son probablemente similares al antepasado común de ambos grupos.
Otra relación con los dinoflagelados viene dada por la presencia en las células de los apicomplejos de un único plasto, denominado apicoplasto, que se encuentra rodeado por tres o cuatro membranas. Se supone que entre sus funciones se incluye la síntesis de lípidos y al parecer es necesario para la supervivencia del organismo. Se considera generalmente que los apicoplastos comparten un origen común con los cloroplastos de los dinoflagelados, aunque algunos estudios sugieren que proceden en última instancia de algas verdes más bien que de algas rojas.[8][9]
Apicomplexa abarca a un grupo de organismos que antes eran clasificados como esporozoos (protozoos parásitos sin flagelos, seudópodos ni cilios). Los otros principales grupos clasificados antes como esporozoos son Ascetosporea (ahora clasificado en Cercozoa), Myxozoa (ahora clasificado entre los animales) y Microsporidia (ahora clasificado entre los hongos). A veces, el término de esporozoo se usa como sinónimo de Apicomplexa y otras veces como subconjunto.
El complejo ciclo de vida de los apicomplejos se entiende mejor en términos de su evolución. Los análisis filogenéticos sugieren que Apicomplexa se originó a partir de Dinoflagellata, un gran grupo de protozoos fotosintéticos. Probablemente los antepasados de los apicomplejos fueron organismos autótrofos que desarrollaron la capacidad de invadir las células intestinales de sus depredadores y posteriormente perdieron la capacidad fotosintética. Algunas especies de dinoflagelados actuales pueden invadir los cuerpos de las medusas y seguir con la fotosíntesis, lo que es posible porque los órganos de las medusas son casi transparentes. En otros organismos con órganos opacos, lo más probable es que la capacidad fotosintética se perdiera rápidamente.
Se cree que los apicomplejos comenzaron infectando la pared intestinal del huésped con una ruta de propagación fecal-oral. En algún punto de la evolución, el parásito evolucionó a la capacidad de infectar el hígado de huésped. Este patrón se observa en el género Cryptosporidium. En algún otro punto de la evolución, un apicomplejo desarrolló la capacidad de infectar a las células de la sangre y de sobrevivir e infectar a los mosquitos. Una vez que la transmisión del mosquito quedó firmemente establecida, se perdió la anterior vía de transmisión fecal-oral. La teoría actual (2007) sugiere que los géneros Plasmodium, Haemoproteus y Hepatocystis evolucionaron a partir de especies de Leukocytozoon. Los parásitos del género Leukocytozoon infectan a los leucocitos (glóbulos blancos) y a las células del hígado y del bazo y se transmiten por medio de moscas negras (Simulium), un extenso género de moscas relacionadas con los mosquitos.
Los leucocitos, la mayoría de los hepatocitos y las células del bazo realizan activamente la fagocitosis de partículas, haciendo la entrada en la célula más fácil para el parásito. El mecanismo de entrada en los eritrocitos de las especies del género Plasmodium está todavía poco clara, teniendo en cuenta que la entrada se realiza en menos de 30 segundos. Todavía no se sabe si este mecanismo evolucionó antes que los mosquitos se convirtieran en los principales vectores de transmisión de Plasmodium.
El patrón de alternancia entre la reproducción sexual y asexual, que puede parecer confusa al principio, es un patrón muy común en las especies parásitas. Las ventajas evolutivas de este tipo de ciclo de vida ya fueron reconocidas por Mendel.
En condiciones favorables, la reproducción asexual es superior a la sexual puesto que el padre está bien adaptado a su medio ambiente y sus descendientes compartirán estos genes. Durante condiciones de estrés, tales como la transferencia a un nuevo huésped, la reproducción sexual es por general superior, ya que produce un barajamiento de los genes que, sobre el promedio de población, da lugar a individuos mejor adaptados al nuevo entorno.
Apicomplexa incluye muchos patógenos importantes para los seres humanos y animales domésticos. En contraste con las bacterias patógenas, estos parásitos son eucariontes y comparten muchas rutas metabólicas con sus huéspedes. Este hecho hace que el tratamiento terapéutico sea extremadamente difícil (una droga que dañe al parásito probablemente también dañará a su huésped). No hay actualmente vacunas eficaces o tratamientos disponibles para la mayoría de las enfermedades causadas por estos parásitos. Un posible objetivo para las drogas es el plástido y, de hecho, los medicamentos existentes que son eficaces contra los apicomplejos, como las tetraciclinas, parecen actuar precisamente contra el plástido.[10]
La investigación biomédica sobre los parásitos es difícil, pues a veces es imposible mantener cultivos vivos del parásito en el laboratorio y manipular estos organismos. Recientemente, varias especies se han seleccionado para secuenciar su genoma. La disponibilidad de estas secuencias genómicas proporciona una nueva oportunidad para aprender más sobre la evolución y la capacidad bioquímica de estos parásitos.
|coautores= (ayuda) Los apicomplejos (Apicomplexa) son un extenso grupo de protistas exclusivamente endoparásitos de animales, con unas 5000 especies descritas. Se caracterizan por la presencia de un orgánulo único denominado complejo apical, una adaptación del organismo que le permite la entrada en la célula hospedadora. Son unicelulares, forman esporas y carecen de estructuras móviles tales como flagelos o pseudópodos, excepto en ciertas etapas de los gametos. Es un grupo diverso incluyendo a organismos como gregarinas, hemogregarinas, coccidios, hemosporidios (Plasmodium) y piroplasmas (Babesia). Entre las enfermedades causadas por estos organismos están la coccidiosis, malaria y babesiosis.
Eosloomad (Sporozoa) oli takson, mis hõlmas kõiki parasiitseid protiste, ning oli seega selgelt polüfüleetiline.
Tänapäeval arvatakse suur osa endistest eosloomade taksonitest hõimkonda Apicomplexa.
Eosloomad (Sporozoa) oli takson, mis hõlmas kõiki parasiitseid protiste, ning oli seega selgelt polüfüleetiline.
Tänapäeval arvatakse suur osa endistest eosloomade taksonitest hõimkonda Apicomplexa.
Itiöeläimet (Apicomplexa) ovat Chromalveolata-kuntaan kuuluva ryhmä yksisoluisia eliöitä. Ne kuuluvat Alveolata-pääjaksoon panssarilevien ja ripsieläinten kanssa. Itiöeläimiä on kuvattu noin 4500 lajia. Itiöeläinten lähimpiä sukulaisia ovat panssarilevät.
Itiöeläimet ovat pitkälle erikoistuneita eläinten solunsisäisiä loisia. Niiden suomenkielinen nimitys johtuu niiden elinkiertoon kuuluvasta itiömäisestä elomuodosta. Tieteellinen nimitys Apicomplexa puolestaan viittaa niiden solun päässä tavattavaan apikaalikompleksi-nimiseen rakenteeseen, jonka avulla eliöt tunkeutuvat isäntäsoluunsa.
Niiden solurakenteessa tavataan myös erikoinen apikoplasti-soluelin, joka on ilmeisesti evoluution kuluessa surkastunut ja lehtivihreänsä menettänyt viherhiukkanen. Apikoplasti tuottaa nykykäsityksen mukaan eräitä itiöeläimen tarvitsemia rasvahappoja. On todettu, että jos apikoplasti tuhotaan, itiöeläin ei kykene infektoimaan uusia isäntäsoluja.
Itiöeläimet aiheuttavat monia sairauksia. Näistä tunnetuimpia ovat:
Itiöeläinten elinkierto on monimutkainen ja siihen kuuluu sekä suvullinen että suvuton lisääntymisvaihe. Tyypillisesti itiöeläimillä suvullisen lisääntymisen tuloksena syntyneet sporotsoiitit tunkeutuvat isäntäsoluihin, joissa ne jakautuvat suvuttomasti ja tuottavat isäntäsolun ulkopuolelle vapautuvia merotsoiitteja. Merotsoiitit infektoivat uusia isäntäsoluja, joista puolestaan vapautuu gametosyyttejä. Gametosyytit kehittyvät sukusoluiksi, joista hedelmöityksen tuloksena syntyy tsygootteja. Näistä tsygooteista muodostuu jälleen uusia isäntiä infektoivia sporotsoiitteja.
Itiöeläimillä voi olla useita isäntäeliöitä, esimerkiksi malariaa aiheuttavilla Plasmodium-lajeilla isäntinä toimivat selkärankainen eläin ja hyttynen. Plasmodium-lajeilla suvullinen lisääntyminen ja hedelmöitys tapahtuvat hyttysen ruuansulatuselimistössä, josta sporotsoiitit hakeutuvat hyttysen sylkirauhasiin. Hyttysen syljen välityksellä sporotsoiitit päätyvät selkärankaisen elimistöön, jossa merotsoiitit muodostuvat maksasoluissa. Merotsoiitit puolestaan infektoivat veren punasoluja, ja tietyssä vaiheessa kehittyy gametosyyttejä, jotka päätyvät hyttysen elimistöön sen imettyä verta.
Bakteereista ja viruksista poiketen itiöeläimet ovat aitotumaisia taudinaiheuttajia. Tämän vuoksi niitä vastaan on vaikeaa kehittää sellaisia lääkkeitä, jotka vaikuttaisivat vain taudinaiheuttajaan, eivätkä olisi potilaalle haitallisia. Malariaa vastaan on tuloksetta yritetty kehittää rokotteita. Sitä tosin pystytään hoitamaan kiniini-, klorokiini- ja amodikiini-nimisillä lääkeaineilla. Itiöeläinten tutkiminen ja kasvattaminen laboratorio-olosuhteissa on myös vaikeaa. Viime vuosina joidenkin itiöeläinten genomia on selvitetty, mikä auttanee tulevaisuudessa ymmärtämään tarkemmin niiden elintoimintoja kehitettäessä niitä vastaan tehoavia lääkeaineita. On esimerkiksi huomattu, että solun toiminnalle välttämättömään apikoplastiin vaikuttavia lääkeaineita on mahdollista kehittää.
Itiöeläimet (Apicomplexa) ovat Chromalveolata-kuntaan kuuluva ryhmä yksisoluisia eliöitä. Ne kuuluvat Alveolata-pääjaksoon panssarilevien ja ripsieläinten kanssa. Itiöeläimiä on kuvattu noin 4500 lajia. Itiöeläinten lähimpiä sukulaisia ovat panssarilevät.
Itiöeläimet ovat pitkälle erikoistuneita eläinten solunsisäisiä loisia. Niiden suomenkielinen nimitys johtuu niiden elinkiertoon kuuluvasta itiömäisestä elomuodosta. Tieteellinen nimitys Apicomplexa puolestaan viittaa niiden solun päässä tavattavaan apikaalikompleksi-nimiseen rakenteeseen, jonka avulla eliöt tunkeutuvat isäntäsoluunsa.
Niiden solurakenteessa tavataan myös erikoinen apikoplasti-soluelin, joka on ilmeisesti evoluution kuluessa surkastunut ja lehtivihreänsä menettänyt viherhiukkanen. Apikoplasti tuottaa nykykäsityksen mukaan eräitä itiöeläimen tarvitsemia rasvahappoja. On todettu, että jos apikoplasti tuhotaan, itiöeläin ei kykene infektoimaan uusia isäntäsoluja.
Itiöeläimet aiheuttavat monia sairauksia. Näistä tunnetuimpia ovat:
Malaria (alaluokka Haemosporasina) Babesioosi (alaluokka Piroplasmasina) Toksoplasmoosi ja kryptosporidioosi (alaluokka Coccidiasina)Les apicomplexes (Apicomplexa), ou sporozoaires (Sporozoa), sont des eucaryotes unicellulaires tous parasites d'animaux ; ils ne possèdent pas de flagelle sauf à certains moments du cycle vital. Celui-ci fait généralement intervenir plusieurs hôtes. La présence d'un ADN chloroplastique vestigial chez certaines espèces laisse penser que ces organismes dérivent d'un ancêtre photosynthétique, ou qu'ils ont acquis et recyclé certains gènes d'un organisme photosynthétique.
Le cycle de vie est constitué par une alternance de générations haploïdes et diploïdes, chacune pouvant se multiplier par schizogonie.
La reproduction sexuée fait intervenir un microgamète mâle qui fusionne avec un macrogamète femelle. Un zygote le plus souvent flagellé est ainsi obtenu. Celui-ci se transforme ensuite en oocyste, forme de résistance car pourvu d'une paroi épaisse. Lorsque les conditions sont réunies, la méiose puis la sporogonie (nombreuses divisions cellulaires) ont lieu, donnant naissance à des formes infectieuses haploïdes (les sporozoïtes).
Plasmodium achiotense Plasmodium achromaticum Plasmodium acuminatum Plasmodium adunyinkai Plasmodium aegyptensis Plasmodium aeuminatum Plasmodium agamae Plasmodium anasum Plasmodium anomaluri Plasmodium arachniformis Plasmodium ashfordi Plasmodium atheruri Plasmodium aurulentum Plasmodium australis Plasmodium attenuatum Plasmodium azurophilum Plasmodium balli Plasmodium bambusicolai Plasmodium basilisci Plasmodium beebei Plasmodium beltrani Plasmodium berghei Plasmodium bertii Plasmodium bigueti Plasmodium biziurae Plasmodium booliati Plasmodium bouillize Plasmodium bowiei Plasmodium brodeni Plasmodium brasilianum Plasmodium brasiliense Plasmodium brumpti Plasmodium brucei Plasmodium brygooi Plasmodium bubalis Plasmodium bucki Plasmodium capistrani Plasmodium cathemerium Plasmodium cephalophi Plasmodium cercopitheci Plasmodium chabaudi Plasmodium chalcidi Plasmodium chiricahuae Plasmodium circularis Plasmodium circumflexum Plasmodium clelandi Plasmodium cordyli Plasmodium cnemaspi Plasmodium cnemidophori Plasmodium coatneyi Plasmodium coggeshalli Plasmodium colombiense Plasmodium columbae Plasmodium corradettii Plasmodium coturnixi Plasmodium coulangesi Plasmodium cuculus Plasmodium cyclopsi Plasmodium cynomolgi Plasmodium diminutivum Plasmodium diploglossi Plasmodium dissanaikei Plasmodium divergens Plasmodium dominicana Plasmodium draconis Plasmodium durae Plasmodium egerniae Plasmodium elongatum Plasmodium eylesi Plasmodium fabesia Plasmodium fairchildi Plasmodium falciparum Plasmodium falconi Plasmodium fallax Plasmodium fieldi Plasmodium fischeri Plasmodium foleyi Plasmodium formosanum Plasmodium forresteri Plasmodium floridense Plasmodium fragile Plasmodium galbadoni Plasmodium garnhami Plasmodium gallinaceum Plasmodium giganteum Plasmodium giovannolai Plasmodium girardi Plasmodium gonderi Plasmodium georgesi Plasmodium gologoense Plasmodium gracilis Plasmodium griffithsi Plasmodium guangdong Plasmodium gundersi Plasmodium guyannense Plasmodium heischi Plasmodium hegneri Plasmodium hermani Plasmodium heteronucleare Plasmodium hexamerium Plasmodium holaspi Plasmodium huffi Plasmodium hylobati Plasmodium icipeensis Plasmodium iguanae Plasmodium inconstans Plasmodium inopinatum Plasmodium inui Plasmodium japonicum Plasmodium jefferi Plasmodium jiangi Plasmodium josephinae Plasmodium joyeuxi Plasmodium juxtanucleare Plasmodium kempi Plasmodium knowlesi Plasmodium kentropyxi Plasmodium lacertiliae Plasmodium lainsoni Plasmodium landauae Plasmodium leanucteus Plasmodium lemuris Plasmodium lepidoptiformis Plasmodium lionatum Plasmodium lophurae Plasmodium loveridgei Plasmodium lutzi Plasmodium lygosomae Plasmodium mabuiae Plasmodium mackerrasae Plasmodium maculilabre Plasmodium maior Plasmodium majus Plasmodium malariae Plasmodium marginatum Plasmodium matutinum Plasmodium melanipherum Plasmodium mexicanum Plasmodium michikoa Plasmodium minasense Plasmodium morulum Plasmodium murinus Plasmodium necatrix Plasmodium nucleophilium Plasmodium octamerium Plasmodium odocoilei Plasmodium osmaniae Plasmodium ovale Plasmodium paddae Plasmodium papernai Plasmodium paranucleophilum Plasmodium parvulum Plasmodium pedioecetii Plasmodium pelaezi Plasmodium percygarnhami Plasmodium pessoai Plasmodium petersi Plasmodium pifanoi Plasmodium pinotti Plasmodium pinorrii Plasmodium pitheci Plasmodium pitmani Plasmodium polare Plasmodium pulmophilum Plasmodium reichenowi Plasmodium relictum Plasmodium rhadinurum Plasmodium rhodaini Plasmodium robinsoni Plasmodium rouxi Plasmodium sandoshami Plasmodium sasai Plasmodium saurocaudatum Plasmodium schweitzi Plasmodium scorzai Plasmodium semiovale Plasmodium semnopitheci Plasmodium shortii Plasmodium silvaticum Plasmodium simium Plasmodium tanzaniae Plasmodium tenue Plasmodium tejerai Plasmodium telfordi Plasmodium tomodoni Plasmodium torrealbai Plasmodium traguli Plasmodium tribolonoti Plasmodium tropiduri Plasmodium tumbayaensis Plasmodium tyrio Plasmodium uilenbergi Plasmodium uluguruense Plasmodium uncinatum Plasmodium uzungwiense Plasmodium watteni Plasmodium wenyoni Plasmodium vacuolatum Plasmodium vastator Plasmodium vaughani Plasmodium vautieri Plasmodium vinckei Plasmodium vivax Plasmodium volans Plasmodium voltaicum Plasmodium yoelii Plasmodium youngi
Plasmodium zonuriaeLes apicomplexes (Apicomplexa), ou sporozoaires (Sporozoa), sont des eucaryotes unicellulaires tous parasites d'animaux ; ils ne possèdent pas de flagelle sauf à certains moments du cycle vital. Celui-ci fait généralement intervenir plusieurs hôtes. La présence d'un ADN chloroplastique vestigial chez certaines espèces laisse penser que ces organismes dérivent d'un ancêtre photosynthétique, ou qu'ils ont acquis et recyclé certains gènes d'un organisme photosynthétique.
Os apicomplexos ou Apicomplexa (tamén Apicomplexia) son un amplo grupo de protistas caracterizados pola presenza nas súas células dun orgánulo especial denominado complexo apical. Son unicelulares, forman esporas e son exclusivamente parasitos de animais. As estruturas móbiles tales como os flaxelos ou pseudópodos están ausentes neles agás en certas fases dos gametos. É un grupo diverso no que se inclúen organismos como os coccidios, gregarinas, piroplasmas, hemogregarinas e plasmodios.[3] Algunhas enfermidades causadas por estes organismos son:
O nome do taxon Apicomplexa deriva do latín apex (extremo) e complexus (pregado) e fai referencia ao seu complexo apical. O antigo taxon Sporozoa (esporozoos) agrupaba a Apicomplexa, Microsporidia e Myxosporida, pero este agrupamento xa non se considera bioloxicamente válido e está desaconsellado.[4]
O primeiro protozoo apicomplexo observado foi visto por Anton van Leeuwenhoek en 1674 e era un ooquiste de Eimeria stiedae na vesícula biliar de coello. O primeiro membro do filum que foi nomeado (por Dufour en 1828) foi Gregarina ovata en insectos dermápteros. Durante o período 1826-1850, foron nomeadas 41 especies de 6 xéneros. No período 1951-1975, nomeáronse 1873 novas especies e 83 novos xéneros.
A maioría dos seus membros ten un ciclo vital complexo, que comprende fases de reprodución asexual e sexual. Tipicamente, os hóspedes destes parasitos contamínanse inxerindo quistes, que se dividen para producir os esporozoítos (esporulación) que entran nas súas células. Finalmente as células reventan, liberando merozoítos (esquizogonia) que infectan novas células. Porén, moitos Apicomplexa teñen máis dun hóspede.
O complexo apical é un orgánulo característico situado nun extremo da célula que presenta unhas vesículas chamadas roptrías e micronemas, que se abren na parte anterior da célula. Estas segregan os encimas que permiten ao parasito entrar noutras células. A extremidade está rodeada por unha banda de microtúbulos, chamada anel polar, e nos Conoidasida hai tamén un cono truncado (funil) de fibrilas chamado conoide.[3] No resto da célula, a excepción dunha pequena boca chamada microporo, a membrana apóiase nunhas vesículas denominadas alvéolos, formando unha estrutura semirríxida.
A presenza de alvéolos e outras características colocan a Apicomplexa no grupo Alveolata. Varios flaxelados relacionados, tales como Perkinsus e Colpodella teñen estruturas similares ao anel polar e foron incluídos antes neste grupo, mais agora parece que son parentes máis próximos dos Dinoflagellata. Son probablemente similares ao devanceiro común de ambos os grupos.
Outra semellanza con Dinoflagellata, é que as células de Apicomplexa conteñen un só plasto, moi peculiar, chamado apicoplasto, rodeado por tres ou catro membranas. Pénsase que entre as súas funcións están a síntese de lípidos, que parece ser necesaria para a súa supervivencia. Considérase xeralmente que comparten unha orixe común cos cloroplastos dos dinoflaxelados, aínda que algúns estudos suxiren que derivan en última instancia de algas verdes e non de algas vermellas.[5][6]
Apicomplexa abrangue un grupo de organismos que antes eran clasificados como esporozoos, protozoos parasitos sen flaxelos, nin pseudópodos nin cilios. A maior parte dos Apicomplexa son móbiles nalgunha parte do seu ciclo de vida. Os outros grupos principais tradicionais de esporozoos eran Ascetosporea (agora clasificado en Cercozoa), Myxozoa (agora clasificado entre os animais) e Microsporidia (agora clasificado entre os fungos). Ás veces, o termo esporozoo úsase como sinónimo de Apicomplexa e outras veces como un subconxunto deles.
Dentro dos apicomplexos distínguense tres grupos principais: coccidios, gregarinas e heemosporidios, cada un cun ciclo de vida un pouco diferente. Os coccidios e as gregarinas parecen estar bastante relacionados.
As gregarinas son xeralmente parasitos de anélidos, artrópodos e moluscos. Atópanse a miúdo no tracto dixestivo dos seus hóspedes pero poden invadir outros tecidos. Nun ciclo de vida típico de gregarina un trofozoíto desenvólvese no seu hóspede orixinando un plasmodio. Este despois divídese en moitos merozoítos por esquizogonia. Os merozoítos libéranse ao lisarse a célula hóspede e invaden novas células. Nun punto do ciclo de vida fórmanse gamontes, que son liberados por lise da célula hóspede e agrúpanse por sizixia. Cada gamonte forma múltiples gametos. Dous gametos fusiónanse formando un oocisto. Os oocistos abandonan o hóspede e son captados por un novo hóspede.
Os coccidios son xeralmente parasitos de vertebrados. Igual ca as gregarinas son normalmente parasitos das células epiteliais do intestino pero poden infectar outros tecidos. O típico ciclo de vida dos coccidios é similar ao das gregarinas pero difire en que se forman cigotos. Algúns trofozoítos agrándanse e convértense en macrogametos e outros divídense repeatidamente para formar microgametos. Os microgametos son móbiles e deben chegar ata o macrogameto para fecundalo. Como resultado da fecundación fórmase un cigoto, o cal á súa vez forma un oocisto, o cal é normalmente liberado fóra do corpo.
Os hemosporidios teñen ciclos de vida máis complexos nos que alternan entre un hóspede artrópodo e outro vertebrado. O trofozoíto parasita eritrocitos ou outros tecidos no hóspede vertebrado. Os microgametos e macrogametos están sempre no sangue. Os gametos son captados polo insecto vector cando succiona sangue. Os microgametos migran ao intestino do insecto e fusiónanse cos macrogametos. O macrogameto fecundado convértese nun oocineto, que penetra noutras partes do corpo do vector, transfórmase en oocisto e divídese inicialmente por meiose e despois por mitose (ciclo de vida haplonte) dando lugar a esporozoítos. Os esporozoítos saen do oocisto e migran ata as glándulas salivares do vector, e serán inxectados coa saliva cando o insecto pique a outro vertebrado.
O complicado ciclo de vida dos apicomplexos enténdese mellor tendo en conta a súa evolución. Apicomplexa crese que se orixinou a partir de Dinoflagellata, un gran grupo de protozoos fotosintéticos. Na actualidade crese que os antepasados dos apicomplexos foron organismos que eran presas doutros e que desenvolveron a capacidade de invadiren as células intestinais dos seus depredadores e posteriormente perderon a capacidade fotosintética. Algunhas especies de dinoflaxelados actuais poden invadir os corpos de celenterados e seguiren facendo a fotosíntese, o que é posible porque os órganos dos hóspede (augamar) son case transparentes. Noutros organismos con órganos opacos, o máis probable é que a capacidade fotosintética se perdese rapidamente.
Crese que os apicomplexos comezaron infectando a parede intestinal do hóspede cunha ruta de propagación fecal-oral. Nalgún punto da súa historia evolutiva, o parasito evolucionou adquirindo a capacidade de infectar outros órganos, como o fígado do hóspede. Este patrón é o que se observa no xénero Cryptosporidium. Nalgún outro momento da evolución, un apicomplexo desenvolveu a capacidade de infectar as células do sangue e de sobrevivir e infectar aos mosquitos. Unha vez que a transmisión utilizando como vector o mosquito quedou firmemente establecida, perdeuse a anterior vía de transmisión fecal-oral. A teoría actual (2007) suxire que os xéneros Plasmodium, Haemoproteus e Hepatocystis evolucionaron a partir de especies de Leukocytozoon. Os parasitos do xénero Leukocytozoon infectan aos leucocitos (glóbulos brancos) e as células do fígado e do bazo e transmítense por medio de moscas negras (Simulium), un amplo xénero de moscas relacionadas cos mosquitos.
Os leucocitos, a maioría dos hepatocitos e as células do bazo realizan activamente a fagocitose de partículas, facendo máis fácil a entrada na célula para o parasito. O mecanismo de entrada nos eritrocitos das especies do xénero Plasmodium está aínda pouco clara, tendo en conta que a entrada se realiza en menos de 30 segundos. Aínda non se sabe se este mecanismo evolucionou antes de que os mosquitos se convertesen nos principais vectores de transmisión de Plasmodium.
O patrón de alternancia entre a reprodución sexual e a asexual, que pode parecer confusa ao primeiro, é un patrón moi común nas especies parasitas. As vantaxes evolutivas deste tipo de ciclo de vida xa foron recoñecidas por Mendel.
En condicións favorables, a reprodución asexual é superior á sexual porque o proxenitor está ben adaptado ao seu medio ambiente e os seus descendentes compartirán os seus xenes. Durante situacións de estrés, tales como a transferencia a un novo hóspede, a reprodución sexual é polo xeral superior, xa que produce unha nova mestura dos xenes que, na media da poboación, dá lugar a individuos mellor adaptados ao novo ambiente.
Apicomplexa inclúe moitos patóxenos importantes para os seres humanos e animais domésticos. A diferenza das bacterias patóxenas, estes parasitos son protistas eucariontes e comparten moitas rutas metabólicas cos seus hóspedes. Este feito fai que o tratamento terapéutico sexa extremadamente difícil (un fármaco que dane ao parasito probablemente tamén danará ao seu hóspede). Non hai actualmente vacinas eficaces ou tratamentos dispoñibles para a maioría das enfermidades causadas por estes parasitos. Un posible obxectivo para os fármacos é o plasto especial que teñen (apicoplasto) e, de feito, os medicamentos existentes que son eficaces contra os apicomplexos, como as tetraciclinas, parecen actuar precisamente contra ese plasto.[7]
A investigación biomédica sobre os parasitos é difícil, porque ás veces é imposible manter cultivos vivos do parasito no laboratorio e manipular estes organismos. Recentemente, seleccionáronse varias especies para secuenciar o seu xenoma. A dispoñibilidade destas secuencias xenómicas proporciona unha nova oportunidade para aprendermos máis sobre a evolución e a bioquímica destes parasitos.
Os apicomplexos ou Apicomplexa (tamén Apicomplexia) son un amplo grupo de protistas caracterizados pola presenza nas súas células dun orgánulo especial denominado complexo apical. Son unicelulares, forman esporas e son exclusivamente parasitos de animais. As estruturas móbiles tales como os flaxelos ou pseudópodos están ausentes neles agás en certas fases dos gametos. É un grupo diverso no que se inclúen organismos como os coccidios, gregarinas, piroplasmas, hemogregarinas e plasmodios. Algunhas enfermidades causadas por estes organismos son:
Malaria (Plasmodium) Babesiose (Babesia) Coccidioses, entre as que están: Criptosporidiose (Cryptosporidium parvum) Ciclosporose (Cyclospora cayetanensis) Toxoplasmose (Toxoplasma gondii) Isosporíase (Isospora belli).O nome do taxon Apicomplexa deriva do latín apex (extremo) e complexus (pregado) e fai referencia ao seu complexo apical. O antigo taxon Sporozoa (esporozoos) agrupaba a Apicomplexa, Microsporidia e Myxosporida, pero este agrupamento xa non se considera bioloxicamente válido e está desaconsellado.
Apicomplexa adalah protista yang memiliki organel unik yang disebut apical complex. Apicomplexa bersifat uniseluler, membentuk spora dan merupakan parasit. Apicomplexa tidak memiliki flagella atau pseudopoda kecuali pada beberapa tahap gamet. Pada apicomplexa terdapat organisme seperti coccidia, gregarine, piroplasm, haemogregarine, dan malaria; beberapa penyakit yang disebabkan oleh organisme apicomplexa adalah:
Apicomplexa adalah protista yang memiliki organel unik yang disebut apical complex. Apicomplexa bersifat uniseluler, membentuk spora dan merupakan parasit. Apicomplexa tidak memiliki flagella atau pseudopoda kecuali pada beberapa tahap gamet. Pada apicomplexa terdapat organisme seperti coccidia, gregarine, piroplasm, haemogregarine, dan malaria; beberapa penyakit yang disebabkan oleh organisme apicomplexa adalah:
Babesiosis (Babesia) Malaria (Plasmodium) Penyakit yang disebabkan Coccidia, termasuk: Kriptosporidiosis (Cryptosporidium parvum) Siklosporiasis (Cyclospora cayetanensis) Toksoplasmosis (Toxoplasma gondii)Gródýr (fræðiheiti: Apicomplexa) er hópur frumdýra sem öll eru sníklar. Þau fjölga sér með gróum og nota oft tvo hýsla eins og mýrarköldusýkillinn, sem notar moskítófluguna til að berast á milli og notar síðan hryggdýr eins og mann til að þroskast í.
Apicomplexa Levine, 1970 è un subphylum di protozoi che comprende organismi endoparassiti, composti in gran parte da un'unica cellula, e dotati di strutture adatte a penetrare nelle cellule di altri esseri viventi e nutrirsene. Il loro nome (dal latino apex (sommità) e complexus (proteso)) deriva dal cosiddetto complesso apicale, una struttura cellulare fatta di fibrille, microtubuli o altri organelli cellulari con il quale questi organismi si introducono nelle cellule degli animali ospiti. In questo insieme sono raggruppate due classi, Sporozoea, suddivisa a sua volta in tre sottoclassi Gregarinia, Coccidea e Hematozoa, e la classe Perkinsea.
Questo gruppo di protozoi si riproduce sessualmente tramite cellule aploidi o tramite cellule diploidi. Il ciclo di vita di questi protozoi può essere molto complesso e richiede diversi organismi ospite. All'interno di questo gruppo esistono numerosi protozoi di fondamentale importanza per l'uomo e diversi animali domestici in quanto portatori di numerose patologie umane e animali, quali la malaria provocata dai protozoi del genere Plasmodium e la toxoplasmosi provocata dal parassita Toxoplasma gondii.
In generale la forma esterna di questi protozoi è di difficile caratterizzazione e va dall'amorfismo (molto simile alle amebe) alla classica forma a goccia. Nel periodo della riproduzione sessuale essi assumono una forma flagellare e i gameti possono variare in grandezza.
Internamente la caratteristica più importante è data sicuramente dal complesso apicale posto nella parte anteriore ed è composto di cinque parti. Da uno o più anelli polari, da un conoide posto all'interno dell'anello polare e composto da vari microtubuli a forma di serpentina, da una serie di micronemi, ovvero organelli di forma tubulare che si sviluppano longitudinalmente, da altri organelli detti roptrie e infine da microtubuli subpellicolari che partono dall'anello polare e si dipanano verso l'interno del protozoo. Le funzioni specifiche di ogni organello che compone il complesso apicale sono tuttavia ancora in fase di studio e restano sconosciute[1]. Tuttavia si ipotizza che le roptrie siano organi addetti alla secrezione di particolari sostanze che collaborano insieme agli altri organelli del complesso apicale per la penetrazione delle cellule dell'organismo ospite di questi parassiti.
Apicomplexa Levine, 1970 è un subphylum di protozoi che comprende organismi endoparassiti, composti in gran parte da un'unica cellula, e dotati di strutture adatte a penetrare nelle cellule di altri esseri viventi e nutrirsene. Il loro nome (dal latino apex (sommità) e complexus (proteso)) deriva dal cosiddetto complesso apicale, una struttura cellulare fatta di fibrille, microtubuli o altri organelli cellulari con il quale questi organismi si introducono nelle cellule degli animali ospiti. In questo insieme sono raggruppate due classi, Sporozoea, suddivisa a sua volta in tre sottoclassi Gregarinia, Coccidea e Hematozoa, e la classe Perkinsea.
Questo gruppo di protozoi si riproduce sessualmente tramite cellule aploidi o tramite cellule diploidi. Il ciclo di vita di questi protozoi può essere molto complesso e richiede diversi organismi ospite. All'interno di questo gruppo esistono numerosi protozoi di fondamentale importanza per l'uomo e diversi animali domestici in quanto portatori di numerose patologie umane e animali, quali la malaria provocata dai protozoi del genere Plasmodium e la toxoplasmosi provocata dal parassita Toxoplasma gondii.
Apicomplexa is in de biologie een groep protisten met de rang van een stam uit de groep van de Chromalveolata, waarvan vrijwel alle soorten parasitair zijn. Voor de Apicomplexa werd vroeger weleens de naam Sporozoa gebruikt. De stam omvat 4000 soorten, maar waarschijnlijk is de lijst met soorten nog lang niet compleet. Apicomplexa kunnen zowel ongewervelde als gewervelde dieren infecteren. Bepaalde soorten Apicomplexa kunnen voor mensen ernstige ziekten veroorzaken zoals malaria. Er zijn ook soorten waarmee wordt geëxperimenteerd om insectenpopulaties onder controle te houden. De soorten die tot de Apicomplexa behoren hebben gemeen dat ze ingewikkelde levenscycli hebben en dat er onderling veel verschil kan zijn tussen de levenscycli. Tijdens de levenscyclus vindt zowel ongeslachtelijke als geslachtelijke voortplanting plaats.
Indeling van Apicomplexa in klassen en onderklassen:
Stam Apicomplexa
Apicomplexa is in de biologie een groep protisten met de rang van een stam uit de groep van de Chromalveolata, waarvan vrijwel alle soorten parasitair zijn. Voor de Apicomplexa werd vroeger weleens de naam Sporozoa gebruikt. De stam omvat 4000 soorten, maar waarschijnlijk is de lijst met soorten nog lang niet compleet. Apicomplexa kunnen zowel ongewervelde als gewervelde dieren infecteren. Bepaalde soorten Apicomplexa kunnen voor mensen ernstige ziekten veroorzaken zoals malaria. Er zijn ook soorten waarmee wordt geëxperimenteerd om insectenpopulaties onder controle te houden. De soorten die tot de Apicomplexa behoren hebben gemeen dat ze ingewikkelde levenscycli hebben en dat er onderling veel verschil kan zijn tussen de levenscycli. Tijdens de levenscyclus vindt zowel ongeslachtelijke als geslachtelijke voortplanting plaats.
Apicomplexa (do latim apex, ponta ou topo + complex, trançado + a, sufixo) é um grande grupo taxonómico de protozoários, com cerca de 5000 espécies, caracterizados pela presença de um tipo de plastídeo, o apicoplasto, e um complexo apical nos estágios esporozoíto e merozoíto de seu ciclo de vida.[1]
São seres unicelulares, formadores de esporos, parasitas intracelulares obrigatórios (agem igual aos vírus) e não apresentam flagelos nem pseudópodes, a não ser em certos gametas. O táxon antigo Sporozoa foi criado por Schrevel em 1971 e agrupado com Apicomplexa junto com Ascetosporea, Microsporidia e Myxosporida. Este agrupamento não é mais válido biologicamente e seu uso é desencorajado.[2]
Muitas espécies são nocivas ao homem causando doenças, como por exemplo:
Possui duas formas de reprodução assexuada: a divisão múltipla e a esporogonia.
Foram raramente estudadas ao nível de subclasse. Morrison usando dados moleculares mostrou que haemosporidia são relacionados à gregarinas e que os piroplasmas e coccídios são grupos irmãos.[3] Outros estudos sugerem que haemosporidia e piroplasmas parecem ser clados irmãos e são mais intimamente relacionados aos coccídios do que as gregarinas.[4]
Transposons parecem ser raros neste filo mas foram identificados no gênero Ascogregarina e Eimeria.[4]
Todos os membros deste filo têm um estágio infeccioso — o esporozoíto — que possui três estruturas distintas no complexo apical. O complexo apical consiste de um conjunto de microtúbulos dispostos em espiral (conoide), uma organela secretora (roptrias), e um ou mais anéis polares. Corpos secretores adicionais, delgados e elétron-densos (micronema) cercados por um ou dois anéis polares podem também estar presentes. É esta estrutura que dá o nome ao filo.
Um outro grupo de organelas esféricas são distribuídos por toda a célula em vez de estar localizada no complexo apical e são conhecidas como grânulos densos. Têm tipicamente um diâmetro médio de cerca de 0,7 micrômetros. A secreção do conteúdo ocorre após a invasão do parasita e posterior localização dentro do vacúolo parasitário, e pode persistir por vários minutos.
Outras descobertas morfológicas que são comuns a todos os membros deste filo incluem:
Replicação:
Outros aspectos comuns neste filo são a reprodução sexual, uso de micrósporos para alimentação, movimento do corpo por flexão ou deslizamento, produção de cistos contendo esporozoítos como forma infectante e a ausência de cílios.
Apicomplexa (do latim apex, ponta ou topo + complex, trançado + a, sufixo) é um grande grupo taxonómico de protozoários, com cerca de 5000 espécies, caracterizados pela presença de um tipo de plastídeo, o apicoplasto, e um complexo apical nos estágios esporozoíto e merozoíto de seu ciclo de vida.
São seres unicelulares, formadores de esporos, parasitas intracelulares obrigatórios (agem igual aos vírus) e não apresentam flagelos nem pseudópodes, a não ser em certos gametas. O táxon antigo Sporozoa foi criado por Schrevel em 1971 e agrupado com Apicomplexa junto com Ascetosporea, Microsporidia e Myxosporida. Este agrupamento não é mais válido biologicamente e seu uso é desencorajado.
Muitas espécies são nocivas ao homem causando doenças, como por exemplo:
Babesiose (Babesia) Malária (Plasmodium) Formas de coccidiose incluem: Criptosporidíase (Cryptosporidium parvum) Ciclosporíase (Cyclospora cayetanensis) Isosporíase (Isospora belli) Toxoplasmose (Toxoplasma gondii)Possui duas formas de reprodução assexuada: a divisão múltipla e a esporogonia.
Sporozoarele reprezintă un grup de protiste asemănătoare animalelor, ce fac parte din încrengătura Apicomplexa. Sunt cunoscute în jur de 25.000 specii. Acestea nu prezintă organite de locomoție, precum cilii sau flagelii. Hrănirea este exclusiv heterotrofă. Toate sporozoarele sunt organisme parazite, care își dobândesc nutrienții pe seama altor organisme. Au un ciclu de viață complex, care implică mai multe gazde. Sunt unicelulare.
Acestea posedă un apicoplast și un complex apical, care le permite să infecteze gazdele. Complexul apical constă din:
Majoritatea sporozoarelor sunt haploide. Reproducerea este asexuată, prin spori, și sexuată, prin microgameți masculini și macrogameți feminini. Aceste prezintă gametogamie, care este urmată de producerea sporilor. În plus, la unele sporozoare poate fi întâlnită sporogonia.
Sporozoarele reprezintă un grup de protiste asemănătoare animalelor, ce fac parte din încrengătura Apicomplexa. Sunt cunoscute în jur de 25.000 specii. Acestea nu prezintă organite de locomoție, precum cilii sau flagelii. Hrănirea este exclusiv heterotrofă. Toate sporozoarele sunt organisme parazite, care își dobândesc nutrienții pe seama altor organisme. Au un ciclu de viață complex, care implică mai multe gazde. Sunt unicelulare.
Apicomplexa är en protistgrupp som tillhör alveolaterna.[1]. Det är en stor grupp protozoer (urdjur) vars kännetecken är närvaron av ett apikomplex (organeller som används för att ta sig in i värdcellen) någon gång under deras livscykel. De är uteslutande parasitiska och saknar organeller som flageller och pseudopodier utom under vissa gametstadier.
Apicomplexa kan orsaka sjukdomar som:
De flesta medlemmarna i stammen har en komplicerad livscykel som varierar mellan sexuell och asexuell reproduktion. I normalfallet infekteras värden genom upptagandet av cystor som delar sig och producerar de sporozoiter som tar sig in i värdcellerna. När cellen spricker sprids merozoiter som infekterar nya celler. Processen kan upprepas flera gånger innan till slut gamonter (moderceller) bildas som skapar gameter (könsceller) som till slut bildar nya cystor. Detta är dock bara ett grundmönster, variationerna är många och många apicomplexa har mer än en värd.
I det apikala komplexet finns vesiklar kallade rhoptrier och mikronemer som öppnar cellen och utsöndrar enzym som gör att parasiten kan ta sig in i andra celler. I cellens ena ände finns ett band av mikrotubuler (kallad "polring") och hos conoidsida dessutom stänger som bildar en tratt ("konoid"). Resten av cellmembranet stöds av vesiklar kallade alveoler[förtydliga] som bildar en halvstyv pellikel.
Alveolerna tillhör de kännemärken som gör att apicomplexa tillhör gruppen alveolater. Flera besläktade flagellater som perkinsus och colpodella liknar apicomplexa och räknades tidigare hit, men betraktas idag som mer besälktade med dinoflagellaterna. Förmodligen liknar de två gruppernas gemensamma anfader.
Ytterligare en likhet är att apicomplexa-celler innehåller en enda plastid kallad apicoplast som omges av tre eller fyra membran. Plastidens antas bland annat stå för syntetisering av lipider och tycks vara livsnödvändig. De förmodas ha ett ursprung gemensamt med dinoflagellaternas kloroplaster även om vissa studier antyder att de ursprungligen härstammar från grönalger snarare än rödalger.
Apicomplexa utgör huvuddelen av den grupp som tidigare kallades sporozoa och som bestod av parasitiska protozoer som saknade flageller, pseudopodier respektive cilier. De flesta apicomplexa är dock mobila. Gruppen innehöll även ascetospora, myxozoa (som man idag vet härstammar från djur) och mikrosporidier (härstammar från svampar). Det förekommer att sporozoa används synonymt med apicomplexa eller någon del av stammen.
Apicomplexa är en protistgrupp som tillhör alveolaterna.. Det är en stor grupp protozoer (urdjur) vars kännetecken är närvaron av ett apikomplex (organeller som används för att ta sig in i värdcellen) någon gång under deras livscykel. De är uteslutande parasitiska och saknar organeller som flageller och pseudopodier utom under vissa gametstadier.
Apicomplexa kan orsaka sjukdomar som:
Babesios (Babesia spp.) Kryptosporidios (Cryptosporidium spp.) Malaria (Plasmodium spp.) Toxoplasmos (Toxoplasma gondii)De flesta medlemmarna i stammen har en komplicerad livscykel som varierar mellan sexuell och asexuell reproduktion. I normalfallet infekteras värden genom upptagandet av cystor som delar sig och producerar de sporozoiter som tar sig in i värdcellerna. När cellen spricker sprids merozoiter som infekterar nya celler. Processen kan upprepas flera gånger innan till slut gamonter (moderceller) bildas som skapar gameter (könsceller) som till slut bildar nya cystor. Detta är dock bara ett grundmönster, variationerna är många och många apicomplexa har mer än en värd.
I det apikala komplexet finns vesiklar kallade rhoptrier och mikronemer som öppnar cellen och utsöndrar enzym som gör att parasiten kan ta sig in i andra celler. I cellens ena ände finns ett band av mikrotubuler (kallad "polring") och hos conoidsida dessutom stänger som bildar en tratt ("konoid"). Resten av cellmembranet stöds av vesiklar kallade alveoler[förtydliga] som bildar en halvstyv pellikel.
Alveolerna tillhör de kännemärken som gör att apicomplexa tillhör gruppen alveolater. Flera besläktade flagellater som perkinsus och colpodella liknar apicomplexa och räknades tidigare hit, men betraktas idag som mer besälktade med dinoflagellaterna. Förmodligen liknar de två gruppernas gemensamma anfader.
Ytterligare en likhet är att apicomplexa-celler innehåller en enda plastid kallad apicoplast som omges av tre eller fyra membran. Plastidens antas bland annat stå för syntetisering av lipider och tycks vara livsnödvändig. De förmodas ha ett ursprung gemensamt med dinoflagellaternas kloroplaster även om vissa studier antyder att de ursprungligen härstammar från grönalger snarare än rödalger.
Apicomplexa utgör huvuddelen av den grupp som tidigare kallades sporozoa och som bestod av parasitiska protozoer som saknade flageller, pseudopodier respektive cilier. De flesta apicomplexa är dock mobila. Gruppen innehöll även ascetospora, myxozoa (som man idag vet härstammar från djur) och mikrosporidier (härstammar från svampar). Det förekommer att sporozoa används synonymt med apicomplexa eller någon del av stammen.
Sınıf Blastocystea
Sınıf Conoidasida
Sporozoa (Apicomplexa), parazitik tekhücreli canlılardır. Protista aleminin bir şubesidir. Özelleşmiş bir hareket yapıları yoktur. Yaşamlarında bir dönemi hareketsiz, spor formunda geçirirler. Sıtma sebebi Plasmodium bu grubun üyesidir. Metagenez ile ürerler. Yaklaşık 5.000 türü tanımlanmıştır.
Parazit bir hücreliler olup sporla ürerler.
Omurgasız ve omurgalı hayvanların hücre içinde ve hücre arasında parazit olarak yaşarlar. Vücutlarının esnek yapısı sayesinde kasılıp gevşeyerek hareket edebilmektedirler.
Kontraktil kofulları yoktur. İnsanda sıtmaya neden olan Plasmodyum sporlu protistalardandır.
.
Всі представники типу — паразити безхребетних і хребетних тварин. В життєвому циклі відбувається чергування безстатевого розмноження (множинний розподіл — шизогонія), статевого процесу і спорогонії. Принаймні частина життєвого циклу проходить всередині клітин господарів. Стадії, що виконують функцію проникнення в клітини (зоїти), мають специфічний комплекс органел — апікальний комплекс. Джгутики у споровиків присутні тільки на стадії гамет.
Тип споровиків включає понад 5000 видів, у тому числі токсоплазма, кокцидії.
У межах типу виділяються наступні таксони:
Apicomplexa (còn được gọi là Apicomplexia) là một ngành lớn gồm những loài ký sinh đơn bào. Hầu hết trong số chúng có một dạng bào quan độc đáo bao gồm một loại lạp thể gọi là apicoplast, và một cấu trúc phức hợp đỉnh. Bào quan là một sự thích nghi mà sinh vật ký sinh đơn bào áp dụng cho hoạt động thẩm thấu của tế bào chủ.
Apicomplexa (còn được gọi là Apicomplexia) là một ngành lớn gồm những loài ký sinh đơn bào. Hầu hết trong số chúng có một dạng bào quan độc đáo bao gồm một loại lạp thể gọi là apicoplast, và một cấu trúc phức hợp đỉnh. Bào quan là một sự thích nghi mà sinh vật ký sinh đơn bào áp dụng cho hoạt động thẩm thấu của tế bào chủ.
Apicomplexa Levine, 1970[1][2]
СинонимыАпикомпле́ксы (лат. Apicomplexa)[1], или споровики́ (лат. Sporozoa)[3] — тип простейших из группы альвеолят (лат. Alveolata). Все представители типа являются облигатными паразитами позвоночных и беспозвоночных животных. Общность плана строения апикомплекс наиболее отчетливо проявляется на стадии зоита и выражается в наличии специфического комплекса органелл — апикального комплекса. Покровы представлены характерной для альвеолят пелликулой. В жизненном цикле большинства представителей типа обнаружен половой процесс. У многих апикомплекс (кокцидий, гемоспоридий, пироплазмид, некоторых грегарин) по крайней мере часть жизненного цикла проходит внутри клеток хозяев.
Тип включает более 5000[4] видов, среди которых встречаются возбудители заболеваний человека и животных (малярийный плазмодий, токсоплазма, криптоспоридии).
Разнообразие жизненных циклов апикомплекс чрезвычайно велико. Все стадии жизненного цикла, за исключением зиготы — гаплоидны. Далее представлена обобщенная (базовая) модель, характерная для многих кокцидий[5].
Заражение хозяина обычно происходит при попадании в его организм экзогенных стадий жизненного цикла — спороцист. В кишечнике хозяина из спороцист выходят спорозоиты, которые проникают в клетки кишечного эпителия. Внутри клетки хозяина паразит (на данном этапе обычно обозначаемый как трофозоит) начинает активно питаться и расти, после чего преобразуется в меронт и приступает к бесполому размножению, проходящему по типу множественного деления — мерогонии[en]. Образовавшееся в результате потомство — мерозоиты могут вновь заражать другие клетки хозяина и проходить мерогонию. Обычно такой цикл повторяется несколько раз.
Следующая стадия жизненного цикла споровиков связана с подготовкой к половому процессу. После очередного деления мерозоиты дают начало макрогамонтам и микрогамонтам, в которых соответственно развиваются макро и микрогаметы. Последние несут жгутики и имеют меньшие размеры. Слияние гамет обычно происходит по типу анизогамии. Образующаяся зигота (ооциста) претерпевает редукционное деление (спорогония) преобразуясь в спороцисту, которая выводится во внешнюю среду для заражения других особей.
В отличие от кокцидий, подавляющее большинство грегарин паразитирует внеклеточно. Экзогенной стадией обычно является ооциста, выходящие из неё спорозоиты прикрепляются передним концом к эпителию хозяина. Характерным отличием жизненного цикла грегарин так же является образование сизигия — объединение гамонтов перед началом гаметогенеза, их совместное инцистирование и формирование стадии гаметоцисты, под покровами которой проходит гаметогенез.
В качестве основных модификаций жизненного цикла споровиков можно выделить утрату стадии мерогонии (характерно для многих грегарин), а также развитие в нескольких хозяевах (широко распространено среди кровяных споровиков)[5].
Зоит представляет собой узкую клетку с крупным ядром, покрытую трёхмембранной пелликулой. Наружная мембрана непрерывна, две внутренние прерываются в области микропоры, предположительно выполняющей функции клеточного рта.
Характерной особенностью зоитов является апикальный комплекс, состоящий из коноида, роптрий и микронем. Коноид — это усеченный конус из микротрубочек, расширяющийся вглубь, участвующий у многих споровиков в процессе проникновения через покровы инфицируемой клетки, а также, возможно, в процессе питания некоторых грегарин[6]. Роптриями называют органеллы, заполненные содержимым, растворяющим покровы клетки и тем облегчающим проникновение зоита внутрь. Роль микронем, предположительно, заключается в синтезе веществ, дополняющих секрет роптрий.
В пределах типа выделяются следующие таксоны:
Иногда в состав споровиков включают в качестве класса группу свободноживущих жгутиконосцев Colpodellea, также обладающих полноценным апикальным комплексом, участвующем, однако, в процессе питания данных протистов, а не в проникновении в клетку хозяина. Данный таксон также часто рассматривают в качестве возможной предковой формы всех остальных апикомплекс[3]
Роль споровиков определяется их паразитическим образом жизни. К этому типу относятся возбудители ряда болезней человека (малярия, токсоплазмоз, кокцидиоз, саркоцистоз) и животных (кокцидиоз, бабезиоз).
Апикомпле́ксы (лат. Apicomplexa), или споровики́ (лат. Sporozoa) — тип простейших из группы альвеолят (лат. Alveolata). Все представители типа являются облигатными паразитами позвоночных и беспозвоночных животных. Общность плана строения апикомплекс наиболее отчетливо проявляется на стадии зоита и выражается в наличии специфического комплекса органелл — апикального комплекса. Покровы представлены характерной для альвеолят пелликулой. В жизненном цикле большинства представителей типа обнаружен половой процесс. У многих апикомплекс (кокцидий, гемоспоридий, пироплазмид, некоторых грегарин) по крайней мере часть жизненного цикла проходит внутри клеток хозяев.
Тип включает более 5000 видов, среди которых встречаются возбудители заболеваний человека и животных (малярийный плазмодий, токсоплазма, криптоспоридии).
頂複門(學名:Apicomplexa,亦作Apicomplexia)是原生真核生物之下一個寄生囊泡虫的门,是一個物種分類眾多的大类。其中大部分含有独特的四层膜细胞器顶质体,屬於色素體的一種,可幫助進入宿主的細胞內。
頂複門均为单细胞,产芽孢,并且完全寄生(英语:endoparasite)于动物[3],就除了 Nephromyces(英语:Nephromyces)屬物種。這個屬的物種與海洋動物共生,過往曾被錯誤歸類為壶菌门真菌的成員[4]。本目物種的移動性構造,例如:鞭毛或偽足等,只會於某些配子階段出現。
过去的分类方法曾经将顶复门内的生物与粘孢子虫和微孢子虫共同归类为Sporozoa(孢子纲),现代分子生物学将后两者分别归入动物界与真菌界后,剩下的大部分归类为顶复门,孢子纲这一分类类群就不再使用了。[5]
頂複門是一个多样化的群体,包括球虫、簇虫、梨形虫、凝血簇虫(英语:haemogregarine)、疟原虫等。有许多传染病的病原体属于这个门,例如:
頂複門的學名Apicomplexa来源于拉丁文单词apex(顶端)和 complexus(複合):特指在頂複門的生命週期還在孢子階段時在細胞內的一組細胞器。
本分類取材於Perkins et al. (2000)[6]。儘管由於柏金絲蟲科(英语:Perkinsidae)被確認為甲藻的姐妹群而不再屬於頂複門,但其餘分類仍然有參考價值。茲詳列如下:
|archive-date=中的日期值 (帮助) 頂複門(學名:Apicomplexa,亦作Apicomplexia)是原生真核生物之下一個寄生囊泡虫的门,是一個物種分類眾多的大类。其中大部分含有独特的四层膜细胞器顶质体,屬於色素體的一種,可幫助進入宿主的細胞內。
頂複門均为单细胞,产芽孢,并且完全寄生(英语:endoparasite)于动物,就除了 Nephromyces(英语:Nephromyces)屬物種。這個屬的物種與海洋動物共生,過往曾被錯誤歸類為壶菌门真菌的成員。本目物種的移動性構造,例如:鞭毛或偽足等,只會於某些配子階段出現。
过去的分类方法曾经将顶复门内的生物与粘孢子虫和微孢子虫共同归类为Sporozoa(孢子纲),现代分子生物学将后两者分别归入动物界与真菌界后,剩下的大部分归类为顶复门,孢子纲这一分类类群就不再使用了。
頂複門是一个多样化的群体,包括球虫、簇虫、梨形虫、凝血簇虫(英语:haemogregarine)、疟原虫等。有许多传染病的病原体属于这个门,例如:
巴倍斯虫病(英语:Babesiosis):由巴倍虫属( Babesia)物種引起; 疟疾:由瘧原蟲(Plasmodium)引起; 隐孢子虫病:由小隱孢子蟲(Cryptosporidium parvum)引起; Cyclosporiasis(英语:Cyclosporiasis) (Cyclospora cayetanensis(英语:Cyclospora cayetanensis)) 等孢子蟲病(英语:Cystoisosporiasis):由一種名為Cystoisospora belli(英语:Cystoisospora belli)(舊作Isospora Belli)的等孢子蟲屬引起; 弓蟲症:由弓形虫(Toxoplasma gondii)引起。頂複門的學名Apicomplexa来源于拉丁文单词apex(顶端)和 complexus(複合):特指在頂複門的生命週期還在孢子階段時在細胞內的一組細胞器。
 分類 ドメ
分類 ドメアピコンプレックス門(Apicomplexa;またはアピコンプレクサ類)は原生生物界の門の1つ。アピコンプレクサ類は生活環のどこかでアピカルコンプレックス(apical complex、頂端複合構造)という構造を持つという点で特徴づけられる原生生物の大きなグループである。寄生性であり、配偶子の時期を例外として、鞭毛や仮足を持たない。
アピコンプレクサ類の生物はコルポデラ類を除いて全てが寄生性である。宿主は後生動物全般で幅広いが、個々の種は比較的宿主特異性が高いと考えられている。寄生部位も腸管、体腔、組織、血球と様々で、細胞内寄生のものも細胞外寄生のものもある。宿主に対して病原性を示すもの多く知られており、マラリア原虫のような著名なヒトの病原体が含まれている。アピコンプレクサ類によって引き起こされる代表的な疾病には以下のようなものがある。
アピコンプレクサ類は無性生殖と有性生殖からなる複雑な生活環を持っており、一般に次の3つの時期に区分できる。
標準的には、宿主はオーシストを摂取することで感染する。まずオーシスト中のスポロゾイトが細胞に侵入してメロゴニーを行い個体数を増やす。生じたメロゾイトは再び周辺の細胞に侵入してメロゴニーを繰り返す(1)。その後、雌雄の生殖母体が分化してガメトゴニーを行い(2)、生じたガメートが受精する(3)とスポロゴニーにより新しいオーシストが生じる(4)。この基本的なパターン以外に様々なバリエーションがあり、また単一宿主のものだけでなく複数の宿主を持つものも多数ある。
アピコンプレクサ類は基本的に単細胞であるが、大きさは数μmから数mmまでと極めて幅がある。細胞膜がアルベオール(alveole、胞室)という小胞に裏打ちされ、やや堅いペリクル(pellicle、外被)を形成している。アルベオールの存在やその他の特徴から、アピコンプレクサ類はアルベオラータというグループに所属させられている。ペリクルは細胞のほぼ全体を覆っているが、飲作用(ピノサイトーシス)を行うミクロポア(micropore、微小孔)の所にはない。
細胞の前端にあるアピカルコンプレックスは、元来はミゾサイトーシスに使われる構造だと考えられているが、大部分の種では宿主細胞との接触や細胞内への侵入に関与していると考えられる。先端は極輪(polar ring)という微小管の帯が取り囲んでおり、多くの原虫にはさらにコノイド(conoid、円錐体)という漏斗状の微小管束がある。開口するロプトリー(rhoptry)とミクロネーム(microneme)という小胞があり、原虫が宿主細胞に侵入するための酵素を分泌している。
もう1つの特徴として、アピコンプレクサ類の細胞にはアピコプラストと呼ばれる細胞小器官があって、3ないし4枚の膜に囲まれていることが挙げられる。その機能は脂質合成などだろうと思われるが、原虫の生存に必須であるらしい。ただしクリプトスポリジウムや(少なくとも一部の)グレガリナ類にはアピコプラストは存在しないと考えられている。アピコプラストと渦鞭毛藻の葉緑体は両方とも紅藻由来だと考えられ、したがって一般的には両分類群の共通祖先から受け継がれたものだと考えられている。しかしアピコプラストは緑藻由来だとする意見もある。
アピコンプレックス門の下位分類は、分子系統解析による再検討が行われている段階である。比較的安定的に受け入れられている分類群は以下の4つであり、これらを綱または亜綱にあてることが多い。
このうち、住血胞子虫類のみ、もしくは住血胞子虫類とピロプラズマ類の2つをコクシジウム類に含めることがある。またピロプラズマ類を住血胞子虫類に含めて扱うこともある。
クリプトスポリジウム(Cryptosporidium)は従来コクシジウム類だと考えられてきたが、最近の分子系統解析によればむしろ特殊化したグレガリナ類であるらしい。実際に培養実験によって、細胞外でグレガリナ類に似た形態をとる時期の存在が判明している。
アピコンプレックス門への所属が議論されるものとして、パーキンサス類とコルポデラ類がある。パーキンサス(ペルキンスス、Perkinsus)は軟体動物に寄生する原虫で、はじめ菌類として分類されていたが、鞭毛虫ステージにアピカルコンプレックス類似の構造が発見されたことでアピコンプレックス門に移された。現在では分子系統解析などにより、アピコンプレクサ類よりも渦鞭毛虫に近縁であると考えられているため、アピコンプレックス門には含めないようになってきている。コルポデラ(Colpodella)は自由遊泳して他の原生生物を吸引消化する鞭毛虫であり、吻(rostrum)がアピカルコンプレックス類似の構造でできている。分子系統解析でもアピコンプレクサ類に近縁であることが示されているが、アピコンプレックス門に含める意見と含めない意見と両論がある。これらの生物はアピコンプレクサ類と渦鞭毛虫の共通祖先に似ているのだろうと考えられている。
アピコンプレックス門は、かつて胞子虫と呼ばれていた寄生性原虫の大部分を含んでいる。もともと胞子虫(Sporozoa Leuckart, 1879)という分類群はグレガリナ類とコクシジウム類からなるものであり、その意味では胞子虫類とアピコンプレクサ類とはほぼ同義である。これ以外に粘液胞子虫類・微胞子虫類・アセトスポラ類をも「胞子虫」として扱っていた期間が長く続いていたが、これはどちらかと言えば便宜上のもので、当時から系統の異なる生物だと考えられていたし、現在ではそれぞれ全く別の生物だと確定している。この経緯については胞子虫の項に詳しい。このような場合、分類学の一般論としては、範囲を狭めて(元に戻して)胞子虫という名を使い続けるのが妥当である。まして胞子虫という名は古典的4分類の1つとして長く親しまれたものであり、アピコンプレックス門という名称が受け入れられるまでには長い論争があった。
獣医学分野で活躍した寄生虫学者Norman D. Levineは、胞子に相当する形態を取らないため胞子虫類でないと疑われていたピロプラズマ類について、アピカルコンプレックスが認められることから胞子虫と近縁と考え、1969年にこれらをまとめてPolannulifera門とすることを提案した。これは翌年Apicomplexa門と改名され、グレガリナ類・コクシジウム類・住血胞子虫類を含む胞子虫綱と、ピロプラズマ綱の2綱が置かれていた。つまり、アピコンプレクサ類は(狭義の)胞子虫類を含みさらに拡大させた概念である。しかし1970年代前半には、ピロプラズマ類は住血胞子虫類と近縁なので独立の綱を与えるべきでなく、したがってアピコンプレクサ類は胞子虫類の同義語で不要と批判された。
ところが1976年に菌類だと考えられていた軟体動物の病原体がアピカルコンプレックスとよく似た構造を持っている事が判明する。Levineはこれにパーキンサスと名付け、アピコンプレックス門を胞子虫綱(Sporozoea、和名では種虫綱と訳し変える)とパーキンサス綱の2綱とした。これが1980年の合意体系で採用され、普及のきっかけとなった。その後の1982年、パーキンサスの構造は渦鞭毛虫によく似ていて、アピカルコンプレックスも本当に相同か疑わしいと批判された。またもやアピコンプレクサ類は胞子虫類の同義語となり不要ということになるが、しかしすでに合意体系で採用されていたこともあり、アピコンプレックス門は普及し続けた。
Levineはその後もアピカルコンプレックスを重視しつづけ、1988年にはアピコンプレックス門にコノイドの有無に着目した2綱(Conoidasida, Aconoidasida)を置くようにし、これによって分類体系から「胞子虫」という名前自体を消してしまった。Levineの死後、分子系統解析によって(狭義の)胞子虫類と(少なくとも一部の)コルポデラ類とが単系統群をなす事が判明しており、アピコンプレックス門という分類群自体は概ね受容されている。一方Conoidasidaという分類群は単系統にはならないため、今後系統を反映した体系に置き換えられることになるだろう。
ウィキメディア・コモンズには、アピコンプレックス門に関するカテゴリがあります。
アピコンプレックス門(Apicomplexa;またはアピコンプレクサ類)は原生生物界の門の1つ。アピコンプレクサ類は生活環のどこかでアピカルコンプレックス(apical complex、頂端複合構造)という構造を持つという点で特徴づけられる原生生物の大きなグループである。寄生性であり、配偶子の時期を例外として、鞭毛や仮足を持たない。
정단복합체충류(Apicomplexa 또는 Apicomplexia) 또는 첨복포자충류(尖複胞子蟲類)[3]는 원생생물의 대형 분류군의 하나이다.
좁은 의미의 정단복합체충류
















































