
腕足動物(わんそくどうぶつ、Brachiopoda)は、2枚の殻を持つ海産の底生無脊椎動物。シャミセンガイ、チョウチンガイなどと呼ばれるものを含む。一見して二枚貝に似るが、体制は大きく異なり、貝類を含む軟体動物門ではなく、独立の腕足動物門に分類される。
化石記録ではカンブリア紀に出現し、古生代を通じて繁栄したグループだが、その後多様性は減少し、現生の種数は比較的少ない。伝統的には無関節綱と有関節綱に分けられてきたが、それとは異なる分類体系も提案されている。
学名のBrachiopodaはギリシャ語で腕を意味するbrachiumと、足を意味するpodaを合わせたもので、和名はその直訳である[1][2]。
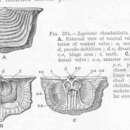
二枚貝のように2枚の殻を持つが、二枚貝類の殻は体の左右に1枚ずつあるのに対して、腕足動物の殻は背腹にあるとされている。殻の成分は分類群によって異なり、有関節類と一部の無関節類は炭酸カルシウム、他はキチン質性のリン酸カルシウムを主成分とする。それぞれの殻は左右対称だが、背側の殻と腹側の殻はかたちが異なる。2枚の殻は、有関節類では蝶番によって繋がるが、無関節類は蝶番を持たず、殻は筋肉で繋がる[2]。殻長は5センチメートル前後のものが多い[1]。
腹殻の後端から肉茎が伸びる。肉茎は体壁が伸びてできたもので、無関節類では体腔や筋肉を含み、伸縮運動をするが、有関節類の肉茎はそれらを欠き、運動の役には立たない。種によっては肉茎の先端に突起があり、海底に固着するときに用いられる[1][2]。肉茎を欠く種もいる[2]。
殻は外套膜から分泌されてできる。外套膜は殻の内側を覆っていて、殻のなかの外套膜に覆われた空間、すなわち外套腔を形成する。外套腔は水で満たされていて、触手冠(英語版)がある。触手冠は口を囲む触手の輪で、腕足動物では1対の腕(arm)に多数の細い触手が生えてできている。有関節類では、この腕は腕骨により支持されるが、無関節類は腕骨を持たず、触手冠は体腔液の圧力で支えられる[1][2]。
消化管はU字型。触手冠の運動によって口に入った餌(後述)は、食道を通って胃、腸に運ばれる。無関節類では、消化管は屈曲して直腸に繋がり、外套腔の内側か右側に開口する肛門に終わるが、有関節類は肛門を欠き、消化管は行き止まり(盲嚢)になる[1]。
循環系は開放循環系だが不完全。腸間膜上に心臓を持つ。真の血管はなく、腹膜で囲われた管がある。血液と体腔液は別になっているとされる[1][2]。ガス交換は体表で行われる[1]。1対か2対の腎管を持ち、これは生殖輸管の役割も果たす。
神経系はあまり発達していない。背側と腹側に神経節があり、2つの神経節は神経環で繋がっている。これらの神経節と神経環から、全身に神経が伸びる[1]。
全種が海洋の底生動物である。多くの種は、肉茎の先端を底質に固着させて体を固定するか、砂に固着させて体を支える支点とする。肉茎を持たない種は、硬い底質に体を直接固定する。体を底質に付着させない種もいる[1]。
餌を取るために、殻をわずかに開き、触手冠の繊毛の運動によって、外套腔内に水流を作り出す。水中に含まれる餌の粒子は、触手表面の繊毛によって、触手の根元にある溝に取り込まれ、口へと運ばれる。主な餌は植物プランクトンだが、小さな有機物なら何でも食べる[1]。
有性生殖のみで繁殖し、無性生殖はまったく知られていない。わずかに雌雄同体のものが知られるが、ほとんどの種は雌雄異体[2]。雌雄異体のものでも、性的二形はあまりない[2]。体外受精で、卵と精子は腎管を通じて海水中に放出され、受精するのが一般的。一部の種では、卵は雌の腎管や外套腔、殻の窪みなどに留まり、そこで受精が起こる。その場合には、受精卵は幼生になるまで、受精した場所で保護される[2]。
受精卵は放射型の全等割を経て発生する。原口は発生過程で閉じてしまい、口は二次的にできる[1][2]。どの種も浮遊する幼生の期間を持つが、幼生の特徴は有関節類と無関節類で大きく異なる[1][2]。無関節類の幼生は触手冠と2枚の殻、肉茎を備え、成体とそれほど変わらない構造を持つ。成体と異なるのは、肉茎が折れ曲がって外套腔内にあることと、外套膜に比べて体と触手冠が不釣合いに大きいことである。大きな触手冠は移動と摂食に用いられる。この幼生は大きな変態を遂げることなく成体になる。一方で、有関節類の幼生には殻がなく、体は頭葉、外套葉、茎葉の3つに分かれている。自力で餌を取ることはできず、卵黄の栄養のみに依存し、浮遊期間は短い。茎葉を使って着生すると、変態して成体になる。
腕足動物は、触手冠などいくつかの特徴を箒虫動物門(ホウキムシ類)、外肛動物門(コケムシ類)と共有するため、この2群とともに触手動物 (Tentaculata) あるいは触手冠動物としてまとめられてきた。触手冠動物は、旧口動物的な特徴もあるものの、放射型の卵割、成体の口が原口に由来しないこと、体の3節構造など新口動物の特徴を持つことから、伝統的には新口動物の1グループとされていた[3][4]。しかし、分子系統学の結果は、触手冠動物を旧口動物に含めることを支持することが多く、こちらが有力となっている[5]。
前口動物のなかで、触手冠動物は、担輪動物(軟体動物など、トロコフォア幼生を持つグループ)と併せて、冠輪動物としてまとめる考えが支持されている[6]。しかし、冠輪動物のなかでの、腕足動物の位置は明らかではない[5]。触手冠動物が単系統群になるという伝統的な考えも疑われている[3]。
キャバリエ=スミスは触手冠動物の3群のなかで、外肛動物だけは異なる系統的位置にあるが、腕足動物と箒虫動物は単系統になると考えて、この2群を亜門として含む腕動物(英語版)門を提唱した[7]。ヘルムカンプらの研究は腕動物の単系統性を支持し、さらに紐形動物(ヒモムシ類)がその姉妹群になると主張した[6]。またコーエンは、箒虫動物は腕足動物門のなかに含まれると述べている[8]。一方で、ダンらは腕足動物、箒虫動物、紐形動物の3群が単系統群になるという点では同じ結論に達しているが、腕足動物にもっとも近縁なのは箒虫動物ではなく、紐形動物と推定した[9]。遠藤は、ミトコンドリアゲノム上の遺伝子の順序に基づいて、腕足動物はむしろ環形動物やユムシ動物に近いと推定し、箒虫動物との近縁性に疑問を呈している[5]。
日本における腕足動物としては、舌殻綱シャミセンガイ目シャミセンガイ科Lingulidaeシャミセンガイ属Lingulaとして
の4種の生息が確認されている。[10]また、同目スズメガイダマシ科Discinidaeについても
の2種の生息が確認されている。 しかし、潮間帯を生息地とするシャミセンガイ類は、水質汚染や干拓により急速に生息地を減らしており、主産地である有明海においても、食用としての漁獲の対象にならないまでに個体数が減少しており、日本における絶滅は不可避である。イカリチョウチン(Craniscus japonicus)をはじめとする頭殻目、タテスジホオズキガイ(Coptothyris grayi)をはじめとする嘴殻目も日本近海の潮間帯から浅海底に生息するが、分布の詳細は明らかでない。
腕足動物の化石は顕生代のどの時代の地層にも見られる。古くは古生代カンブリア紀初期の地層から発見されている。はじめは無関節類しか見られないが、カンブリア紀中期になると有関節類も登場する。有関節類の登場以降、無関節類の数は減少し、デボン紀以降はわずかしか見られない。オルドビス紀には有関節類が多様化し、デボン紀にその多様性は最大となる。しかし古生代末(P-T境界)の大量絶滅で、腕足動物の多様性は大部分が失われた。その後、腕足動物はかつてのように繁栄することなく、衰退傾向にある[11]。腕足動物の化石種は1万3000種記載されているのに対し、現生種は350種程度に留まっている[2]。腕足動物の多様性の減少は、生態の似た二枚貝類との競争に敗れたためと考えられている[1]。
現生のミドリシャミセンガイを含むシャミセンガイ属は、生きている化石(古くから大きく形態を変化させることなく生きてきた生物)の代表例としてよく知られている[12]。これと同じ属の化石(異論はあるが、いずれにしてもよく似た形態を持つ化石)が、オルドビス紀・シルル紀から見つかっているためである。同じ属に分類されるほどではないが、現生種に似た化石はさらに遡り、カンブリア紀からも見つかっている。
グリーンランドのシリウス・パセットで見つかったハルキエリアの化石は、体の両端背側に殻を備えていた。サイモン・コンウェイ・モリス(英語版)は、この殻が腕足動物のものに似ていることを指摘し、腕足動物はハルキエリアのような動物から進化したと推測した。彼の考えによれば、両端に殻を持つハルキエリアの体が折りたたまれるように変化し、2枚の殻が向かい合うようになったことで、腕足動物の体制が起源したという[13]。彼はほかに、腕足動物の持つ剛毛がハルキエリアの持つ硬皮に由来すると考えられること、一部の現生の腕足動物では幼生が変態するときに、体が折りたたまれるようになることを根拠に挙げている。ハルキエリアの硬皮は環形動物の剛毛に由来するとも考えられることから、この仮説は腕足動物と環形動物が近縁であるとする仮説に整合する[4]。一方で、ハルキエリアを軟体動物に近いと考える意見もある[5]。
ところで、このコンウェイ・モリスの主張が正しければ、腕足動物の殻は2枚とも背側にある(腹側は内側に折り込まれる)ことになり、2枚の殻が背腹にあるという伝統的な考えは正しくないことになる[5]。
大プリニウスの『博物誌』には、「二形、白と黒、男と女という特徴を有する」石への言及がある。16世紀の古生物学者は、これに相当する化石を発見し、女性器を思わせるその外見から、子宮石と命名した(陰門石、外陰石などと呼ばれることもあった)。この化石は女性器のみならず、男性器のように見える部分も持ち、17世紀には、子宮の異常や男性の精力減退を改善すると信じられたこともあった。18世紀半ばまでには、子宮石は腕足動物の殻の内側に形成された雌型化石だと正しく認識されるようになった。女性器を思わせる割れ目は殻の内側の隆起を、男性器に見える突起は窪みを写し取ったものである[14]。
伝統的には有関節綱と無関節綱の2綱に大別されてきたが、3つの亜門(舌殻亜門、頭殻亜門、嘴殻亜門)に分類する体系が広く用いられるようになっている[8]。この体系では、嘴殻亜門は伝統的な分類の有関節綱に対応する。無関節綱は、キチン質性のリン酸カルシウムの殻を持つ舌殻亜門と、炭酸カルシウムの殻を持つ頭殻亜門に二分されている。
ほかに、殻の成分を重視して、炭酸カルシウムの殻を持つ頭殻類と嘴殻類をCalciataとしてまとめ、リン酸カルシウムの殻を持つ舌殻類と並べる体系も提案されている[15]。以下には、蝶番の有無による伝統的な2綱の体系と、3亜門の体系を示す。
以下は伝統的な体系の1例である[16]。同じ2綱に分ける体系でも、下位の分類については異なるものもある[17]。絶滅群は冒頭に†を付した。
以下は3亜門に分類する体系である[18]。なお、コーエンはホウキムシ類を腕足動物門の第4の亜門として含める体系を提案している[8]が、本項ではホウキムシ類は腕足動物に含めていない。
人間生活と直接に関わることは少ない。日本や東南アジアでは、ミドリシャミセンガイなどのシャミセンガイ属が食用にされている[19]。
腕足動物(わんそくどうぶつ、Brachiopoda)は、2枚の殻を持つ海産の底生無脊椎動物。シャミセンガイ、チョウチンガイなどと呼ばれるものを含む。一見して二枚貝に似るが、体制は大きく異なり、貝類を含む軟体動物門ではなく、独立の腕足動物門に分類される。
化石記録ではカンブリア紀に出現し、古生代を通じて繁栄したグループだが、その後多様性は減少し、現生の種数は比較的少ない。伝統的には無関節綱と有関節綱に分けられてきたが、それとは異なる分類体系も提案されている。
学名のBrachiopodaはギリシャ語で腕を意味するbrachiumと、足を意味するpodaを合わせたもので、和名はその直訳である。
_01.png)








