Brakiopoda, filum Brachiopoda, adalah hewan laut yang memiliki "katup" keras (cangkang) pada permukaan atas dan bawah, tidak seperti pengaturan kiri dan kanan di moluska bivalvia. Katup brakiopoda berengsel di bagian belakang, sedangkan bagian depan dapat dibuka untuk makan atau tertutup untuk perlindungan. Dua kelompok utama diakui, artikulata dan inartikulata. Brakiopoda artikulata memiliki engsel bergigi dan otot membuka dan menutup secara sederhana, sementara brakiopoda inartikulata memiliki engsel tanpa gigi dan sistem yang lebih kompleks dari otot yang digunakan untuk menjaga kedua bagian sejajar. Dalam brakiopoda, sebuah pedikel seperti tangkai menonjol dari sebuah lubang di salah satu katup, yang dikenal sebagai katup pedikel, menambatkan hewan ke dasar laut tetapi bebas dari lumpur yang akan menghalangi bukaan.
Kata "brakiopoda" dibentuk dari kata dalam bahasa Yunani Kuno βραχίων ("lengan") dan πούς ("kaki").[2] Mereka sering dikenal sebagai "kerang lentera", karena kerang melengkung kelas Terebratulida terlihat agak seperti lampu minyak tembikar.[1]
Rentang hidup berkisar dari tiga sampai lebih dari tiga puluh tahun. Gamet matang (ova atau sperma) mengapung dari gonad ke selom utama dan kemudian keluar ke dalam rongga mantel. Larva brachiopoda inartikulata adalah miniatur bentuk dewasa, dengan lofofor yang memungkinkan larva untuk mencari makan dan berenang selama berbulan-bulan sampai hewan menjadi cukup berat hidup di dasar laut. Larva planktonik spesies artikulata tidak menyerupai bentuk dewasa, melainkan terlihat seperti gumpalan dengan kantung kuning telur, dan tetap di antara plankton untuk hanya beberapa hari sebelum meninggalkan kolom air pada metamorfosis.
Selain klasifikasi tradisional Brachiopoda ke inartikulata dan artikulata, dua pendekatan muncul pada 1990-an: satu pendekatan mengelompokkan inartikulata Craniida dengan brakiopoda artikulata, karena keduanya menggunakan bahan yang sama di lapisan mineral dari cangkang mereka; pendekatan lain membuat Craniida kelompok ketiga, sebagai lapisan organik luar mereka berbeda dengan di salah satu dari dua lainnya. Namun, beberapa ahli taksonomi percaya terlalu dini untuk menyarankan tingkat yang lebih tinggi dari klasifikasi seperti ordo dan merekomendasikan pendekatan bawah-ke-atas yang mengidentifikasi genera dan kemudian kelompok-kelompok ini menjadi kelompok-kelompok menengah. Secara tradisional, brakiopoda telah dianggap sebagai anggota, atau sebagai kelompok saudara, dari Deuterostomia, sebuah superfilum yang mencakup chordata dan echinodermata. Salah satu jenis analisis hubungan evolusi dari Brachiopoda selalu menempatkan brakiopoda sebagai protostom sementara jenis lain telah dibagi antara menempatkan brakiopoda antara protostom atau deuterostom.
Disarankan pada tahun 2003 bahwa brakiopoda telah berevolusi dari nenek moyang mirip dengan Halkieria, hewan periode Kambrium seperti-siput dengan "zirah rantai" di punggungnya dan cangkang di depan dan bagian belakang; ia berpikir bahwa brakiopoda leluhur mengubah cangkangnya menjadi sepasang katup dengan melipat bagian belakang tubuhnya di bawah depannya. Namun, fosil baru ditemukan pada tahun 2007 dan 2008 menunjukkan bahwa "zirah rantai" dari tommotiid membentuk tabung dari hewan sesil; satu tommotiid mirip phoronida, yaitu kerabat dekat atau subkelompok brakiopoda, sedangkan tommotiid lainnya melahirkan dua piring simetris yang mungkin merupakan bentuk awal dari katup brakiopoda. Garis keturunan dari brakiopoda yang memiliki baik fosil dan taksa yang masih ada muncul di periode Kambrium awal, Ordovisium, dan Karbon. Garis keturunan lainnya telah muncul dan kemudian menjadi punah, kadang-kadang selama kepunahan massal parah. Di puncak mereka di era Paleozoikum, brakiopoda adalah hewan penyaring dan pembangun-karang yang paling melimpah, dan menduduki relung ekologi lainnya, termasuk berenang dengan gaya jet-propulsi dari kerang. Fosil brakiopoda telah menjadi indikator yang berguna dari perubahan iklim selama Paleozoikum. Namun, setelah peristiwa kepunahan Perm-Trias, brakiopoda pulih hanya sepertiga dari keanekaragaman mereka sebelumnya. Sebuah studi pada tahun 2007 menyimpulkan brakiopoda yang sangat rentan terhadap kepunahan Perm-Trias, karena mereka dibangun berkapur bagian keras (terbuat dari kalsium karbonat) dan memiliki tingkat metabolisme yang rendah dan sistem pernafasan lemah. Sering dianggap bahwa brakiopoda menurun setelah kepunahan Perm-Trias dan kalah bersaing dengan bivalvia, tetapi sebuah studi pada tahun 1980 menemukan spesies brakiopoda dan kerang meningkat dari Paleozoikum ke zaman modern, dengan bivalvia meningkat lebih cepat; setelah peristiwa kepunahan Perm-Trias, brakiopoda menjadi untuk pertama kalinya kurang beragam dari bivalvia.
Brakiopoda hanya hidup di laut, dan sebagian besar spesies menghindari lokasi dengan arus yang kuat atau gelombang. Larva spesies artikulata menetap dengan cepat dan membentuk populasi padat di daerah didefinisikan dengan baik sedangkan larva spesies inartikulata berenang untuk sampai satu bulan dan memiliki kisaran luas. Brakiopoda sekarang tinggal terutama di air dingin dan cahaya rendah. Ikan dan krustasea tampaknya menemukan daging brakiopoda tidak enak dan jarang menyerang mereka. Di antara brakiopoda, hanya lingulid (Lingula sp.[3]) telah dipancing secara komersial, pada skala yang sangat kecil. Salah satu spesies brakiopoda (Coptothyrus adamsi) mungkin menjadi ukuran kondisi lingkungan sekitar terminal minyak sedang dibangun di Rusia di pantai Laut Jepang.
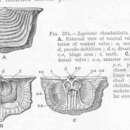
Brakiopoda modern panjangnya berkisar dari 1 sampai 100 mm, dan sebagian besar spesies sekitar 10 sampai 30 mm.[1] Brakiopoda terbesar yang diketahui - Gigantoproductus dan Titanaria, mencapai lebar 30 sampai 38 cm - hidup di akhir Karbon Awal.[4] Masing-masing memiliki dua katup (bagian cangkang) yang menutupi permukaan dorsal dan ventral hewan, tidak seperti moluska bivalvia yang cangkangnya menutupi permukaan lateral. Katup-katup itu disebut brakialis dan pedikel. Katup brakialis memiliki brachia ("lengan") di permukaan dalamnya yang menjadi nama dari filum ini, dan yang mendukung lofofor, digunakan untuk makan dan respirasi. Katup pedikel memiliki perlekatan ke pedikel seperti-tangkai di permukaan dalamnya; dengan pedikel, sebagian besar brakiopoda menempel pada substrat. (R. C. Moore, 1952) Katup brakialis dan pedikel sering disebut dorsal ("atas") dan ventral ("bawah"), tetapi beberapa ahli paleontologi menganggap istilah "dorsal" dan "ventral" tidak relevan karena mereka percaya bahwa katup "ventral" dibentuk oleh lipatan dari permukaan atas ke bagian bawah tubuh. Katup pedikel biasanya lebih besar dari katup brakialis. Dalam kebanyakan spesies brakiopoda artikulata, kedua katup itu cembung, permukaan sering memperlihatkan jalur pertumbuhan dan/atau ornamen lainnya. Namun, lingulid inartikulata, yang menggali ke dasar laut, memiliki katup yang halus, datar dan ukuran dan bentuk yang hampir sama. (R. C. Moore, 1952)
Brakiopoda artikulata ("bersendi") memiliki gigi dan pengaturan soket dimana katup pedikel dan brakialis berengsel, mengunci katup terhadap perpindahan lateral. Brakiopoda inartikulata tidak memiliki gigi yang cocok dan soket; katup mereka disatukan hanya dengan otot. (R. C. Moore, 1952)
Semua brakiopoda memiliki otot adduktor yang ditetapkan di dalam katup pedikel dan yang menutup katup dengan menarik pada bagian dari katup brakialis menjelang engsel. Otot-otot ini memiliki keduanya "cepat" serat yang menutup katup dalam keadaan darurat dan "menangkap" serat yang lebih lambat tetapi dapat menjaga katup ditutup untuk waktu yang lama. Brakiopoda artikulata membuka katup dengan cara otot abduktor, juga dikenal sebagai diduktor, yang terletak lebih jauh ke belakang dan tarik pada bagian katup brakialis balik engsel. Brakiopoda inartikulata menggunakan mekanisme membuka berbeda, di mana otot-otot mengurangi panjang selom (rongga tubuh utama) dan membuatnya menonjol keluar, mendorong katup terpisah. Kedua kelas membuka katup untuk sudut sekitar 10°. Set yang lebih kompleks dari otot yang digunakan oleh brakiopoda inartikulata juga dapat mengoperasikan katup gunting, mekanisme yang digunakan lingulid untuk menggali.[5]
Setiap katup terdiri dari tiga lapisan, sebuah periostrakum luar yang terbuat dari senyawa organik dan dua lapisan terbiomineralisasi. Brakiopoda artikulata memiliki periostrakum terluar terbuat dari protein, "lapisan utama" dari kalsit (bentuk dari kalsium karbonat) di bawah itu, dan terdalam campuran protein dan kalsit.[5] Cangkang brakiopoda tdk jelas memiliki urutan lapisan yang sama, tetapi komposisi mereka adalah berbeda dari brakiopoda artikulata dan juga bervariasi di antara kelas brakiopoda artikulata. Lingulid dan discinid, yang memiliki pedikel, memiliki matriks glikosaminoglikan (polisakarida tidak bercabang dan panjang), di mana bahan-bahan lain yang tertanam: kitin di periostrakum;[5] apatit yang mengandung kalsium fosfat dalam lapisan terbiomineralisasi utama;[6] dan campuran kompleks di lapisan terdalam, yang mengandung kolagen dan protein lainnya, kitinofosfat dan apatit.[5][7] Craniidae, yang tidak memiliki pedikel dan melekatkan dirinya langsung ke permukaan keras, memiliki periostrakum kitin dan lapisan mineral kalsit.[5][8] Cangkang bisa tumbuh secara holoperiferal, di mana materi baru ditambahkan di seluruh bagian pinggir, atau secara miksoperiferal, di mana materi baru ditambahkan ke daerah posterior dari cangkang dengan arah anterior, tumbuh ke arah cangkang lainnya.[9]
Brakiopoda, seperti moluska, memiliki mantel epitel yang mensekresikan dan melapisi cangkang, dan membungkus organ-organ internal. Tubuh brakiopoda hanya menempati sekitar sepertiga dari ruang internal di dalam cangkang, terdekat dengan engsel. Sisa ruang dilapisi dengan lobus mantel, ekstensi yang menutup ruang berisi air di mana lofofor berada.[5] Selom meluas ke masing-masing lobus sebagai jaringan kanal, yang membawa nutrisi ke tepi mantel.[10]
Sel yang relatif baru dalam alur pada tepi mantel mensekresikan material yang memperluas periostrakum. Sel-sel ini secara bertahap dipindah ke bawah mantel oleh sel yang lebih baru dalam alur, dan beralih ke mensekresikan bahan mineral dari katup cangkang. Dengan kata lain, di tepi katup periostrakum diperpanjang terlebih dahulu, dan kemudian diperkuat oleh perluasan lapisan mineral di bawahnya.[10] Dalam kebanyakan spesies tepi mantel juga memiliki bulu bergerak, sering disebut chaetae atau setae, yang dapat membantu mempertahankan hewan dan dapat bertindak sebagai sensor. Dalam beberapa brakiopoda sekelompok chaetae membantu untuk menyalurkan aliran air masuk dan keluar dari rongga mantel.[5]
Dalam kebanyakan brakiopoda, divertikula (ekstensi berongga) dari mantel menembus lapisan mineral katup ke periostraka. Fungsi dari divertikula ini tidak pasti dan disarankan bahwa mereka mungkin ruang penyimpanan untuk bahan kimia seperti glikogen, dapat mensekresi penolak untuk mencegah organisme menempel ke cangkang atau mungkin membantu dalam respirasi.[5] Percobaan menunjukkan bahwa konsumsi oksigen brakiopoda menurun jika jeli petroleum dioleskan pada cangkang, menyumbat divertikula.[10]
Seperti bryozoa dan phoronida, brakiopoda memiliki lofofor, mahkota tentakel di mana silia (rambut-rambut halus) membuat arus air yang memungkinkan mereka untuk menyaring partikel makanan dari air. Namun lofofor bryozoa atau phoronida adalah cincin dari tentakel terpasang pada tangkai tunggal yang dapat ditarik,[11][12] sedangkan bentuk dasar dari lofofor brakiopoda adalah berbentuk U, membentuk brachia ("lengan") yang menjad asal dari nama filum.[5] Lofofor brakiopoda tidak dapat ditarik dan menempati hingga dua-pertiga dari ruang internal, di daerah paling depan katup menganga ketika dibuka. Untuk menyediakan kapasitas penyaringan yang cukup dalam ruang terbatas ini, lofofor dari brakiopoda yang lebih besar dilipat dalam bentuk yang sedang sampai sangat kompleks-loop dan gulungan umum, dan lofofor beberapa spesies menyerupai tangan dengan jari-jari terentang.[5] Dalam semua spesies lofofor didukung oleh kartilago dan oleh rangka hidrostatik (dengan kata lain oleh tekanan dari cairan internal),[10] dan cairan mencapai tentakel.[5] Beberapa brakiopoda artikulata juga memiliki brachidium, dukungan berkapur untuk lofofor yang melekat pada bagian dalam katup brakialis.[10]
Tentakel menanggung silia (rambut halus dapat bergerak) di tepi dan di sepanjang pusat. Pemukulan silia luar menggerakkan arus air dari ujung tentakel ke dasar, di mana air itu keluar. Partikel makanan yang bertabrakan dengan tentakel terjebak oleh lendir, dan silia di bawah bagian tengah menggerakkan campuran ini ke dasar tentakel.[13] Sebuah alur brakialis mengelilingi dasar dari tentakel, dan silianya sendiri melewatkan makanan di sepanjang alur menuju mulut.[5] Metode yang digunakan oleh brakiopoda dikenal sebagai "mengumpulkan ke hulu", karena partikel makanan ditangkap saat mereka memasuki bidang silia yang menciptakan arus makan. Metode ini digunakan oleh phoronida dan bryozoa, dan juga oleh pterobranchia. Entoprocta menggunakan mahkota tentakel yang hampir sama, tetapi padat dan aliran berjalan dari dasar ke ujung, membentuk sistem "mengumpulkan ke hilir" yang menangkap partikel makanan karena mereka akan keluar.[14]
Kebanyakan spesies modern menempel pada permukaan keras dengan pedikel ("tangkai") silindris, merupakan perpanjangan dari dinding tubuh. Pedikel memiliki kutikula berkitin ("kulit" non-seluler) dan menonjol melalui pembukaan di engsel.[5] Namun, beberapa genera seperti inartikulata Crania dan artikulata Lacazella tidak memiliki pedikel, dan melekatkan bagian belakang katup "pedikel" ke permukaan sehingga bagian depan sedikit naik dari permukaan.[1][5] Dalam beberapa genera artikulata seperti Neothyris dan Anakinetica, pedikel layu ketika dewasa tumbuh dan akhirnya berbaring longgar di permukaan. Dalam genera ini cangkang menebal dan berbentuk sehingga pembukaan katup yang menganga tetap bebas dari sedimen.[1]
Pedikel spesies inartikulata adalah ekstensi dari selom utama, yang merupakan tempat organ internal. Sebuah lapisan otot memanjang melapisi epidermis dari pedikel tersebut.[5] Anggota ordo Lingulida memiliki pedikel panjang, yang mereka gunakan untuk menggali ke dalam substrat lunak, untuk mengangkat cangkang ke pembukaan liang untuk mencari makan, dan untuk menarik kembali cangkang ketika terganggu.[10] Lingulid menggerakkan tubuhnya naik turun pada bagian dua pertiga atas dari liang, sementara sepertiga sisanya ditempati hanya oleh pedikel, dengan bohlam di ujung yang membangun tambatan "konkret".[15] Namun, pedikel dari ordo Discinida pendek dan menempel pada permukaan keras.[5]
Sebuah pedikel artikulata tidak memiliki selom, dibangun dari bagian yang berbeda dari tubuh larva, dan memiliki inti yang terdiri dari jaringan ikat. Otot di bagian belakang tubuh dapat meluruskan, membengkokkan atau bahkan memutar pedikel. Ujung dari pedikel umumnya memiliki ekstensi seperti akar atau papila pendek ("benjolan"), yang menempel pada permukaan keras. Namun, brakiopoda artikulata dari genus Chlidonophora menggunakan pedikel bercabang untuk berjangkar di sedimen. Pedikel muncul dari katup pedikel, baik melalui takik di engsel atau, dalam spesies di mana katup pedikel lebih panjang dari katup brakialis, dari lubang di mana katup pedikel ganda kembali menyentuh katup brakialis. Beberapa spesies berdiri dengan ujung depan ke atas, sementara yang lain berbaring horisontal dengan katup pedikel paling atas.[5]
Aliran air memasuki lofofor dari sisi katup terbuka dan keluar di bagian depan binatang. Dalam lingulid pintu masuk dan keluar saluran dibentuk oleh kelompok chaetae yang berfungsi sebagai corong.[5] Pada brakiopoda lainnya saluran masuk dan keluar diatur oleh bentuk lofofor.[10] Lofofor menangkap partikel makanan, terutama fitoplankton (organisme fotosintetik kecil), dan mengantarkan mereka ke mulut melalui alur brakialis sepanjang dasar dari tentakel.[5] Mulut ada di dasar lofofor.[16] Makanan melewati mulut, faring berotot ("tenggorokan") dan esofagus ("kerongkongan"),[5] yang semuanya dilapisi dengan silia dan sel-sel yang mengeluarkan mukus dan enzim pencernaan.[10] Dinding lambung memiliki seka ("kantong") bercabang di mana makanan dicerna, terutama dalam sel.[5]
Nutrisi diangkut oleh selom, termasuk lobus mantel, oleh silia.[10] Limbah yang dihasilkan oleh metabolisme dipecah menjadi amonia, yang dieliminasi oleh difusi melalui mantel dan lofofor.[5] Brakiopoda memiliki metanefridia, digunakan oleh banyak filum untuk mengekskresikan amonia dan limbah terlarut lainnya. Namun, brakiopoda tidak memiliki podosit, yang melakukan tahap pertama ekskresi dalam proses ini,[17] dan metanefridia brakiopoda tampaknya hanya digunakan untuk mengeluarkan sperma dan ovum.[5]
Mayoritas makanan yang dikonsumsi oleh brakiopoda dapat dicerna, dengan limbah padat yang dihasilkan sangat sedikit.[18] Silia dari lofofor dapat mengubah arah untuk mengeluarkan partikel terisolasi materi tak dicerna. Jika hewan menemukan gumpalan besar materi yang tidak diinginkan, silia yang melapisi saluran masuk berhenti dan tentakel yang berkontak dengan gumpalan bergerak terpisah untuk membentuk kesenjangan besar dan kemudian perlahan-lahan menggunakan silia mereka untuk membuang gumpalan ke lapisan mantel. Lapisan mantel memiliki silia sendiri, yang membersihkan gumpalan keluar melalui pembukaan antara katup. Jika lofofor tersumbat, aduktor menutup katup dengan tajam, yang menciptakan "bersin" yang membersihkan penghalang.[10] Dalam beberapa brakiopoda inartikulata saluran pencernaan adalah berbentuk U dan berakhir dengan anus yang menghilangkan padatan dari bagian depan dinding tubuh.[16] Brakiopoda inartikulata lain dan semua brakiopoda artikulata memiliki usus melengkung yang berakhir membabi buta, tanpa anus.[5] Hewan ini menggabungkan limbah padat dengan mukus dan secara berkala "membersinkan" itu, menggunakan kontraksi tajam otot usus.[10]
Lofofor dan mantel adalah satu-satunya permukaan yang menyerap oksigen dan melepaskan karbon dioksida. Oksigen tampaknya didistribusikan oleh cairan dari selom, yang beredar melalui mantel dan didorong baik oleh kontraksi lapisan selom atau dengan gerakan silia. Dalam beberapa spesies sebagian oksigen dibawa oleh pigmen pernapasan hemeritrin, yang diangkut dalam sel selomosit.[5] Konsumsi oksigen maksimum dari brakiopoda rendah, dan persyaratan minimum mereka tidak terukur.
Brakiopoda juga memiliki darah tak berwarna, diedarkan oleh jantung berotot yang ada di bagian dorsal tubuh di atas perut.[5] Darah melewati pembuluh yang mencapai bagian depan dan belakang tubuh, dan bercabang ke organ-organ termasuk lofofor di depan dan usus, otot, gonad dan nefridia di bagian belakang. Sirkulasi darah tampaknya tidak benar-benar tertutup, dan cairan selom dan darah setidaknya bercampur untuk suatu derajat.[10] Fungsi utama dari darah mungkin untuk membawa nutrien.[5]
"Otak" dari artikulata dewasa terdiri dari dua ganglia, satu di atas dan yang lainnya di bawah kerongkongan. Inartikulata dewasa hanya memiliki ganglion bawah.[19] Dari ganglia dan komisura di mana mereka bergabung, saraf lari ke lofofor, lobus mantel dan otot-otot yang mengoperasikan katup. Tepi mantel mungkin memiliki konsentrasi terbesar sensor. Meskipun tidak secara langsung terhubung ke neuron sensorik, chaetae mantel itu mungkin mengirim sinyal taktil ke reseptor pada epidermis mantel. Banyak brakiopoda menutup katup mereka jika bayangan muncul di atas mereka, tetapi sel-sel yang bertanggung jawab untuk ini tidak diketahui. Beberapa Brachiopoda memiliki statokista, yang mendeteksi perubahan posisinya.[5]
Rentang hidup berkisar dari 3 sampai lebih dari 30 tahun.[1] Dewasa dari sebagian besar spesies adalah dari satu jenis kelamin sepanjang hidup mereka. Gonad adalah massa gamet (ovum atau sperma) yang sedang berkembang, dan sebagian besar spesies memiliki empat gonad, dua di setiap katup.[5] Gonad pada artikulata terletak pada saluran lobus mantel, sedangkan pada inartikulata beradaa di dekat usus.[10] Gamet matang mengapung ke dalam selom utama dan kemudian keluar ke dalam rongga mantel melalui metanefridia, yang terbuka pada kedua sisi mulut. Sebagian besar spesies melepaskan baik ovum dan sperma ke dalam air, tetapi betina dari beberapa spesies menjaga embrio di ruang induk sampai larva menetas.[5]
Pembelahan sel dalam embrio adalah radial (sel terbentuk di tumpukan cincin langsung di atas satu sama lain), holoblastik (sel terpisah, meskipun berdampingan) dan regulatif (jenis jaringan di mana sel berkembang dikendalikan oleh interaksi antara sel-sel yang berdekatan, bukannya kaku dalam setiap sel).[20][5] Sementara beberapa hewan mengembangkan mulut dan anus dengan memperdalam blastopor, sebuah "penyok" di permukaan embrio awal, blastopor dari brakiopoda menutup, dan mulut dan dubur mereka berkembang dari bukaan baru. [5]
Larva inartikulata berenang sebagai plankton selama berbulan-bulan[1] dan seperti miniatur dewasa, dengan katup, lobus mantel, pedikel yang tergulung di dalam rongga mantel, dan lofofor kecil, yang digunakan untuk makan dan berenang[5]—kecuali bahwa Craniidae tidak memiliki pedikel.[10] Ketika cangkang menjadi lebih berat, juvenil tenggelam ke bawah dan menjadi dewasa sesil.[5] Larva spesies artikulata hidup hanya pada kuning telur, dan tetap di antara plankton untuk hanya beberapa hari.[1] Jenis larva ini memiliki lobus paling depan bersilia yang menjadi tubuh dan lofofor, lobus belakang yang menjadi pedikel, dan mantel seperti rok, dengan hem ke arah belakang. Pada metamorfosis menjadi dewasa, pedikel menempel ke permukaan, lobus depan mengembangkan lofofor dan organ lainnya, dan mantel menggulung di atas lobus depan dan mulai mensekresi cangkang.[5] Di laut dingin, pertumbuhan brakiopoda musiman dan berat badannya sering turun di musim dingin. Variasi dalam pertumbuhan ini sering membentuk garis pertumbuhan kerang. Anggota beberapa genera telah bertahan selama satu tahun di akuarium tanpa makanan.[1]
Genera fosil brakiopoda memiliki keragaman besar tetapi hanya sedikit karakteristik rangka, sedangkan genera modern memiliki keragaman jauh lebih rendah tetapi memberikan karakteristik bertubuh lunak serta kerangka—dan kedua set spesimen memiliki keterbatasan yang membuatnya sulit untuk menghasilkan klasifikasi komprehensif brakiopoda. Filum ini juga telah mengalami evolusi konvergen yang signifikan dan pembalikan (di mana kelompok yang lebih baru tampaknya telah kehilangan ciri khas yang terlihat dalam kelompok sebelumnya, kembali ke karakteristik yang terakhir terlihat pada kelompok yang lebih tua). Oleh karena itu beberapa ahli taksonomi brakiopoda percaya terlalu dini untuk menentukan tingkat yang lebih tinggi dari klasifikasi seperti ordo, dan merekomendasikan pendekatan bawah-ke-atas yang mengidentifikasi genera dan kemudian kelompok-kelompok ini menjadi kelompok-kelompok menengah.[21]
Namun, ahli taksonomi lainnya percaya bahwa beberapa pola karakteristik cukup stabil untuk membuat tingkat klasifikasi yang lebih tinggi berharga, meskipun ada pandangan yang berbeda tentang apakah tingkat klasifikasi yang lebih tinggi harus ada.[21] Klasifikasi "tradisional" didefinisikan pada tahun 1869; dua pendekatan lebih lanjut didirikan pada 1990-an:[7][22]
Sekitar 330 spesies yang hidup diakui,[7] dikelompokkan ke dalam lebih dari 100 genera. Sebagian besar dari brakiopoda modern adalah rhynchonelliformea (Articulata, tetapi tidak meliputi Craniida).[1]
Brakiopoda hanya hidup di laut. Sebagian besar spesies menghindari lokasi dengan arus yang kuat atau gelombang, dan situs khas termasuk overhang berbatu, celah-celah dan gua-gua, lereng curam landas benua, dan di bagian bawah lautan dalam. Namun, beberapa spesies artikulata melekat pada rumput laut atau di situs yang sangat terlindung di zona intertidal. Brakiopoda terkecil yang masih hidup, Gwynia, panjangnya hanya sekitar 1 mm, dan tinggal di kerikil.[1] Rhynchonelliformea (Articulata tidak mencakup Craniida), yang larvanya mengkonsumsi hanya kuning telurnya dan menetap dan berkembang dengan cepat, mengkhususkan diri di daerah tertentu dan membentuk populasi padat yang bisa mencapai ribuan individu per meter. Dewasa muda sering menempel pada cangkang hewan yang lebih matang. Di sisi lain, brakiopoda inartikulata, yang larvanya berenang untuk sampai satu bulan sebelum menetap, memiliki kisaran luas. Anggota dari genus diskinoid Pelagodiscus memiliki distribusi kosmopolitan.[1]
Tingkat metabolisme brakiopoda adalah antara sepertiga dan sepersepuluh dari bivalvia. Sementara brakiopoda berlimpah di laut hangat dan dangkal selama periode Kapur, mereka kalah berkompetisi dari bivalvia, dan sekarang tinggal terutama dalam kondisi dingin dan cahaya gelap.[25]
Kerang brakiopoda sesekali menunjukkan bukti kerusakan oleh predator, dan kadang-kadang perbaikannya. Ikan dan krustasea tampaknya menemukan daging brakiopoda tidak enak.[1] Catatan fosil menunjukkan bahwa predator pengebor seperti gastropoda menyerang moluska dan bulu babi 10 sampai 20 kali lebih sering daripada pada brakiopoda, menunjukkan bahwa predator menyerang brakiopoda karena kesalahan atau ketika mangsa lain langka.[26] Di perairan di mana makanan langka, siput Capulus ungaricus mencuri makanan dari bivalvia, siput, cacing tabung, dan brakiopoda. [27]
Di antara brakiopoda hanya lingulid yang telah dipancing secara komersial, dan hanya pada skala yang sangat kecil.[28] Brakiopoda jarang menetap pada permukaan buatan, mungkin karena mereka rentan terhadap polusi. Hal ini dapat membuat populasi Coptothyrus adamsi berguna sebagai ukuran kondisi lingkungan di sekitar terminal minyak yang sedang dibangun di Rusia di pantai Laut Jepang.[29]
Lebih dari 12.000 spesies fosil diakui,[7] dikelompokkan ke dalam lebih dari 5.000 marga. Sementara brakiopoda modern yang terbesar panjangnya 100 milimeter,[1] beberapa fosil mencapai lebar 200 milimeter.[31] Brakiopoda paling awal yang dikonfirmasi telah ditemukan dari periode Kambrium awal, bentuk inartikulata muncul pertama, diikuti segera setelahnya oleh bentuk artikulata.[32] Tiga spesies tidak termineralisasi juga telah ditemukan di Kambrium, dan tampaknya mewakili dua kelompok berbeda yang berevolusi dari nenek moyang termineralisasi.[33] Inartikulata Lingula sering disebut "fosil hidup", karena genera yang sangat mirip telah ditemukan dari periode Ordovisium. Di sisi lain, brakiopoda artikulata telah berdiversifikasi besar-besaran, dan mengalami kepunahan massal parah[34]—tapi artikulata Rhynchonellida dan Terebratulida, kelompok masa kini yang paling beragam, muncul pada awal Ordovisium dan Karbon, berturut-turut.[30]
Sejak tahun 1991 Claus Nielsen telah mengajukan hipotesis tentang perkembangan brakiopoda, diadaptasi pada tahun 2003 oleh Cohen dan rekan-rekannya sebagai hipotesis tentang evolusi awal Brachiopoda. Hipotesis "lipatan brakiopoda" ini menunjukkan bahwa brakiopoda berevolusi dari nenek moyang yang mirip dengan Halkieria,[16] hewan seperti-siput dengan "zirah rantai" di punggungnya dan cangkang di bagian depan dan belakang.[35] Hipotesis ini mengusulkan bahwa brakiopoda pertama mengubah cangkangnya menjadi sepasang katup dengan melipat bagian belakang tubuhnya di bawah depannya.[16]
Namun, fosil dari tahun 2007 dan setelahnya telah mendukung interpretasi baru dari tommotiid Kambrium Awal dan hipotesis baru bahwa brakiopoda berevolusi dari tommotiid. "Armor mail" dari tommotiid adalah terkenal tetapi tidak dalam bentuk dirakit, dan secara umum diasumsikan bahwa tommotiid adalah hewan seperti-siput yang mirip dengan Halkieria, kecuali bahwa baju besi tommotiid dibuat dari senyawa organofosfatik sedangkan milik Halkieria dibuat dari kalsit. Namun fosil dari tommotiid baru, Eccentrotheca, menunjukkan zirah mantel dirakit yang membentuk tabung, yang menunjukkan bahwa binatang itu sesil, bukannya merayap seperti siput. Tabung organofosfatik Eccentrotheca menyerupai milik phoronida,[36] hewan sesil yang makan dengan lofofor dan dianggap baik kerabat dekat atau sub-kelompok brakiopoda.[37] Paterimitra, fosil lain sebagian besar dirakit ditemukan pada tahun 2008 dan dijelaskan pada tahun 2009, memiliki dua piring simetris di bagian bawah, seperti katup brakiopoda tetapi tidak sepenuhnya memenutup tubuhnya.[38]
Di puncak mereka di era Paleozoikum, brakiopoda adalah hewan penyaring dan pembangun-karang paling melimpah,[39] dan menempati relung ekologi lainnya, termasuk berenang dengan gaya jet-propulsi dari kerang.[1] Namun, setelah peristiwa kepunahan Perm-Trias, secara informal dikenal sebagai "Great Dying",[39] brakiopoda pulih hanya sepertiga dari keanekaragaman mereka sebelumnya.[39] Hal ini sering berpikir bahwa brakiopoda benar-benar menurun dalam keragaman, dan bahwa dalam beberapa cara bivalvia mengalahkan mereka. Namun, pada tahun 1980 Gould dan Calloway menghasilkan analisis statistik yang menyimpulkan bahwa: baik brakiopoda dan bivalvia meningkat dari Paleozoikum ke zaman modern, tetapi bivalvia meningkat lebih cepat; kepunahan Perm-Trias itu cukup parah untuk bivalvia tetapi menghancurkan untuk brakiopoda, sehingga brakiopoda untuk pertama kalinya kurang beragam dari bivalvia dan keragaman mereka setelah Permian meningkat dari dasar yang sangat rendah; tidak ada bukti bahwa bivalvia mengalahkan brakiopoda, dan peningkatan atau penurunan jangka pendek untuk kedua kelompok muncul pada waktu yang sama.[40] Pada tahun 2007 Knoll dan Bambach menyimpulkan bahwa brakiopoda adalah salah satu dari beberapa kelompok yang paling rentan terhadap kepunahan Perm-Trias, karena semua memiliki bagian keras berkapur (terbuat dari kalsium karbonat) dan memiliki tingkat metabolisme yang rendah dan sistem pernapasan lemah.[41]
Fosil brakiopoda adalah indikator yang berguna dari perubahan iklim selama era Paleozoikum. Ketika suhu global yang rendah, seperti dalam sebagian besar dari Ordovisium, perbedaan besar dalam suhu antara khatulistiwa dan kutub menciptakan koleksi yang berbeda dari fosil di lintang yang berbeda. Di sisi lain, periode hangat, seperti pada Silur, menciptakan perbedaan kecil dalam suhu, dan semua lautan di lintang rendah sampai menengah dikolonisasi oleh beberapa spesies brakiopoda yang sama.[42]
Dari sekitar tahun 1940-an ke 1990-an, pohon keluarga berdasarkan fitur embriologis dan morfologi menempatkan brakiopoda di dalam atau sebagai kelompok saudara dari Deuterostomia,[43][44] superfilum yang mencakup chordata dan echinodermata.[45] Pemeriksaan lebih dekat menemui kesulitan di dasar dari brakiopoda berafiliasi dengan deuterostom:[46]
Nielsen memandang brakiopoda dan phoronida yang berkerabat dekat berafiliasi dengan deuterostoma pterobranchia karena lofofor mereka didorong oleh satu silium per sel, sedangkan yang dari bryozoa, yang dianggapnya sebagai protostom, memiliki beberapa silia per sel.[51] Namun, pterobranchia adalah hemichordata dan mungkin berkaitan erat dengan echinodermata, dan tidak ada bukti bahwa nenek moyang terbaru dari pterobranchia dan hemichordata lain atau nenek moyang terbaru dari hemichordata dan echinodermata adalah sesil dan makan dengan tentakel.[46]
Dari tahun 1988 dan seterusnya analisis berdasarkan filogeni molekuler, yang membandingkan fitur biokimia seperti kesamaan dalam DNA, telah menempatkan brakiopoda dalam Lophotrochozoa, sebuah superfilum protostom yang meliputi moluska, Annelida dan cacing pipih tetapi tidak termasuk superfilum protostom Ecdysozoa, yang anggotanya termasuk artropoda.[43][46] Kesimpulan ini bulat antara studi filogeni molekuler, yang menggunakan berbagai pilihan gen: rDNA, gen Hox, gen protein mitokondria, gen protein nukleus tunggal dan set gen protein nukleus.[52]
Beberapa penelitian gabungan pada tahun 2000 dan 2001, menggunakan kedua data molekuler dan morfologi, mendukung brakiopoda sebagai Lophotrochozoa,[53][54] sementara yang lain pada tahun 1998 dan 2004 menyimpulkan bahwa brakiopoda adalah deuterostom.[52]
Phoronida makan dengan lofofor, meliang atau menatah pada permukaan, dan membangun tabung tiga-lapis yang terbuat dari polisakarida, mungkin kitin, dicampur dengan partikel dengan bahan dari dasar laut. Secara tradisional mereka telah dianggap sebagai filum terpisah, tetapi studi filogeni molekuler semakin rinci antara tahun 1997 dan 2000 telah menyimpulkan bahwa phoronida adalah sub-kelompok brakiopoda.[37] Namun, sebuah analisis pada tahun 2005 menyimpulkan bahwa phoronida adalah sub-kelompok bryozoa.[55]
Sementara semua studi filogeni molekuler dan setengah studi gabungan hingga 2008 menyimpulkan bahwa brakiopoda adalah lophotrochozoa, mereka tidak bisa mengidentifikasi filum lophotrochozoa yang merupakan kerabat terdekat dari brakiopoda—kecuali phoronida, yang merupakan sub-kelompok Brachiopoda.[37][52] Namun, pada tahun 2008 dua analisis menemukan bahwa kerabat terdekat lophotrochozoa brakiopoda adalah nemertea. Para penulis menemukan ini mengejutkan, karena nemertea mengalami penyibakan spiral pada tahap awal pembelahan sel dan membentuk larva trokofor, sementara brakiopoda memiliki penyibakan radial dan larva yang tidak menunjukkan tanda telah berevolusi dari trokofor.[56][57] Studi lain pada tahun 2008 juga menyimpulkan bahwa brakiopoda berhubungan erat dengan nemertea, meragukan gagasan bahwa brakiopoda adalah bagian dari klad Lophophorata dalam lophotrochozoa.[52]
Morfologi brakiopoda
Sebuah brakiopoda Karbon Neospirifer condor, dari Bolivia. Spesimen adalah 7 cm.
Rhynchotrema dentatum, sebuah brakiopoda Rhynchonellida dari Cincinnatian (Ordovisium Atas) dari tenggara Indiana.
Sebuah brakiopoda spiriferid Devonian dari Ohio yang menjadi inang substrat untuk koloni hederellid. Spesimen lebar 5 cm.
Syringothyris sp.; sebuah brakiopoda spiriferid dari Formasi Logan (Karbon Bawah) dari Wooster, Ohio (cetakan internal).
Brakiopoda Petrocrania melekat pada brakiopoda strophomenid; Atas Ordovician dari tenggara Indiana.
Lingula ditemukan di dekat Ozamis City, Filipina.
Cetakan brakiopoda di Formasi Lock Haven
Hercosestria cribrosa Cooper & Grant 1969 (Roadian, Guadalupian, Permian Tengah); Glass Mountains, Texas.
Interior katup ventral brakiopoda productid; Roadian, Guadalupian (Permian Tengah); Glass Mountains, Texas.
Terebratella sanguinea (Leach, 1814)
|accessdate= (bantuan)
|date= (bantuan)
Brakiopoda, filum Brachiopoda, adalah hewan laut yang memiliki "katup" keras (cangkang) pada permukaan atas dan bawah, tidak seperti pengaturan kiri dan kanan di moluska bivalvia. Katup brakiopoda berengsel di bagian belakang, sedangkan bagian depan dapat dibuka untuk makan atau tertutup untuk perlindungan. Dua kelompok utama diakui, artikulata dan inartikulata. Brakiopoda artikulata memiliki engsel bergigi dan otot membuka dan menutup secara sederhana, sementara brakiopoda inartikulata memiliki engsel tanpa gigi dan sistem yang lebih kompleks dari otot yang digunakan untuk menjaga kedua bagian sejajar. Dalam brakiopoda, sebuah pedikel seperti tangkai menonjol dari sebuah lubang di salah satu katup, yang dikenal sebagai katup pedikel, menambatkan hewan ke dasar laut tetapi bebas dari lumpur yang akan menghalangi bukaan.
Kata "brakiopoda" dibentuk dari kata dalam bahasa Yunani Kuno βραχίων ("lengan") dan πούς ("kaki"). Mereka sering dikenal sebagai "kerang lentera", karena kerang melengkung kelas Terebratulida terlihat agak seperti lampu minyak tembikar.
Rentang hidup berkisar dari tiga sampai lebih dari tiga puluh tahun. Gamet matang (ova atau sperma) mengapung dari gonad ke selom utama dan kemudian keluar ke dalam rongga mantel. Larva brachiopoda inartikulata adalah miniatur bentuk dewasa, dengan lofofor yang memungkinkan larva untuk mencari makan dan berenang selama berbulan-bulan sampai hewan menjadi cukup berat hidup di dasar laut. Larva planktonik spesies artikulata tidak menyerupai bentuk dewasa, melainkan terlihat seperti gumpalan dengan kantung kuning telur, dan tetap di antara plankton untuk hanya beberapa hari sebelum meninggalkan kolom air pada metamorfosis.
Selain klasifikasi tradisional Brachiopoda ke inartikulata dan artikulata, dua pendekatan muncul pada 1990-an: satu pendekatan mengelompokkan inartikulata Craniida dengan brakiopoda artikulata, karena keduanya menggunakan bahan yang sama di lapisan mineral dari cangkang mereka; pendekatan lain membuat Craniida kelompok ketiga, sebagai lapisan organik luar mereka berbeda dengan di salah satu dari dua lainnya. Namun, beberapa ahli taksonomi percaya terlalu dini untuk menyarankan tingkat yang lebih tinggi dari klasifikasi seperti ordo dan merekomendasikan pendekatan bawah-ke-atas yang mengidentifikasi genera dan kemudian kelompok-kelompok ini menjadi kelompok-kelompok menengah. Secara tradisional, brakiopoda telah dianggap sebagai anggota, atau sebagai kelompok saudara, dari Deuterostomia, sebuah superfilum yang mencakup chordata dan echinodermata. Salah satu jenis analisis hubungan evolusi dari Brachiopoda selalu menempatkan brakiopoda sebagai protostom sementara jenis lain telah dibagi antara menempatkan brakiopoda antara protostom atau deuterostom.
Disarankan pada tahun 2003 bahwa brakiopoda telah berevolusi dari nenek moyang mirip dengan Halkieria, hewan periode Kambrium seperti-siput dengan "zirah rantai" di punggungnya dan cangkang di depan dan bagian belakang; ia berpikir bahwa brakiopoda leluhur mengubah cangkangnya menjadi sepasang katup dengan melipat bagian belakang tubuhnya di bawah depannya. Namun, fosil baru ditemukan pada tahun 2007 dan 2008 menunjukkan bahwa "zirah rantai" dari tommotiid membentuk tabung dari hewan sesil; satu tommotiid mirip phoronida, yaitu kerabat dekat atau subkelompok brakiopoda, sedangkan tommotiid lainnya melahirkan dua piring simetris yang mungkin merupakan bentuk awal dari katup brakiopoda. Garis keturunan dari brakiopoda yang memiliki baik fosil dan taksa yang masih ada muncul di periode Kambrium awal, Ordovisium, dan Karbon. Garis keturunan lainnya telah muncul dan kemudian menjadi punah, kadang-kadang selama kepunahan massal parah. Di puncak mereka di era Paleozoikum, brakiopoda adalah hewan penyaring dan pembangun-karang yang paling melimpah, dan menduduki relung ekologi lainnya, termasuk berenang dengan gaya jet-propulsi dari kerang. Fosil brakiopoda telah menjadi indikator yang berguna dari perubahan iklim selama Paleozoikum. Namun, setelah peristiwa kepunahan Perm-Trias, brakiopoda pulih hanya sepertiga dari keanekaragaman mereka sebelumnya. Sebuah studi pada tahun 2007 menyimpulkan brakiopoda yang sangat rentan terhadap kepunahan Perm-Trias, karena mereka dibangun berkapur bagian keras (terbuat dari kalsium karbonat) dan memiliki tingkat metabolisme yang rendah dan sistem pernafasan lemah. Sering dianggap bahwa brakiopoda menurun setelah kepunahan Perm-Trias dan kalah bersaing dengan bivalvia, tetapi sebuah studi pada tahun 1980 menemukan spesies brakiopoda dan kerang meningkat dari Paleozoikum ke zaman modern, dengan bivalvia meningkat lebih cepat; setelah peristiwa kepunahan Perm-Trias, brakiopoda menjadi untuk pertama kalinya kurang beragam dari bivalvia.
Brakiopoda hanya hidup di laut, dan sebagian besar spesies menghindari lokasi dengan arus yang kuat atau gelombang. Larva spesies artikulata menetap dengan cepat dan membentuk populasi padat di daerah didefinisikan dengan baik sedangkan larva spesies inartikulata berenang untuk sampai satu bulan dan memiliki kisaran luas. Brakiopoda sekarang tinggal terutama di air dingin dan cahaya rendah. Ikan dan krustasea tampaknya menemukan daging brakiopoda tidak enak dan jarang menyerang mereka. Di antara brakiopoda, hanya lingulid (Lingula sp.) telah dipancing secara komersial, pada skala yang sangat kecil. Salah satu spesies brakiopoda (Coptothyrus adamsi) mungkin menjadi ukuran kondisi lingkungan sekitar terminal minyak sedang dibangun di Rusia di pantai Laut Jepang.
_01.png)




.jpg)

_slab_2.jpg)











