الأسماء في صفحات التنقل
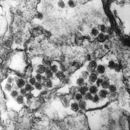
Віріони ретровірусів сферичної форми, розміром 90 — 120 нм, мають суперкапсиду ліпопротеїнової природи з шипоподібними пепломерами. Капсид побудований за типом кубічної симетрії. Ретровіруси містять до 2% РНК, близько 65% білку , 30% ліпідів та 2% вуглеводів. Характерною структурною особливістю у віріонів ретровірусів є унікальна серцевина. Остання представлена власною капсулою (внутрішня білкова мембрана) та нуклеотидом, до складу якого входить РНК і білок. За будовою серцевини онковіруси поділяються на три типи: С, В та D. Віріони типу С (діаметр близько 100 нм.) мають крупну (74-80 нм.), розташовану центрально, серцевину. У віріонів типу В і D серцевини розташовані ексцентрично. У перших вона округлої а у останніх циліндричної форми.
Вірус термолабільний — при 56˚С руйнується протягом 15-ти хвилин, при 74˚С. за 16 секунд, чутливий до ефіру і хлороформу, руйнується при заморожуванні — відтаванні.
Геном ретровірусів, на відміну від всіх останніх представників царства вірусів, диплоїдний — представлений двома однаковими молекулами РНК, молекулярною масою близько 3 МД. Розмір геному 7-11 kb. Він містить чотири гени : (5'→3') gag, pol, env, src.
Стратегія реплікації ядерна:
У складі ретровірусів виявляється від 11 до 13 структурних білків. Найменше білків локалізовано у серцевині. Головний внутрішній білок Р27-Р30 розташований у мембрані, що оточує РНК. Два - п'ять білків гліколізовані і знаходяться у складі ліпопротеїнової оболонки. З ними пов'язана типоспецифічна характеристика вірусів. Групоспецифічні антигени знаходяться у серцевині вірусу. Основним групоспецифічним антигеном є головний внутрішній білок Р27 - Р30. У віріонах онковірусів виявлено ряд білків з ферментативною активністю, зокрема РНК- залежна ДНК- полімераза, ДНК- залежна ДНК-полімераза, нуклеази, протеїнкінази і інші. Зворотна транскриптаза (РНК- залежна ДНК – полімераза) зв'язана з нуклеоїдом віріонів і представлена у онковірусів 10-20 молекулами на віріон. Молекулярна маса цього ферменту у віріонах типу С становить 70 кД, у В та D - 100-105кД. У онковірусів птахів ідентифіковано дві форми зворотної транскриптази : L (65кД) та В (105кД). У віріоні вони утворюють комплекс LB. Ліпіди знаходяться у складі ліпопротеїдної оболонки і мають клітинне походження. Вуглеводи входять до складу глікопротеїдів Глікопротеїди є типоспецифічними антигенами, беруть участь в реакції нейтралізації. Основним є глікопротеїд зовнішньої мембрани – SU.
Господар – хордові – примати, парнокопитні, непарнокопитні та хижаки.
Шляхи передачі в природі: трансмісивний шлях, парентеральний, вертикальний.
Переносники відсутні.
Географіче розповсюдження: по всьому світу.
Патогенез. У природних умовах патогенні для великої рогатої худоби. В експериментальних - захворюють телята, вівці. У природних умовах зараження чутливих тварин може відбуватись пренатально та частіше постнатально. Механізм пренатального зараження може являти собою передачу вірусного геному через гамети (генетична чи хромосомна трансляція ) або передачу сформованого вірусу (епігенетична екстрахромосомна трансляція). Постнатальна передача здійснюється через молоко чи під час прямого контакту. У останньому випадку вірус заноситься в організм з різноманітними секретами і екскретами або комахами чи контамінованими об'єктами (ін'єкційними голками, хірургічними інструментами тощо). Доведено, що вірус тривалий час персистує в організмі кліщів і може передаватись через них великій рогатій худобі.
Контактна передача — основний шлях розповсюдження лейкозу в природних умовах. Це потрібно мати на увазі тим, хто маніпулює в несприятливих по лейкозу стадах тварин.
Тропізм: найчастіше клітини імунної системи.
Патологічні зміни: знищують популяцію імунокомпетентних клітин, що призводить до загибелі особин від опортуністичних інфекцій. Порушення геному можуть викликати рак.
Вакцини для профілактики розробляються. Лікування ВІЛ – комбінована терапія з азидотимідином та інгібітором протеази ВІЛ. У природних умовах певне значення має колостральний імунітет, який триває кілька місяців і залежить від ряду обставин, зокрема від кількості молозивних імуноглобулінів.
У дорослих інфікованих тварин, попри наявність віруснейтралізуючих антитіл і інших факторів захисту, інфекційний процес не зупиняється. Штучна імунізація поки-що не набула широкого розповсюдження. Є повідомлення щодо позитивних результатів при використанні інактивованих препаратів. Двократне введення інактивованої 0,05%-ним АЕН, або 0,02% ДЕІ вакцини телятам обумовлено індукцію високих титрів віруснейтралізуючих антитіл та резистентність до експериментального зараження.