الأسماء في صفحات التنقل
Retrovirusse is enkelstring-RNS-virusse met ’n positiewe polariteit en ’n DNS-tussenstadium. By die infeksie van die gasheer vervaardig die virus DNS van sy RNS-genoom – die teenoorgestelde proses as normaalweg, waar DNS in RNS omgesit word (dus "retro", omgekeerd).
Hul naam word aangedui met "RT" agteraan gevoeg. ’n Voorbeeld is die MI-virus, wat ’n ssRNA-RT-virus is.
Un retrovirus és un virus que el seu genoma en fase extracel·lular és d'ARN. Utilitza una transcriptasa inversa per transcriure el seu genoma d'ARN a ADN i poder integrar-lo en el genoma de l'hoste. El virus en si mateix és solament un recipient per emmagatzemar el seu ARN. La transcripció inversa té lloc en el citosol de la cèl·lula. El genoma d'un retrovirus integrat en el genoma de l'hoste es coneix com a provirus.
El genoma d'un retrovirus conté com a mínim tres gens:
S'han identificat quatre retrovirus humans que ataquen les cèl·lules CD4 (HTLV 1 i 2, HIV 1 i 2). Una altra característica comuna a tots els retrovirus és el recobriment lipídic que envolta la càpsida. Això explica per què els retrovirus poden ser eliminats amb el simple gest de rentar-se les mans amb sabó.
Estudis d'aquests virus van ser els primers que van demostrar la síntesi d'ADN amb plantilles d'ARN, un mode fonamental de transmissió de material genètic que succeeix tant en eucariotes com en procariotes.
Aquests són virus amb embolcall que presenten un genoma de RNA monocatenari i es repliquen de manera inusual a través d'una forma intermèdia d'ADN bicatenari. Aquest procés es duu a terme mitjançant un enzim: la retrotranscriptasa o transcriptasa inversa, que dirigeix la síntesi d'ADN a través d'ARN i té una importància extraordinària en la manipulació genètica. Un cop s'ha passat d'ARN monocatenari a ADN s'insereix dins de l'ADN propi de la cèl·lula infectada on es comporta com un gen més.[1] Per tant, s'inclouen en el Grup VI de la Classificació de Baltimore.
Els retrovirus són responsables de moltes malalties, incloent alguns càncers i el SIDA (VIH).[2] Els científics poden modificar genèticament els retrovirus i utilitzar-los en teràpia gènica com a vectors.
La família inclou els següents gèneres:
Els leucovirus són un antic gènere de virus morfològicament similars a virus RNA, que produeixen neoplàsies i leucèmies en els animals, entre els quals hi ha el virus de la leucèmia aviària i el de la leucèmia murina. Actualment s'inclouen dins els retrovirus.[3]
El genoma del virus pren la forma d'un ARNm de polaritat positiva, inclosa la cap 5 'i la poly-A 3' dins del virió. Un cop dins de la cèl·lula de l'hoste, la cadena d'ARN se sotmet a la transcripció inversa en el citosol i és integrat en el genoma de l'hoste, moment en què l'ADN retroviral s'anomena provirus.
En el cas del VIH, el genoma consta de dues molècules d'ARN de cadena simple i polaritat positiva. Les molècules d'ARN estan físicament unides mitjançant ponts d'hidrogen en els seus extrems 5 ', el que fa que sigui difícil l'encapsidación de més de 2 molècules en un provirus.
L'organització genòmica és sempre la mateixa, 5'-GagPol-env-3 ', ia més depenent del tipus de retrovirus, hi ha gens accessoris que se solapen amb els gens principals.
Malgrat la immensa variabilitat entre els diferents tipus de retrovirus, podem dir que la partícula viral es compon de:
El cicle de replicació comprèn diverses etapes comunes a tots els retrovirus. En una fase inicial o primerenca, el virus s'uneix a receptors específics de la cèl·lula gràcies a la glicoproteïna de superfície. Les membranes vírica i cel·lular es fusionen i la càpside viral entri a la cèl·lula.
Els enzims víriques estan associades a l'ARN genòmic formant un complex nucleoproteic. La síntesi d'ADN víric, incloent-hi les LTR, es produeix al citoplasma a través de la RT. L'activitat Arnasa H de la RT degrada la cadena d'ARN i gairebé simultàniament empra la de l'ADNc com a motlle per sintetitzar una segona cadena d'ADN, convertint-lo en bicatenari. Aquest roman unit al complex nucleoproteic, amb el que passa al nucli cel·lular i, mitjançant la Integrasa (IN), l'ADN viral s'integra (provirus) en el genoma cel·lular, on pot romandre per un temps indefinit. Després ve la fase de transcripció dels gens, originant ARN que serveixen com a genoma del nou virió i ARN missatgers per a les poliproteïnes. Les proteïnes de gag i pol s'associen amb l'ARN viral formant un "core" intracel·lular, mentre que les proteïnes de env s'insereixen a la membrana plasmàtica de la cèl·lula. Després de l'acoblament, es produeix la sortida de la cèl·lula per gemmació, durant la qual el virus adquireix la doble capa lipídica de l'embolcall. Estudis recents suggereixen que la sortida de la partícula viral es produeix en llocs determinats de la cèl·lula. Sembla que Gag migraría cap a zones de la membrana riques en determinats lípids, són les anomenades basses o raft. Les proteïnes associades a aquests raft que s'incorporen a l'embolicada del virus tindrien també un paper important en la replicació viral. Finalment es produeix la maduració de les partícules gemadas mitjançant el processament de les poliproteïnes per la Proteasa (PR) viral que talla les poliproteïnes precursores. Els principals productes són les proteïnes de la matriu (p17), càpside (p24), nucleocàpside (p7) i la p6 (proteïna). La proteasa és només activa dins del virió. La maduració del VIH constitueix un mecanisme important per al disseny d'antiretrovirals.
Hi ha 4 retrovirus humans identificats: el virus de la immunodeficiència humana de tipus 1 (VIH-1), el de tipus 2 (HIV-2) i els virus limfotròpic de cèl·lules T humans de tipus I i II (HTLV-I i HTLV - II). Tots tenen com a diana els limfòcits T. Els virus de la immunodeficiència humana produeixen la lisi de les cèl·lules que infecten provocant una severa immunodepressió. Els virus HTLVI / II produeixen la immortalització dels limfòcits infectats, generant una replicació descontrolada d'aquests, i per tant una limfoproliferación.
La síndrome d'immunodeficiència humana o la SIDA, és l'expressió final de la infecció pel VIH. La infecció per aquest virus ocasiona la destrucció del sistema immunitari a més de manifestacions neurològiques i tumorals. Aquestes manifestacions clíniques es deu al tropisme tant macrofàgics com limfocitari del virus. Presenta una preferència per infectar limfòcits TCD4 +, en què la replicació és activa i molt agressiva, el que provoca com a característica de la infecció una profunda immunosupressió. La fisiopatologia de la SIDA és un procés complex, on hi ha implicats mecanismes patogènics tan diferents que alguns fins avui no han estat completament compresos. Els principals mecanismes de transmissió de la infecció per VIH són sexual, parenteral i vertical.
Igual que el VIH-1, el VIH-2 és un lentivirus. El seu genoma està compost per 2 cadenes simples d'ARN de polaritat positiva, i també conté l'enzim RT, que permet la integració del material genètic del virus, com a forma de provirus en el genoma de la cèl·lula que infecta, que és generalment els limfòcits T CD4 +. Comparteix amb el VIH-1 un 40-50% d'homologia genética11, el que fa necessari disposar de tècniques de biologia molecular específiques per a diagnosticar la infecció per VIH-2. La distribució geogràfica de l'HIV-2 es limita pràcticament al continent africà i sembla que la patologia produïda és molt més lleu i lenta que la causada pel VIH-112. El mecanisme de transmissió és igual que el VIH-1.
El virus limfotròpic de cèl·lules T humà és un retrovirus que pertany a la família Oncovirinae. L'HTLV-I va ser el primer retrovirus oncògens humà conocido13. Provoca una hemopatia maligna anomenada leucèmia / limfoma de cèl·lules T de l'adult (ATL) i desenvolupa també una mielopatia subaguda anomenada paraparèsia espàstica Tropical (TSP) o mielopatia associada a HTLV-I (HAM) 14,15. L'HTLV-II és un virus a la recerca de malaltia, no té una patologia clarament definida tot i que se l'ha associat amb diversos síndromes neurològics i mielopatia subagudes. El material genètic està format per 2 molècules d'ARN de cadena simple i polaritat positiva. L'HTLV-I té com a diana principal els limfòcits TCD4 + i el HTLV-II els limfòcits TCD8 +. L'HTLV una vegada que ha infectat la cèl·lula pot romandre silent integrat en forma de provirus o començar a replicar-se. Es creu que el principal mecanisme de transmissió de la infecció per HTLV és a partir de mitosi de les cèl·lules que infecta. Aquesta expansió clonal dóna lloc al que s'anomena càrrega proviral16. L'HTLV necessita el contacte cèl·lula-cèl·lula per produir la infecció. Els principals mecanismes de transmissió de la infecció per virus HTLV són per via sexual, via parenteral i via vertical.
Un retrovirus és un virus que el seu genoma en fase extracel·lular és d'ARN. Utilitza una transcriptasa inversa per transcriure el seu genoma d'ARN a ADN i poder integrar-lo en el genoma de l'hoste. El virus en si mateix és solament un recipient per emmagatzemar el seu ARN. La transcripció inversa té lloc en el citosol de la cèl·lula. El genoma d'un retrovirus integrat en el genoma de l'hoste es coneix com a provirus.
El genoma d'un retrovirus conté com a mínim tres gens:
gag, que codifica les proteïnes estructurals i del nucli del virus. pol, que codifica la transcriptasa inversa, la proteasa i la integrasa. env, que codifica les proteïnes que recobreixen el virus.S'han identificat quatre retrovirus humans que ataquen les cèl·lules CD4 (HTLV 1 i 2, HIV 1 i 2). Una altra característica comuna a tots els retrovirus és el recobriment lipídic que envolta la càpsida. Això explica per què els retrovirus poden ser eliminats amb el simple gest de rentar-se les mans amb sabó.
Estudis d'aquests virus van ser els primers que van demostrar la síntesi d'ADN amb plantilles d'ARN, un mode fonamental de transmissió de material genètic que succeeix tant en eucariotes com en procariotes.
Aquests són virus amb embolcall que presenten un genoma de RNA monocatenari i es repliquen de manera inusual a través d'una forma intermèdia d'ADN bicatenari. Aquest procés es duu a terme mitjançant un enzim: la retrotranscriptasa o transcriptasa inversa, que dirigeix la síntesi d'ADN a través d'ARN i té una importància extraordinària en la manipulació genètica. Un cop s'ha passat d'ARN monocatenari a ADN s'insereix dins de l'ADN propi de la cèl·lula infectada on es comporta com un gen més. Per tant, s'inclouen en el Grup VI de la Classificació de Baltimore.
Els retrovirus són responsables de moltes malalties, incloent alguns càncers i el SIDA (VIH). Els científics poden modificar genèticament els retrovirus i utilitzar-los en teràpia gènica com a vectors.
Retroviry (v užším smyslu, tj. čeleď Retroviridae) jsou obalené RNA viry s jednovláknovou nukleovou kyselinou, které jsou schopné pomocí reverzní transkriptázy přepsat svou genetickou informaci do DNA, kterou následně mohou včlenit (inzertovat) do genomu hostitelské buňky. Reverzní transkripce byla poprvé objevena právě u těchto virů, dnes je však známo více virových čeledí s touto schopností.
Retroviry jsou viry obratlovců, mají vysokou hostitelskou specifitu a u svých hostitelů vyvolávají celoživotní infekce. Podle druhu viru jsou původci nádorových onemocnění nebo napadají lymfatickou tkáň. Pro člověka je patogenní T-lymfotropní virus a virus HIV, který způsobuje syndrom získaného selhání imunity, AIDS.
V širším smyslu se jako retroviry označují veškeré viry s reverzní transkriptázou, tedy Baltimorovy skupiny VI a VII.
Baltimoreova klasifikace virů řadí retroviry (v užším smyslu, čeleď Retroviridae) do skupiny VI, mezi ssRNA viry s reverzní transkriptázou.
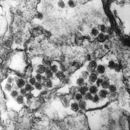
Viriony jsou obalené, v průměru kolem 100 nm velké, kulovitého tvaru. Kapsida uvnitř je pravděpodobně izohedrálně symetrická, z perikapsidálního obalu zralých virových částic vyčnívají na povrch glykoproteinové, asi 8 nm dlouhé peplomery, které tvoří jakési bodce pravidelně čnící z povrchu viru.
Virový genom je tvořen dvěma molekulami pozitivní jednovláknové RNA (+ssRNA), tvoří asi 2 % hmotnosti celé virové částice. Mimo této RNA se uvnitř virionu nachází také tRNA, která slouží jako primer k syntéze komplementární negativní DNA v infikované buňce, a reverzní transkriptáza a další enzymy.
Retroviry se replikují v cytoplasmě a dozrávají pučením z cytoplasmatické membrány napadené buňky. Převedením své genetické informace na dvouvláknovou DNA pomocí reverzní transkriptázy také získaly schopnost integrovat se do buněčného genomu jako tzv. provirus, neaktivní forma viru, která je však předávána všem potomkům nakažené buňky. V této formě je také dokonale chráněn před imunitním systémem hostitele – infekce retroviry je vždy doživotní, i když sama infekce nemusí nutně znamenat smrt nositele viru.
V současnosti (2018) je známo 11 rodů čeledi Retroviridae ve dvou podčeledích:[1]
Původní jeden rod Spumavirus byl rozdělen do 5 rodů, nazvaných podle hostitelů:
Do virů s reverzní transkriptázou patří (v závorce Baltimorova skupina):[1]
Retroviry (v užším smyslu, tj. čeleď Retroviridae) jsou obalené RNA viry s jednovláknovou nukleovou kyselinou, které jsou schopné pomocí reverzní transkriptázy přepsat svou genetickou informaci do DNA, kterou následně mohou včlenit (inzertovat) do genomu hostitelské buňky. Reverzní transkripce byla poprvé objevena právě u těchto virů, dnes je však známo více virových čeledí s touto schopností.
Retroviry jsou viry obratlovců, mají vysokou hostitelskou specifitu a u svých hostitelů vyvolávají celoživotní infekce. Podle druhu viru jsou původci nádorových onemocnění nebo napadají lymfatickou tkáň. Pro člověka je patogenní T-lymfotropní virus a virus HIV, který způsobuje syndrom získaného selhání imunity, AIDS.
V širším smyslu se jako retroviry označují veškeré viry s reverzní transkriptázou, tedy Baltimorovy skupiny VI a VII.
Retrovirus (retroviridae) er en gruppe af revers transkriberende RNA-virus. De bruger en revers transkriptase – et DNA polymerase-enzym – der i cellen oversætter retrovirusens enkeltstrengede RNA til dobbeltstrenget DNA. Dette DNA integreres i værtscellens eget DNA og danner så grundlaget for produktion af nye viruspartikler.
Der findes mange retrovira, der inficerer dyr, en del af disse er oncogene. Der findes kun fire humanpatogene retrovirus: HTLV-1, og -2, samt HIV-1 og -2. HIV-1(human immunodeficiency virus (HIV)) er den bedst kendte, da den er årsagen til AIDS.
Familien retroviridae har to underfamilier
Retrovirus (retroviridae) er en gruppe af revers transkriberende RNA-virus. De bruger en revers transkriptase – et DNA polymerase-enzym – der i cellen oversætter retrovirusens enkeltstrengede RNA til dobbeltstrenget DNA. Dette DNA integreres i værtscellens eget DNA og danner så grundlaget for produktion af nye viruspartikler.
Der findes mange retrovira, der inficerer dyr, en del af disse er oncogene. Der findes kun fire humanpatogene retrovirus: HTLV-1, og -2, samt HIV-1 og -2. HIV-1(human immunodeficiency virus (HIV)) er den bedst kendte, da den er årsagen til AIDS.
Retroviren (Retroviridae) sind eine große Familie behüllter Viren mit Einzel(+)-strängigem-RNA-Genom, deren Erbinformation (ss(+)-RNA) dementsprechend in Form von Ribonukleinsäure vorliegt. Anders als bei „normalen“ RNA-Viren aber muss die RNA von Retroviren zunächst einmal mittels reverser Transkription in ein DNA-Molekül umgeschrieben werden, bevor sie als solches in das Genom der Wirtszelle eingebaut und dort aktiv werden kann.
Retroviren können grob in einfache und komplexe Retroviren unterteilt werden. Des Weiteren unterscheidet man zwischen infektiösen exogenen Retroviren (teilweise mit XRV abgekürzt) und den endogenen Retroviren (ERV), die vertikal über die Keimbahn vererbt werden und auf diese Weise Bestandteil des Wirtszell-Genoms werden.
Retroviren infizieren vornehmlich tierische Zellen und sind bei Wirbeltieren allgegenwärtig: Sie infizieren Säugetiere, Vögel, Amphibien, Reptilien und Fische, sind dabei aber meist sehr wirtsspezifisch. Zu ihnen gehören die Erreger einiger weit verbreiteter Infektionskrankheiten, die sowohl bei Menschen als auch bei Tieren pandemisch bzw. epidemisch auftreten. Als Auslöser von Krankheiten beim Menschen sind unter anderem HIV und HTLV-1 bekannt.
Historisch wurden die Retroviren zunächst nach ihrem elektronenmikroskopischen Erscheinungsbild in Typ A-, B-, C- oder D-Retroviren eingeteilt. Später folgte eine Klassifikation, die auch biochemische Eigenschaften und den Zelltropismus berücksichtigte. Die Klassifikation unterschied Onkornaviren, Spumaviren und die Lentiviren.
Die aktuelle, zurzeit verbindliche Taxonomie durch das International Committee on Taxonomy of Viruses (ICTV) unterteilt die Retroviren vor allem aufgrund ihrer genetischen Verwandtschaftsverhältnisse wie folgt in zwei Unterfamilien und elf Gattungen:
Familie: Retroviren (Retroviridae)
Beim Menschen sind bisher fünf Retroviren bekannt: Humanes T-lymphotropes Virus 1 (HTLV-1) und Humanes T-lymphotropes Virus 2 (HTLV-2), beide Deltaretroviren; Humanes Immundefizienz-Virus-1 (HIV-1), Humanes Immundefizienz-Virus Typ II (HIV-2), beide Lentiviren; sowie Xenotropic murine leukemia virus-related virus (XMRV), ein Gammaretrovirus. Die menschlichen Retroviren sind denen anderer Primaten so eng verwandt, dass häufig beide Gruppen unter der Bezeichnung Primaten-Retroviren zusammengefasst werden. Es wird davon ausgegangen, dass die entsprechenden menschlichen Retroviren durch Übertragung von Affen-Retroviren auf den Menschen entstanden sind. Bei HTLV-1 und HTLV-2 hat diese Übertragung wohl schon vor Jahrtausenden stattgefunden, für HIV-1 und HIV-2 wahrscheinlich im 20. Jahrhundert. Die Foamyviren wurden in verschiedene Gattungen aufgespalten und bilden jetzt eine Unterfamilie der Retroviren (ICTV-Stand November 2018).
Die Unterteilung in einfache und komplexe Retroviren erfolgt nach der Genomorganisation bzw. der Translation von akzessorischen Proteinen. Dabei umfassen erstere Alpha-, Gamma-, Epsilon- und die meisten Betaretroviren, letztere die Deltaretroviren sowie Lenti- und Foamyviren.
Die Retroviridae gehören mit weiteren Virusfamilien (Belpaoviridae, Metaviridae und Pseudoviridae) zu den revers transkribierenden RNA-Viren der Virusordnung Ortervirales, die noch die Caulimoviridae als revers transkribierenden DNA-Viren gehören. Die Caulimoviridae werden manchmal mit den nicht näher verwandten, aber ebenfalls revers transkribierenden DNA-Viren der Hepadnaviridae als Pararetroviren (Baltimore-Gruppe 7) klassifiziert.
Seit dem 19. Jahrhundert waren bei Haustieren Krankheiten wie die Bovine Leukose oder Jaagsiekte bei Schafen bekannt, ihre Ursache blieb jedoch unklar. 1904 wurde für die erste durch Retroviren verursachte Krankheit, die Ansteckende Blutarmut der Einhufer, durch zwei französische Tierärzte, Vallée und Carré, gezeigt, dass sie sich mit einem Filtrat auf andere Pferde übertragen ließ. Onkogene (Tumoren auslösende) Retroviren wurden seit 1908 untersucht, als die dänischen Pathologen Vilhelm Ellermann und Oluf Bang zeigten, dass sich durch zellfreie Filtrate Hühnerleukämie auf andere Hühner übertragen ließ und die damit die erste ansteckende Krebserkrankung beschrieben.[3] Dieses Virus ist heute als aviäres Leukosevirus (ALV) bekannt, zunächst wurde die Leukose jedoch nicht als echte Leukämie angesehen und Leukämien wurden nicht als echte Tumoren betrachtet. Diese frühen Untersuchungen blieben innerhalb der Wissenschaftsgemeinde weitgehend unbeachtet, erst viel später konnte ihre Bedeutung im Zusammenhang mit Retroviren erkannt werden. Peyton Rous stellte 1911 fest, dass mit filtrierten Extrakten aus Hühnersarkomen gesunde Hühner infiziert werden konnten, die daraufhin ebenfalls Tumoren entwickelten. Das Virus wurde später nach ihm Rous-Sarkom-Virus (RSV) genannt und Rous erhielt 1966, 54 Jahre nach seiner Erstbeschreibung, den Nobelpreis. Erst 1961 wurde festgestellt, dass Rous-Sarkom-Viren Ribonukleinsäure (RNA) enthalten, sie wurden daher (bis 1974) als RNA-Tumorviren bezeichnet.
Die Entdeckung, dass Viren Tumoren auslösen können, bestätigte sich 1936 auch bei Säugetieren: John J. Bittner beschrieb das Maus-Mammatumorvirus (MMTV). 1951 wurde das Murine Leukämievirus (MLV) isoliert und erstmals die vertikale Übertragung von Eltern auf die Nachkommen beschrieben. 1964 wurde von Howard M. Temin die Provirus-Hypothese vorgeschlagen, da beobachtet wurde, dass Zellen, die durch RSV „transformiert“ wurden (Tumor-Eigenschaften erhielten), auch in Abwesenheit des Virus die transformierten Eigenschaften beibehielten. Aus diesem Grund postulierte Temin in Anlehnung an temperente Bakteriophagen, von denen man bereits wusste, dass sie ins Genom ihres Wirts integrieren können, dass die RNA-Tumorviren dies ebenfalls tun. Bereits 1960 war von André Lwoff vorgeschlagen worden, dass DNA-Tumorviren (Polyomaviren) in das Genom ihres Wirts integrieren können. 1968 wurde gezeigt, dass diese Annahme zutrifft. Dass auch RNA-Tumorviren über die Keimbahn vererbt werden können, wurde weiterhin als bizarr betrachtet.[4]
Endogene Retroviren wurden gegen Ende der 1960er Jahre entdeckt. Die Vermutung, dass ganze virale Genome durch ihre Wirte nach den Mendelschen Regeln weitervererbt werden, war eine völlig neue Vorstellung, und die Provirus-Hypothese von Temin wurde immer noch nicht allgemein akzeptiert, teilweise sogar für unmöglich gehalten. Die Reverse Transkriptase, durch die RNA in DNA umgeschrieben wird, wurde 1970 nachgewiesen, und die Familie der RNA-Tumorviren als Folge 1974 in Retroviren umbenannt. Temins Provirus-Hypothese erwies sich nach der Entdeckung der Reversen Transkriptase endgültig als zutreffend.
Zu Beginn der 1970er Jahre wurden auch die ersten viralen Proteine beschrieben und im Verlauf der darauffolgenden Jahre der Replikationszyklus der Retroviren nach und nach in groben Zügen aufgeklärt. 1978 wurden die LTR-Regionen (Long Terminal Repeats) im Genom der Retroviren entdeckt, zwei Jahre später wurde ein Jumping scheme für den komplexen Vorgang der reversen Transkription vorgeschlagen. Die Technik der DNA-Sequenzierung, die Anfang der 1980er Jahre aufkam, führte 1981 zur ersten Publikation der vollständigen genomischen Sequenz eines Retrovirus, des Moloney murine leukemia virus.
1980 erfolgte die Erstbeschreibung des humanen T-Zell-Leukämie-Virus Typ 1 (HTLV-1), des ersten Retrovirus, das den Menschen infiziert, nachdem viele Jahre lang erfolglos in allen möglichen Tumorgeweben des Menschen nach Retroviren gesucht wurde. Kurze Zeit darauf entdeckten Luc Montagnier und Françoise Barré-Sinoussi (Nobelpreis für Medizin 2008) das HIV-1, HIV-2 folgte 1986. Spätestens seitdem 1988 klar wurde, dass HIV die Ursache der Immunschwächeerkrankung AIDS ist, entwickelte sich die Retrovirologie von einer eher exotischen Grundlagenforschung zu dem am intensivsten beforschten Gebiet in der Virologie mit großer Bedeutung für die Gesundheitswissenschaften.
Infektiöse Retrovirus-Partikel haben einen Durchmesser von etwa 100 nm. Sie besitzen ein Kapsid, das von einer Virushülle umgeben ist, die aus der Zytoplasmamembran der Wirtszelle abgeschnürt wurde und mit viralen Glykoproteinen durchsetzt ist, sowie einen „Kern“ innerhalb des Kapsids aus weiteren Proteinen und einem Ribonukleinsäure-Komplex.
Das einzelsträngige RNA-Genom der Retroviren ist linear und 7–12 Kilobasenpaare (kb) groß. Retroviren sind die einzigen RNA-Viren, die diploid angelegt sind, d. h. jedes Retrovirus hat eine zusätzliche Kopie seines Genoms. Sie werden von den wirtseigenen Transkriptions-Enzymen übersetzt und synthetisiert und benötigen eine spezifische zelluläre tRNA. Das provirale Genom eines einfachen Retrovirus enthält in der Regel drei Gene und zwei Long Terminal Repeats (LTRs), die sich am Anfang und am Ende befinden und Informationen zur Steuerung der Expression der viralen Gene enthalten. Bei den drei Genen handelt es sich um gag (Gruppenspezifisches Antigen), pol (Polymerase) und env (envelope). gag codiert die Matrix-, Kapsid- und Nukleokapsidproteine. pol codiert die viralen Enzyme Protease, Reverse Transkriptase (mit RNase H) und Integrase. Bei den Beta- und Deltaretroviren hat die Protease ein eigenes Leseraster (pro) und bei den Alpharetroviren befindet sich die Information für die Protease im gag-Gen. env codiert die Proteine der Hülle. An regulatorischen Sequenzen gibt es im 5'-Bereich eine mit ψ (psi) bezeichnete Sequenz, die ein Signal für das Verpacken der RNA in die Viruspartikel ist, eine Primerbindungsstelle (PBS), an die sich die jeweilige tRNA anlagern kann und einen Promotor. Im 3'-Bereich finden sich ein oder mehr Polypurintrakte, die bei der reversen Transkription essentiell sind.
Komplexe Retroviren, wie z. B. das zu den Lentiviren gehörende HI-Virus, das zu den Deltaretroviren gehörende HTLV oder die Foamyviren enthalten noch weitere, regulatorische Gene, die als akzessorische Gene bezeichnet werden. Bei HIV-1 sind dies tat, rev, vif, nef, vpu und vpr, bei HTLV-1 rex, rof, tax, tof und bei den Foamyviren tas und bet.
Der Lebenszyklus eines Retrovirus besteht aus mehreren Schritten: Infektion der Zelle, Reverse Transkription, Überwindung der Kernhülle, Integration ins Wirtsgenom, Expression der viralen Proteine und des RNA-Genoms und die Bildung neuer Viruspartikel.
Nachdem das Glykoprotein der Virushülle spezifisch an seine(n) zellulären Rezeptor(en) gebunden hat, verschmilzt die virale Membran mit der Membran der Zelle und entlässt das Kapsid in das Innere der Zelle, das Cytoplasma. Was mit dem Kapsid im Cytoplasma geschieht, ist noch nicht in allen Einzelheiten geklärt, vermutlich zerfällt es in seine einzelnen Bausteine und gibt die beiden ssRNA-Stränge und die im Inneren enthaltenen Proteine, wie zum Beispiel die Reverse Transkriptase ins Cytoplasma der Wirtszelle frei.
Hauptartikel: Reverse Transkriptase
Retroviren sind die einzigen einsträngig-plusstrangorientierten RNA-Viren, bei denen das Genom nicht sofort als Matrize (mRNA) bei der Infektion benutzt werden kann: Wenn das Virus diese RNA in die zu befallende Zelle eingebracht hat, muss die RNA in doppelsträngige DNA umgeschrieben werden. Dieser Vorgang wird reverse Transkription genannt. Dazu bringt das Virus die Reverse Transkriptase in seinen Viruspartikeln mit. Diese schreibt die einzelsträngige RNA des Virus zuerst in einzelsträngige DNA und anschließend in doppelsträngige DNA um. Bei der reversen Transkription werden die beiden LTR-Sequenzen generiert, die für den weiteren Ablauf der Infektion essenziell sind.
Normalerweise verläuft die Transkription an der DNA als Matrize, wobei ein komplementärer RNA-Strang synthetisiert wird; eine Ausnahme stellen die Retroviren und die Retroelemente (auch Klasse-I-Transposons genannt) dar. Da der Prozess durch die fehlende Korrekturlese-Fähigkeit der Reversen Transkriptase relativ ungenau ist, erfolgen häufige Mutationen des Virus. Diese ermöglichen eine schnelle Anpassung des Virus an antivirale Medikamente und damit eine Ausbildung von Resistenzen.
Einige Gattungen der Retroviren, beispielsweise Gammaretroviren, können die Kernhülle nicht aktiv überwinden. Sie infizieren daher nur Zellen, die sich in Teilung befinden und nutzen den Moment der Zellteilung zur Integration, wenn das Genom nicht durch die Kernhülle geschützt ist. Andere Gattungen wie beispielsweise die Alpharetroviren und vor allem die Lentiviren können die Kernhülle aktiv überwinden und damit auch ruhende Zellen infizieren. Der Eintritt in den Zellkern erfolgt nach der Bildung des Präintegrationskomplexes (PIC) im Cytoplasma. Da die Kernporen kleiner sind als der PIC, der etwa die Größe eines Ribosoms hat, muss es sich um einen aktiven Transportprozess handeln. An diesem noch nicht vollständig verstandenen Prozess sind sowohl zelluläre als auch virale Proteine beteiligt. Ein weithin akzeptiertes Modell beschreibt den Eintritt in den Zellkern durch die Kernporen mit Hilfe von karyophilen Signalen der im PIC enthaltenen Proteine.[5]
Hauptartikel: Integrase
Die Integration des viralen Genoms in das Wirtsgenom ist ein essentieller Schritt im Replikationszyklus des Virus. Er wird katalysiert durch ein Enzym namens Integrase, das in allen Retroviren und Retrotransposons vorkommt. Die Integrase bindet an die virale und die Wirts-DNA und bildet mit diesen einen Komplex, der als Präintegrationskomplex (PIC, von engl. preintegration complex) bezeichnet wird. Theoretisch kann die Integration an jedem Ort im Wirtsgenom erfolgen, je nach Art des Retrovirus zeigen sich jedoch bestimmte bevorzugte chromosomale Bereiche. Was genau den Ort der Integration beeinflusst, ist noch nicht völlig geklärt, sicher ist jedoch, dass die Aminosäuresequenz der Integrase eine Rolle spielt, wodurch sich spezifische Interaktionsmöglichkeiten zwischen Integrase und Faktoren im Chromatin ergeben. In der Abbildung rechts ist die Abfolge der DNA-Strangbrüche und anschließender Neuzusammensetzung bei der Integration des viralen Genoms dargestellt. In grau sind die Integrase-Monomere, in rot die virale DNA und in schwarz die Wirts-DNA abgebildet, die roten Punkte schließlich die 5'-Enden der viralen DNA:
Nach der Integration werden zelluläre Transkriptionsfaktoren und die RNA-Polymerase II der Zelle rekrutiert, um die Provirus-DNA zu transkribieren. Die dafür notwendigen Promotor- und Enhancer-Strukturen liegen in der LTR des Provirus. Bei komplexen Retroviren wirken manche viralen Proteine (z. B. tat) zusätzlich als Transkriptionsverstärker. Es entstehen verschiedene mRNAs durch alternatives Spleißen.
Die entstehenden mRNAs werden ins Cytoplasma transportiert. Dort werden die verschiedenen viralen Proteine translatiert. Die Synthese der Env-Proteine erfolgt an der Membran des Endoplasmatischen Retikulums, so dass die Env-Proteine direkt in der Zellmembran verankert werden, wo sie sich zu Trimeren zusammenlagern. Alle anderen Virusproteine werden an freien Ribosomen synthetisiert. Die Gag- und Gag/Pol-Vorläuferproteine werden aminoterminal myristyliert und lagern sich an der Zellmembran an. An der Zellmembran findet dann die Partikelbildung statt: Gag- und Gag/Pol-Vorläuferproteine akkumulieren sich und interagieren außer miteinander auch mit Glykoproteinen der Zelle. Nur die ungespleißten mRNAs, von denen Gag und Pol translatiert wurden, besitzen das Verpackungssignal Psi und die Leader-Sequenz. Mit Hilfe des Psi-Signals binden die mRNAs an die Zinkfingermotive der Nucleokapsidproteine- so wird sichergestellt, dass nur ungespleißte und damit vollständige Genome in das Viruspartikel verpackt werden. Bei Kontakt mit den mRNAs stülpt sich die Membran an der Zelloberfläche ein und schnürt ein unreifes Viruspartikel ab. Erst innerhalb dieses Partikels beginnt die virale Protease, sich zu Dimeren zusammenzulagern, sich in einem autokatalytischen Schnitt aus den Vorläuferproteinen herauszuschneiden und dann die Gag- und Gag/Pol-Vorläuferproteine in die einzelnen Komponenten (Matrix-, Kapsid-, Nukleokapsid, Reverse Transkriptase und Integrase) zu spalten. Innerhalb des Partikels lagern sich die Kapsidproteine zu einem konischen Kapsid zusammen. Erst am Ende dieses Reifungsprozesses ist der Partikel infektiös.
Hauptartikel: Retroelement, Retrotransposon, Endogenes Retrovirus
Der genaue Entstehungszeitpunkt der ersten retrovirusartigen Partikel ist unklar, die ältesten Sequenzen legen ein Alter von mindestens 250 Millionen Jahren nahe, vermutlich sind sie noch wesentlich älter.[6] Retroviren haben sich wahrscheinlich aus Retrotransposons entwickelt. Das bedeutet, dass sie eine infektiöse, weiterentwickelte Form der Retroelemente darstellen. Demnach wäre die reverse Transkription einer der ältesten Mechanismen in der Retrovirusentwicklung, der möglicherweise bereits in der RNA-Welt entstanden ist. Große Ähnlichkeit besteht jedenfalls zwischen Retroviren und den Transposons aus verschiedenen Lebewesen, beispielsweise den Ty-Elementen der Bäckerhefe und den Copia- und Ulysses-Elementen aus Drosophila melanogaster. Diese Retrotransposons codieren eine Reverse Transkriptase und haben eine ähnliche Struktur wie die Retroviren.
Hinweise, dass die Organisation der DNA bei Bakterien fundamental von der bei Archaeen und komplexen Zellen (Eukaryoten) abweicht, geben Anlass zu der These, dass deren zelluläre Urvorfahren (LUGA oder LUCA) noch der RNA-Welt angehört haben. Die Speicherung der Erbinformation in der DNA wird dann als eine Fähigkeit angesehen, die zunächst von Retroviren ‚erfunden‘ wurde und die dann zelluläre Organismen mehrmals durch Übertragung von solchen erworben haben. Daraus resultieren die Bakterien einerseits, die Archaeen und Eukaryoten andererseits. (Der grundsätzliche Aufbau der Ribosomen als Eiweißfabriken und der Genetische Code stimmen dagegen bei allen zellulären Organismen so gut überein, dass bereits der LUCA darüber verfügt haben muss.)[7]
Die Integration in das Genom ihres Wirts ist eines der ungewöhnlichsten und bemerkenswertesten Merkmale von Retroviren. Die Vielzahl an ähnlichen Sequenzen in Wirbeltieren und Retroviren zeigt, dass Retroviren in der Vergangenheit schon sehr oft auch die Zellen der Keimbahn ihrer Wirte infiziert haben. Derart an die Nachkommen vererbte Retroviren werden als endogene Retroviren (ERV) bezeichnet, um sie von den horizontal weitergegebenen, exogenen Retroviren zu unterscheiden. Durch die zunehmende Zahl sequenzierter Organismen wurden auch immer mehr endogene Retroviren entdeckt. Die Menge retroviraler DNA schwankt bei Wirbeltieren zwischen 5 und 10 %, das menschliche Genom besteht zu etwa 8 % aus retroviralen Sequenzen. Diese Daten geben einen Einblick in die lange Wirts-Virus-Coevolution.[8] Mit den Endogenen Retroviren wurden aus den infektiösen Viren, die aus Transposons entstanden, wieder Teile der Genome.
Bisher wurden im menschlichen Genom 31 verschiedene ERV-„Familien“ beschrieben, die wahrscheinlich auf 31 verschiedene Fälle von Keimbahninfektionen durch Retroviren zurückgehen (im Englischen als genome invasion event (Genominvasionsereignis) bezeichnet). Diesem Ausgangsereignis folgte eine Erhöhung der ERV-Kopienzahl, entweder durch Reinfektion der Keimbahnzellen oder durch Retrotransposition innerhalb der Zelle. Im Lauf der Generationen nimmt die Aktivität der ERVs immer weiter ab, da sich Mutationen ansammeln und ganze Abschnitte der ERVs verloren gehen können, bis schließlich die Aktivität der Viren ganz aufhört. Das Alter der einzelnen ERV-Familien oder ERV-Linien kann anhand der Größe und Form der Phylogenetischen Stammbäume der Linien abgeschätzt werden. Die meisten der humanen ERV-Linien (HERV) entstanden demnach vor der evolutionären Entwicklung von Altwelt- und Neuweltaffen vor etwa 25 bis 30 Millionen Jahren.
Von Retroviren werden sehr viele verschiedene Lebewesen infiziert. Die betroffenen Spezies reichen von Muscheln bis zum Menschen, wobei die meisten unter den Wirbeltieren zu finden sind. Retroviren verursachen in ihren Wirten eine große Zahl verschiedenartiger Krankheiten, darunter Tumoren (Lymphome, Sarkome), Neurologische Erkrankungen und Immunschwächen. Einige dieser Erkrankungen verursachen große Schäden in der Landwirtschaft, weil Nutztiere betroffen sind, oder sind die Ursache für menschliche Pandemien (AIDS). Andere Infektionen bleiben symptomlos, weshalb diese Retroviren als apathogen angesehen werden.
Manche durch Retroviren induzierte Krankheiten von Nagetieren stellen Modellsysteme dar, anhand derer die Infektionsmechanismen der Retroviren sowie die Entstehung der durch manche Retroviren verursachten Tumoren im Detail untersucht werden können. Die moderne Tumorbiologie fußt teilweise auf Daten, die aufgrund dieser Modelle erzeugt werden konnten.
Sehr viele heute bekannte Onkogene wurden zunächst bei Experimenten mit tierischen Retroviren, die diese Onkogene in ihr Genom aufgenommen hatten, entdeckt. Beispiele sind:[9][10][11]
Retroviren (Retroviridae) sind eine große Familie behüllter Viren mit Einzel(+)-strängigem-RNA-Genom, deren Erbinformation (ss(+)-RNA) dementsprechend in Form von Ribonukleinsäure vorliegt. Anders als bei „normalen“ RNA-Viren aber muss die RNA von Retroviren zunächst einmal mittels reverser Transkription in ein DNA-Molekül umgeschrieben werden, bevor sie als solches in das Genom der Wirtszelle eingebaut und dort aktiv werden kann.
Retroviren können grob in einfache und komplexe Retroviren unterteilt werden. Des Weiteren unterscheidet man zwischen infektiösen exogenen Retroviren (teilweise mit XRV abgekürzt) und den endogenen Retroviren (ERV), die vertikal über die Keimbahn vererbt werden und auf diese Weise Bestandteil des Wirtszell-Genoms werden.
Retroviren infizieren vornehmlich tierische Zellen und sind bei Wirbeltieren allgegenwärtig: Sie infizieren Säugetiere, Vögel, Amphibien, Reptilien und Fische, sind dabei aber meist sehr wirtsspezifisch. Zu ihnen gehören die Erreger einiger weit verbreiteter Infektionskrankheiten, die sowohl bei Menschen als auch bei Tieren pandemisch bzw. epidemisch auftreten. Als Auslöser von Krankheiten beim Menschen sind unter anderem HIV und HTLV-1 bekannt.
Un retrovirus es un virus que son genòma en fasa extracellular es d'ARN. Utiliza una transcriptasa invèrsa per transcriure son genòma de l'ARN a l'ADN e poder l'integrar en lo genòma del òste. La transcripcion inversa se debana en lo citosòl de la cellula infectada. Lo genòma d'un retrovirus integrat dins lo genòma del òste s'apela un provirus. Un exemple de retrovirus es lo virus dau SIDA o virus de l'immunodeficiéncia umana.
Retrovirus je pripadnik virusne porodice Retroviridae – virusa koji se umnožavaju u ćeliji domaćina putem procesa reverzne transkripcije. To je jednolančani pozitivno smisleni RNK virus sa DNK međustepenom, a ćelije domaćina napada kao parazit. Unutar ćelije domaćina u citoplazmi, virus koristi svoj enzim reverznu transkriptazu za proizvodnju DNK iz RNK genoma, suprotno od uobičajenog obrasca, tj. retro (=unazad). Ova nova DNK je tada uključena u genom ćelije domaćina pomoću enzima integraze, koja usmjerava retrovirusnu DNK zvanu provirus. Ćelije domaćina zatim prihvataju virusnu DNK kao dio vlastitog genoma, prevodi i prepisuje virusne gene zajedno sa vlastitim genima, uključujući i proizvodnju proteina potrebnih za sklapanje novih kopija virusa. Zato je teško otkriti virus prilikom zaraze domaćina. U tom trenutku, infekcija će istrajati na neodređeno vrijeme.
Najpoznatiji virus iz ove grupe je HIV (Human Immunodeficiency Virus - ljudski virus imunodeficijencije). Svi retrovirusi koriste reverznu transkriptazu za sintezu DNK iz RNK. Inhibicijom reverzne transkriptaze se zaustavlja razmnožavanje retrovirusa, pa su razvijeni odgovarajući lijekovi (inhibitori reverzne tranksriptaze).
Kod većine virusa, DNK se transkribirana u RNK, koja se zatim prevodi na jezik aminokiselina, tj. proteina. Međutim, retrovirusi djeluju drugačije - njihova RNK je obrnuto transkribovana u DNK, koja je ukjlučena u genom ćelije domaćina (kada postaje provirus), a zatim prolazi kroz uobičajene transkripcijske i translacijske procese ekspresije virusnih gena. Dakle, informacija retrovirusnih gena se koristi za stvaranje odgovarajućeg proteina u slijedu: RNK → DNK → RNK → polipeptid. To proširuje osnovne procese koje je prepoznao Francis Crick, (jedan gen-jedan peptid), u kojima je formuliran niz: DNK → RNK → peptid, (proteini su izrađeni od jednog ili više polipeptidnih lanaca npr., hemoglobin sadrži četiri lanca peptida).
Retrovirusi su dokazani kao vrijedni istraživački alati u molekularnoj biologiji, a uspješno se koriste u sistemima prenosa gena.[1]
Virioni retrovirusa se sastoje od obavijenih čestica promjera oko 100 nm. Oni također sadrže dvije identične jednolančane molekule RNK dužine 7-10 kilobaza. Iako virioni različitih retrovirusa nemaju istu morfologiju ili biologijawbiologiju, svi sastavni dijelovi viriona su vrlo slični.[2]
Glavni dijelovi viriona su:
Virusni omotač, koji se sastoji od lipida (koji se dobija od plazmi membrane domaćina tokom procesa pupoljenja), kao i glikoproteina kodiranog sa env genima. Omotač ima tri različite uloge: zaštita od ekstracelularnog okruženja preko lipidnog dvosloja, omogućavajući retrovirusu ulazak / izlazak u/iz ćelije kroz endosomske membrane čovjeka i sposobnost da izravno uđe u ćelije, stapanjem sa svojim membranama.
Retroviridae is a faimily o enveloped viruses that replicate in a host cell throu the process o reverse transcription.
Retrovirus je pripadnik virusne porodice Retroviridae – virusa koji se umnožavaju u ćeliji domaćina putem procesa reverzne transkripcije. To je jednolančani pozitivno smisleni RNK virus sa DNK međustepenom, a ćelije domaćina napada kao parazit. Unutar ćelije domaćina u citoplazmi, virus koristi svoj enzim reverznu transkriptazu za proizvodnju DNK iz RNK genoma, suprotno od uobičajenog obrasca, tj. retro (=unazad). Ova nova DNK je tada uključena u genom ćelije domaćina pomoću enzima integraze, koja usmjerava retrovirusnu DNK zvanu provirus. Ćelije domaćina zatim prihvataju virusnu DNK kao dio vlastitog genoma, prevodi i prepisuje virusne gene zajedno sa vlastitim genima, uključujući i proizvodnju proteina potrebnih za sklapanje novih kopija virusa. Zato je teško otkriti virus prilikom zaraze domaćina. U tom trenutku, infekcija će istrajati na neodređeno vrijeme.
Najpoznatiji virus iz ove grupe je HIV (Human Immunodeficiency Virus - ljudski virus imunodeficijencije). Svi retrovirusi koriste reverznu transkriptazu za sintezu DNK iz RNK. Inhibicijom reverzne transkriptaze se zaustavlja razmnožavanje retrovirusa, pa su razvijeni odgovarajući lijekovi (inhibitori reverzne tranksriptaze).
Kod većine virusa, DNK se transkribirana u RNK, koja se zatim prevodi na jezik aminokiselina, tj. proteina. Međutim, retrovirusi djeluju drugačije - njihova RNK je obrnuto transkribovana u DNK, koja je ukjlučena u genom ćelije domaćina (kada postaje provirus), a zatim prolazi kroz uobičajene transkripcijske i translacijske procese ekspresije virusnih gena. Dakle, informacija retrovirusnih gena se koristi za stvaranje odgovarajućeg proteina u slijedu: RNK → DNK → RNK → polipeptid. To proširuje osnovne procese koje je prepoznao Francis Crick, (jedan gen-jedan peptid), u kojima je formuliran niz: DNK → RNK → peptid, (proteini su izrađeni od jednog ili više polipeptidnih lanaca npr., hemoglobin sadrži četiri lanca peptida).
Retrovirusi su dokazani kao vrijedni istraživački alati u molekularnoj biologiji, a uspješno se koriste u sistemima prenosa gena.
Retroviridae is a faimily o enveloped viruses that replicate in a host cell throu the process o reverse transcription.
रेट्रोवायरस (Retrovirus) आरएनए वायरस का एक कुल है जिसकी सदस्य जातियाँ अन्य जीवों की कोशिकाओं में घुसकर उनके डीएनए में अपने अनुवांशिक जीनोम का आरएनए की प्रति डाल देती हैं। इस से वह कोशिका अपनी ही सामग्री से रेट्रोवायरस की अन्य प्रतियाँ बनाने पर विवश हो जाती है और उस रेट्रोवायरस का प्रजनन-केन्द्र बनकर उसकी संख्या बढ़ाने लगती है। मानवों में एड्स फैलाने वाली एचआइवी वायरस जातियाँ इसी रेट्रोवायरस कुल की सदस्य हैं।[1][2][3]
रेट्रोवायरस (Retrovirus) आरएनए वायरस का एक कुल है जिसकी सदस्य जातियाँ अन्य जीवों की कोशिकाओं में घुसकर उनके डीएनए में अपने अनुवांशिक जीनोम का आरएनए की प्रति डाल देती हैं। इस से वह कोशिका अपनी ही सामग्री से रेट्रोवायरस की अन्य प्रतियाँ बनाने पर विवश हो जाती है और उस रेट्रोवायरस का प्रजनन-केन्द्र बनकर उसकी संख्या बढ़ाने लगती है। मानवों में एड्स फैलाने वाली एचआइवी वायरस जातियाँ इसी रेट्रोवायरस कुल की सदस्य हैं।
A retrovirus is a type of virus that inserts a DNA copy of its RNA genome into the DNA of a host cell that it invades, thus changing the genome of that cell.[2] After invading a host cell's cytoplasm, the virus uses its own reverse transcriptase enzyme to produce DNA from its RNA genome, the reverse of the usual pattern, thus retro (backwards). The new DNA is then incorporated into the host cell genome by an integrase enzyme, at which point the retroviral DNA is referred to as a provirus. The host cell then treats the viral DNA as part of its own genome, transcribing and translating the viral genes along with the cell's own genes, producing the proteins required to assemble new copies of the virus. Many retroviruses cause serious diseases in humans, other mammals, and birds.[3]
Retroviruses have many subfamilies in three basic groups.
The specialized DNA-inflitration enzymes in retroviruses make them valuable research tools in molecular biology, and they have been used successfully in gene delivery systems.[6]
Evidence from endogenous retroviruses (inherited provirus DNA in animal genomes) suggests that retroviruses have been infecting vertebrates for at least 450 million years.[7]
Virions, viruses in the form of independent particles of retroviruses, consist of enveloped particles about 100 nm in diameter. The outer lipid envelope consists of glycoprotein.[8] The virions also contain two identical single-stranded RNA molecules 7–10 kilobases in length. The two molecules are present as a dimer, formed by base pairing between complementary sequences. Interaction sites between the two RNA molecules have been identified as a "kissing stem-loop".[3] Although virions of different retroviruses do not have the same morphology or biology, all the virion components are very similar.[9]
The main virion components are:

The retroviral genome is packaged as viral particles. These viral particles are dimers of single-stranded, positive-sense, linear RNA molecules.[10]
Retroviruses (and orterviruses in general) follow a layout of 5'–gag–pro–pol–env–3' in the RNA genome. gag and pol encode polyproteins, each managing the capsid and replication. The pol region encodes enzymes necessary for viral replication, such as reverse transcriptase, protease and integrase.[19] Depending on the virus, the genes may overlap or fuse into larger polyprotein chains. Some viruses contain additional genes. The lentivirus genus, the spumavirus genus, the HTLV / bovine leukemia virus (BLV) genus, and a newly introduced fish virus genus are retroviruses classified as complex. These viruses have genes called accessory genes, in addition to gag, pro, pol and env genes. Accessory genes are located between pol and env, downstream from the env, including the U3 region of LTR, or in the env and overlapping portions. While accessory genes have auxiliary roles, they also coordinate and regulate viral gene expression. In addition, some retroviruses may carry genes called oncogenes or onc genes from another class. Retroviruses with these genes (also called transforming viruses) are known for their ability to quickly cause tumors in animals and transform cells in culture into an oncogenic state.[20]
The polyproteins are cleaved into smaller proteins each with their own function. The nucleotides encoding them are known as subgenes.[18]

When retroviruses have integrated their own genome into the germ line, their genome is passed on to a following generation. These endogenous retroviruses (ERVs), contrasted with exogenous ones, now make up 5–8% of the human genome.[21] Most insertions have no known function and are often referred to as "junk DNA". However, many endogenous retroviruses play important roles in host biology, such as control of gene transcription, cell fusion during placental development in the course of the germination of an embryo, and resistance to exogenous retroviral infection. Endogenous retroviruses have also received special attention in the research of immunology-related pathologies, such as autoimmune diseases like multiple sclerosis, although endogenous retroviruses have not yet been proven to play any causal role in this class of disease.[22]
While transcription was classically thought to occur only from DNA to RNA, reverse transcriptase transcribes RNA into DNA. The term "retro" in retrovirus refers to this reversal (making DNA from RNA) of the usual direction of transcription. It still obeys the central dogma of molecular biology, which states that information can be transferred from nucleic acid to nucleic acid but cannot be transferred back from protein to either protein or nucleic acid. Reverse transcriptase activity outside of retroviruses has been found in almost all eukaryotes, enabling the generation and insertion of new copies of retrotransposons into the host genome. These inserts are transcribed by enzymes of the host into new RNA molecules that enter the cytosol. Next, some of these RNA molecules are translated into viral proteins. The proteins encoded by the gag and pol genes are translated from genome-length mRNAs into Gag and Gag–Pol polyproteins. In example, for the gag gene; it is translated into molecules of the capsid protein, and for the pol gene; it is translated into molecules of reverse transcriptase. Retroviruses need a lot more of the Gag proteins than the Pol proteins and have developed advanced systems to synthesize the required amount of each. As an example, after Gag synthesis nearly 95 percent of the ribosomes terminate translation, while other ribosomes continue translation to synthesize Gag–Pol. In the rough endoplasmic reticulum glycosylation begins and the env gene is translated from spliced mRNAs in the rough endoplasmic reticulum, into molecules of the envelope protein. When the envelope protein molecules are carried to the Golgi complex, they are divided into surface glycoprotein and transmembrane glycoprotein by a host protease. These two glycoprotein products stay in close affiliation, and they are transported to the plasma membrane after further glycosylation.[3]
It is important to note that a retrovirus must "bring" its own reverse transcriptase in its capsid, otherwise it is unable to utilize the enzymes of the infected cell to carry out the task, due to the unusual nature of producing DNA from RNA.
Industrial drugs that are designed as protease and reverse-transcriptase inhibitors are made such that they target specific sites and sequences within their respective enzymes. However these drugs can quickly become ineffective due to the fact that the gene sequences that code for the protease and the reverse transcriptase quickly mutate. These changes in bases cause specific codons and sites with the enzymes to change and thereby avoid drug targeting by losing the sites that the drug actually targets.
Because reverse transcription lacks the usual proofreading of DNA replication, a retrovirus mutates very often. This enables the virus to grow resistant to antiviral pharmaceuticals quickly, and impedes the development of effective vaccines and inhibitors for the retrovirus.[23]
One difficulty faced with some retroviruses, such as the Moloney retrovirus, involves the requirement for cells to be actively dividing for transduction. As a result, cells such as neurons are very resistant to infection and transduction by retroviruses. This gives rise to a concern that insertional mutagenesis due to integration into the host genome might lead to cancer or leukemia. This is unlike Lentivirus, a genus of Retroviridae, which are able to integrate their RNA into the genome of non-dividing host cells.
Two RNA genomes are packaged into each retrovirus particle, but, after an infection, each virus generates only one provirus.[24] After infection, reverse transcription occurs and this process is accompanied by recombination. Recombination involves template strand switching between the two genome copies (copy choice recombination) during reverse transcription. From 5 to 14 recombination events per genome occur at each replication cycle.[25] Genetic recombination appears to be necessary for maintaining genome integrity and as a repair mechanism for salvaging damaged genomes.[24]
The DNA formed after reverse transcription (the provirus) is longer than the RNA genome because each of the terminals have the U3 - R - U5 sequences called long terminal repeat (LTR). Thus, 5' terminal has the extra U3 sequence, while the other terminal has the U5 sequence.[15] LTRs are able to send signals for vital tasks to be carried out such as initiation of RNA production or management of the rate of transcription. This way, LTRs can control replication, hence, the entire progress of the viral cycle.[27] Although located in the nucleus, the non-integrated retroviral cDNA is a very weak substrate for transcription. For this reason, an integrated provirus is a necessary for permanent and an effective expression of retroviral genes.[10]
This DNA can be incorporated into host genome as a provirus that can be passed on to progeny cells. The retrovirus DNA is inserted at random into the host genome. Because of this, it can be inserted into oncogenes. In this way some retroviruses can convert normal cells into cancer cells. Some provirus remains latent in the cell for a long period of time before it is activated by the change in cell environment.
Studies of retroviruses led to the first demonstrated synthesis of DNA from RNA templates, a fundamental mode for transferring genetic material that occurs in both eukaryotes and prokaryotes. It has been speculated that the RNA to DNA transcription processes used by retroviruses may have first caused DNA to be used as genetic material. In this model, the RNA world hypothesis, cellular organisms adopted the more chemically stable DNA when retroviruses evolved to create DNA from the RNA templates.
An estimate of the date of evolution of the foamy-like endogenous retroviruses placed the time of the most recent common ancestor at> 450 million years ago.[28]
Gammaretroviral and lentiviral vectors for gene therapy have been developed that mediate stable genetic modification of treated cells by chromosomal integration of the transferred vector genomes. This technology is of use, not only for research purposes, but also for clinical gene therapy aiming at the long-term correction of genetic defects, e.g., in stem and progenitor cells. Retroviral vector particles with tropism for various target cells have been designed. Gammaretroviral and lentiviral vectors have so far been used in more than 300 clinical trials, addressing treatment options for various diseases.[6][29] Retroviral mutations can be developed to make transgenic mouse models to study various cancers and their metastatic models.
Retroviruses that cause tumor growth include Rous sarcoma virus and mouse mammary tumor virus. Cancer can be triggered by proto-oncogenes that were mistakenly incorporated into proviral DNA or by the disruption of cellular proto-oncogenes. Rous sarcoma virus contains the src gene that triggers tumor formation. Later it was found that a similar gene in cells is involved in cell signaling, which was most likely excised with the proviral DNA. Nontransforming viruses can randomly insert their DNA into proto-oncogenes, disrupting the expression of proteins that regulate the cell cycle. The promoter of the provirus DNA can also cause over expression of regulatory genes. Retroviruses can cause diseases such as cancer and immunodeficiency. If viral DNA is integrated into host chromosomes, it can lead to permanent infections. It is therefore important to discover the body's response to retroviruses. Exogenous retroviruses are especially associated with pathogenic diseases. For example, mice have mouse mammary tumor virus (MMTV), which is a retrovirus. This virus passes to newborn mice through mammary milk. When they are 6 months old, the mice carrying the virus get mammary cancer because of the retrovirus. In addition, leukemia virus I (HTLV-1), found in human T cell, has been found in humans for many years. It is estimated that this retrovirus causes leukemia in the ages of 40 and 50.[30] It has a replicable structure that can induce cancer. In addition to the usual gene sequence of retroviruses, HTLV-1 contains a fourth region, PX. This region encodes Tax, Rex, p12, p13 and p30 regulatory proteins. The Tax protein initiates the leukemic process and organizes the transcription of all viral genes in the integrated HTLV proviral DNA. [31]
Exogenous retroviruses are infectious RNA- or DNA-containing viruses that are transmitted from one organism to another. In the Baltimore classification system, which groups viruses together based on their manner of messenger RNA synthesis, they are classified into two groups: Group VI: single-stranded RNA viruses with a DNA intermediate in their life cycle, and Group VII: double-stranded DNA viruses with an RNA intermediate in their life cycle.
All members of Group VI use virally encoded reverse transcriptase, an RNA-dependent DNA polymerase, to produce DNA from the initial virion RNA genome. This DNA is often integrated into the host genome, as in the case of retroviruses and pseudoviruses, where it is replicated and transcribed by the host.
Group VI includes:
The family Retroviridae was previously divided into three subfamilies (Oncovirinae, Lentivirinae, and Spumavirinae), but are now divided into two: Orthoretrovirinae and Spumaretrovirinae. The term oncovirus is now commonly used to describe a cancer-causing virus. This family now includes the following genera:
Note that according to ICTV 2017, genus Spumavirus has been divided into five genera, and its former type species Simian foamy virus is now upgraded to genus Simiispumavirus with not less than 14 species, including new type species Eastern chimpanzee simian foamy virus.[32]
Both families in Group VII have DNA genomes contained within the invading virus particles. The DNA genome is transcribed into both mRNA, for use as a transcript in protein synthesis, and pre-genomic RNA, for use as the template during genome replication. Virally encoded reverse transcriptase uses the pre-genomic RNA as a template for the creation of genomic DNA.
Group VII includes:
The latter family is closely related to the newly proposed
whilst families Belpaoviridae, Metaviridae, Pseudoviridae, Retroviridae, and Caulimoviridae constitute the order Ortervirales.[34]
Endogenous retroviruses are not formally included in this classification system, and are broadly classified into three classes, on the basis of relatedness to exogenous genera:
Retroviruses have been the focus of several recent claims and assertions which have been largely discredited by the science community. An initial study in 2009 seemed to make new findings which might change some of the established knowledge on this topic. However, although later research disproved some of the claims made about retroviruses, there are several controversial figures who continue to make claims which overall are considered to not have any valid basis or consensus in support of these claims.[35][36][37]
Antiretroviral drugs are medications for the treatment of infection by retroviruses, primarily HIV. Different classes of antiretroviral drugs act on different stages of the HIV life cycle. Combination of several (typically three or four) antiretroviral drugs is known as highly active antiretroviral therapy (HAART).[38]
Feline leukemia virus and Feline immunodeficiency virus infections are treated with biologics, including the only immunomodulator currently licensed for sale in the United States, Lymphocyte T-Cell Immune Modulator (LTCI).[39]
A retrovirus is a type of virus that inserts a DNA copy of its RNA genome into the DNA of a host cell that it invades, thus changing the genome of that cell. After invading a host cell's cytoplasm, the virus uses its own reverse transcriptase enzyme to produce DNA from its RNA genome, the reverse of the usual pattern, thus retro (backwards). The new DNA is then incorporated into the host cell genome by an integrase enzyme, at which point the retroviral DNA is referred to as a provirus. The host cell then treats the viral DNA as part of its own genome, transcribing and translating the viral genes along with the cell's own genes, producing the proteins required to assemble new copies of the virus. Many retroviruses cause serious diseases in humans, other mammals, and birds.
Retroviruses have many subfamilies in three basic groups.
Oncoretroviruses (cancer-causing retroviruses) include human T-lymphotropic virus (HTLV) causing a type of leukemia in humans, and murine leukemia viruses (MLVs) in mice. Lentiviruses (slow viruses) include HIV-1 and HIV-2, the cause of acquired immune deficiency syndrome (AIDS) in humans. Spumaviruses (foamy viruses) are benign and not linked to any disease in humans or animals.The specialized DNA-inflitration enzymes in retroviruses make them valuable research tools in molecular biology, and they have been used successfully in gene delivery systems.
Evidence from endogenous retroviruses (inherited provirus DNA in animal genomes) suggests that retroviruses have been infecting vertebrates for at least 450 million years.
Retroviridae es una familia de virus que comprende los retrovirus. Son virus con genoma de ARN monocatenario de polaridad positiva y se replican de manera inusual a través de una forma intermedia de ADN bicatenario. Este proceso se lleva a cabo mediante una enzima: la retrotranscriptasa o transcriptasa inversa, que dirige la síntesis de ADN a través de ARN y posee una importancia extraordinaria en la manipulación genética. Una vez que se ha pasado de ARN monocatenario a ADN, se inserta dentro del ADN propio de la célula infectada donde se comporta como un gen más (véase Ciclo reproductivo de los virus).[1] Por tanto, se incluyen en el Grupo VI de la Clasificación de Baltimore.
Los retrovirus son responsables de muchas enfermedades, incluyendo algunos cánceres y el sida (VIH).[2] Existen diversos grupos de investigación que han intentado modificar genéticamente los retrovirus para usarlos en terapia génica como vectores, pero se han encontrado con diversos problemas.
La familia incluye los siguientes géneros:
El genoma del virus toma la forma de un ARNm de polaridad positiva, incluida la cap 5' y la poly-A 3' dentro del virión. Una vez dentro de la célula del huésped, la cadena de ARN se somete a la transcripción inversa en el citosol y es integrado en el genoma del huésped, momento en que el ADN retroviral se denomina provirus.
En el caso del VIH, el genoma consta de dos moléculas de ARN de cadena simple y polaridad positiva. Las moléculas de ARN están físicamente unidas mediante puentes de hidrógeno en sus extremos 5', lo que hace que sea difícil la encapsidación de más de 2 moléculas en un provirus.
La organización genómica es siempre la misma, 5'-Gag-Pol-env-3', y además dependiendo del tipo de retrovirus, hay genes accesorios que se solapan con los genes principales.
A pesar de la inmensa variabilidad entre los distintos tipos de retrovirus, podemos decir que la partícula viral se compone de:
El ciclo de replicación comprende varias etapas comunes a todos los retrovirus. En una fase inicial o temprana, el virus se une a receptores específicos de la célula gracias a la glicoproteína de superficie. Las membranas vírica y celular se fusionan y la cápside viral entra en la célula.
Las enzimas víricas permanecen asociadas al ARN genómico formando una complejo nucleoproteico. La síntesis de ADN vírico, incluyendo las LTR (Long Terminal Repeat;Repetición Terminal Larga) , se produce en el citoplasma a través de la Transcriptasa reversa RT. La actividad ARNasa H de la RT degrada la hebra de ARN y casi simultáneamente emplea la del ADNc como molde para sintetizar una segunda hebra de ADN, convirtiéndolo en bicatenario. Este permanece unido al complejo nucleoproteico, con el que pasa al núcleo celular y, mediante la Integrasa (IN), el ADN viral se integra (provirus) en el genoma celular, donde puede permanecer por un tiempo indefinido (en ocasiones luego de esta fase, puede pasar a transformarse en un retrovirus endógeno si logra infectar una célula germinal).
Luego viene la fase de trascripción de los genes, originando ARN que sirven como genoma del nuevo virión y ARN mensajeros para las poliproteínas. Las proteínas de gag y pol se asocian con el ARN viral formando un “core” intracelular, mientras que las proteínas de env se insertan en la membrana plasmática de la célula.
Tras el ensamblaje, se produce la salida de la célula por gemación, durante la cual el virus adquiere la doble capa lipídica de su envoltura. Estudios recientes sugieren que la salida de la partícula viral se produce en lugares determinados de la célula. Parece ser que Gag migraría hacia zonas de la membrana ricas en determinados lípidos; son las llamadas balsas o raft. Las proteínas asociadas a estos raft que se incorporan a la envuelta del virus desempeñarían también un papel importante en la replicación viral.
Finalmente se produce la maduración de las partículas gemadas mediante el procesamiento de las poliproteínas por la Proteasa (PR) viral que corta las poliproteínas precursoras. Los principales productos son las proteínas de la matriz (p17), cápside (p24), nucleocápside (p7) y la p6 (proteína). La proteasa es solamente activa dentro del virión. La maduración del VIH constituye un mecanismo importante para el diseño de antirretrovirales.
Existen 5 [cita requerida] retrovirus humanos identificados: el virus de la inmunodeficiencia humana de tipo 1 (VIH-1), el de tipo 2 (VIH-2) (Véase Subtipos de VIH) y los virus linfotrópicos de células T humanas de tipo I y II (HTLV-I y HTLV-II). Todos se hospedan en los linfocitos T. Los virus de la inmunodeficiencia humana producen la lisis de las células que infectan provocando una severa inmunodepresión. Los virus HTLVI/ II producen la inmortalización de los linfocitos infectados, generando una replicación descontrolada de los mismos, y por lo tanto una linfoproliferación.[3]
El síndrome de inmunodeficiencia adquirida o sida, es la expresión final de la infección por el VIH. La infección por este virus ocasiona la destrucción del sistema inmunitario además de manifestaciones neurológicas y tumorales. Estas manifestaciones clínicas se deben al tropismo tanto macrofágico como linfocitario del virus. Presenta una preferencia para infectar a linfocitos TCD4+, en los que la replicación es activa y muy agresiva, lo que provoca como característica de la infección una profunda inmunosupresión. La fisiopatología del sida es un proceso complejo, donde existen implicados mecanismos patogénicos tan diferentes que algunos hasta hoy no han sido completamente comprendidos. Los principales mecanismos de transmisión de la infección por VIH son sexual, parenteral y sanguínea.
Al igual que el VIH-1, el VIH-2 es un lentivirus. Su genoma está compuesto por 2 cadenas simples de ARN de polaridad positiva, y también contiene la enzima RT, que permite la integración del material genético del virus, como forma de provirus en el genoma de la célula que infecta, que es generalmente los linfocitos T CD4+. Comparte con el VIH-1 un 40-50% de homología genética, lo que hace necesario disponer de técnicas de biología molecular específicas para diagnosticar la infección por VIH-2. La distribución geográfica del VIH-2 está restringida prácticamente al continente africano y parece ser que la patología producida es mucho más leve y lenta que la causada por el VIH-1. El mecanismo de transmisión es igual que el VIH-1.
El virus linfotrópico de células T humano es un retrovirus que pertenece a la subfamilia Oncovirinae.
El HTLV-I fue el primer retrovirus oncógeno humano conocido. Puede provocar una hemopatía maligna denominada leucemia/linfoma de células T del adulto (ATL) y también una mielopatía subaguda denominada Paraparesia Espástica Tropical (PET) o mielopatía asociada al HTLV-I (HAM).
El HTLV-II es un virus que no tiene una patología claramente definida, aunque se lo ha asociado con diversos síndromes neurológicos y mielopatías subagudas.
El material genético está formado por 2 moléculas de ARN de cadena simple y polaridad positiva. El HTLV-I tiene como diana principal los linfocitos TCD4+ y el HTLV-II los linfocitos TCD8+.
El HTLV, una vez que ha infectado a la célula, puede permanecer latente integrado en forma de provirus o comenzar a replicarse. Se cree que el principal mecanismo de trasmisión de la infección por HTLV es a partir de mitosis de las células que infecta. Esta expansión clonal da lugar a lo que se denomina carga proviral. El HTLV necesita el contacto célula-célula para producir la infección;los principales mecanismos de transmisión de la infección por virus HTLV son por vía sexual, vía parenteral y vía vertical.
El XMRV o Xenotropic Murine Retrovirus es un virus del tipo gamma-retrovirus. Se cree que es causante de cáncer de próstata y Síndrome de fatiga crónica.
La terapia génica consiste en insertar copias funcionales de un gen defectuoso en el genoma de un individuo. Se han considerado los retrovirus como vectores génicos pero existen muchos problemas en su utilización.
Retroviridae es una familia de virus que comprende los retrovirus. Son virus con genoma de ARN monocatenario de polaridad positiva y se replican de manera inusual a través de una forma intermedia de ADN bicatenario. Este proceso se lleva a cabo mediante una enzima: la retrotranscriptasa o transcriptasa inversa, que dirige la síntesis de ADN a través de ARN y posee una importancia extraordinaria en la manipulación genética. Una vez que se ha pasado de ARN monocatenario a ADN, se inserta dentro del ADN propio de la célula infectada donde se comporta como un gen más (véase Ciclo reproductivo de los virus). Por tanto, se incluyen en el Grupo VI de la Clasificación de Baltimore.
Los retrovirus son responsables de muchas enfermedades, incluyendo algunos cánceres y el sida (VIH). Existen diversos grupos de investigación que han intentado modificar genéticamente los retrovirus para usarlos en terapia génica como vectores, pero se han encontrado con diversos problemas.
Retroviirused (Retroviridae) on viiruste sugukond, mille genoom koosneb kahest RNA molekulist ja mille genoomis kodeeritud revertaasi ehk pöördtranskriptaasi abil kopeerib viirus end DNA vormi ja suudab integraasi abil liituda peremeesorganismi pärilikkusainega.
Integreerunud retroviiruse genoomi nimetatakse proviiruseks.
Sugukonda Retroviridae liigitatakse järgmised viiruste alamsugukonnad[1]:
Retroviirused (Retroviridae) on viiruste sugukond, mille genoom koosneb kahest RNA molekulist ja mille genoomis kodeeritud revertaasi ehk pöördtranskriptaasi abil kopeerib viirus end DNA vormi ja suudab integraasi abil liituda peremeesorganismi pärilikkusainega.
Integreerunud retroviiruse genoomi nimetatakse proviiruseks.
Erretrobirusak Retroviridae familian sailkatzen diren birusak dira. Birus hauen ezaugarri nabarmena kate bakarreko ARN izatean datza. ARN hori entzima berezi bati esker erreplikatzen da, alderantzizko transkriptasa-ri esker, hain zuzen ere:
Alderantzizko transkriptasak ARN molekula ADN-n eraldatzen du, transkriptasaren ohiko jarduera kontrako norabidean eginez.
Sortutako ADNa zelularen genoman txertatzen da, beste gene baten antzera egonik, eta zelularen genomarekin batera erreplikatuz. Zelularen material genetikoan kokatuta dagoen birusaren ADN-ri probirus deritzo, ziklo lisogenikoaren profago-arekin alderatu baitaiteke.
Erretrobirusek estalki bat dute kanpoaldean, kapsidearen gainetik. Estalkia glikoproteinaz osatuta dago.
Birus hauek zelularen hazkuntza eralda dezakete, zelulak normalak zelula tumoralak bihurtuz. Minbizi mota batzuk eragin ditzaketenez onkobirusen kategorian sartzen dira.
Erretrobirus baten genomak gutxienez hiru gene hauek izaten ditu:
Erretrobirusen barnean hainbat genero daude, Lentivirus, Deltaretrovirus eta Spumavirus garrantzitsuenak izanik. Gizakia infektatzen dutenen artean hauek dira aipagarrienak:
Erretrobirusak oso erabiliak izan dira terapia genetikoan, bektore moduan.
Retrovirukset ovat kuorellisia viruksia, joiden genomi koostuu kahdesta yksijuosteisesta RNA-molekyylistä. Kun retrovirus infektoi isäntäsolun, viruksen oma käänteiskopioijaentsyymi valmistaa solussa viruksen RNA-koodia vastaavaa DNA:ta, joka kiinnittyy isäntäsolun perimään muuttuen provirukseksi.
Retroviruksien nimi (lat. retro ’takaperin’) johtuu juuri niiden omalaatuisesta kyvystä muodostaa DNA:ta RNA:n perusteella, kun yleisemmin eliökunnassa RNA syntyy transkriptiossa DNA-koodin perusteella. Käänteiskopioijaentsyymin löytämisestä myönnettiin Nobelin lääketieteen palkinto vuonna 1975.
Käänteiseen transkriptioon ei sisälly samaa oikaisulukua kuin normaaliin transkriptioon. Siksi retroviruksiin syntyy usein mutaatioita, minkä takia niitä vastaan on vaikeaa kehittää rokotteita. Esimerkiksi HIV on retrovirus.
Retrovirukset ovat eläinkunnassa varsin kauan tunnettuja ja laajalle levinneitä viruksia, jotka aiheuttavat monissa eläimissä kasvaimia. Kanoissa ja kissoissa niiden on todettu aiheuttavan leukemiaa, hiirissä rintasyöpää.
Ihmisiin tarttuvia retroviruksia ovat mm. kaksi eri leukemioihin liittyvää tyyppiä, HTLV-1 ja -2, sekä HIV.
Endogeenisten retrovirusten (ERV) perusteella voidaan nykyään päätellä eliöiden sukulaisuussuhteita. Simpanssilla ja ihmisellä on useita samoja endogeenisia retroviruksia.[1]
Retrovirukset ovat kuorellisia viruksia, joiden genomi koostuu kahdesta yksijuosteisesta RNA-molekyylistä. Kun retrovirus infektoi isäntäsolun, viruksen oma käänteiskopioijaentsyymi valmistaa solussa viruksen RNA-koodia vastaavaa DNA:ta, joka kiinnittyy isäntäsolun perimään muuttuen provirukseksi.
Retroviruksien nimi (lat. retro ’takaperin’) johtuu juuri niiden omalaatuisesta kyvystä muodostaa DNA:ta RNA:n perusteella, kun yleisemmin eliökunnassa RNA syntyy transkriptiossa DNA-koodin perusteella. Käänteiskopioijaentsyymin löytämisestä myönnettiin Nobelin lääketieteen palkinto vuonna 1975.
Käänteiseen transkriptioon ei sisälly samaa oikaisulukua kuin normaaliin transkriptioon. Siksi retroviruksiin syntyy usein mutaatioita, minkä takia niitä vastaan on vaikeaa kehittää rokotteita. Esimerkiksi HIV on retrovirus.
Retrovirukset ovat eläinkunnassa varsin kauan tunnettuja ja laajalle levinneitä viruksia, jotka aiheuttavat monissa eläimissä kasvaimia. Kanoissa ja kissoissa niiden on todettu aiheuttavan leukemiaa, hiirissä rintasyöpää.
Ihmisiin tarttuvia retroviruksia ovat mm. kaksi eri leukemioihin liittyvää tyyppiä, HTLV-1 ja -2, sekä HIV.
Endogeenisten retrovirusten (ERV) perusteella voidaan nykyään päätellä eliöiden sukulaisuussuhteita. Simpanssilla ja ihmisellä on useita samoja endogeenisia retroviruksia.
Les Retroviridae (rétrovirus) sont une famille de virus qui regroupe les sous-familles suivantes : Orthoretrovirinae (en) et Spumaretrovirinae (en). Ce sont des virus à ARN monocaténaire de polarité positive infectant les vertébrés. Ils se distinguent notamment par la présence d'une enzyme virale : la transcriptase inverse (TI, ou encore RT pour reverse transcriptase), qui rétrotranscrit leur génome d'ARN en ADN pour être intégré par la suite dans le génome de la cellule hôte. La TI a la particularité de commettre relativement facilement des erreurs, ce qui fait que certains rétrovirus ont une grande variabilité génétique. Les Retroviridae disposent d'un fort pouvoir oncogène.
Le virus de l'immunodéficience humaine (VIH), responsable du Sida, est un rétrovirus. À cause de sa grande variabilité, il est très compliqué pour les chercheurs d'élaborer un vaccin, car les glycoprotéines transmembranaires de l'enveloppe du virus (GP120 et GP41) mutent constamment.

Cycle de vie d'un rétrovirus endogène:
Ce sont des virus enveloppés, de 125 génome est diploïde, les deux brins monocaténaires d'ARN sont reliés par des ponts hydrogènes à leur extrémité 5'.
Pour pouvoir s'intégrer dans le génome de la cellule hôte et synthétiser ses protéines virales via la machinerie cellulaire, qui est bicaténaire, la transcriptase inverse va synthétiser les deuxièmes brins manquants pour donner un ADN et non plus un ARN. Le brin d'ARN étant monocaténaire, des erreurs de transcription surviennent fréquemment (il n'y a pas de contrôle possible à l'aide du nucléotide complémentaire) ; si certaines aboutissent à un ADN improductif, d'autres sont viables et engendrent des mutants qui peuvent éventuellement différer par leur signature antigénique. Cette grande variabilité rend difficile la vaccination.
Le génome se décompose en différentes régions, ayant chacune un rôle bien défini. Orienté de 5' vers 3' :
Puis, suivent les trois gènes de structure :
Enfin, la dernière région :
Ils sont l'un des moteurs de la biodiversité, par leurs propriétés de transposition[2]. Les rétrovirus sont un cas particulier, avec une « transposition » qui propage dans ce cas une infection à partir de la cellule pénétrée par le virus aux cellules et organes voisins[2].
Un rétrotransposon peut être converti en agent infectieux par l'acquisition d'une glycoprotéine d'enveloppe (« capture d’enveloppe »), ce qui a permis l'apparition de rétrovirus infectieux de mammifères[2]. Ces captures expliqueraient les parentés étroites de protéines Env (protéines d'enveloppe) de différents rétrovirus pourtant phylogéniquement distants. Comprendre ces « étymologies » rétrovirales est nécessaire pour l'établissement de la phylogénie des rétrovirus et la physiopathologie des infections qu'ils produisent, et peut-être pour la découverte de nouveaux rétroéléments[2].
Certains rétrovirus possèdent, en plus, un oncogène. Cet oncogène code une protéine de transformation. Cette protéine confère aux virus un fort pouvoir de transformation leur permettant d'induire des tumeurs très rapidement (quelques semaines) après l'infection. Les protéines oncogéniques peuvent être de différentes natures :
On classe les rétrovirus en deux grandes catégories :
Seuls les rétrovirus exogènes sont formellement classifiés par l'International Committee on Taxonomy of Viruses (ICTV)[3] et sont regroupés dans deux sous-familles :
Ces deux sous-familles comprennent au total sept genres : Alpharetrovirus, Betaretrovirus, Gammaretrovirus, Deltaretrovirus, Epsilonretrovirus, Lentivirus et Spumavirus.
Selon leur aspect infectieux, les rétrovirus sont classés dans trois catégories :
L'ADN de toutes les espèces, mammifères y compris, comporte des génomes viraux laissés par des virus, probablement à l'occasion d'une infection de cellules germinales, puis dupliqués avec l'ADN de génération en génération. Ces rétrovirus endogènes peuvent être présents en très grand nombre dans les génomes, où ils sont présents en de multiples exemplaires. Ils appartiennent à ce qu'on appelle les répétitions dispersées. Le génome humain contient environ 450 000 exemplaires de rétrovirus endogènes, plus ou moins délétés ou mutés depuis leur insertion dans la lignée germinale humaine[4].
L'expression des gènes des rétrovirus endogènes est normalement inhibée par une protéine spécialisée dite « KAP1 », dès le début de l'embryogenèse[5]. L'étude des rétrovirus endogènes est un enjeu médical, car ils peuvent être source de diverses maladies (dont des cancers) quand leur génome est exprimé, malgré les protections naturelles mises en place au cours de l'évolution.
Dès le début, le caractère oncogène des rétrovirus a été observé. C'est ainsi qu'en 1908, le virus de la leucémie aviaire (ALV) a été transmis d'un poulet à un autre par les Danois Vilhelm Ellerman et Oluf Bang après transfert de tissus infectés. En 1911, c'est au tour de Peyton Rous de mettre en évidence le virus du sarcome de Rous (RSV), qui, contrairement à l'ALV, induit une tumeur quelques semaines après l'infection, contre plusieurs mois pour l'ALV[6].
Mais il faudra attendre plus de quarante ans pour observer un rétrovirus chez les mammifères, avec la découverte du virus de la leucémie murine (MLV) en 1957 par Ludwig Gross et enfin 1981 pour la découverte du HTLV-1 par Robert Gallo, faisant de ce virus le premier rétrovirus humain identifié. Très rapidement d'autres virus sont identifiés : HTLV-2 en 1982 et surtout le VIH en 1983.
La découverte de ce dernier et la pandémie que l'on connaît depuis ont poussé les institutions de recherche publiques et privées, ainsi que l'industrie pharmaceutique, à faire des rétrovirus les virus les plus étudiés au monde. Une nouvelle classe d'antiviraux a été mise au point, pour s'attaquer à des particularités des rétrovirus, et regroupe ainsi ceux appelés antirétroviraux.
Des manipulations génétiques ont montré que l'on pouvait utiliser des rétrovirus pour amener des gènes particuliers dans une cellule humaine. Les chercheurs espèrent ainsi introduire dans l'ADN de personnes atteintes de déficiences génétiques héréditaires les gènes qui leur font défaut. On pourrait ainsi infecter positivement un organisme humain et le guérir de maladies génétiques. Une maîtrise totale de ces techniques pourrait à long terme permettre de réintroduire dans le génome humain les gènes déficients communs à l'espèce humaine, comme ceux permettant de synthétiser certaines vitamines (vitamine C par exemple), les acides aminés essentiels et les acides gras essentiels. Ces nutriments doivent actuellement être obligatoirement apportés par la nourriture.[réf. nécessaire]
Il y a sans doute une question bioéthique de fond pour réguler ces manipulations.[réf. nécessaire]
Víreas timpeall 100 nm ar leithead le truaill sheachtrach mórthimpeall ar an gcroíleacán. Bíonn an t-eolas géiniteach stóráilte ar mhóilín amháin aonduail RNA ann. Airí sainiúil de is ea go bhfuil einsím ar leith, cúltrascrioptáis, taobh istigh den pháirteagal víreasach.
Un retrovirus é un virus de ARN que se replica nunha célula hóspede facendo unha copia en ADN do seu xenoma. Os máis típicos pertencen á familia Retroviridae. Entre eles está o VIH causante da SIDA. O virus usa primeiramente o seu encima característico, a reversotranscritase para producir ADN a partir do seu xenoma de ARN, o que é o inverso do procedemento normal (transcrición: ADN→ARN). Este novo ADN de orixe viral é despois incorporado ao xenoma da célula hóspede pola acción dun encima integrase. A célula despois trata ao ADN proviral incorporado como parte do seu propio xenoma, e segue as súas instrucións fielmente, fabricando as proteínas víricas codificadas nos xenes virais necesarias para a ensamblaxe e formación de novas copias do virus. Os retrovirus son virus con envoltura membranosa, que toman da célula infectada.
Unha variedade especial dos retrovirus son os retrovirus endóxenos (ERV) que están integrados no xenoma do hóspede e se herdan dunha xeración a outra.
Hai algúns virus con reversotranscrición que son virus de ADN e non pertencen á familia Retroviridae ou relacionadas (ver grupo VII en "Clasificación" máis abaixo).
O retrovirus almacena o seu material xenético en forma de ARNm (que ten os extremos modificados 5' cap e 3'poliA) dentro do virión, e este serve como medio para liberar o xenoma vírico dentro da célula, e iniciar a infección. Unha vez que está na célula hóspede, o ARN sofre a reversotranscrición no citoplasma e despois integra o seu ADN no xenoma celular. Cando está integrado recibe o nome de provirus.
O ADN proviral do retrovirus pode permanecer integrado moito tempo e non é detectado polo sistema inmunitario. Porén, o provirus nun determinado momento transcribe o seu ADN a ARN para expresar os xenes do virus e producir novas copias do virus. Por tanto, a información contida nun xene retroviral flúe na secuencia: ARN → ADN → ARN → proteína.
Os retrovirus son valiosas ferramentas de investigación en bioloxía molecular e foron utilizados con éxito en sistemas de transferencia de xenes.[1]
Os virións dos retrovirus son partículas envoltas duns 100 nm de diámetro. Os virións conteñen dúas moléculas idénticas de ARN monocatenario de 7-10 quilobases (kb) de lonxitude. Aínda que os virións de diferentes retrovirus non teñen a mesma morfoloxía ou bioloxía, todos os compoñentes do virión son moi semellantes.[2]
Os principais compoñentes do virión de retrovirus son:
Cando os retrovirus integran os seus propios xenomas en células da liña xerminal, os seus xenomas pasan de xeración en xeración. Deste modo orixínanse retrovirus endóxenos (ERVs), que hoxe forman o 5-8% do xenoma humano, e que en xeral levan moito tempo no xenoma. Contrastan cos retrovirus exóxenos, que son dos que trata principalmente este artigo.[6] A maioría das insercións de retrovirus endóxenos non teñen función coñecida e formarían parte do que se denominou "ADN lixo". Porén, moitos retrovirus endóxenos xogan importantes papeis na bioloxía do hóspede, como o control da transcrición de xenes, fusión celular durante o desenvolvemento da placenta no curso da xerminación dun embrión, e resistencia a infeccións por retrovirus exóxenos. Os retrovirus endóxenos recibiron tamén a atención dos investigadores da área da inmunoloxía.[7]
A transcrición ten lugar do ADN ao ARN, pero a reversotranscrición transcribe ARN a ADN. O prefixo "retro" en retrovirus refírese a esta inversión do fluxo xenético do dogma central da bioloxía molecular. A actividade de reversotrinscritase, ademais de nos retrovirus, foi atopada tamén nas células eucariotas, nas que permite a xeración e inserción de novas copias de retrotransposóns no xenoma do hóspede.
Cando o retrovirus inserta o seu xenoma no da célula hóspede, este transcríbese e orixina ARNms que saen ao citosol, onde son traducidos ás proteínas necesarias para formar os novos virus. É importante notar que un retrovirus debe traer con el a súa propia reversotranscritase dentro da súa cápside, xa que non pode utilizar os encimas das células infectadas.
As drogas terapéuticas denominadas inhibidores da protease e da reversotranscritase teñen como dianas sitios e secuencias específicos nos seus respectivos encimas. Porén, estas drogas poden perder rapidamente a súa efectividade debido a que as secuencias xénicas que codifican para a protease e a reversoptranscritase mutan rapidamente. Estes cambios de bases orixinan codóns e sitios específicos nos encimas, que cambian, e, por tanto, evitan ser as dianas das drogas ao perderen os sitios que as drogas utilizan para unirse a eles.
Como a reversotranscrición carece da habitual actividade de corrección de probas da replicación do ADN. Os retrovirus mutan con gran frecuencia. Isto permite que o virus aumente a súa resistencia aos fármacos antivirais rapidamente, e impide o desenvolvemento de vacinas efectivas e inhibidores para os retrovirus.[8]
Un inconveniente de moitos retrovirus, como o retrovirus da leucemia murina Moloney, implica o requirimento de que as células estean dividíndose activamente para que se produza a transdución, o que dificulta o seu uso na terapia xénica. Como resultado, as células como as neuronas son moi resistentes á infección e á transdución por retrovirus, xa que non se dividen. Sempre existe a preocupación de que a mutaxénese insercional debida á integración no xenoma do hóspede poida orixinar un cancro ou leucemia. Isto é diferente ao que fan os Lentiviridae, un subgrupo dentro dos Retroviridae que pode integrar o seu ARN no xenoma de células hóspede que non se están a dividir.
Hai tres modos de transmisión entre os retrovirus:
Os xenomas de retrovirus xeralmente conteñen tres marcos de lectura abertos que codifican as proteínas que poden encontrarse no virus maduro. Son:
A posesión dunha copia funcional do xene env é o que fai que os retrovirus sexan distintos dos retroelementos.[10] O xene env realiza tres funcións distintas: permite que o retrovirus entre e saia das células hóspede por medio do tráfico de membranas endosómicas na célula, protección do ambiente extracelular por medio da bicapa lipídica, e a capacidade de entrar nas células. A capacidade do retrovirus de unirse á súa célula hóspede utilizando receptores específicos da superficie celular dálla o compoñente de superficie (SU) do env, e a capacidade do retrovirus de entrar na célula por medio da fusión de membranas débese ao compoñente transmembrana ancorado na membrana (TM). Deste xeito, a proteína env é a que permite que o retrovirus sexa infeccioso.
É o ADN vírico incorporado ao xenoma da célula hóspede en forma de provirus, que pode pasar á proxenie da célula. Cando a célula se divide ten que replicar previamente o seu xenoma e ao facelo replica tamén o provirus integrado, polo que as células fillas levan tamén o provirus. O ADN dalgúns retrovirus insírese aleatoriamente no xenoma da célula hóspede, polo que pode ser inserido nun oncoxene. Deste modo algúns retrovirus poden converter as células normais en células cancerosas. Algúns provirus permanecen latentes na célula durante longos períodos antes de que son activados por algún cambio no seu ambiente.
O estudo dos retrovirus levou á primeira demostración da síntese de ADN a partir de moldes de ARN, un modo fundamental de transferir o material xenético entre especies. Especulouse que nas fases máis iniciais da evolución das células os procesos de reversotranscrición do ARN ao ADN utilizados polos retrovirus son os que puideron orixinar a aparición do ADN como material xenético. Neste modelo que se enmarca na hipótese do mundo de ARN, os organismos celulares adoptaron o ADN (molécula quimicamente máis estable) cando os retrovirus evolucionaron para crear ADN a partir de moldes de ARN.
Os vectores gammarretrovirais e lentivirais utilizados na terapia xénica median a modificación xenética estable de células tratadas por medio da súa integración no xenoma celular. Esta trecnoloxía é útil non só na investigación, senón tamén para a terapia xénica clínica que trata de corrixir os defectos xenéticos das células de forma permanente; por exemplo, en células nai proxenitoras. Desenvolvéronse partículas vector retrovirais que infectan determinados tipos de células. Os vectores gammarretrovirais e lentivirais foron xa usados ata o momento en máis de 300 ensaios clínicos, para tratar diversas doenzas.[1][11]
Os retrovirus que causan o crecemento de tumores inclúen o virus do sarcoma de Rous e o virus do tumor mamario de rato. O cancro pode ser desencadeado por protooncoxenes que foron incorporados por erro no ADN proviral ou pola disrupción de protooncoxenes celulares. O virus do sarcoma de Rous contén o xene src que desencadea a formación de tumores. Máis tarde atopouse que un xene similar das células estaba implicado na sinalización celular, o cal era moi probablemente extraído co ADN proviral. Os virus non transformantes poden inserir o seu ADN aleatoriamente en protooncoxenes, interrompendo a expresión de proteínas que regulan o ciclo celular. O promotor do ADN do provirus pode tamén causar a sobreexpresión de xenes regulatorios.
Os virus que presentan reversotranscrición poden clasificarse da seguinte maneira:
Non se transmiten á proxenie directamente polo xenoma, xa que non están definitivamente integrados nel (só o fan durante as infeccións), polo que chegan ao corpo desde o exterior causando a infección. O grupo inclúe os seguintes xéneros:
Estes xéneros estaban antes clasificados en tres subfamilias (Oncovirinae, Lentivirinae, e Spumavirinae), pero co coñecemento que se ten actualmente dos retrovirus esta división xa non parece apropiada. (Porén, o termo oncovirus aínda se usa correntemente).
Os retrovirus clasificáronse en dous grupos na clasificación de Baltimore, que son os grupos VI e VII de virus. Os detalles da reversotranscrición non son iguais en todos eles.
Son virus de ARN. Todos os membros do grupo VI usan unha reversotranscritase codificada polo virus, que é unha ADN polimerase ARN dependente, para producir ADN a partir do xenoma inicial viral de ARN. O ADN é a miúdo integrado no xenoma do hóspede, como no caso dos retrovirus e pseudovirus, nos que o xenoma é replicado e transcrito polo hóspede.
O grupo VI inclúe as seguintes familias:
Ambas as familias do grupo VII teñen xenomas de ADN nas partículas que infectan as células. O xenoma de ADN é transcrito tanto a un ARNm (usado na síntese proteica) coma a un ARN prexenómico (usado como molde durante a replicación do xenoma vírico por reversotranscrición). Unha reversotranscritase codificada polo virus utiliza o ARN prexenómico como molde para a creación do ADN viral xenómico para as novas copias do virus.
O grupo VII inclúe as seguintes familias:
Os retrovirus endóxenos (ERV)s non están formalmente incluídos neste sistema de clasificación, xa que están integrados no xenoma e pasan directamente de xeración en xeración formando parte do xenoma e nunca están no exterior. Deberon orixinarse por infeccións retrovirais nos antepasados das especies hai moito tempo. Clasifícanse en tres clases baseándose na proximidade aos xéneros de retrovirus exóxenos:
As drogas antirretrovirais son medicacións usadas para o tratamento de infeccións por retrovirus, principalmente o VIH. Hai distintas clases de drogas antirretrovirais que actúan en diferentes estadios do ciclo de replicación do VIH. A combinación de varias drogas antirretrovirais (xeralmente tres ou catro) é unha activa terapia antirretroviral.[12]
As infeccións polo virus da leucemia felina e o virus da inmunodeficiencia felina poden tratarse con produtos como o inmunomodulador chamado inmunomodulador célula T-linfocito (LTCI).[13]
Un retrovirus é un virus de ARN que se replica nunha célula hóspede facendo unha copia en ADN do seu xenoma. Os máis típicos pertencen á familia Retroviridae. Entre eles está o VIH causante da SIDA. O virus usa primeiramente o seu encima característico, a reversotranscritase para producir ADN a partir do seu xenoma de ARN, o que é o inverso do procedemento normal (transcrición: ADN→ARN). Este novo ADN de orixe viral é despois incorporado ao xenoma da célula hóspede pola acción dun encima integrase. A célula despois trata ao ADN proviral incorporado como parte do seu propio xenoma, e segue as súas instrucións fielmente, fabricando as proteínas víricas codificadas nos xenes virais necesarias para a ensamblaxe e formación de novas copias do virus. Os retrovirus son virus con envoltura membranosa, que toman da célula infectada.
Unha variedade especial dos retrovirus son os retrovirus endóxenos (ERV) que están integrados no xenoma do hóspede e se herdan dunha xeración a outra.
Hai algúns virus con reversotranscrición que son virus de ADN e non pertencen á familia Retroviridae ou relacionadas (ver grupo VII en "Clasificación" máis abaixo).
O retrovirus almacena o seu material xenético en forma de ARNm (que ten os extremos modificados 5' cap e 3'poliA) dentro do virión, e este serve como medio para liberar o xenoma vírico dentro da célula, e iniciar a infección. Unha vez que está na célula hóspede, o ARN sofre a reversotranscrición no citoplasma e despois integra o seu ADN no xenoma celular. Cando está integrado recibe o nome de provirus.
O ADN proviral do retrovirus pode permanecer integrado moito tempo e non é detectado polo sistema inmunitario. Porén, o provirus nun determinado momento transcribe o seu ADN a ARN para expresar os xenes do virus e producir novas copias do virus. Por tanto, a información contida nun xene retroviral flúe na secuencia: ARN → ADN → ARN → proteína.
Os retrovirus son valiosas ferramentas de investigación en bioloxía molecular e foron utilizados con éxito en sistemas de transferencia de xenes.
Retrovirus merupakan salah satu golongan virus yang terdiri dari satu benang tunggal RNA (bukannya DNA). Setelah menginfeksi sel, virus tersebut akan membentuk replika DNA dari RNA-nya dengan menggunakan enzim reverse transcriptase.
Terdapat pada kera-kera kecil, atau kera besar macam gorila atau simpanse yang ada di benua Afrika, serta orangutan yang ada di Sumatra dan Kalimantan.
Ada tiga golongan retrovirus yang ditemukan pada primata yaitu oncornaviruses, lentiviruses, dan spumaviruses. Meskipun jumlahnya sangat sedikit, ketiga golongan virus tersebut beresiko menular pada manusia baik melalui gigitan, urin maupun feses (kotoran). Berikut virus-virus tersebut:
Ada empat jenis ornocavirus yang terdapat pada non human primata(ordo) (NHP) yaitu Simian T-lymphotropic virus (STLV), Gibbon ape leukemia virus (GaLV), Simian sarcoma virus, dan Simian retrovirus Type D (SRV). Simian T-lymphotropic virus (STLV) sangat mirip dengan Human T-cell leukemia virus (HTLV) yang banyak sekali terdapat di Asia, Afrika maupun Amerika. Meskipun kasus kejadiannya tidak banyak, HTLV dapat menyebabkan leukemia pada sel T dewasa atau lymphoma pada manusia yang terinfeksi. Selain itu, strain virus HTLV I juga berkaitan dengan tropical spastic paraparesis yaitu suatu gangguan saraf yang langka. Hal yang amat mengkhawatirkan, saat ini telah diketahui bahwa HTLV ternyata berasal dari STLV purba yang menular antar spesies yang berbeda. Bahkan sebuah survei yang dilakukan oleh Verschoor et al. (1998) terhadap 143 orangutan di Kalimantan Tengah menunjukkan adanya dua ekor orangutan yang terinfeksi oleh virus HTLV I. Dengan demikian, peluang virus golongan ini untuk menginfeksi manusia semakin besar. Gibbon ape leukemia virus (GaLV) juga dapat mengakibatkan leukemia meskipun hewan yang dijangkiti masih tampak sehat. Virus ini dapat berpindah antar spesies. Simian sarcoma virus, yang kemungkinan merupakan mutan dari GaLV diketahui menginfeksi monyet wooly yang serumah dengan gibbon. Simian retrovirus Type D (SRV) terdiri dari beberapa jenis virus. Virus ini biasanya menyerang monyet dan menyebabkan penurunan sistem kekebalan tubuh. Namun, monyet yang terserang virus ini tetap terlihat sehat. Antibodi terhadap retrovirus tipe D telah dilaporkan pada 2 dari 247 orang yang sehari-hari berhubungan dengan primata non manusia.
Salah satu golongan lentivirus yang amat berbahaya adalah Simian immunodeficiency virus (SIV). Virus ini berkerabat erat dengan HIV (Human Immunodeficiency Virus). Virus HIV 1 berasal dari strain SIV simpanse. Sedangkan virus HIV 2 berasal dari SIV sooty mangabeys. Ada sejumlah besar monyet Afrika baik yang liar maupun tangkapan yang terinfeksi oleh SIV. Jenis strainnya berbeda-beda, sesuai dengan jenis spesiesnya. Sebagian besar hewan yang terinfeksi oleh virus ini, tetap terlihat sehat. Primata Asia bukanlah induk semang alami dari SIV. Dengan demikian, apabila terkena SIV, primata Asia (termasuk orangutan) akan sangat mudah mengalami penurunan kekebalan tubuh. Saat ini ada 0.06% (2 dari 3123) manusia yang biasa bekerja dengan primata yang terinfeksi oleh virus ini. Satu di antara kedua orang tersebut selanjutnya menunjukkan hasil uji serologi yang negatif, tetapi yang lainnya tetap positif. Namun mereka berdua tidak menunjukkan gejala penyakit.
Spuma virus yang terdapat pada primata adalah Simian Foamy Virus (SFV). Virus ini banyak ditemukan pada primata dunia baru maupun lama. Ada 3,7% atau 11 dari 296 orang yang biasa berhubungan dengan primata telah terinfeksi oleh virus ini.
Virus dan Cara Penularan
(STLV) hubungan seksual dan air susu induk
(GaLV) urin, feses dan kemungkinan hubungan seksual
hubungan seksual, gigitan, dari induk ke anak
Simian Retrovirus-1 tipe D (SRV-1) ditemukan pertama kali antara tahun 1981-1983 sebagai penyebab sindrom imunosupresif yang diderita banyak primata di New England Primate Research Center (NEPRC) dan California Regional Primate Centres (CRPC), Amerika Serikat. Virus SRV-1 ini tidak berkerabat dekat dengan HIV-1, tetapi memiliki kemiripan dengan Mason Pfizer monkey virus (MPMV) yang diisolasi dari sel tumor payudara monyet betina. Pada spesies primata M. nemestrina, ditemukan SRV-2 yang cukup jauh kekerabatannya dibandingkan SRV-1 dan MPMV (SRV-3). SRV-2 tidak hanya menyebabkan imunosupresi fatal, tetapi juga diikuti dengan proliferasi sel vaskuler dan mesenkimal yang disebut fibromatosis retroperitoneal (RF). Infeksi SRV tidak menunjukkan aktivasi imunitas yang tinggi pada awal terjadinya infeksi, tetapi virus ini menyerang berbagai sel pertahanan tubuh seperti sel T CD4+ dan sel T CD8, sel B, makrofaga, dan sel epitelial di saluran pencernaan, kelenjar saliva, dan choroid plexus. Pada tahap akhir infeksi SRV, terjadi penurunan limfoid pada kelenjar limfoid dan infeksi oportunistik seperti yang juga terjadi pada tahap akhir dari infeksi HIV. Infeksi SRV pada primata menjadi model studi mekanisme supresi sistem imun yang menarik para ahli karena kemampuan reproduksi virus yang tinggi, transmisinya cepat, dan waktu turnover yang diperlukan relatif pendek (7-10 hari). Untuk mendeteksi keberadaan virus ini yang bersifat onkogenik ini, dapat digunakan uji serologis seperti ELISA dan western blot, analisis PCR, ataupun isolasi virus secara langsung dengan menumbuhkan peripheral blood mononuclear cell (PBMC) asal primata dan sel raji secara bersamaan.
hubungan seksual, gigitan
(SIV)
gigitan yang dalam
Hingga saat AAZV mengeluarkan panduan untuk penanganan primata yang bukan manusia (15 Februari 2004), belum ada laporan resmi tentang terjadinya penyakit pada manusia akibat tertular virus-virus ini. Namun, karena virus adalah material genetik yang dapat bermutasi dengan mudah, sebaiknya kita tetap berhati-hati.
Retrovirus merupakan salah satu golongan virus yang terdiri dari satu benang tunggal RNA (bukannya DNA). Setelah menginfeksi sel, virus tersebut akan membentuk replika DNA dari RNA-nya dengan menggunakan enzim reverse transcriptase.
Terdapat pada kera-kera kecil, atau kera besar macam gorila atau simpanse yang ada di benua Afrika, serta orangutan yang ada di Sumatra dan Kalimantan.
I Retroviridae sono una famiglia di retrovirus provvisti di peplos, con capside isometrico e con genoma costituito da due molecole di RNA monocatenario a polarità positiva. Sono gli unici virus con genoma diploide.
Il genoma dei retroviridae è caratterizzato dalla presenza di tre sequenze del tutto peculiari, che assicurano lo svolgimento delle operazioni essenziali per la sopravvivenza del virus stesso. Queste sequenze sono:
Oltre ad esse, i diversi generi di retroviridae codificano anche per alcune proteine con funzione regolatoria o accessoria, particolarmente importanti nel virus HIV.
Il primo retrovirus individuato fu il virus della leucemia aviaria. Il più noto è il virus dell'immunodeficenza umana (HIV), indicato come agente eziologico dell'AIDS. Più in generale, i retrovirus umani noti che infettano l'uomo sono i virus HTLV, appartenenti al genere Deltaretrovirus che sono causa della Leucemia delle cellule T, ed i Lentivirus di cui fanno parte anche i due virus HIV-1, HIV-2. Precedentemente HIV-1 e HIV-2 furono chiamati da Robert Gallo con il nome di HTLV-III e HTLV-IV rispettivamente a causa della loro somiglianza con i Deltaretrovirus da lui identificati. Successivamente vennero inseriti in un loro genere i Lentivirus, con i loro attuali nomi: HIV-1 e HIV-2.
La replicazione dei retroviridae avviene nel citoplasma della cellula infettata. Essendo dotato di pericapside, il virione è in grado di fondersi con la membrana cellulare della cellula e di introdurvi quindi il nucleo-capside. Questo contiene al suo interno alcune molecole dell'enzima trascrittasi inversa, che è in grado di retrotrascrivere il genoma del virus da ssRNA+ a dsDNA. Il DNA neosintetizzato viene quindi integrato nel genoma della cellula ospitante (provirus), dove può rimanere silente per periodi di tempo molto variabili (molto lunghi nei Lentivirus, più rapidi in altri). È da sottolineare la necessità d'azione da parte della trascrittasi inversa per l'espressione del genoma virale, poiché l'RNA presente nel virione al momento della penetrazione, seppur a polarità positiva, non possiede una sequenza tale da permettere l'inizio della traduzione. Il DNA neosintetizzato invece è trascritto funzionalmente in mRNA e duplicato tramite l'apparato replicativo cellulare.
Una volta prodotti gli mRNA virali nel nucleo, approfittando dell'apparato trascrizionale della cellula, questi sono tradotti nel citoplasma dai ribosomi cellulari in proteine funzionali. Quindi il virus provvede all'assemblaggio del capside e del pericapside. In ultimo, le nuove unità di genoma virale vengono assemblate con il capside neoformato, che gemma attraverso la membrana plasmatica cellulare e, ricoperto così del pericapside, provvede a infettare nuove cellule.
I Retroviridae sono una famiglia di retrovirus provvisti di peplos, con capside isometrico e con genoma costituito da due molecole di RNA monocatenario a polarità positiva. Sono gli unici virus con genoma diploide.
Il genoma dei retroviridae è caratterizzato dalla presenza di tre sequenze del tutto peculiari, che assicurano lo svolgimento delle operazioni essenziali per la sopravvivenza del virus stesso. Queste sequenze sono:
gag, che codifica le proteine strutturali del nucleo-capside virale pol, che codifica per la trascrittasi inversa necessaria alla replicazione del virus env, che codifica le proteine del pericapside virale.Oltre ad esse, i diversi generi di retroviridae codificano anche per alcune proteine con funzione regolatoria o accessoria, particolarmente importanti nel virus HIV.
Retrovirus est virus cum RNA, non DNA. Informatio genetica retroviri ergo transcribitur in DNA, si virus se multiplicat in cellula infecta. Est encymum nomine reverse transcriptase, quid transcriptionem fungitur. Post transcriptionem informatio virogenica continet, transcribitur in mRNA ut DNA alia.
Retrovirorum genera sunt:
Endogenua retrovira tribus generibus discribuntur:
Retrovirus est virus cum RNA, non DNA. Informatio genetica retroviri ergo transcribitur in DNA, si virus se multiplicat in cellula infecta. Est encymum nomine reverse transcriptase, quid transcriptionem fungitur. Post transcriptionem informatio virogenica continet, transcribitur in mRNA ut DNA alia.
Retrovīruss ir RNS saturošs vīruss, kas vairojas, transkribējot savu RNS par DNS un iekļaujot šo DNS šūnas hromosomās. Retrovīrusi ir nozīmīga vēzi izraisošo vīrusu grupa.
Retrovirussen zijn virussen die hun erfelijk materiaal opslaan in de vorm van RNA. Bij de infectie van een gastheercel wordt de in het virale RNA vastgelegde genetische code naar DNA gekopieerd. De naam is gekozen omdat dit een omkering is van het normale procedé, waarbij de DNA-volgorde wordt omgezet in een RNA-volgorde.
Omdat de gastheercel in het algemeen niet in staat is om deze omzetting van RNA in DNA te maken, bevat het virus een gen voor reverse-transcriptase. Het enzym reverse-transcriptase (letterlijk: 'enzym voor achteruit overschrijven') zorgt ervoor dat het virale RNA kan worden gekopieerd in DNA in de gastheercel. Door middel van het enzym integrase kan dit DNA worden ingebouwd in het genoom van de gastheercel. Van enkelstrengs viraal RNA wordt door het enzym een streng DNA gemaakt, deze streng vormt samen met de virale RNA-streng een RNA-DNA-hybride. Nu bevat de enkele DNA-streng de genetische informatie van het virus. Het virale RNA wordt losgekoppeld van het hybride waarna in het ribosoom van de gastheercel een nieuwe DNA-streng gevormd wordt. Deze is complementair aan het viraal DNA dat zonet gesynthetiseerd is. De DNA-strengen combineren tot één DNA-dubbelstreng. Een restrictie-enzym knipt hierna aan het 5'- en 3'-eind van het ds-DNA (dubbelstrengs-DNA) een DNA-end. Hierdoor kan het DNA ingebouwd worden in het DNA van de gastheercel. Omdat het viraal DNA aanwezig is in het gastheercel-DNA kan constant de virale genetische code afgelezen worden en worden de eiwitbouwstenen van het virus gemaakt. Nieuw gesynthetiseerd viraal RNA vormt samen met een pakketje eiwitten een eiwitmantel, de eiwitmantel+RNA verlaat de gastheercel door middel van insnoering. Een nieuw virus is geboren en is nu in staat om andere cellen te infecteren.
Een zeer bekend retrovirus is het human immunodeficiency virus (hiv), de veroorzaker van aids en de variant voor katten: kattenaids. Retrovirussen vergroten ook vaak de kans op kanker (oncogeen), men noemt deze oncovirussen.
Indeling van de familie Retroviridae volgens de International Committee on Taxonomy of Viruses[1][2]:
Retrovirussen zijn virussen die hun erfelijk materiaal opslaan in de vorm van RNA. Bij de infectie van een gastheercel wordt de in het virale RNA vastgelegde genetische code naar DNA gekopieerd. De naam is gekozen omdat dit een omkering is van het normale procedé, waarbij de DNA-volgorde wordt omgezet in een RNA-volgorde.
Omdat de gastheercel in het algemeen niet in staat is om deze omzetting van RNA in DNA te maken, bevat het virus een gen voor reverse-transcriptase. Het enzym reverse-transcriptase (letterlijk: 'enzym voor achteruit overschrijven') zorgt ervoor dat het virale RNA kan worden gekopieerd in DNA in de gastheercel. Door middel van het enzym integrase kan dit DNA worden ingebouwd in het genoom van de gastheercel. Van enkelstrengs viraal RNA wordt door het enzym een streng DNA gemaakt, deze streng vormt samen met de virale RNA-streng een RNA-DNA-hybride. Nu bevat de enkele DNA-streng de genetische informatie van het virus. Het virale RNA wordt losgekoppeld van het hybride waarna in het ribosoom van de gastheercel een nieuwe DNA-streng gevormd wordt. Deze is complementair aan het viraal DNA dat zonet gesynthetiseerd is. De DNA-strengen combineren tot één DNA-dubbelstreng. Een restrictie-enzym knipt hierna aan het 5'- en 3'-eind van het ds-DNA (dubbelstrengs-DNA) een DNA-end. Hierdoor kan het DNA ingebouwd worden in het DNA van de gastheercel. Omdat het viraal DNA aanwezig is in het gastheercel-DNA kan constant de virale genetische code afgelezen worden en worden de eiwitbouwstenen van het virus gemaakt. Nieuw gesynthetiseerd viraal RNA vormt samen met een pakketje eiwitten een eiwitmantel, de eiwitmantel+RNA verlaat de gastheercel door middel van insnoering. Een nieuw virus is geboren en is nu in staat om andere cellen te infecteren.
Een zeer bekend retrovirus is het human immunodeficiency virus (hiv), de veroorzaker van aids en de variant voor katten: kattenaids. Retrovirussen vergroten ook vaak de kans op kanker (oncogeen), men noemt deze oncovirussen.
Retrovirus (Retroviridae) er ein familie av enkelttråda RNA-virus som nyttar dobbelttråda DNA som mellomsteg i økslinga. Dei har membranar[1] og positiv RNA-polaritet, og vert delte inn i dei to underfamiliane Orthoretrovirinae og Spumavirinae. DNA-utgåva av genomet til retrovirus har ein storleik på mellom 8 og 12 kilobasar, avhengig av virus. Dei har ryggradsdyr som vertar, inkludert pattedyr, fiskar, fuglar, krypdyr og amfibium.[2]
HIV-viruset er eit døme på eit retrovirus.[2]
Under økslinga nyttar retrovirus enzymet revers transkriptase (oppdaga i 1970) til å laga eit DNA-mellomsteg. Ein integrase vert deretter nytta til å lima inn mellomsteget i genomet til sella som vert infisert, og lagar slik eit provirus.[2][3] Denne prosessen lagar brot i DNA-molekylet proviruset vart sett inn i og som truleg lyt vølast av protein frå vertssella.[4] Etter innsetjinga i vertssellegenomet kan proviruset styra avskriving av DNA.[2]
Retroviruspartiklar knoppar seg ut av sellemembranen til ei vertsselle i ei umogen form. Fleire steg katalyserte av virusproteasar gjer partikkelen mogen slik at han kan gå inn i nye vertsseller.[5]
Alle retrovirus har dei tre gena gag, pro, pol og env.[2] gag kodar for strukturelle protein.[5] pro kodar for virusproteasen.[2] pol kodar for revers transkriptase og integrase.[2] env kodar for glykoprotein i virusmembranen som gjer det mogeleg for viruset å gå inn i ei vertsselle.[2][6]
Om sella retroviruset infiserer er ei kjønnsselle, kan det innsette virusgenomet gå i arv over fleire generasjon. Slike sekvendar vert kalla indre (endogene) retrovirus. Det er vanleg at genoma til ryggradsdyr inneheld hundretusenvis av uverksame DNA-sekvensar frå retrovirusgenom.[2] Det er rekna at 5 til 8 % av menneskegenomet er slike restar etter genoma til retrovirus.[7]
Retrovirus (Retroviridae) er ein familie av enkelttråda RNA-virus som nyttar dobbelttråda DNA som mellomsteg i økslinga. Dei har membranar og positiv RNA-polaritet, og vert delte inn i dei to underfamiliane Orthoretrovirinae og Spumavirinae. DNA-utgåva av genomet til retrovirus har ein storleik på mellom 8 og 12 kilobasar, avhengig av virus. Dei har ryggradsdyr som vertar, inkludert pattedyr, fiskar, fuglar, krypdyr og amfibium.
HIV-viruset er eit døme på eit retrovirus.
Retrovirus er en type virus som har RNA som arvestoff. De fleste av retrovirusene forårsaker sykdommer som blant annet aids og enkelte kreftformer hos pattedyr.
Disse virusene angriper levende celler (i motsetning til bakteriofager, som angriper bakterier) og binder seg til celleoverflaten, for så å sende inn RNA-et sitt i cellen.
RNA-tråden fra viruset bygger opp en DNA-tråd, som binder seg til vertscellens DNA. Deretter er alt som trengs en liten trigger (feks. ved en infeksjon), og cellen setter igang produksjon av nye virus.
Flere celler ødelegges og immunforsvaret reduseres.
Retrovirus er en type virus som har RNA som arvestoff. De fleste av retrovirusene forårsaker sykdommer som blant annet aids og enkelte kreftformer hos pattedyr.
Disse virusene angriper levende celler (i motsetning til bakteriofager, som angriper bakterier) og binder seg til celleoverflaten, for så å sende inn RNA-et sitt i cellen.
RNA-tråden fra viruset bygger opp en DNA-tråd, som binder seg til vertscellens DNA. Deretter er alt som trengs en liten trigger (feks. ved en infeksjon), og cellen setter igang produksjon av nye virus.
Flere celler ødelegges og immunforsvaret reduseres.
Retrowirusy (Retroviridae) – rodzina wirusów RNA (których materiał genetyczny zawarty jest w kwasie rybonukleinowym), które przeprowadzają proces odwrotnej transkrypcji.
Retrowirusy wywołują wiele chorób, w tym AIDS i niektóre nowotwory. Genom retrowirusa zawiera dwie identyczne kopie jednoniciowego RNA i koduje odwrotną transkryptazę (inaczej rewertazę), która ma zdolność przepisywania informacji z RNA na DNA.
Najdokładniej poznanym retrowirusem jest wirus HIV.
Wyróżnia się 7 rodzajów retrowirusów:[1]
Retrowirusy (Retroviridae) – rodzina wirusów RNA (których materiał genetyczny zawarty jest w kwasie rybonukleinowym), które przeprowadzają proces odwrotnej transkrypcji.
Retrowirusy wywołują wiele chorób, w tym AIDS i niektóre nowotwory. Genom retrowirusa zawiera dwie identyczne kopie jednoniciowego RNA i koduje odwrotną transkryptazę (inaczej rewertazę), która ma zdolność przepisywania informacji z RNA na DNA.
Najdokładniej poznanym retrowirusem jest wirus HIV.
Retroviridae é uma família de vírus que possuem genoma constituído por RNA fita simples senso positivo e que replicam o RNA viral por meio de um processo denominado transcrição reversa, onde moléculas de DNA dupla fita (dsDNA) são geradas a partir de RNA, pela ação da enzima transcriptase reversa. Esta família viral pertence ao grupo VI (vírus ssRNA-RT) no Sistema de Classificação de Baltimore.[1][2] Ledy do Horto dos Santos Oliveira [nota 1]
De maneira geral, o ciclo viral dos retrovírus ocorre da seguinte maneira:[3]
_RNA_synthesis.svg)
Os seguintes gêneros são incluídos dentro da família Retroviridae :[6]
Quando utilizados como vetores na terapia gênica somática, alteram de forma permanente a célula hospedeira, integrando-se ao genoma. São utilizados especialmente em células dos músculos liso e esquelético, da medula óssea, fibroblastos e hepatócitos.[carece de fontes?]
São retrovírus com comportamento infeccioso típico, disseminando-se horizontalmente por contacto. Muitos retrovírus exógenos são recombinantes produzidos em laboratório ou por co-infecção casual de um animal, não se encontrando como provírus endógeno na natureza.[8]
Uma cópia DNA completa do genoma (provírus), em determinadas situações, em muitas espécies de retrovírus, pode ser transmitida ao DNA da linhagem germinal materna à prole por herança mendeliana. Perpetua-se, assim, esse DNA em todas as células de um indivíduo em algumas espécies de vertebrados. Ex: retrovírus PERV-A e PERV-B em suínos. Esses genomas provirais são controlados pelos genes reguladores das células e normalmente são silenciosos devido à mutações, deleções ou transposições. Seqüências endógenas de retrovírus representam 8% do genoma humano. Sugere-se que esses provírus possam ser ativados por diversos fatores como radiações, exposição a substâncias químicas mutagênicas ou carcinogênicas, hormônios e etc.[9]
Retroviridae é uma família de vírus que possuem genoma constituído por RNA fita simples senso positivo e que replicam o RNA viral por meio de um processo denominado transcrição reversa, onde moléculas de DNA dupla fita (dsDNA) são geradas a partir de RNA, pela ação da enzima transcriptase reversa. Esta família viral pertence ao grupo VI (vírus ssRNA-RT) no Sistema de Classificação de Baltimore.
Un retrovirus este un virus ARN care se poate replica în celula gazdă. Folosește propria enzimă numită reverstranscriptza pentru a produce ADN din ARN și astfel „întoarce” șablonul inițial, de unde și prefixul „retro” (invers). Noua secvență de ADN este incorporată în genomul gazdei cu ajutorul unei enzime numite integrază. Celula gazdă se comportă cu noua secvență de ADN ca și cum aceasta ar face parte din propriile sale instrucțiuni, pe care le urmează necondiționat, făcând proteinele necesare pentru asamblarea noilor copii ale virusului. O categorie specială de retrovirusuri o constituie retrovirusurile endogene care sunt integrate in genomul gazdei și pot fi moștenite și transmise de la o generație la alta. Virusul păstrează informația sa genetică sub forma de ARN mesager; se integrează în genomul celulei gazdă, devine un parazit și declanșează infecții. Informația unui retrovirus produce proteine urmând pașii ARN -> ADN -> ARN-> proteine.
Se estimează că aproximativ 8% din genomul uman e reprezentat de cod genetic provenit de la retrovirusuri.[1]
Un retrovirus este un virus ARN care se poate replica în celula gazdă. Folosește propria enzimă numită reverstranscriptza pentru a produce ADN din ARN și astfel „întoarce” șablonul inițial, de unde și prefixul „retro” (invers). Noua secvență de ADN este incorporată în genomul gazdei cu ajutorul unei enzime numite integrază. Celula gazdă se comportă cu noua secvență de ADN ca și cum aceasta ar face parte din propriile sale instrucțiuni, pe care le urmează necondiționat, făcând proteinele necesare pentru asamblarea noilor copii ale virusului. O categorie specială de retrovirusuri o constituie retrovirusurile endogene care sunt integrate in genomul gazdei și pot fi moștenite și transmise de la o generație la alta. Virusul păstrează informația sa genetică sub forma de ARN mesager; se integrează în genomul celulei gazdă, devine un parazit și declanșează infecții. Informația unui retrovirus produce proteine urmând pașii ARN -> ADN -> ARN-> proteine.
Se estimează că aproximativ 8% din genomul uman e reprezentat de cod genetic provenit de la retrovirusuri.
Poddružina Orthoretrovirinae
Poddružina Spumaretrovirinae
Retrovirusi so kroglasti virusi s premerom od 80 do 100 nm, imajo ovojnico z 8 nm velikimi peplomeri, notranjo vijačno nukleokapsido obdaja ikozaedrična kapsida. Genom je iz dveh enakih molekul linearno pozitivne polarne enovijačne RNK.[1] Virus HIV je tipičen predstavnik.
Retrovirusi imajo enovijačno, pozitivno polarno molekulo RNK, ki ne deluje kot mRNK, ampak jo virusna od RNK odvisna polimeraza DNK (reverzna transkriptaza) prepiše v dvojnovijačno molekulo DNK, ki se kot provirus vgradi v celično DNK. Provirus celična polimeraza RNK II prepiše v molekulo RNK, ki ima dvojno vlogo – hkrati je virusna genomska RNK in policitronska mRNK. Poliproteine, ki nastanejo, virusne proteaze cepijo v posamezne virusne proteine. Genomske RNK se združujejo v pare in predstavljajo diploidni genom novih virusov.[2][3]
Glede na njihov genom lahko retroviruse razdelimo na enostavne in kompleksne (Coffin in sod., 1997). Tako enostavni kot kompleksni retrovirusi imajo gag (specifični antigeni – kapsidni proteini), pro (proteaza), pol (polimeraza) in env (ovojnica – glikoproteini) gene.[4] Virusna DNK vsebuje dolga ponavljajoča se zaporedja na obeh koncih, t. i. LTR zaporedja (Long terminal repetas). LTR lahko razdelimo na U3, R in U5 regije. Virusni promotorji in prepisovalni ojačevalci se nahajajo v U3 regiji. R regija je nujna za reverzno prepisovanje in podvojevanje retrovirusov. U5 regija vsebuje zaporedja, katera pospešujejo reverzna prepisovanja. Navzdol (downstream) za 5' LTR je začetno vezavno mesto (PBS), katerega sekvenca je komplementarna delu celične tRNA. Različni virusi uporabljajo različne tRNA za začetek delovanja reverzne transkriptaze.[5]
Signal za sestavljanje (ψ) oziroma kapsidizacijo (E) so zaporedja, ki interagirajo z virusnimi proteini, da se doseže specifično sestavljanje virusne RNK. Pri MLV (murine leukemia virus) ψ najdemo v 5' neprevedljivi regiji, med PBS (tRNA vezavno mesto) in gag odprtim bralnim okvirjem, kjer je med drugim tudi idealno mesto za vstavljanje tuje RNK v virusne delce.[3]
Blizu 5' konca virusnega genoma se nahaja gag gen, ki nosi zapis za Gag poliproteine, ki sodelujejo pri izgradnji kapside. Zapis za reverzno transkriptazo in integrazo (omogoči vključitev nastale virusne DNK v celično DNK) se nahaja v pol regiji. Pri vseh retrovirusih najdemo env gen, ki nosi zapis za proteine ovojnice. Le ti se cepijo v transmembransko in površinsko domeno (SU). Sekvence, katere imajo zapis za virusno proteazo (Pro) vedno najdemo med gag in pol. Večinoma se prepisujejo kot del Gag poliproteina ali pa kot Gag-Pol poliproteina.[5]
Retrovirusi so kroglasti virusi s premerom od 80 do 100 nm, imajo ovojnico z 8 nm velikimi peplomeri, notranjo vijačno nukleokapsido obdaja ikozaedrična kapsida. Genom je iz dveh enakih molekul linearno pozitivne polarne enovijačne RNK. Virus HIV je tipičen predstavnik.
Retrovirus, en klass av virus som har ett hölje och vars genom (arvsmassa) genomgår omvänd transkription som innebär att virusets RNA omvandlas till DNA. DNA-kopian infogas i värdcellens genom. En annan typisk egenskap som kännetecknar retrovirus är deras förmåga att alltid integrera det virala genomet i cellens genom. Efter integrationen tillverkas nya kopior av virusets RNA, samt de proteiner som virusets gener kodar för. Dessa sätts sedan ihop till nya viruspartiklar.
Det enzym som behövs för att tillverka DNA från RNA kallas omvänt transkriptas. Retroviruspartikeln bär själv med sig detta enzym, och den omvända transkriptionen sker i den infekterade cellens cytosol. Virusets enzym saknar den "korrekturläsningsmekanism" som vanliga enzym för DNA-transkription har. Därför muteras viruset lätt, vilket i sin tur medför att viruset lätt utvecklar resistens mot läkemedel mot virusinfektioner.
I vanliga fall sker informationsflöden från DNA till RNA och därifrån eventuellt till protein; detta kallas inom molekylär genetik för den centrala dogmen. Retroviruset, som ser till att informationsflödet går från RNA till DNA, bryter mot detta. Retro i namnet syftar på detta tvärtom (eng. reverse) mot det vanliga.
Subfamiljer till retrovirus är:
Retrovirus, en klass av virus som har ett hölje och vars genom (arvsmassa) genomgår omvänd transkription som innebär att virusets RNA omvandlas till DNA. DNA-kopian infogas i värdcellens genom. En annan typisk egenskap som kännetecknar retrovirus är deras förmåga att alltid integrera det virala genomet i cellens genom. Efter integrationen tillverkas nya kopior av virusets RNA, samt de proteiner som virusets gener kodar för. Dessa sätts sedan ihop till nya viruspartiklar.
Det enzym som behövs för att tillverka DNA från RNA kallas omvänt transkriptas. Retroviruspartikeln bär själv med sig detta enzym, och den omvända transkriptionen sker i den infekterade cellens cytosol. Virusets enzym saknar den "korrekturläsningsmekanism" som vanliga enzym för DNA-transkription har. Därför muteras viruset lätt, vilket i sin tur medför att viruset lätt utvecklar resistens mot läkemedel mot virusinfektioner.
I vanliga fall sker informationsflöden från DNA till RNA och därifrån eventuellt till protein; detta kallas inom molekylär genetik för den centrala dogmen. Retroviruset, som ser till att informationsflödet går från RNA till DNA, bryter mot detta. Retro i namnet syftar på detta tvärtom (eng. reverse) mot det vanliga.
Subfamiljer till retrovirus är:
Lentivirus (HIV, SIV, FIV) Gammaretrovirus (XMRV) Spumavirus OnkovirusAltaile: Orthoretrovirinae
Altaile: Spumaretrovirinae
Retrovirüsler, Retroviridae virüs ailesine ait olan bütün virüsler.
RNA genomu taşıyan zaflı virüslerdir ve DNA aracılığıyla ikileşme yapabilirler. Diploit olan tek RNA virisüdür. ters transkriptaz enzimiyle RNA'dan DNA dönüşümü yapabilirler. 90 - 120 nm çapında, tek iplikçikli RNA'ya sahip, kompleks yapılı, zarflı virüslerdir. HIV virüsü bu ailedendir. Genetik bilgiyi değiştirebilen virüslerin genel adıdır. Bu ailede Oncovirinae, Lentivirinae ve Spumavirinae alt aileleri bulunur. Bu ailenin insan için en önemli virüsleri Human T Lenfotropik Virüs (HTLV) ve AIDS etkeni olan Human Immunodeficiency Virüs (HIV) bulunur.
Tıbbi virolojiye giriş ve genel prensipler
Retrovirüsler, Retroviridae virüs ailesine ait olan bütün virüsler.
RNA genomu taşıyan zaflı virüslerdir ve DNA aracılığıyla ikileşme yapabilirler. Diploit olan tek RNA virisüdür. ters transkriptaz enzimiyle RNA'dan DNA dönüşümü yapabilirler. 90 - 120 nm çapında, tek iplikçikli RNA'ya sahip, kompleks yapılı, zarflı virüslerdir. HIV virüsü bu ailedendir. Genetik bilgiyi değiştirebilen virüslerin genel adıdır. Bu ailede Oncovirinae, Lentivirinae ve Spumavirinae alt aileleri bulunur. Bu ailenin insan için en önemli virüsleri Human T Lenfotropik Virüs (HTLV) ve AIDS etkeni olan Human Immunodeficiency Virüs (HIV) bulunur.
Віріони ретровірусів сферичної форми, розміром 90 — 120 нм, мають суперкапсиду ліпопротеїнової природи з шипоподібними пепломерами. Капсид побудований за типом кубічної симетрії. Ретровіруси містять до 2% РНК, близько 65% білку , 30% ліпідів та 2% вуглеводів. Характерною структурною особливістю у віріонів ретровірусів є унікальна серцевина. Остання представлена власною капсулою (внутрішня білкова мембрана) та нуклеотидом, до складу якого входить РНК і білок. За будовою серцевини онковіруси поділяються на три типи: С, В та D. Віріони типу С (діаметр близько 100 нм.) мають крупну (74-80 нм.), розташовану центрально, серцевину. У віріонів типу В і D серцевини розташовані ексцентрично. У перших вона округлої а у останніх циліндричної форми.
Вірус термолабільний — при 56˚С руйнується протягом 15-ти хвилин, при 74˚С. за 16 секунд, чутливий до ефіру і хлороформу, руйнується при заморожуванні — відтаванні.
Геном ретровірусів, на відміну від всіх останніх представників царства вірусів, диплоїдний — представлений двома однаковими молекулами РНК, молекулярною масою близько 3 МД. Розмір геному 7-11 kb. Він містить чотири гени : (5'→3') gag, pol, env, src.
Стратегія реплікації ядерна:
У складі ретровірусів виявляється від 11 до 13 структурних білків. Найменше білків локалізовано у серцевині. Головний внутрішній білок Р27-Р30 розташований у мембрані, що оточує РНК. Два - п'ять білків гліколізовані і знаходяться у складі ліпопротеїнової оболонки. З ними пов'язана типоспецифічна характеристика вірусів. Групоспецифічні антигени знаходяться у серцевині вірусу. Основним групоспецифічним антигеном є головний внутрішній білок Р27 - Р30. У віріонах онковірусів виявлено ряд білків з ферментативною активністю, зокрема РНК- залежна ДНК- полімераза, ДНК- залежна ДНК-полімераза, нуклеази, протеїнкінази і інші. Зворотна транскриптаза (РНК- залежна ДНК – полімераза) зв'язана з нуклеоїдом віріонів і представлена у онковірусів 10-20 молекулами на віріон. Молекулярна маса цього ферменту у віріонах типу С становить 70 кД, у В та D - 100-105кД. У онковірусів птахів ідентифіковано дві форми зворотної транскриптази : L (65кД) та В (105кД). У віріоні вони утворюють комплекс LB. Ліпіди знаходяться у складі ліпопротеїдної оболонки і мають клітинне походження. Вуглеводи входять до складу глікопротеїдів Глікопротеїди є типоспецифічними антигенами, беруть участь в реакції нейтралізації. Основним є глікопротеїд зовнішньої мембрани – SU.
Господар – хордові – примати, парнокопитні, непарнокопитні та хижаки.
Шляхи передачі в природі: трансмісивний шлях, парентеральний, вертикальний.
Переносники відсутні.
Географіче розповсюдження: по всьому світу.
Патогенез. У природних умовах патогенні для великої рогатої худоби. В експериментальних - захворюють телята, вівці. У природних умовах зараження чутливих тварин може відбуватись пренатально та частіше постнатально. Механізм пренатального зараження може являти собою передачу вірусного геному через гамети (генетична чи хромосомна трансляція ) або передачу сформованого вірусу (епігенетична екстрахромосомна трансляція). Постнатальна передача здійснюється через молоко чи під час прямого контакту. У останньому випадку вірус заноситься в організм з різноманітними секретами і екскретами або комахами чи контамінованими об'єктами (ін'єкційними голками, хірургічними інструментами тощо). Доведено, що вірус тривалий час персистує в організмі кліщів і може передаватись через них великій рогатій худобі.
Контактна передача — основний шлях розповсюдження лейкозу в природних умовах. Це потрібно мати на увазі тим, хто маніпулює в несприятливих по лейкозу стадах тварин.
Тропізм: найчастіше клітини імунної системи.
Патологічні зміни: знищують популяцію імунокомпетентних клітин, що призводить до загибелі особин від опортуністичних інфекцій. Порушення геному можуть викликати рак.
Вакцини для профілактики розробляються. Лікування ВІЛ – комбінована терапія з азидотимідином та інгібітором протеази ВІЛ. У природних умовах певне значення має колостральний імунітет, який триває кілька місяців і залежить від ряду обставин, зокрема від кількості молозивних імуноглобулінів.
У дорослих інфікованих тварин, попри наявність віруснейтралізуючих антитіл і інших факторів захисту, інфекційний процес не зупиняється. Штучна імунізація поки-що не набула широкого розповсюдження. Є повідомлення щодо позитивних результатів при використанні інактивованих препаратів. Двократне введення інактивованої 0,05%-ним АЕН, або 0,02% ДЕІ вакцини телятам обумовлено індукцію високих титрів віруснейтралізуючих антитіл та резистентність до експериментального зараження.
Retrovirus là một từ để gọi các loại virus mà vật chất di truyền của chúng là phân tử RNA. Khi chúng xâm nhập vào cơ thể thì có khả năng thực hiện quá trình phiên mã ngược nhờ enzim đặc biệt là enzyme phiên mã ngược - Reverse Transciptase. DNA sau đó là kết hợp vào gen của vật chủ bởi một enzyme tích hợp. Vi rút này sau đó sao chép như là một phần của DNA của tế bào chủ. Retrovirus là vi rút bao bọc es thuộc về gia họ virus "Retroviridae". Một biến thể đặc biệt của retrovirus retrovirus nội sinh được tích hợp vào bộ gen của vật chủ và di truyền qua nhiều thế hệ.
Màng của retrovirus có glycoprotein, protein dùng để liên kết với thụ quan protein trên tế bào vật chủ. Có hai sợi của RNA bên trong tế bào, chứa 3 enzyme: protease, reverse transcriptase và integrase. Bước đầu của quá trình sao chép là sự liên kết của glycoprotein với thụ quan protein. Khi đã có liên kết với nhau, màng của retrovirus sẽ phân giải thành một bộ phận của tế bào chủ để sợi RNA và enzyme có thể xâm nhập tế bào. Trong tế bào, enzyme reverse transcriptase tạo một dây DNA phụ từ sợi RNA đã bị phân giải của retrovirus. Sợi DNA này được gọi là cDNA. Sau đó, cADN làm khuôn để tổng hợp ra sợi ADN thứ hai bổ sung với nó (cADN→ADN) để xâm nhập vào nhân tế bào chủ bằng enzyme integrase. Sợi cADN này cũng có thể làm khuôn để tổng hợp trở lại bộ gene của virus (cADN→ARN). Ribosome được dùng để sao chép mRNA của virus thành chuỗi axit amin để tạo protein trong màng lưới nội bào. Bước này cũng sẽ tạo enzyme và vỏ protein của virus. RNA của virus sẽ được hình thành trong nhân. Những mảnh này sau đó sẽ tập hợp lại và ra khỏi màng tế bào, trở thành một retrovirus mới. Cứ thế, retrovirus này lại tấn công vào các tế bào chủ khác.
Việc phát hiện ra retrovirus có mang nhiều ý nghĩa vì tính chất lịch sử, kỹ thuật, và quan trọng nhất, cách sử dụng retrovirus để xử lý những vấn đề sức khỏe cộng đồng do chính nó gây ra.
Về tính chất lịch sử, qua nhiều nghiên cứu ban đầu về sinh học tế bào và phân tử, con người đã tìm ra DNA, RNA và protein. Sau đó, họ nhận thấy một khuynh hướng chung: DNA à RNA à protein. Lúc đó, người ta đã tin đây là quy luật, không thể đảo ngược.
Sau đó, retrovirus được phát hiện. Retrovirus đặc biệt ở chỗ nó có thể tạo DNA từ RNA, ngược lại so với những gì người thời ấy luôn tin tưởng. Họ đã nghi ngờ, nhưng những nghiên cứu sau đó đã chứng minh đây là thật.
Con người biết quá ít về retrovirus. Retrovirus càng trở nên nguy hiểm khi họ nhận ra rằng: nếu một cơ thể bị nhiễm virus này, retrovirus sẽ dùng bộ máy tế bào của vật chủ, khiến tế bào phân ra và tạo nên vô số bản sao virus. Virus này lại tiếp tục tấn công vào các tế bào khác của vật chủ, có thể dẫn đến tình trạng ung thư.
Để chống lại HIV, chúng ta thường dùng các phương pháp nhằm chống lại sự tạo thành DNA của retrovirus.
Retrovirus được nhắc đến nhiều trong công nghệ sinh học vì chúng ta muốn khai thác khả năng của nó. Những bệnh về gene ngày một nhiều, và nguyên nhân chính của chúng thường là một lỗi rất nhỏ trong DNA. Lỗi này có thể ngăn lại quá trình tổng hợp của một protein quan trọng, hoặc khiến protein hoạt động sai cách. Nếu chúng ta có thể thêm lại DNA đúng, khiến protein hoạt động bình thường, bệnh có thể được chữa khỏi. Điều này đòi hỏi sự khéo léo vì virus có thể dễ dàng vượt qua các sự bảo hộ mà ta đã dựng lên. Nhưng phương pháp này cũng có nhiều khả năng thành công, và rất nhiều người đang coi retrovirus như một phép chữa bệnh. Chúng ta biết được về retrovirus càng nhiều thì lại càng có thể chữa trị cho những người bị nhiễm chúng tốt hơn.
http://www.madsci.org/posts/archives/2001-12/1008703762.Vi.r.html
Retrovirus là một từ để gọi các loại virus mà vật chất di truyền của chúng là phân tử RNA. Khi chúng xâm nhập vào cơ thể thì có khả năng thực hiện quá trình phiên mã ngược nhờ enzim đặc biệt là enzyme phiên mã ngược - Reverse Transciptase. DNA sau đó là kết hợp vào gen của vật chủ bởi một enzyme tích hợp. Vi rút này sau đó sao chép như là một phần của DNA của tế bào chủ. Retrovirus là vi rút bao bọc es thuộc về gia họ virus "Retroviridae". Một biến thể đặc biệt của retrovirus retrovirus nội sinh được tích hợp vào bộ gen của vật chủ và di truyền qua nhiều thế hệ.
Retroviridae
Подсемейства Группа по БалтиморуVI: оцРНК-ОТ-вирусы
Ретрови́русы[2] (лат. Retroviridae, от лат. retro — обратный) — семейство РНК-содержащих вирусов, заражающих преимущественно позвоночных. Наиболее известный и активно изучаемый представитель — вирус иммунодефицита человека.
После инфицирования клетки ретровирусом в цитоплазме начинается синтез вирусного ДНК-генома с использованием вирионной РНК в качестве матрицы. Все ретровирусы используют для репликации своего генома механизм обратной транскрипции: вирусный фермент обратная транскриптаза (или ревертаза) синтезирует одну нить ДНК на матрице вирусной РНК, а затем уже на матрице синтезированной нити ДНК достраивает вторую, комплементарную ей нить. Образуется двунитевая молекула ДНК, которая интегрируется в хромосомную ДНК клетки во время клеточного деления, когда нет ядерной оболочки, (исключением является ВИЧ, ДНК которого активно проникает в ядро) и далее служит матрицей для синтеза молекул вирусных РНК. Эти РНК выходят из клеточного ядра и в цитоплазме клетки упаковываются в вирусные капсиды, способные инфицировать новые клетки.
По одной из гипотез, ретровирусы могли произойти от ретротранспозонов — подвижных участков генома эукариот[3].
Вирионы сферической формы размером 80—100 нм, покрыты внешней липопротеиновой оболочкой, имеющей ворсинки длиной 8—10 нм. Внутри икосаэдрального капсида находится спиральный рибонуклеопротеид. Наружная оболочка, капсидная мембрана и нуклеоид на разрезе вириона расположены концентрически. Чувствительны к эфиру, термолабильны, относительно резистентны к УФ-лучам. Характерной чертой семейства является наличие в составе вириона РНК-зависимой ДНК-полимеразы, иначе называемой обратной транскриптазой. Это и послужило основой для названия семейства (от лат. retro — обратный). Вирионы имеют 6 структурных белков, из них 4 внутренних (капсидных) негликолизированных и 2 гликопротеина оболочки.
Основными структурными генами кодирующими трансляцию белков, из которых в последующем строится вирус, являются gag (group-specific antigens), pol (polymerase), env (envelope). К регуляторным генам относятся: tat (трансактиватор всех вирусных белков), rev (регулятор экспрессии вирионных белков), vif (вирионный инфекционный фактор), vpr (функции остаются неясными), nef (негативный фактор экспрессии), vpx (функции неизвестны).
Капсидные белки несут группоспецифические межвидовые антигены и являются основой для разделения вирусов на роды и подроды. Гликопротеиды являются типоспецифическими антигенами, участвуют в реакции нейтрализации. Геном ретровирусов представлен однонитчатой РНК с молекулярной массой 7 мегадальтон и состоит из двух копий, каждая из которых является полноценным геномом и содержит одинаковую генетическую информацию, однако неизвестно, обе ли они функциональны. Нуклеиновая кислота онковирусов имеет гомологию с клеточной ДНК своего вида хозяина. Вирионная РНК неинфекционна. Вирусная РНК транскрибируется в ковалентно связанную двунитчатую ДНК, которая интегрируется с клеточной ДНК в виде ДНК-провируса. Провирус, экстрагированный из клетки, обладает инфекционностью. Многие вирусы этого семейства вызывают неопластические процессы, главным образом лейкемии и саркомы ряда видов животных. Нормальные клетки некоторых видов животных содержат интегрированные копии соответствующих видов онковирусов. Они могут никак не проявляться или активируются некоторыми физическими и химическими факторами, а возможно, и при инфекции другими онковирусами. Часто встречаются дефектные вирусы, размножающиеся с помощью вируса-помощника. Передаются вертикально и горизонтально.
Ретровирус раздваивает функции своего генетического материала: инфекционную функцию, то есть функцию самораспространения, выполняет вирусная РНК, а функцию экспрессии вирусных генов и синтеза молекул РНК, которые затем перенесут генетическую информацию в другие клетки, выполняет вирусная ДНК.
Попадая внутрь клетки в ходе вирусной инфекции, ретровирусная РНК служит матрицей для синтеза ДНК путём хорошо теперь известного процесса обратной транскрипции. Эта ДНК встраивается в геномную ДНК и с этого момента становится неотъемлемой частью генома клетки. А вирус становится провирусом. Провирус для животной клетки это то же самое, что профаг для бактериальной. Идеи лизогении, по-видимому, и привели Говарда Темина, который вместе с Дэвидом Балтимором открыл обратную транскрипцию, к идее провирусного состояния ретровирусов. По информационному содержанию ДНК-вариант генома ретровируса отличается от РНК-варианта только тем, что ДНК содержит не короткие концевые повторы, а длинные концевые повторы, LTR.
Находясь в составе геномной ДНК, вирусные гены транскрибируются под контролем LTR.
LTR, long terminal repeats. Последовательности LTR включают в себя последовательности STR. Возникновение LTR очень важно для экспрессии вирусных генов. Они содержат вирусные регуляторные транскрипционные элементы: промотор, энхансер, и другие. Например, некоторые вирусы содержат элементы, определяющие зависимость вирусной транскрипции от наличия определённых гормонов. LTR и являются теми регуляторными сигналами, которые вирус использует для эксплуатации клеточной транскрипционной машины в своих целях.
Продуктом транскрипции является полноразмерная вирусная РНК. Она должна транслироваться. И здесь вирусу необходимо решить такую проблему: нужно синтезировать много белков, а РНК одна. И в клетках эукариот РНК моноцистронны, то есть предназначены для синтеза только одного белка. Синтез белка в большинстве случаев начинается с ближайшего к кэп-сайту инициирующего кодона[4].
Если просмотреть открытую рамку считывания от этого ближайшего инициирующего кодона, то мы увидим, что если бы вирус пользовался традиционными способами экспрессии, то он смог бы синтезировать только полипептид GAG. А дальше идёт стоп- кодон. Как быть с POL и ENV? Кроме того, эти полипептиды очень длинны, а в вирусе содержатся гораздо более короткие. Проблема решается несколькими способами.
Во-первых, с помощью сплайсинга эта одна РНК превращается в нашем упрощённом варианте ещё в одну, более короткую. При этом последовательности, кодирующие ENV полипептид, оказываются рядом с инициирующим кодоном, ближайшим к кэп-сайту, и начинают транслироваться.
Во-вторых, разными для разных ретровирусов способами они ухитряются обойти стоп-кодон после открытой рамки считывания GAG и синтезировать сплавленный полипетид GAG-POL, который содержит последовательности обеих групп белков.
В-третьих, полученные длинные полипептиды подвергаются процессингу и разрезаются на множество белков, которые и функционируют либо в роли регуляторных, как, например, обратная транскриптаза, либо в роли структурных, как, например, белки оболочки зрелых вирусов.
Иными словами, ретровирусы используют гибкую тоталитарную систему для весьма тонкой регуляции синтеза большого разнообразия белков под контролем одного промотора.
Ретровирусы могут применяться в качестве векторов, например, в генотерапии. Механизм проникновения внутрь клеток при помощи слияния мембран. У них есть ряд преимуществ: широкий круг хозяев, стабильность в интегрированном в геном хозяина состоянии. Недостатки: сложно добиться высокого титра, заражает только делящиеся клетки.[источник не указан 940 дней]
На март 2018 года в семейство включают 2 подсемейства с 6 и 5 родами соответственно[5]:
Ретрови́русы (лат. Retroviridae, от лат. retro — обратный) — семейство РНК-содержащих вирусов, заражающих преимущественно позвоночных. Наиболее известный и активно изучаемый представитель — вирус иммунодефицита человека.
После инфицирования клетки ретровирусом в цитоплазме начинается синтез вирусного ДНК-генома с использованием вирионной РНК в качестве матрицы. Все ретровирусы используют для репликации своего генома механизм обратной транскрипции: вирусный фермент обратная транскриптаза (или ревертаза) синтезирует одну нить ДНК на матрице вирусной РНК, а затем уже на матрице синтезированной нити ДНК достраивает вторую, комплементарную ей нить. Образуется двунитевая молекула ДНК, которая интегрируется в хромосомную ДНК клетки во время клеточного деления, когда нет ядерной оболочки, (исключением является ВИЧ, ДНК которого активно проникает в ядро) и далее служит матрицей для синтеза молекул вирусных РНК. Эти РНК выходят из клеточного ядра и в цитоплазме клетки упаковываются в вирусные капсиды, способные инфицировать новые клетки.
По одной из гипотез, ретровирусы могли произойти от ретротранспозонов — подвижных участков генома эукариот.
逆转录病毒科(学名:Retroviridae)又稱「反轉錄病毒科」,分爲雙鏈DNA逆轉錄病毒和正鏈RNA逆轉錄病毒。它们增殖時都有將遺傳物質RNA逆轉錄爲DNA的過程(前者逆轉錄反鏈RNA,後者逆轉錄正鏈RNA),多具有逆转录酶。本文主要介紹單鏈RNA逆轉錄病毒。
逆轉錄病毒科下有Alpharetrovirus,Betaretrovirus,Gammaretrovirus,Deltaretrovirus,Epsilonretrovirus,Lentivirus及Spumavirus共七屬。於1908年於丹麥証明、1911年於美國進行人工實驗証實此病毒科之存在。在感染受害细胞时,逆转录病毒首先将其RNA逆转录为DNA(因此被称为逆转录病毒),然后将这段逆转录的基因插入细胞基因中,由细胞的转录机构转换为病毒的蛋白质和RNA。逆转录病毒一般在其外壳外还有一层脂肪。
逆轉錄病毒大小約為100nm(70~110nm,有時至200nm),外面為一層具有表面突起的脂蛋白套膜。套膜內有一二十面體的核蛋白衣(nucleo capsid),核蛋白衣內有一螺旋結構的核糖核蛋白(ribonucleoprotein),當以負染色電子顯微鏡觀察時,許多反轉錄病毒,尤其是引起動物白血病及腫瘤者,都是多形性病毒顆粒,對滲透壓的改變非常敏感,當樣品處理時常會使病毒破碎。此科的亞科致腫瘤RNA病毒(Oncovirinae),可於許多細胞內繁殖達到很高的力價而不改變細胞的性狀。尤其是在薄切片時經常可看到直徑為85~110nm的C顆粒,大部分哺乳動物都有相關的C顆粒。
在薄切片時,C顆粒因其核心殼(core shell)及其染色較深的核蛋白衣呈同心狀排列而類似靶心。B型致腫瘤RNA病毒如鼠乳房瘤病毒(mouse mammary tumor virus),因其病毒核心位置較偏而與C型不同,其與外殼間有一電子透明的空間。B型致腫瘤RNA病毒於負染色時較C型穩定而不易破碎。A型顆粒為圓形或油炸甜圈餅形中心為電子透明,直徑約75nm常存在於細胞質高基氏體附近或內質網之空隙內(cisternae of the endoplasmic reticulum)。
逆转录病毒至少含下列三种基因:
比较著名的逆转录病毒有艾滋病病毒等。
大多数逆转录病毒造成的疾病都是比较慢性的,有些病毒虽然不直接导致疾病,但可以导致癌症,有些病毒可以在其基因插入细胞基因内而不导致疾病。在人类进化的过程中有许多逆转录病毒将它们的基因插入人的基因内并成为人的基因的一部分。[來源請求]
由此可以将逆转录病毒分为三类:
由于逆转录过程比较不稳定,因此逆转录病毒的变异过程比较快。这使研究针对逆转录病毒的免疫抗体的过程非常困难。
大多数针对逆转录病毒的治疗方法(比如目前拥有的针对艾滋病的药物)主要是针对病毒的逆转录过程。但由于病毒迅速的变异过程,病毒往往会很快就产生对药物的阻抗。
反轉錄病毒科(Retroviridae)
在台灣,家禽白血病病毒(Avian leukosis viruses),貓白血病病毒(Feline leukemia virus 1996),牛白血病病毒(Bovine leukemia virus),羊關節炎腦炎病毒(caprine arthritis-encephalitis virus 1984),馬傳染性貧血症病毒(Equine infectious anemia virus 1992),均為台灣重要的疾病,感染情形非常普遍。
逆转录病毒科(学名:Retroviridae)又稱「反轉錄病毒科」,分爲雙鏈DNA逆轉錄病毒和正鏈RNA逆轉錄病毒。它们增殖時都有將遺傳物質RNA逆轉錄爲DNA的過程(前者逆轉錄反鏈RNA,後者逆轉錄正鏈RNA),多具有逆转录酶。本文主要介紹單鏈RNA逆轉錄病毒。
スプマウイルス亜科 spumavirinae
レンチウイルス亜科 Lentivirinae
オンコウイルス亜科 oncvirinae
オルソレトロウイルス亜科 Orthoretrovirinae
レトロウイルス科(retrovirus)とは、RNAウイルス類の中で逆転写酵素を持つ種類の総称。ウイルス核酸は、一本鎖RNAの+鎖である。単にレトロウイルスと呼ばれることも多い。
レトロウイルス科のウイルスは直径約100nmの球状の形をしており、脂質のエンベロープに覆われている。ウイルス粒子のコアには逆転写酵素と通常2組の同一なRNAゲノムを持つ。共通の遺伝子としては、プロモーター活性のある2つのLTR(long terminal repeat)と呼ばれる塩基配列の繰り返し領域に挟まれて、gag(構造タンパク質をコード)、pro(プロテアーゼをコード)、pol(逆転写酵素などをコード)、env(エンベロープのタンパク質をコード)を持っている。
ウイルスのエンベロープが細胞膜の受容体と結合することで、細胞内にRNAと逆転写酵素が侵入する。
その後逆転写酵素が作用し、ウイルスRNAを鋳型にマイナス鎖DNAを合成する。そして合成されたマイナス鎖DNAを鋳型にプラス鎖DNAが合成され、一本鎖RNAが二本鎖DNAに変換される。
その後二本鎖DNAは宿主細胞のDNAに組み込まれ、プロウイルスと呼ばれる状態になる。プロウイルスは恒常的に発現している状態となっており、ウイルスRNAやメッセンジャーRNAが次々と合成されていく。メッセンジャーRNAはウイルス蛋白を合成させ、完成したウイルスは宿主細胞から発芽していく。
このレトロウイルスは近現代において「天然のナノマシン」として、様々な遺伝子治療や遺伝子研究に利用されている。
レトロウイルス科は以下の亜科に分けられる。
細胞を培養すると泡状 (spuma) の細胞変性効果が見られることから名づけられたウイルス。病原性は不明。
発病までの進行がゆっくり (lente) であることを意味するウイルス。
腫瘍 (onc) を引き起こすウイルス。
ガンマレトロウイルス属にはマウス白血病ウイルス、猫白血病ウイルス、細網内皮症ウイルスがある。 XMRV (Xenotropic murine leukemia virus-related virus)は前立腺がん、慢性疲労症候群との関連が示唆されたが、後に人の疾病との関連は否定された[1]。
B型肝炎ウイルス (HBV: hepatitis B virus) はDNA→RNA→DNAと逆転写を経て複製するためレトロイドともよばれ、HIVの治療薬でもある逆転写酵素阻害薬の一部が効果を示す(エンテカビル、アデフォビル、ラミブジンは日本でも保険適応がある)。
レトロウイルス科(retrovirus)とは、RNAウイルス類の中で逆転写酵素を持つ種類の総称。ウイルス核酸は、一本鎖RNAの+鎖である。単にレトロウイルスと呼ばれることも多い。
레트로바이러스(영어: Retrovirus, 문화어: 레트롤비루스)는 유전자로써 RNA를 소유하고 있는 RNA 바이러스의 한 유형이다. 숙주를 감염시키면 자신의 RNA를 역전사 효소를 사용해 상보적 DNA로 역전사하고 이 상보적 DNA를 인테그레이스를 사용해 DNA를 합성하는 역전사의 기작을 보이기 때문에 분자생물학의 중심원리를 따르지 않는 가장 대표적인 존재로 분류된다. 레트로바이러스는 유전 물질인 핵산이 뉴클레오캡시드 형태로 되어있다.
레트로바이러스의 감염과 증식은 다음과 같은 과정을 통해 일어난다.
대개의 레트로바이러스는 숙주 세포를 죽이지 않고 이용만 하기 때문에, 면역 체계가 바이러스만 인식해서 공격하는 것만으로는 레트로바이러스에 이미 감염된 세포를 없애지 못하므로 거의 대부분 만성 감염으로 진행하게 된다. 따라서 혈액 중의 바이러스 숫자는 극히 적지만 감염 자체는 계속 진행되는 현상이 나타난다. 따라서 치료를 위해서는 레트로바이러스의 감염 자체를 막는 방법 또는 레트로바이러스에 감염된 세포를 파괴하는 방법이 필요하다. 인간에 감염되는 대표적인 레트로바이러스로는 HIV(Human Immunodeficiency Virus) 가 있다.
종양 성장을 일으키는 레트로바이러스에는 라우스육종바이러스(Rous sarcoma virus) 및 생쥐유방종양바이러스(Mouse mammary tumor virus)가 포함된다. 암은 프로바이러스 DNA에 통합된 원 종양 유전자 또는 세포 원 종양 유전자의 파괴에 의해 유발된다. 라우스육종바이러스에는 종양 형성을 유발하는 src 유전자가 포함되어 있다. 나중에 세포에서 유사한 유전자가 세포 신호 전달에 관여한다는 것이 밝혀졌다. 비형질 전환 바이러스는 DNA를 무작위로 원형 종양 유전자에 삽입하여 세포 주기를 조절하는 단백질의 발현을 방해 할 수 있다. 프로바이러스 DNA의 촉진유전자는 또한 조절 유전자의 과발현을 야기 할 수 있다.
레트로바이러스(영어: Retrovirus, 문화어: 레트롤비루스)는 유전자로써 RNA를 소유하고 있는 RNA 바이러스의 한 유형이다. 숙주를 감염시키면 자신의 RNA를 역전사 효소를 사용해 상보적 DNA로 역전사하고 이 상보적 DNA를 인테그레이스를 사용해 DNA를 합성하는 역전사의 기작을 보이기 때문에 분자생물학의 중심원리를 따르지 않는 가장 대표적인 존재로 분류된다. 레트로바이러스는 유전 물질인 핵산이 뉴클레오캡시드 형태로 되어있다.











_tr%C3%A4nger_in_i_T-hj%C3%A4lparcellerna_och_f%C3%B6r%C3%B6kar_sig.JPG)

{kind=link}
{kind=link}