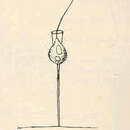
The choanoflagellates are a group of free-living unicellular and colonial flagellate eukaryotes considered to be the closest living relatives of the animals. Choanoflagellates are collared flagellates, having a funnel shaped collar of interconnected microvilli at the base of a flagellum. Choanoflagellates are capable of both asexual and sexual reproduction.[8] They have a distinctive cell morphology characterized by an ovoid or spherical cell body 3–10 µm in diameter with a single apical flagellum surrounded by a collar of 30–40 microvilli (see figure). Movement of the flagellum creates water currents that can propel free-swimming choanoflagellates through the water column and trap bacteria and detritus against the collar of microvilli, where these foodstuffs are engulfed. This feeding provides a critical link within the global carbon cycle, linking trophic levels. In addition to their critical ecological roles, choanoflagellates are of particular interest to evolutionary biologists studying the origins of multicellularity in animals. As the closest living relatives of animals, choanoflagellates serve as a useful model for reconstructions of the last unicellular ancestor of animals.
Choanoflagellate is a hybrid word from Greek χοάνηcode: ell promoted to code: el khoánēcode: ell promoted to code: el meaning "funnel" (due to the shape of the collar) and the Latin word flagellum.
Each choanoflagellate has a single flagellum, surrounded by a ring of actin-filled protrusions called microvilli, forming a cylindrical or conical collar (choanoscode: ell promoted to code: el in Greek). Movement of the flagellum draws water through the collar, and bacteria and detritus are captured by the microvilli and ingested.[9] Water currents generated by the flagellum also push free-swimming cells along, as in animal sperm. In contrast, most other flagellates are pulled by their flagella.
In addition to the single apical flagellum surrounded by actin-filled microvilli that characterizes choanoflagellates, the internal organization of organelles in the cytoplasm is constant.[10] A flagellar basal body sits at the base of the apical flagellum, and a second, non-flagellar basal body rests at a right angle to the flagellar base. The nucleus occupies an apical-to-central position in the cell, and food vacuoles are positioned in the basal region of the cytoplasm.[10][11] Additionally, the cell body of many choanoflagellates is surrounded by a distinguishing extracellular matrix or periplast. These cell coverings vary greatly in structure and composition and are used by taxonomists for classification purposes. Many choanoflagellates build complex basket-shaped "houses", called lorica, from several silica strips cemented together.[10] The functional significance of the periplast is unknown, but in sessile organisms, it is thought to aid attachment to the substrate. In planktonic organisms, there is speculation that the periplast increases drag, thereby counteracting the force generated by the flagellum and increasing feeding efficiency.[12]
Choanoflagellates are either free-swimming in the water column or sessile, adhering to the substrate directly or through either the periplast or a thin pedicel.[13] Although choanoflagellates are thought to be strictly free-living and heterotrophic, a number of choanoflagellate relatives, such as members of Ichthyosporea or Mesomycetozoa, follow a parasitic or pathogenic lifestyle.[14] The life histories of choanoflagellates are poorly understood. Many species are thought to be solitary; however, coloniality seems to have arisen independently several times within the group and colonial species retain a solitary stage.[13]
.png)
Over 125 extant species of choanoflagellates[9] are known, distributed globally in marine, brackish and freshwater environments from the Arctic to the tropics, occupying both pelagic and benthic zones. Although most sampling of choanoflagellates has occurred between 0 and 25 m (0 and 82 ft), they have been recovered from as deep as 300 m (980 ft) in open water[15] and 100 m (330 ft) under Antarctic ice sheets.[16] Many species are hypothesized to be cosmopolitan on a global scale [e.g., Diaphanoeca grandis has been reported from North America, Europe and Australia (OBIS)], while other species are reported to have restricted regional distributions.[17] Co-distributed choanoflagellate species can occupy quite different microenvironments, but in general, the factors that influence the distribution and dispersion of choanoflagellates remain to be elucidated.
A number of species, such as those in the genus Proterospongia, form simple colonies,[9] planktonic clumps that resemble a miniature cluster of grapes in which each cell in the colony is flagellated or clusters of cells on a single stalk.[10][18] In October 2019, scientists found a new band behaviour of choanoflagellates: they apparently can coordinate to respond to light.[19]
The choanoflagellates feed on bacteria and link otherwise inaccessible forms of carbon to organisms higher in the trophic chain.[20] Even today, they are important in the carbon cycle and microbial food web.[9] There is some evidence that choanoflagellates feast on viruses as well.[21]

Choanoflagellates grow vegetatively, with multiple species undergoing longitudinal fission;[11] however, the reproductive life cycle of choanoflagellates remains to be elucidated. A paper released in August 2017 showed that environmental changes, including the presence of certain bacteria, trigger the swarming and subsequent sexual reproduction of choanoflagellates.[8] The ploidy level is unknown;[23] however, the discovery of both retrotransposons and key genes involved in meiosis[24] previously suggested that they used sexual reproduction as part of their life cycle. Some choanoflagellates can undergo encystment, which involves the retraction of the flagellum and collar and encasement in an electron dense fibrillar wall. On transfer to fresh media, excystment occurs; though it remains to be directly observed.[25]
Evidence for sexual reproduction has been reported in the choanoflagellate species Salpingoeca rosetta.[26][27] Evidence has also been reported for the presence of conserved meiotic genes in the choanoflagellates Monosiga brevicollis and Monosiga ovata.[28]
The Acanthoecid choanoflagellates produce an extracellular basket structure known as a lorica. The lorica is composed of individual costal strips, made of a silica-protein biocomposite. Each costal strip is formed within the choanoflagellate cell and is then secreted to the cell surface. In nudiform choanoflagellates, lorica assembly takes place using a number of tentacles once sufficient costal strips have been produced to comprise a full lorica. In tectiform choanoflagellates, costal strips are accumulated in a set arrangement below the collar. During cell division, the new cell takes these costal strips as part of cytokinesis and assembles its own lorica using only these previously produced strips.[29]
Choanoflagellate biosilicification requires the concentration of silicic acid within the cell. This is carried out by silicon transporter (SiT) proteins. Analysis of choanoflagellate SiTs shows that they are similar to the SiT-type silicon transporters of diatoms and other silica-forming stramenopiles. The SiT gene family shows little or no homology to any other genes, even to genes in non-siliceous choanoflagellates or stramenopiles. This suggests that the SiT gene family evolved via a lateral gene transfer event between Acanthoecids and Stramenopiles. This is a remarkable case of horizontal gene transfer between two distantly related eukaryotic groups, and has provided clues to the biochemistry and silicon-protein interactions of the unique SiT gene family.[30]
Dujardin, a French biologist interested in protozoan evolution, recorded the morphological similarities of choanoflagellates and sponge choanocytes and proposed the possibility of a close relationship as early as 1841.[12] Over the past decade, this hypothesized relationship between choanoflagellates and animals has been upheld by independent analyses of multiple unlinked sequences: 18S rDNA, nuclear protein-coding genes, and mitochondrial genomes (Steenkamp, et al., 2006; Burger, et al., 2003;[14] Wainright, et al., 1993). Importantly, comparisons of mitochondrial genome sequences from a choanoflagellate and three sponges confirm the placement of choanoflagellates as an outgroup to Metazoa and negate the possibility that choanoflagellates evolved from metazoans (Lavrov, et al., 2005). Finally, a 2001 study of genes expressed in choanoflagellates have revealed that choanoflagellates synthesize homologues of metazoan cell signaling and adhesion genes.[31] Genome sequencing shows that, among living organisms, the choanoflagellates are most closely related to animals.[9] Because choanoflagellates and metazoans are closely related, comparisons between the two groups promise to provide insights into the biology of their last common ancestor and the earliest events in metazoan evolution. The choanocytes (also known as "collared cells") of sponges (considered among the most basal metazoa) have the same basic structure as choanoflagellates. Collared cells are found in other animal groups, such as ribbon worms,[32] suggesting this was the morphology of their last common ancestor. The last common ancestor of animals and choanoflagellates was unicellular, perhaps forming simple colonies; in contrast, the last common ancestor of all eumetazoan animals was a multicellular organism, with differentiated tissues, a definite "body plan", and embryonic development (including gastrulation).[9] The timing of the splitting of these lineages is difficult to constrain, but was probably in the late Precambrian,>600 million years ago.[9]
External relationships of Choanoflagellatea.[33]
Opisthokonta Holomycota Holozoa Filozoa ChoanozoaChoanoflagellatea
The choanoflagellates were included in Chrysophyceae until Hibberd, 1975.[34] Recent molecular phylogenetic reconstruction of the internal relationships of choanoflagellates allows the polarization of character evolution within the clade. Large fragments of the nuclear SSU and LSU ribosomal RNA, alpha tubulin, and heat-shock protein 90 coding genes were used to resolve the internal relationships and character polarity within choanoflagellates.[18] Each of the four genes showed similar results independently and analysis of the combined data set (concatenated) along with sequences from other closely related species (animals and fungi) demonstrate that choanoflagellates are strongly supported as monophyletic and confirm their position as the closest known unicellular living relative of animals.
Previously, Choanoflagellida was divided into these three families based on the composition and structure of their periplast: Codonosigidae, Salpingoecidae and Acanthoecidae. Members of the family Codonosigidae appear to lack a periplast when examined by light microscopy, but may have a fine outer coat visible only by electron microscopy. The family Salpingoecidae consists of species whose cells are encased in a firm theca that is visible by both light and electron microscopy. The theca is a secreted covering predominately composed of cellulose or other polysaccharides.[35] These divisions are now known to be paraphyletic, with convergent evolution of these forms widespread. The third family of choanoflagellates, the Acanthoecidae, has been supported as a monophyletic group. This clade possess a synapomorphy of the cells being found within a basket-like lorica, providing the alternative name of "Loricate Choanoflagellates". The Acanthoecid lorica is composed of a series of siliceous costal strips arranged into a species-specific lorica pattern."[10][12]
The choanoflagellate tree based on molecular phylogenetics divides into three well supported clades.[18] Clade 1 and Clade 2 each consist of a combination of species traditionally attributed to the Codonosigidae and Salpingoecidae, while Clade 3 comprises species from the group taxonomically classified as Acanthoecidae.[18] The mapping of character traits on to this phylogeny indicates that the last common ancestor of choanoflagellates was a marine organism with a differentiated life cycle with sedentary and motile stages.[18]
Choanoflagellates;[4]
The genome of Monosiga brevicollis, with 41.6 million base pairs,[9] is similar in size to filamentous fungi and other free-living unicellular eukaryotes, but far smaller than that of typical animals.[9] In 2010, a phylogenomic study revealed that several algal genes are present in the genome of Monosiga brevicollis. This could be due to the fact that, in early evolutionary history, choanoflagellates consumed algae as food through phagocytosis.[36] Carr et al. (2010)[28] screened the M. brevicollis genome for known eukaryotic meiosis genes. Of 19 known eukaryotic meiotic genes tested (including 8 that function in no other process than meiosis), 18 were identified in M. brevicollis. The presence of meiotic genes, including meiosis specific genes, indicates that meiosis, and by implication, sex is present within the choanoflagellates.
The genome of Salpingoeca rosetta is 55 megabases in size.[37] Homologs of cell adhesion, neuropeptide and glycosphingolipid metabolism genes are present in the genome. S. rosetta has a sexual life cycle and transitions between haploid and diploid stages.[27] In response to nutrient limitation, haploid cultures of S. rosetta become diploid. This ploidy shift coincides with mating during which small, flagellated cells fuse with larger flagellated cells. There is also evidence of historical mating and recombination in S. rosetta.
S. rosetta is induced to undergo sexual reproduction by the marine bacterium Vibrio fischeri.[26] A single V. fischeri protein, EroS fully recapitulates the aphrodisiac-like activity of live V. fisheri.
The single-cell amplified genomes of four uncultured marine choanoflagellates, tentatively called UC1–UC4, were sequenced in 2019. The genomes of UC1 and UC4 are relatively complete.[38]
An EST dataset from Monosiga ovata was published in 2006.[39] The major finding of this transcriptome was the choanoflagellate Hoglet domain and shed light on the role of domain shuffling in the evolution of the Hedgehog signaling pathway. M. ovata has at least four eukaryotic meiotic genes.[28]
The transcriptiome of Stephanoeca diplocostata was published in 2013. This first transcriptome of a loricate choanoflagellate[30] led to the discovery of choanoflagellate silicon transporters. Subsequently, similar genes were identified in a second loricate species, Diaphanoeca grandis. Analysis of these genes found that the choanoflagellate SITs show homology to the SIT-type silicon transporters of diatoms and have evolved through horizontal gene transfer.
An additional 19 transcriptomes were published in 2018. A large number of gene families previously thought to be animal-only were found.[40]

Monosiga brevicollis under PCM

Salpingoeca under PCM

Salpingoeca sp. section under TEM

Desmarella moniliformis colony under PCM

Codosiga colony under light microscopy

Sphaeroeca colony (approx. 230 individuals) under light microscopy.
The choanoflagellates are a group of free-living unicellular and colonial flagellate eukaryotes considered to be the closest living relatives of the animals. Choanoflagellates are collared flagellates, having a funnel shaped collar of interconnected microvilli at the base of a flagellum. Choanoflagellates are capable of both asexual and sexual reproduction. They have a distinctive cell morphology characterized by an ovoid or spherical cell body 3–10 µm in diameter with a single apical flagellum surrounded by a collar of 30–40 microvilli (see figure). Movement of the flagellum creates water currents that can propel free-swimming choanoflagellates through the water column and trap bacteria and detritus against the collar of microvilli, where these foodstuffs are engulfed. This feeding provides a critical link within the global carbon cycle, linking trophic levels. In addition to their critical ecological roles, choanoflagellates are of particular interest to evolutionary biologists studying the origins of multicellularity in animals. As the closest living relatives of animals, choanoflagellates serve as a useful model for reconstructions of the last unicellular ancestor of animals.
