nomes no trilho de navegação
BAKULOVİRUSLAR (Baculoviridae) –onurğasız heyvanları, başlıca olaraq buğumayaqlıları zədələyən viruslar fəsiləsi.
2 cinsi, təqribən 100-ədək növü var. Çubuqşəkilli virus hissəcikləri (virionlar) təqribən 10 müxtəlif zülalın daxil olduğu lipoprotein qılafından və özək, yaxud nukleokapsiddən (uz. 200–450 nm, diametri 30–100 nm) ibarətdir. Sonuncu həlqəvi ikisaplı DNT molekulu (virusun genomu) və bir zülal növü ilə təmsil olunmuşdur. Genom, əksəriyyəti virionların tərkibinə daxil olmayan, lakin virusların çoxalması üçün zəruri sayılan 100–200-ədək zülalı kodlaşdırır. Virus DNT-sinin sintezi və nukleokapsidlərin əmələ gəlməsi yoluxmuş hüceyrənin nüvəsində baş verir. Bu zaman nukleokapsidlərin çoxu 15-dən artıq zülalın iştirakı ilə formalaşan spesifik törəmələr – okklyuzilər şəklində nüvə də qalır. Qalan hissədən hüceyrədən xaricə çıxan virus hissəcikləri əmələ gəlir. Virus hissəcikləri plazmatik membranların fraqmentlərini tutaraq onları öz qılaflarına birləşdirir. Bakulovirusdan gen terapiyasında genetik informasiya daşıyıcıları kimi istifadə olunur.
"Bakuloviruslar". Azərbaycan Milli Ensiklopediyası: [25 cilddə]. III cild (25 000 nüs.). Bakı: “Azərbaycan Milli Ensiklopediyası” Elmi Mərkəzi. baş red. M. K. Kərimov. 2011. səh. 94. ISBN 978-9952-441-07-9.
BAKULOVİRUSLAR (Baculoviridae) –onurğasız heyvanları, başlıca olaraq buğumayaqlıları zədələyən viruslar fəsiləsi.
2 cinsi, təqribən 100-ədək növü var. Çubuqşəkilli virus hissəcikləri (virionlar) təqribən 10 müxtəlif zülalın daxil olduğu lipoprotein qılafından və özək, yaxud nukleokapsiddən (uz. 200–450 nm, diametri 30–100 nm) ibarətdir. Sonuncu həlqəvi ikisaplı DNT molekulu (virusun genomu) və bir zülal növü ilə təmsil olunmuşdur. Genom, əksəriyyəti virionların tərkibinə daxil olmayan, lakin virusların çoxalması üçün zəruri sayılan 100–200-ədək zülalı kodlaşdırır. Virus DNT-sinin sintezi və nukleokapsidlərin əmələ gəlməsi yoluxmuş hüceyrənin nüvəsində baş verir. Bu zaman nukleokapsidlərin çoxu 15-dən artıq zülalın iştirakı ilə formalaşan spesifik törəmələr – okklyuzilər şəklində nüvə də qalır. Qalan hissədən hüceyrədən xaricə çıxan virus hissəcikləri əmələ gəlir. Virus hissəcikləri plazmatik membranların fraqmentlərini tutaraq onları öz qılaflarına birləşdirir. Bakulovirusdan gen terapiyasında genetik informasiya daşıyıcıları kimi istifadə olunur.
Baculoviridae és una família de virus que es pot dividir en dos gèneres: nucleopolyhedrovirus (NPV) i granulovirus (GV). Mentre que els GV contenen només dues càpsides nucleiques per embolcall víric, els NPV contenen sia una simple càpside sia una múltiple (SNPV o MNPV) per embolcall.[1]
Els baculovirus tenen tropismes específics cap als invertebrats amb més de 600 espècies d'hostes animals descrites. No es coneix que es repliquin en mamífers i altres animals vertebrats.
El primer registre d'aquesta família de virus és del segle VI per la malaltia que produïren a la larva de la papallona de la seda a la Xina. Cap a la dècada de 1940 es va estudiar i es va fer ús d'aquests virus com a plaguicida insecticida del tipus biològic. Des de la dècada de 1990 es fan servir en laboratori per a produir una proteïna útil en el cultiu cel·lular i en vacunes per a humans i altres animals.[2]
Index of Viruses - Baculoviridae (2006). In: ICTVdB - The Universal Virus Database, version 4. Büchen-Osmond, C (Ed), Columbia University, New York, USA. http://www.ncbi.nlm.nih.gov/ICTVdb/Ictv/fs_index.htm
Baculoviridae és una família de virus que es pot dividir en dos gèneres: nucleopolyhedrovirus (NPV) i granulovirus (GV). Mentre que els GV contenen només dues càpsides nucleiques per embolcall víric, els NPV contenen sia una simple càpside sia una múltiple (SNPV o MNPV) per embolcall.
Els baculovirus tenen tropismes específics cap als invertebrats amb més de 600 espècies d'hostes animals descrites. No es coneix que es repliquin en mamífers i altres animals vertebrats.
Die Familie Baculoviridae (Baculoviren) umfasst doppelsträngig-zirkuläre filamentöse DNA-Viren. Sie befallen ausschließlich Wirbellose, ihr Hauptwirt sind Mottenlarven, jedoch sind über 600 Wirte beschrieben. Baculoviren werden in der Gentechnologie zur Erzeugung von komplexen Eukaryotenproteinen in Zellkulturen sowie zur Vektorklonierung verwendet. Auch werden sie in der Landwirtschaft zur Kontrolle von Schadinsekten eingesetzt.
Die Familie ist Mitglied der 2021 neu geschaffenen Ordnung Lefavirales in der ebenfalls neuen Klasse Naldaviricetes. Diese Taxonomie löst provisorische Bezeichnungen ab wie beispielsweise „Baculo-like viruses“.[2]
Der Name "Baculo" geht auf lateinisch baculum ‚Stock, Stab‘ zurück und bezieht sich auf die Morphologie des Nukleokapsids.[3]
Sie sind weltweit verbreitet und wurden erstmals im 16. Jahrhundert beschrieben, als "Welkkrankheit" bei Seidenraupen. In den 1940er Jahren wurden sie als Biopestizide in Getreidefeldern eingesetzt, um Schadinsekten zu dezimieren. Durch spätere Untersuchungen wurden sie in den 1990er Jahren zur Produktion von Eukaryonten-Proteinen in Insektenzellkulturen eingesetzt, da sie dort einfach vermehrt werden können. Die Viren sind an Wirbellose angepasst mit über 600 möglichen Wirtsarten. Hauptsächlich befallen sie Mottenlarven, aber sie wurden auch bei Pflanzenwespen, Moskitos und Shrimps gefunden.
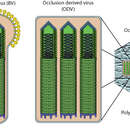
In seinem Lebenszyklus kommt das Virus in zwei Formen vor, dem ODV (englisch Occlusion Derived Virus), mit Proteinhülle, und dem BV (englisch Budding Virus). Die ODV-Form ist für die Erstinfektion notwendig, später werden dann BV-Partikel gebildet.[4]
Der Hauptinfektionsweg verläuft über kontaminierte Blätter, die von Insekten gefressen werden. Das Virus gelangt in den Verdauungstrakt, wo sich die Proteinhülle (ODV-Form) aufgrund der basischen Umgebung auflöst. Die freien Viren heften sich nun an die Darmepithelzellen und werden über Endocytose in ein Endosom aufgenommen, aus dem sie sich befreien und nun als Nukleokapside, vermutlich durch Aktin-Filamente, in den Zellkern transportiert werden. Hier erfolgt die Replikation des Virus. Nun werden unbehüllte Viren (BV) erzeugt und das Virus infiziert die umliegenden (basolateralen) Epithelzellen. Die BV-Partikel umhüllen sich jedoch auch locker mit Zellmembran, die Glycoproteine des Virus enthält.
Die Infektion kann in drei Phasen unterteilt werden: die frühe Phase (0–6 Stunden), die späte Phase (6–24 h) und die sehr späte Phase (18–24 bis 72 h nach der Infektion). Bis zur späten Phase werden noch unbehüllte BV Formen freigesetzt, in der letzten Phase jedoch nur ODV Formen, die durch die Kernmembran knospen und mit Viren gefüllte Partikel bilden, die mit OB-(Occlusion Body)-Protein gefüllt sind. Diese besitzen nun eine größere Resistenz gegen Umweltfaktoren als die BV-Form und dienen der Verbreitung des Virus auf den nächsten Wirt. Da beim Austritt der Viren die Zelle lysiert wird, kommt es im gesamten Insekt zu einer regelrechten Verflüssigung, wobei nur die Chitinhülle intakt bleibt. Diese platzt später auf und verstreut die Viren auf der gesamten Blattoberfläche. Diese Eigenschaft gab der durch dieses Virus verursachten Erkrankung auch den englischen Namen “wilting disease” ‚Welkkrankheit‘.
Das bestuntersuchte Baculovirus ist Autographa californica multicapsid nucleopolyhedrovirus (AcMNPV). Es wurde erstmals aus einer Gammaeulenart (Autographa californica) isoliert, die auf Luzerne lebt. Das Genom hat 134 kBp (kilo Basenpaare) und besitzt 154 Offene Leserahmen (englisch open reading frames, ORFs). Das wichtigste Kapsidprotein ist VP39, es bildet mit einigen kleineren Proteinen das Nukleokapsid (21 nm × 260 nm), das die DNA umhüllt, die wiederum vom Protein p6.9 umgeben ist.
Die BV-Partikel benötigen für eine Infektion das Glycoprotein gp64 (Genprodukt 64), welches an den Enden des fadenförmigen Virions vorkommt. Dieses Protein findet sich nicht in den ODV-Partikeln, hier gibt es einige Proteine die nur bei ODV vorkommen. Unterschiede gibt es auch in der Zusammensetzung der Lipide der Hülle, bei BV kommt Phosphatidylserin vor, ODV enthält Phosphatidylcholine und Phosphatidylethanolamine.
Im Laufe der Evolution des Virus hat sich das gp64 verändert. Ld139, (auch Baculovirus-F-Protein) aus dem Schwammspinner Lymantria dispar (LdMNPV) ist vermutlich ein ursprüngliches Membranfusionsprotein (zur Freisetzung des Virus aus der Wirtszelle), das durch gp64 als nicht "gleichwertiges" Protein ersetzt wurde, da Ld139 und gp64 im Genom vieler Baculovirus-Arten vorkommen (Maulbeerspinner, Gammaeule, Douglasien-Bürstenspinner (Familie der Trägspinner)).
Gp64 selbst ist ein homotrimeres Protein, das an den Polen der filamentösen BV-Partikeln sitzt. Es besteht aus 512 Aminosäuren, mit vier Glykolisierungssstellen an Asparagin-Resten und eine N-terminale Signalsequenz von 20 Aminosäuren, weiters Oligomerisations- und Fusionsdomänen, sowie eine hydrophobe Transmembrandomäne am C-terminalen Ende von sieben Aminosäuren. Es wird in der frühen und in der späten Phase der Infektion gebildet mit einem Maximum 24 bis 26 Stunden nach der Infektion. Die Trimerisation (Zusammenlagerung von je 3 gp64-Einheiten) über interne Cystinbindungen ist wichtig für den Transport zur Zelloberfläche, da nur 33 % des gp64 die Oberfläche erreichen und Monomere innerhalb der Zelle abgebaut werden.
Dieses Protein ist essentiell für die Knospung des Virions aus der Zelle, sowie für die Infektion von weiteren Zellen im Infektionszyklus. Seine Hauptaufgabe ist die über den pH-Wert regulierte Fusion mit dem Endosom. Obwohl gp64 notwendig ist für die Virusvermehrung, können Mutanten, denen gp64 fehlt, über Substitution mit Ld130 oder dem G-protein des "Vesikulären Stomatitis-Virus" (Vesicular stomatitis virus, VSV) wieder funktionsfähig gemacht werden.
Innerhalb der Familie wurden früher zwei Gattungen unterschieden: Nucleopolyhedrovirus (NPV) und Granulovirus (GV, jetzt Gattung Betabaculovirus). Bei Betabaculovirus kommt pro Hülle nur ein Nukleokapsid, bei den aus der Gattung Nucleopolyhedrovirus neu unterteilten Gattungen Alpha-, Gamma- und Deltabaculovirus kommen entweder einzelne (SNPV) oder mehrere (MNPV) Nukleokapside pro Hülle vor. Die Virusteilchen (Virionen, Nukleokapsid und Hülle) sind nun abermals in eine Granulin (GV)- oder Polyhedrin (NPV)-Matrix eingebettet. Die Betabaculoviren besitzen nur ein Virion pro Granulinmatrix, bei den anderen Gattungen sind mehrere Virionen in der Polyhedrinmatrix.
Nach ICTV (Master Species List#35, Stand April 2020) besteht die Familie aus den folgenden vier Gattungen (die Typusspezies und ggf. weitere Arten sind mit angegeben):
Die Familie Baculoviridae (Baculoviren) umfasst doppelsträngig-zirkuläre filamentöse DNA-Viren. Sie befallen ausschließlich Wirbellose, ihr Hauptwirt sind Mottenlarven, jedoch sind über 600 Wirte beschrieben. Baculoviren werden in der Gentechnologie zur Erzeugung von komplexen Eukaryotenproteinen in Zellkulturen sowie zur Vektorklonierung verwendet. Auch werden sie in der Landwirtschaft zur Kontrolle von Schadinsekten eingesetzt.
Die Familie ist Mitglied der 2021 neu geschaffenen Ordnung Lefavirales in der ebenfalls neuen Klasse Naldaviricetes. Diese Taxonomie löst provisorische Bezeichnungen ab wie beispielsweise „Baculo-like viruses“.
Baculoviridae is a family of viruses. Arthropods, among the most studied being Lepidoptera, Hymenoptera and Diptera, serve as natural hosts. Currently, 85 species are placed in this family, assigned to four genera.[1][2][3]
Baculoviruses are known to infect insects, with over 600 host species having been described. Immature (larval) forms of lepidopteran species (moths and butterflies) are the most common hosts, but these viruses have also been found infecting sawflies, and mosquitoes. Although baculoviruses are capable of entering mammalian cells in culture,[4] they are not known to be capable of replication in mammalian or other vertebrate animal cells.
Starting in the 1940s, they were used and studied widely as biopesticides in crop fields. Baculoviruses contain a circular, double-stranded DNA (dsDNA) genome ranging from 80 to 180 kbp.
The earliest records of baculoviruses can be found in the literature from as early as the 16th century in reports of "wilting disease" infecting silkworm larvae.[5] Starting in the 1940s, the viruses were used and studied widely as biopesticides in crop fields. Since the 1990s, they have been employed to produce complex eukaryotic proteins in insect cell cultures (see Sf21, High Five cells). These recombinant proteins have been used in research and as vaccines in both human and veterinary medical treatments (for example, the most widely used vaccine for prevention of H5N1 avian influenza in chickens was produced in a baculovirus expression vector). More recently, baculoviruses were found to transduce mammalian cells with a suitable promoter.[6]
The baculovirus lifecycle involves two distinct forms of virus. Occlusion-derived virus (ODV) is present in a protein matrix (polyhedrin or granulin) and is responsible for the primary infection of the host, while the budded virus (BV) is released from the infected host cells later during the secondary infection.
Baculoviruses have very species-specific tropisms among the invertebrates with over 700 host species having been described. Immature (larval) forms of lepidopteran species are the most common hosts, but these viruses have also been found infecting sawflies and mosquitoes. Reports of baculovirus infections of shrimp and beetles (e.g. Oryctes rhinocerus) were found to be nudiviruses, a closely related lineage to the baculoviruses.
Typically, the initial infection occurs when a susceptible host insect feeds on plants that are contaminated with the occluded form of the virus. The protein matrix dissolves in the alkaline environment of the host midgut (stomach), releasing ODVs that then fuse to the columnar epithelial cell membranes of the host intestine and are taken into the cell in endosomes. Nucleocapsids escape from the endosomes and are transported to nucleus. This step is possibly mediated by actin filaments. Viral transcription and replication occur in the cell nucleus and new BV particles are budded out from the basolateral side to spread the infection systemically. During budding, BV acquires a loosely fitting host cell membrane with expressed and displayed viral glycoproteins.
After baculovirus infection, three distinct phases occur:
While BV is produced in the late phase, the ODV form is produced in the very late phase, acquiring the envelope from host cell nucleus and embedded in the matrix of occlusion body protein. These occlusion bodies are released when cells lyse to further spread baculovirus infection to next host. The extensive lysis of cells frequently causes the host insect to literally disintegrate, thus the reason for the historic name "wilting disease". The complete ODV-polyhedrin particles are resistant to heat and light inactivation, whereas the naked BV virion is more sensitive to environment.
When infecting a caterpillar, the advanced stages of infection cause the host to feed without resting, and then to climb to the higher parts of trees, including exposed places they would normally avoid due to the risk of predators. This is an advantage for the virus since (when the host dissolves) it can drip down onto leaves, which will be consumed by new hosts.[7]
The virus is unable to infect humans in the way it does insects, because human stomachs are acid-based and NPV requires an alkaline digestive system in order to replicate. It is possible for the virus crystals to enter human cells, but not to replicate to the point of causing illness.[8]
The most studied baculovirus is Autographa californica multicapsid nucleopolyhedrovirus (AcMNPV). The virus was originally isolated from the alfalfa looper (a lepidopteran) and contains a 134 kbp genome with 154 open reading frames. The major capsid protein VP39 together with some minor proteins forms the nucleocapsid (21 nm x 260 nm) that encloses the DNA with p6.9 protein.
BV acquires its envelope from the cell membrane and requires a glycoprotein, gp64, to be able to spread systemic infection. This protein forms structures called peplomers on one end of the budded virus particle, but is not found on ODVs (although several other proteins are only associated with the ODV form). Some differences also exist in the lipid composition of the viral envelope of the two forms. While the BV envelope consists of phosphatidylserine, ODV contains phosphatidylcholine and phosphatidylethanolamine.
A nucleocapsid assembly-essential element (NAE) was identified in the AcMNPV genome. The NAE is an internal cis-element within the ac83 gene. The nucleocapsid assembly is not dependent on the ac83 protein product.[9]
During periods of evolution, the baculoviral envelope glycoproteins have undergone changes. Ld130, also known as baculovirus F-protein from Lymantria dispar (LdMNPV) is suggested to be an ancestral envelope fusion protein which has been replaced by non-orthologous gene replacement with gp64 in AcMNPV, Bombyx mori (BmNPV) and Orgyia pseudotsugata (OpMNPV) while they still retain the ld130 gene.
Gp64 is a homotrimeric membrane glycoprotein that is polarly present on the rod-shaped virion. It consists of 512 amino acids (aa) with four glycosylation sites at asparagine residues and has a N-terminal signal sequence (20 aa), oligomerization and fusion domain and a hydrophobic transmembrane domain near the C-terminus (7 aa).
It is produced in both early and late phases of the infection cycle with a maximal rate of synthesis occurring in 24–26 h after infection. Trimerization with intermolecular cysteine-bonds seems to be a crucial step for protein transport to cell surface, since only 33% of synthesized protein reaches cell surface, as monomeric gp64 is degraded within the cells.
Gp64 is essential for efficient budding of the virion and for the cell-to-cell transmission during the infection cycle, as well as viral entry, i.e. causing viral trophism and endosome-mediated uptake to the cell. The major function of the gp64 envelope protein is to cause the pH-mediated envelope fusion to the endosome. Although gp64 has variety of essential functions, it has been reported that gp64-null baculoviruses can be substituted with other viral glycoproteins such as Ld130, G-protein of Vesicular stomatitis virus. These substitutions will result in functional virons.
Baculovirus expression in insect cells represents a robust method for producing recombinant glycoproteins or membrane proteins.[10][11][12] Baculovirus-produced proteins are currently under study as therapeutic cancer vaccines with several immunologic advantages over proteins derived from mammalian sources.[13]
Lymantria dispar (commonly known as the gypsy moth), a serious pest of forest trees, has been successfully contained by releasing gypsy moth baculovirus (NPV) preparations. Certain species of sawfly (Neodiprion sertifer, N. lecontei, N. pratti pratti, etc.), have also been successfully controlled by NPV treatments specific to them.
Members of the genus Heliothis—cosmopolitan insects that attack at least 30 food- and fibre-yielding crop plants—have been controlled by the application of Baculovirus heliothis. In 1975, the United States Environmental Protection Agency registered the B. heliothis preparations.
NPV preparations have also been used commercially against pests such as Trichoplusia (under the biotrol-VTN brand name) and the cotton leafworm (under the biotrol-VSE brand name).
Baculoviruses are incapable of replicating within the cells of mammals and plants.[14] They have a restricted range of hosts they can infect that is typically restricted to a limited number of closely related insect species. Because baculoviruses are not harmful to humans, they are considered a safe option for use in research applications. They are also used as biological agents as in the case of the Indian mealmoth, a grain-feeding pest.[15]
The name of this family has been derived from the Latin word baculus, meaning "stick". The family has been divided into four genera: Alphabaculovirus (lepidopteran-specific nucleopolyhedroviruses), Betabaculovirus (lepidopteran-specific granuloviruses), Gammabaculovirus (hymenopteran-specific nucleopolyhedroviruses), and Deltabaculovirus (dipteran-specific nucleopolyhedroviruses).[16]
Nuclear Polyhedrosis Virus (NPV) was once listed by the International Committee on Taxonomy of Viruses as a subgenus of Eubaculovirinae, but the term now refers to 35 species of the family Baculoviridae, mostly alphabaculoviruses, but also one deltabaculovirus and two gammabaculoviruses.
The polyhedral capsid from which the virus gets its name is an extremely stable protein crystal that protects the virus in the external environment. It dissolves in the alkaline midgut of moths and butterflies to release the virus particle and infect the larva.[17] An example of an insect that it infects is the fall webworm.[18]
Baculoviruses are thought to have evolved from the Nudiviridae family of viruses 310 million years ago.[19]
Baculoviridae is a family of viruses. Arthropods, among the most studied being Lepidoptera, Hymenoptera and Diptera, serve as natural hosts. Currently, 85 species are placed in this family, assigned to four genera.
Baculoviruses are known to infect insects, with over 600 host species having been described. Immature (larval) forms of lepidopteran species (moths and butterflies) are the most common hosts, but these viruses have also been found infecting sawflies, and mosquitoes. Although baculoviruses are capable of entering mammalian cells in culture, they are not known to be capable of replication in mammalian or other vertebrate animal cells.
Starting in the 1940s, they were used and studied widely as biopesticides in crop fields. Baculoviruses contain a circular, double-stranded DNA (dsDNA) genome ranging from 80 to 180 kbp.
Baculoviridae es una familia de virus infectivos para artrópodos. Poseen un genoma con ADN de cadena doble como ácido nucleico, por lo que se incluyen en el Grupo I de la Clasificación de Baltimore. El genoma es circular con un tamaño de 80-180 kpb. Se caracterizan por albergar dicha información genética en una cápside recubierta de envoltura viral y estructuralmente definida por una simetría cilíndrica, de un tamaño (con envoltura) de 40-110 por 200-400 nm; y por ensamblar los viriones maduros en el núcleo como compartimento celular.
Se conocen cuatro géneros:
Los baculovirus exhiben un tropismo muy específico con la especie de invertebrado que infectan, con más de 600 especies de huéspedes descritas. Los huéspedes más comunes son las larvas de polillas, pero estos virus también se pueden encontrar infectando moscas sierra, mosquitos y camarones. No se conocen especies que infecten células de mamíferos o de otros animales vertebrados.
Los baculovirus son patógenos específicos que atacan a los insectos y otros artrópodos, por lo que uno de sus usos más frecuentes es como insecticida biológico, es decir, como agentes para controlar las plagas que afectan a las cosechas. El comportamiento de los baculovirus es similar al del virus herpes que afecta a los humanos, ya que el virus tiene una doble estrategia que le permite permanecer asociado al huésped sin causar ninguna enfermedad, pero, por otra parte, algún elemento todavía desconocido sirve de desencadenante para que el virus se reactive, empiece a crecer en el huésped y acabe matándolo.
Los baculovirus presentan dos formas distintas durante su ciclo vital. Los virus derivados ocluidos (ODV) son responsables de la infección primaria del huésped y están presentes en una matriz proteica (polihedrina o granulina), mientras que los virus de gemación (BV) se liberan de las células huésped durante la infección secundaria.
Típicamente, la infección inicial se produce cuando el huésped, un insecto susceptible, se alimenta de las plantas que están contaminadas con la forma de oclusión del virus. La proteína de la matriz se disuelve en el medio alcalino del estómago del huésped, liberando ODVs que entonces se fusionan con la membrana celular del epitelio columnar del intestino del huésped y se introducen en la célula en endosomas. Las nucleocápsides escapan de los endosomas y son transportados al núcleo. Este paso es posiblemente mediado por filamentos de actina. La transcripción y replicación viral se produce en el núcleo de la célula y nuevas partículas BV se desprenden por gemación desde el lado basolateral que propagan la infección sistémicamente. Durante la gemación, las partículas BV adquieren partes de la membrana de la célula huésped con glicoproteínas virales expresadas.
La infección por baculovirus puede dividirse en tres fases: temprana (0-6 horas de la infección), tardía (6-24 horas) y muy tardía (18-24 a 72 horas). La forma BV se produce en la fase tardía, mientras que la ODV lo hace en la fase muy tardía. Esta adquiere la envoltura a partir del núcleo de la célula huésped y es embebida en la matriz de proteína del cuerpo de oclusión. Estos cuerpos de oclusión son liberados cuando las células se lisan para propagar la infección a otros huéspedes. La lisis extensiva de las células con frecuencia hace que el insecto huésped literalmente se derrita. Para garantizar su supervivencia en la naturaleza, las partículas de ODV-polihedrina son resistentes a inactivación por luz y calor, mientras que BV es más sensible a las condiciones del medio ambiente.
El baculovirus más estudiado es Nucleopolyhedrovirus multicápside de Autographa californica (AcMNPV). El virus fue aislado originalmente del lepidóptero Autographa californica y contiene un genoma de 134 kpb de longitud con 154 marcos abiertos de lectura (ORF). La principal proteína de la cápside, VP39, junto con algunas proteínas pequeñas, forma la nucleocápside (21 nm x 260 nm) que encierra el ADN con proteína p6.9.
La forma BV adquiere su envoltura de la membrana celular y requiere una glicoproteína gp64 para poder difundir la infección sistémica. Esta proteína forma estructuras denominadas peplómeros sobre un extremo de las partículas BV, pero no se encuentra en las ODV (aunque varias otras proteínas se presentan únicamente en la forma ODV). También existen algunas diferencias en la composición de lípidos de la envoltura viral de las dos formas. Mientras que la envoltura de BV consta de fosfatidilserina, ODV contiene fosfatidilcolina y fosfatidiletanolamina.
Baculoviridae es una familia de virus infectivos para artrópodos. Poseen un genoma con ADN de cadena doble como ácido nucleico, por lo que se incluyen en el Grupo I de la Clasificación de Baltimore. El genoma es circular con un tamaño de 80-180 kpb. Se caracterizan por albergar dicha información genética en una cápside recubierta de envoltura viral y estructuralmente definida por una simetría cilíndrica, de un tamaño (con envoltura) de 40-110 por 200-400 nm; y por ensamblar los viriones maduros en el núcleo como compartimento celular.
Se conocen cuatro géneros:
Betabaculovirus. Contienen una única nucleocápside por envoltura y los viriones están envueltos en una matriz de granulina. Además, contienen un único virión por cuerpo de oclusión. Alphabaculobirus, Deltabaculovirus y Gammabaculovirus. Pueden contener una o varias nucleocápsides por envoltura y los viriones (incluyendo la envoltura) están adicionalmente envueltos en una matriz de polihedrina. Además, contienen múltiples viriones por cuerpo de oclusión.Los baculovirus exhiben un tropismo muy específico con la especie de invertebrado que infectan, con más de 600 especies de huéspedes descritas. Los huéspedes más comunes son las larvas de polillas, pero estos virus también se pueden encontrar infectando moscas sierra, mosquitos y camarones. No se conocen especies que infecten células de mamíferos o de otros animales vertebrados.
Los baculovirus son patógenos específicos que atacan a los insectos y otros artrópodos, por lo que uno de sus usos más frecuentes es como insecticida biológico, es decir, como agentes para controlar las plagas que afectan a las cosechas. El comportamiento de los baculovirus es similar al del virus herpes que afecta a los humanos, ya que el virus tiene una doble estrategia que le permite permanecer asociado al huésped sin causar ninguna enfermedad, pero, por otra parte, algún elemento todavía desconocido sirve de desencadenante para que el virus se reactive, empiece a crecer en el huésped y acabe matándolo.
Bakuloviirused (Baculoviridae) on viiruste sugukond, kelle looduslikud peremeesorganismid kuuluvad lülijalgsete, liblikaliste, kiletiivaliste, kahetiivaliste ning kümnejalaliste hulka.[1][2]
On uuritud 600 selgrootute liiki, keda bakuloviirused nakatavad. Kõige tüüpilisemad kandjad on ööliblikate vastsed, kuid bakuloviirused nakatavad ka pidevkehalisi, pistesääsklasi ning krevette. Bakuloviirused suudavad siseneda ka imetajate [3] või teiste selgroogsete loomade rakkudesse, kuid ei suuda seal paljuneda.
Tuntakse 55 liiki bakuloviirusi, kes jagunevad nelja perekonda.[4]
1940. aastatel hakati uurima võimalusi kasutada bakuloviirusi biopestitsiididena viljapõldudel. Tänapäeal on neile leitud mitmeid biotehnoloogilisi rakendusi ja nende põhja on arendatud välja BacMam-süsteem.
Bakuloviiruste genoom on 80–180 tuhande aluspaari suurune tsirkulaarne DNA kaksikahel.
Bakuloviirusi mainitakse kirjanduses esmakordselt 16. sajandil seoses siidiussidel esineva närbumistõvega. Alates 1940. aastatest on bakuloviirusi laialdaselt uuritud ja kasutatud biopestitsiididena. Alates 1990. aastatest on neid kasutatud keeruliste eukarüootsete valkude tootmiseks putukarakukultuurides. Nõnda toodetud rekombinantseid valke on kasutatud nii teaduses kui ka vaktsiinidena inimeste ja loomade raviks. Näiteks on levinuim H5N1 linnugripi tüve vaktsiin kanadele toodetud bakuloviiruste ekpressioonivektorit kasutades. Hiljuti on avastatud, et bakuloviirusi kasutades saab imetajarakkudesse viia sobivaid promootoreid.[5] Need potentsiaalselt kliinilised kasutusalad on alates 1995. aastast viinud bakuloviiruste intensiivsema uurimiseni.
Bakuloviiruse paljunemistsükkel koosneb kahest erinevast viiruse vormist. Oklusioonikehaga viirused on kaitstud valgulise kestaga, mis koosneb polühedriinist või granuliinist. Oklusioonikehaga viirused vastutavad esmase nakatamise eest, samas kui pungunud bakuloviirused vabanevad peremeesrakust hiljem sekundaarse nakatamise käigus. Bakuloviirustel on väga liigispetsiifilised nakatamismehanismid. Kirjeldatud on üle 600 erineva peremeesorganismi. Ööliblikate vastsed on kõige tüüpilisemad peremeesorganismid, kuid bakuloviirused nakatavad ka pidevkehalisi, pistesääsklasi ning krevette.
Looduses toimub esmane nakatamine üldiselt siis, kui putukas sööb taime, mis on saastunud bakuloviiruse oklusioonikehaga vormiga. Valguline kest laguneb peremeesorganismi kõhu aluselises keskkonnas. Vabaneb viirusosake, mis liitub soolestikus epiteelraku membraaniga ning siseneb rakku endosoomis. Nukleokapsiidid lahkuvad endosoomidest ning transporditakse rakutuuma. Seda etappi aitavad läbi viia tõenäoliselt aktiinifilamendid.
Rakutuumas toimub bakuloviiruse transkriptsioon ja replikatsioon ning uued bakuloviirused punguvad läbi basolateraalse membraanidomeeni ja levitavad nakkust edasi. Pungumise käigus jääb viirusosakest ümbritsema peremeesraku membraan, millel on ekspresseeritud viiruse glükoproteiinid. Bakuloviiruse nakatumise saab jagada kolme selgelt eristuvasse faasi:
Hilise faasi jooksul vabanevad rakkudest pungunud bakuloviirused, samas kui oklusioonikehaga viirusi toodetakse väga hilises faasis. Oklusioonikehaga viirused saavad rakutuumast kaasa membraani ja koos sellega sisestatakse need valkudest koosnevasse oklusioonikeha maatriksisse. Oklusioonikehaga viirused vabanevad rakkude purunemisel ning nakatavad järgmisi peremeesorganisme. Nakkuse tõttu puruneb samal ajal suur hulk rakke, mis viib terve organismi lagunemisele. Terved oklusioonikeha-polühedriin viirusosakesed on vastupidavad ning ei kaota aktiivsust temperatuuri ja valguse mõjul. Pungunud bakuloviirused võivad keskkonnategurite toimel kaotada nakkusvõimelisuse palju kergemini.

Enim uuritud bakuloviirus on Autographa californica multikapsiidne nukleopolühedroviirus (AcMNPV). See viirus eraldati esmakordselt tähtöölaste perekonda kuuluvast liigist Autographa californica. Viiruse genoom koosneb 134 tuhandest aluspaarist. Viiruse nukleokapsiid koosneb peamiselt valgust nimega VP39 ning mõningatest väiksematest valkudest. Nukleokapsiidi pikkus on 260 nm ja diameeter on 21 nm. Bakuloviirus saab oma membraani rakumembraani osana ning sellel peab olema ekspresseeritud glükoproteiin gp64, et viirusosake oleks nakatamisvõimeline. See valk moodustab peplomeerideks kutsutud struktuure pungunud bakuloviiruse ühte otsa, kuid neid ei moodustu oklusioonikehaga viiruse pinnale. Oklusioonikehaga viirusega seostatakse aga mõningaid teisi valke. Kummagi viiruse vormi membraanid erinevad natuke ka lipiidse koostise poolest. Pungunud bakuloviiruste membraan koosneb fosfatitüülseriinist, kuid oklusioonikehaga viiruste membraanis esinevad fosfatitüülkoliin ning fosfatitüületanoolamiin.
Evolutsiooni käigus on bakuloviiruse membraani glükoproteiinid muutunud. Arvatakse, et ld130 on membraanvalk, mis on gp64 eellane ning mille on funktsionaalselt asendanud mitteortogonaalse geeniasenduse käigus gp64, kuid ld130 kodeeriv geen on viiruse genoomis endiselt olemas. Gp64 on homotrimeerne membraani glükoproteiin, mis esineb pulgakujulises viirusosakeses polaarselt. See valk koosneb 512 aminohappejäägist. Valk on glükosüleeritud nelja asparagiini jäägi juurest, sellel on 20 aminohappejäägi pikkune signaaljärjestus N-terminaalses otsas, oligomerisatsiooni-ja liitumisdomeen ja hüdrofoobne transmembraanne seitsmest aminohappejäägist koosnev domeen C-terminali lähedal. Seda valku toodetakse kogu nakatumistsükli jooksul, kusjuures kõige rohkem toodetakse seda vahemikus 24–26 tundi pärast nakatumist. Trimeriseerumine molekulidevaheliste tsüsteiinijääkide vaheliste disulfiidsildade kaudu on võtmeetapp selleks, et valk jõuaks membraanile. Ainult 33% toodetud valgust jõuab membraanile, samas kui monomeerne gp64 lagundatakse. Gp64 on hädavajalik, et viirus saaks punguda, jõuda järgmise rakuni ning sellesse endosoomi kaudu siseneda. Gp64 põhjustab pH mõjul viiruse membraani liitumise endosoomiga. Kuigi gp64-l on palju tähtsaid omadusi, on avastatud, et gp64 asendamisel mõne teise viirustele omase glükoproteiiniga, näiteks Ld130-ga, jääb viirusosake endiselt toimivaks.
Putukakultuurides on bakuloviiruste abil on võimalik toota rekombinantseid glükoproteiine.[6][7] Sel viisil toodetud valke uuritakse, eesmärgiga kasutada neid vähivastaste vaktsiinidena ning neil on mitmeid eeliseid võrreldes imetajarakukultuurides toodetud valkudega.[8] Bakuloviirusi võib kasutada ka imetajarakkudes valkude ekspresseerimiseks BacMam-süsteemi kasutades. Nii saab bakuloviiruste abil panna rakud tootma nii retseptoreid, biosensoreid kui muid valke.
Bakuloviiruste efektiivseks kasutamiseks on vajalik teada viirusosakeste hulka ruumalaühikus. Arvulise hinnangu andmiseks kasutatakse mitut ühikut: nakatamisvõimelisi viirusosakesi milliliitris ehk ivp/ml ja viiruslaiku moodustavaid viirusosakesi milliliitris ehk pfu/ml.
Pfu/ml määratakse viiruslaigu meetodil. Huvipakkuvast proovist valmistatakse sobiv lahjendus, nakatatakse sellega viirusele sobiva rakuliini monokihti, eemaldatakse teatud aja möödudes viirus ning kaetakse rakud agaroosgeeliga. Nakatumisel rakud purunevad ja neist moodustuvad laigud loendatakse.[9][10] Ivp/ml määramiseks kasutatakse erinevaid meetodeid, mille eesmärgiks on määrata, milline viiruse kogus tapab peremeesorganismidest 50% või tekitab tsütopaatilise muutuse.
Bakuloviirused võivad siseneda ka imetajarakkudesse, sealhulgas inimese rakkudesse ning viia sinna oma DNA, kuid ei ole seal võimelised paljunema ja nakkust levitama.[11] Bakuloviirused saavad nakatada ja paljuneda vaid väikese hulga sarnaste liikide organismides. Kuna bakuloviirused ei kujuta endast inimesele ohtu, peetakse neid ohutuks kasutada.
Bakuloviirused jagunevad nelja perekonda: alfabakuloviirused (liblikaliste-spetsiifilised nukleopolühedroviirused), beetabakuloviirused (liblikaliste-spetsiifilised granuloviirused), gammabakuloviirused ja deltabakuloviirused.[12]
Arvatakse, et bakuloviirused arenesid nudiviiruste perekonnast 310 miljonit aastat tagasi.[13]
|displayauthors= suggested (juhend) Bakuloviirused (Baculoviridae) on viiruste sugukond, kelle looduslikud peremeesorganismid kuuluvad lülijalgsete, liblikaliste, kiletiivaliste, kahetiivaliste ning kümnejalaliste hulka.
On uuritud 600 selgrootute liiki, keda bakuloviirused nakatavad. Kõige tüüpilisemad kandjad on ööliblikate vastsed, kuid bakuloviirused nakatavad ka pidevkehalisi, pistesääsklasi ning krevette. Bakuloviirused suudavad siseneda ka imetajate või teiste selgroogsete loomade rakkudesse, kuid ei suuda seal paljuneda.
Tuntakse 55 liiki bakuloviirusi, kes jagunevad nelja perekonda.
1940. aastatel hakati uurima võimalusi kasutada bakuloviirusi biopestitsiididena viljapõldudel. Tänapäeal on neile leitud mitmeid biotehnoloogilisi rakendusi ja nende põhja on arendatud välja BacMam-süsteem.
Bakuloviiruste genoom on 80–180 tuhande aluspaari suurune tsirkulaarne DNA kaksikahel.
Bakulovirukset ovat hyvin laajalti levinnyt virusryhmä, joka infektoi lajispesifisesti niveljalkaisia (hyönteiset, hämähäkit ja äyriäiset). Viruksia tunnetaan yli 500 erilaista kantaa, joista useimmin käytetty ja bakulovirusten mallikappale on Autographa californica multicapsid nucleopolyhedro virus (AcMNPV).
1900-luvun puolivälissä bakulovirusten hyönteisiä infektoivat ominaisuudet huomattiin ja niiden käyttö tehokkaana biopestisidinä aloitettiin 1975. Käyttö sai myös aikaan runsaasti tutkimusta, jonka myötä selvitettiin bakulovirusten molekylaarista biologiaa, patologiaa ja ekologiaa. Bakulovirusten käytön turvallisuutta ja patogeenisuutta selkärankaisiin tutkittiin myös laajalti tuohon aikaan. Yhteenvetona on todettu, että erilaisissa turvallisuustesteissä ei havaittu virusten kykenevän infektioon tai aiheuttamaan allergisia reaktioita selkärankaisissa, eikä niissä myöskään todettu vasta-aineiden muodostumista koe-eläimille tai ihmisille ruoansulatuskanavan, hengitysteiden tai ihon kautta tapahtuvien altistumisten jälkeen.
Ensimmäinen bakuloviruksiin pohjautuva ekspressiovektori, jolla kyettiin tuottamaan proteiineja hyönteissoluissa esiteltiin vuonna 1983. Silloin käytetyimpiä solutyyppejä olivat Spodoptera frugiperda –peräiset SF-21 ja SF-9. Uudempana solutyyppinä ovat tulleet käyttöön Trichoplusia ni -peräiset High Five -solut. Bakulovirusten käytön proteiiniekspressioon tekee oivalliseksi se, että hyönteissolut ovat aitotumallisia (eukaryootti) soluja, ja siten kykenevät suorittamaan useille aitotumallisten solujen proteiineille tärkeät modifikaatiot paremmin kuin paljon käytetyt alkeistumalliset (prokaryootti) bakteerisolut. Esimerkkeinä modifikaatioista ovat mm. signaalipeptidin poistaminen, N-glykosylaatio, asylaatio, fosforylaatio ja karboksimetylaatio. Bakuloviruksia on alettu käyttää tästä syystä myös peptidinäyttötekniikan sovellutuksissa, joissa esiteltävän proteiinin post-translationaaliset modifikaatiot ovat oleellisia selektion kannalta.
1980-luvulla havaittiin, että bakulovirukset kykenevät penetroitumaan useisiin nisäkässolulinjoihin, mutta eivät kyenneet saamaan aikaan infektiota. Vuonna 1995 esiteltiin ensimmäisen kerran havainto, että muokatut AcMNPV:t kykenivät toimittamaan erillistä geenimateriaalia nisäkässoluihin. 1998 suoritettiin ensimmäinen ex vivo geeninsiirto hepatosyytti-solulinjaan ja vuonna 2000 julkaistiin tulokset joissa bakulovirusten todettiin kykenevän myös siirtämään geneettistä materiaalia in vivo adenoviruksiin verrattavalla tehokkuudella. Näiden havaintojen myötä tutkimus bakulovirusten käyttämiseksi geeniterapia-vektorina on kiihtynyt. Bakuloviruksilla on monia hyviä puolia perinteisesti käytettyihin geeniterapiavektoreihin verrattuna: kykenemättömyys jakautua nisäkässoluissa, suuri DNA-kapasiteetti, helppo valmistus ja alhainen sytotoksisuus. Huonoina puolina voidaan mainita mm. veren komplementin bakuloviruksia inaktivoiva vaikutus sekä ohimenevä (eng. transient) ekspressio. 2000-luvun puolivälin jälkeen virusten tutkiminen geeni- ja soluterapiakäyttöön on lisääntynyt. Uusimpina modifikaatioina ovat mm. lentiviruksiin siirretty gp64 pintaproteiini ja avidiini-gp64 fuusioproteiinilla päällystetyt bakulovirukset, joita on yhdessä raudasta koostuvien nanopartikkeleiden kanssa kyetty seuramaan MRIssä ensimmäisenä viruksena maailmassa.
Bakulovirukset koostuvat kahdesta suvusta, Nuclear polyhedrosis- viruksista (NPV) ja Granulosis- viruksista (GV). NPV:t tuottavat hyönteissoluissa infektion myötä suuria polyhedriini-proteiinimatrikseja, joiden sisällä on useita viruksia ja GV:t partikkeleita, joiden sisällä on vain yksi virioni.
Bakulovirukset ovat hyönteissoluja infektoivia viruksia ja varsin lajispesifisiä. AcMNPV infektoi spesifisesti yli 30:tä lepidoptera –suvun hyönteislajia. Tämä suuri isäntälajien määrä on kuitenkin poikkeuksellista bakulovirusten keskuudessa. Infektoidessaan hyönteissoluja bakulovirukset esiintyvät kahdessa erilaisessa muodossa: niin sanottuna okluusioperäisenä viruksena (ODV: occlusion derived virus, joskus myös nimellä PDV eli polyhedron derived virus), joka sisältää useita viruskapsideita polyhedriinimatriksiin ympäröitynä, joka taas on päällystynyt vaipalla. Toinen muoto on silmikoituva virus (budded virus, lyhyemmin BV, joskus myös extra-cellular virion eli ECV, ekstrasellulaarinen virus), jolloin yksittäistä viruskapsidia ympäröi oma vaippa proteiineineen. Edellä mainittu ODV-muoto on tarpeen suojaamaan fyysisesti viruksia luonnossa, esim. puiden lehdillä. ODV-muodon polyhedriinimatriksi suojaa viruksia hyvin tehokkaasti ympäristön vaikutuksilta ja lisää siten virusten infektiivisyyttä ekosysteemissä. Joutuessaan hyönteisen ruuansulatuksen alkaaliseen ympäristöön polyhedriinimatriksi hajoaa vapauttaen ODV-virionit. Nämä ensimmäisen aallon virionit ovat hyvin tehokkaita tartuttamaan suoliston epiteelisoluja, mutta ne eivät ole juurikaan tehokkaita levittämään infektiota ympärilleen.
Silmikoituva virus -tyyppi on taas tarpeen kun primäärinen infektio hyönteisen ruuansulatuselimistön soluihin on saavutettu. BV-virukset ovat erikoistuneet solusta-soluun infektioon, jonka tarkoituksena on levittää systeemistä infektiota ympäri hyönteisen elimistöä. Solujen lyysautumisen alkaminen johtaa ODV- muodon tuotannon käynnistymiseen infektion myöhäisessä vaiheessa ja pian isännän kuolemaan. Polyhedriinin suojaamat ODV- virukset jatkavat infektion levittämistä ekosysteemissä. Kahden erilaisen muodon käyttö viruksen lisääntymisessä lienee syy sen hyvään menestykseen luonnossa.
Bakuloviruksen soluunotto on tärkeä vaihe niin sen luontaisen kiertokulun kuin erilaisten geeniterapian sovellustenkin takia. Soluunoton kaikkia vaiheita ei toistaiseksi tunneta, eikä myöskään olla täysin selvillä siitä, miten viruksen kapsidi päätyy solun tumaan. Soluunotto hyönteisissä on kuitenkin analoginen mekanismeiltaan nisäkässoluissa tapahtuvaan soluunottoon. Seuraavat mekanismit ovat siis tämänhetkisen tiedon mukaan yhteisiä AcMNPV:n tunkeutumisessa sekä hyönteissoluihin että nisäkässoluihin: tunkeutuminen soluun alkaa sitoutumisella isäntäsolun toistaiseksi tunnistamattomaan reseptoriin plasmamembraanilla, reseptorin kuroutumisella sisäänpäin ja endosyyttisen vesikkelin muodostumisella. Endosomin happamoituminen saa aikaan viruksen vaipan ja endosomin membraanin fuusion, joka vapauttaa nukleokapsidin sytoplasmaan. Viruksen kapsidi kuljetetaan tämän jälkeen tumaan replikaatiota varten.
Silmikoituva virus (BV) on rakenteeltaan sauvamainen ja sitä ympäröi membraani, jonka toisessa päässä on gp64–vaippaproteiineja peplomeerimäisenä rakenteena. Membraanin sisältä löytyy viruksen kapsidi, jonka sisällä viruksen sirkulaarinen genomi on. Okluusioperäinen virus (ODV) sisältää puolestaan useita kapsideita, ilman gp64-proteiinia, yhteisessä polyhedriiniproteiinista muodostuvassa matriisissa. AcMNPV:n genomi on 133 894 emäsparin mittainen ja muodostuu kaksisäikeisestä DNA:sta. Se koodaa potentiaalisesti 150 proteiinia.
Polyhedriini ja granuliini-proteiini (AcMNPV:n orf8 eli open reading frame 8, luettava geeni 8) ovat 245 aminohapon muodostamia proteiineja. Polyhedriini on myös hyvin konservoitunut proteiini eri bakuloviruskantojen keskuudessa. AcMPV:n polyhedriinipromoottori polh on hyvin käytetty proteiiniekspressiossa. Tämä johtuu siitä, että jopa puolet solun proteiineista voi infektion aikana muodostua tämän promoottorin alla. Polyhedriini ei ole oleellinen viruksen replikaatiolle tai sen infektiivisyydelle.
ODV-muotoisen viruksen polyhedrarakennetta ympäröi vaippa (eng. polyhedron envelope eli PE tai polyhedron calyx). Tämän vaipan pääasiallinen rakenneproteiini on PEP (Polyhedron envelope protein, pp34, orf131). Polyhedra-rakenteen toinen oleellinen proteiini on p10 (orf 137), jonka ekspressio tapahtuu myöhemmin infektion aikana. Myös se on oleellinen polyhedran stabiiliuden ja rakenteen kannalta.
BV:llä ja ODV:llä on hyvin samankaltaiset kapsidit, vain proteiini ODV-EC27:n on osoitettu löytyvän yksinomaan okluusiomuodosta. AcMNPV:n proteiini p6,9 (orf100) on DNA:han sitoutuva proteiini, joka on osallisena viruksen DNA:n pakkautumisessa sen kapsidin sisään. Se toimii spermassa histoneita korvaavien protamiinien tavoin ja on aminohappokoostumukseltaan samankaltainen. Se on oleellinen viruksen DNA:n pakkautumisessa kapsidin sisälle.
Mainitsemisen arvoinen kapsidiproteiini AcMNPV:lle on myös vp-39 (orf89), jota löytyy satunnaisesti kautta nukleokapsidin. Tämä viittaisi siihen, että se on kapsidin rakennekomponentti, erikoistuneen rakenteen sijaan. Proteiinin kysteiinitähteet ovat konservoituneet eri viruslajien keskuudessa, joka viittaisi niiden olevan oleellisia rakenteen kannalta.
Kapsidin päädyssä oleva ORF1629 (orf9) on erikoistunut kuppimainen rakenne, joka sulkee viruksen kapsidin. Viruksen kapsidin orientaatio infektion ja soluuntunkeutumisen aikana vaikuttaa kytkeytyvän ORF1629:n kautta kapsidin sisällä olevan DNA:n vapautumiseen kapsidista.
Bakuloviruksia ympäröivä vaippa koostuu kaksoiskerroksellisesta lipidikalvosta ja siihen liittyneistä proteiineista. Olennainen osa bakulovirusten rakennetta ovat niiden vaipan proteiinit, joilla on erilaisia rakennetta stabiloivia tai infektion kannalta olennaisia tehtäviä. ODV -ja BV -muotojen välillä on havaittavissa selviä eroja vaipan koostumuksen suhteen; ODV:n vaippa sisältää enemmän tyydyttyneitä rasvahappoja, kolesterolia ja tiheämmin proteiineja kuin BV:n vaippa. ODV:n vaippa koostuu tumakalvosta, kun taas BV:llä vaippa on peräisin solumembraanista. Tämän lisäksi osa eroista johtunee viruksen vaipan proteiinien osallistumisesta selektiivisesti membraanin ottamiseen isäntäsolun ulko- tai sisäpinnalta. BV-muotoinen virus, joka levittää infektiota systeemisesti, tarvitsee liikkuvamman vaipan ja erilaisen proteiinikoostumuksen kuin ODV, jonka vaipan proteiinit osallistuvat polyhedriinimatriksin kristallisoitumiseen.
Viruksen membraaniin on assosioitunut useita erilaisia proteiineja, niistä ODV-E66:n (orf46) tarkoituksena vaikuttaa olevan proteiinien targetointi, säilyttäminen ja/tai siirto ODV-muodon vaipalle. Samantapainen rakenne ja tehtävä vaikuttaa olevan myös ODV-E25:llä (orf94). p74 (orf138) -proteiinia ekspressoidaan infektion loppuvaiheessa ja geenin deleetio aiheuttaa virukselle kykenemättömyyden infektoida hyönteisiä ruuansulatuskanavan kautta.
Gp64 (orf128) on ainoa proteiini, joka löytyy yksinomaan BV-muodosta. Proteiini on lukukehyksen ennusteen mukaan 59 kDa:n kokoinen, mutta valmis proteiini on vahvasti glykolysoitu ja asyloitu ollen kooltaan 64 kDa. Geeniä säädellään aikaisella ja myöhäisellä (eng. late ja early) promoottorilla ja sen tuottama gp-64 proteiini on tärkein pohja fuusioproteiineille peptidinäyttötekniikassa. Gp64 on rakenteeltaan ja toiminnaltaan parhaiten tutkittu proteiini bakuloviruksissa, johtuen sen sopivuudesta erilaisiin tuotanto- ja näyttömenetelmiin.
Immunologisten ja biokemiallisten tutkimusten mukaan gp64 kerääntyy plasmamembraanille infektion alku- ja loppuvaiheissa. Viruksen nukleokapsidit tulevat vaipallisiksi työntyessään ulos solusta, jolloin viruksen vaippa muodostuu gp64-modifioidusta solumembraanista ja gp64-proteiineista. Itse viruksessa proteiinit ovat konsentroituneet viruspartikkelin toiseen päähän peplomeeri-rakenteiksi ollen olennaisia viruksen solusta-soluun infektiossa.
Gp64-proteiini sisältää N-terminaalisen signaalipeptidin ja C-terminaalisen ankkuridomeenin. Baculoviridae-heimon keskuudessa proteiini on erittäin konservoitunut. Runsaan glykolysaation lisäksi proteiini on asyloity, merkittävimmin palmitiinihapolla. Valmis gp64 trimerisoituu nopeasti disulfidisidosten kautta ja on membraanilla trimeerisessä muodossa. Trimerisaatiossa on osallisena proteiinin leusiini-vetoketju (amfipaattinen alfa-heliksi) rakenne.
Bakulovirukset ovat hyvin laajalti levinnyt virusryhmä, joka infektoi lajispesifisesti niveljalkaisia (hyönteiset, hämähäkit ja äyriäiset). Viruksia tunnetaan yli 500 erilaista kantaa, joista useimmin käytetty ja bakulovirusten mallikappale on Autographa californica multicapsid nucleopolyhedro virus (AcMNPV).
1900-luvun puolivälissä bakulovirusten hyönteisiä infektoivat ominaisuudet huomattiin ja niiden käyttö tehokkaana biopestisidinä aloitettiin 1975. Käyttö sai myös aikaan runsaasti tutkimusta, jonka myötä selvitettiin bakulovirusten molekylaarista biologiaa, patologiaa ja ekologiaa. Bakulovirusten käytön turvallisuutta ja patogeenisuutta selkärankaisiin tutkittiin myös laajalti tuohon aikaan. Yhteenvetona on todettu, että erilaisissa turvallisuustesteissä ei havaittu virusten kykenevän infektioon tai aiheuttamaan allergisia reaktioita selkärankaisissa, eikä niissä myöskään todettu vasta-aineiden muodostumista koe-eläimille tai ihmisille ruoansulatuskanavan, hengitysteiden tai ihon kautta tapahtuvien altistumisten jälkeen.
Ensimmäinen bakuloviruksiin pohjautuva ekspressiovektori, jolla kyettiin tuottamaan proteiineja hyönteissoluissa esiteltiin vuonna 1983. Silloin käytetyimpiä solutyyppejä olivat Spodoptera frugiperda –peräiset SF-21 ja SF-9. Uudempana solutyyppinä ovat tulleet käyttöön Trichoplusia ni -peräiset High Five -solut. Bakulovirusten käytön proteiiniekspressioon tekee oivalliseksi se, että hyönteissolut ovat aitotumallisia (eukaryootti) soluja, ja siten kykenevät suorittamaan useille aitotumallisten solujen proteiineille tärkeät modifikaatiot paremmin kuin paljon käytetyt alkeistumalliset (prokaryootti) bakteerisolut. Esimerkkeinä modifikaatioista ovat mm. signaalipeptidin poistaminen, N-glykosylaatio, asylaatio, fosforylaatio ja karboksimetylaatio. Bakuloviruksia on alettu käyttää tästä syystä myös peptidinäyttötekniikan sovellutuksissa, joissa esiteltävän proteiinin post-translationaaliset modifikaatiot ovat oleellisia selektion kannalta.
1980-luvulla havaittiin, että bakulovirukset kykenevät penetroitumaan useisiin nisäkässolulinjoihin, mutta eivät kyenneet saamaan aikaan infektiota. Vuonna 1995 esiteltiin ensimmäisen kerran havainto, että muokatut AcMNPV:t kykenivät toimittamaan erillistä geenimateriaalia nisäkässoluihin. 1998 suoritettiin ensimmäinen ex vivo geeninsiirto hepatosyytti-solulinjaan ja vuonna 2000 julkaistiin tulokset joissa bakulovirusten todettiin kykenevän myös siirtämään geneettistä materiaalia in vivo adenoviruksiin verrattavalla tehokkuudella. Näiden havaintojen myötä tutkimus bakulovirusten käyttämiseksi geeniterapia-vektorina on kiihtynyt. Bakuloviruksilla on monia hyviä puolia perinteisesti käytettyihin geeniterapiavektoreihin verrattuna: kykenemättömyys jakautua nisäkässoluissa, suuri DNA-kapasiteetti, helppo valmistus ja alhainen sytotoksisuus. Huonoina puolina voidaan mainita mm. veren komplementin bakuloviruksia inaktivoiva vaikutus sekä ohimenevä (eng. transient) ekspressio. 2000-luvun puolivälin jälkeen virusten tutkiminen geeni- ja soluterapiakäyttöön on lisääntynyt. Uusimpina modifikaatioina ovat mm. lentiviruksiin siirretty gp64 pintaproteiini ja avidiini-gp64 fuusioproteiinilla päällystetyt bakulovirukset, joita on yhdessä raudasta koostuvien nanopartikkeleiden kanssa kyetty seuramaan MRIssä ensimmäisenä viruksena maailmassa.
Les Baculoviridae (baculovirus) sont une famille de virus en forme de bâtonnet[2], spécifiques des arthropodes, qui compte quatre genres (Alphabaculovirus, Betabaculovirus, Deltabaculovirus, Gammabaculovirus) regroupant 49 espèces.
Les baculovirus ont un tropisme spécifique pour les invertébrés. Ces virus peuvent infecter plus de 600 espèces d’insectes comme les larves des symphytes, moustiques, mais aussi de crustacés comme les crevettes. On ne connait pas de baculovirus capables d’infecter les mammifères ou d’autres vertébrés.
Le génome des baculovirus est constitué d’ADN bicaténaire, circulaire, d'une taille comprise entre 80 et 180 kpB.
Les baculovirus sont utilisés en biotechnologie pour la production de protéines recombinantes. Ces virus permettent d’introduire le gène codant la protéine à produire dans des cultures de cellules d’insectes.
Depuis les années 1940, on a beaucoup étudié les baculovirus pour leur pouvoir insecticide et on les a parfois utilisés comme biopesticides. C'est le cas, par exemple, d'Anticarsia gemmatalis Multiple Nucleopolyhedrovirus (AgMNPV, genre Alphabaculovirus), commercialisé au Brésil pour lutter contre la chenille du soja, Anticarsia gemmatalis[3].
Les Baculoviridae (baculovirus) sont une famille de virus en forme de bâtonnet, spécifiques des arthropodes, qui compte quatre genres (Alphabaculovirus, Betabaculovirus, Deltabaculovirus, Gammabaculovirus) regroupant 49 espèces.
Les baculovirus ont un tropisme spécifique pour les invertébrés. Ces virus peuvent infecter plus de 600 espèces d’insectes comme les larves des symphytes, moustiques, mais aussi de crustacés comme les crevettes. On ne connait pas de baculovirus capables d’infecter les mammifères ou d’autres vertébrés.
Le génome des baculovirus est constitué d’ADN bicaténaire, circulaire, d'une taille comprise entre 80 et 180 kpB.
Les baculovirus sont utilisés en biotechnologie pour la production de protéines recombinantes. Ces virus permettent d’introduire le gène codant la protéine à produire dans des cultures de cellules d’insectes.
Depuis les années 1940, on a beaucoup étudié les baculovirus pour leur pouvoir insecticide et on les a parfois utilisés comme biopesticides. C'est le cas, par exemple, d'Anticarsia gemmatalis Multiple Nucleopolyhedrovirus (AgMNPV, genre Alphabaculovirus), commercialisé au Brésil pour lutter contre la chenille du soja, Anticarsia gemmatalis.
Baculoviridae é unha familiade virus comunmente chamados baculovirus, que teñen como hóspedes naturais artrópodos, como insectos lepidópteros, himenópteros, dípteros e crustáceos decápodos. Actualmente existen nesta familia 66 especies, repartidas en 4 xéneros.[1][2]
Os baculovirus infectan invertebrados, e describíronse unhas 600 especies de hóspedes. As formas inmaduras (larvarias) de especies de avelaíñas son os hóspedes máis comúns, pero estes virus tamén infectan sínfitos, mosquitos, e gambas. Aínda que os baculovirus poden entrar en células de mamíferos en cultivo[3] non poden replicarse en células de vertebrados.
A partir da década de 1940 foron estudados e utilizados como biopesticidas en agricultura. Os baculovirus conteñen un xenoma de ADN bicatenario de entre 80 e 180 kbp.
As mencións máis antigas da probable presenza de baculovirus poden encontrarse xa na literatura desde inicios do século XVII en informes dunha enfermidade que afectaba os vermes da seda. A partir da década de 1940 foron moi usados e estudados como biopesticidas nos campos agrícolas. A partir da década de 1990 foron utilizados para producir proteínas eucariotas complexas en cultivos celulares de células de insectos (ver Sf21, Hi-5). Estas proteínas recombinantes foron utilizadas en investigación como vacinas tanto en humanos coma en tratamentos veterinarios (por exemplo, a vacina máis amplamente utilizada para a prevención da gripe aviaria por H5N1 en polos foi producida nun vector de expresión baculovirus). Máis recentemente atopouse que os baculovirus poden transducir células de mamíferos cun promotor axeitado.[4] Estes usos médicos aplicados ou potenciais fixeron que aumentase o número de publicacións sobre baculovirus desde 1995.
O ciclo de vida dos baculovirus implica dúas formas distitnas do virus. O virus derivado da oclusión (ODV) está presente nunha matriz proteica (poliedrina ou granulina) e é responsable da infección primaria do hóspede, mentres que o virus evaxinado (BV) libérase das células hóspede infectadas posteriormente durante a infección secundaria.
Os baculovirus teñen tropismos moi específicos de especie entre os invertebrados e describíronse unhas 700 especies hóspede. As forma inmaduras (larvas) das avelaíñas son os hóspedes máis comúns, pero estes virus infectan tamén a sínfitos, mosquitos e gambas.
Tipicamente, a infección inicial ocorre cando un insecto hóspede susceptible se alimenta de plantas que están contaminadas coa forma ocluída do virus. A matriz proteica disólvese no ambiente alcalino do estómago do hóspede, liberando ODVs que despois se fusionan coas membranas das células epiteliais columnares do intestino do hóspede e penetran na célula dentro de endosomas. As nucleocápsides escapan dos endosomas e son transportadas ao núcleo. Este paso está mediado posiblemente por filamentos de actina. A transcrición viral e a replicación ocorren no núcleo celular e evaxínanse novas partículas de BV desde a membrana basolateral para espallar a infección sistematicamente. Durante a evaxinación, os BV adquiren unha membrana da célula hóspede que está axustada de forma frouxa e contén glicoproteínas virais.
A infección por baculovirus pode ser dividida en tres fases:
Aínda que os BV se producen na fase tardía, a forma ODV prodúcese na fase moi tardía, na que adquire a envoltura do núcleo da célula hóspede e é incrustada na matriz da proteína do corpo de inclusión. Estes corpos de inclusión son liberados cando as células se lisan para espallar a infección por baculovirus aos seguintes hóspedes. A extensa lise de células causa frecuentemente que o insecto hóspede literalmente se desintegre. As partículas completas de ODV-poliedrina son resistentes á inactivación pola calor e a luz, mentres que o virión BV espido é máis sensible ás condicións do ambiente.
Cando infectan unha eiruga, os estadios avanazados da infección causan que o hóspede se alimente sen parar e despois gabee ás partes máis altas das árbores, includíndo sitios expostos que normalmente evitan debido ao risco que representan os predadores. Esta é unha vantaxe para o virus se (cando o hóspede se disolve) este pinga sobre as follas de máis abaixo que son consumidas por novos hóspedes.[5]
O baculovirus máis estudado é o Nucleopoliedrovirus multicápside de Autographa californica (AcMNPV, Autographa californica multicapsid nucleopolyhedrovirus). O virus foi illado orixinalmente do lepidóptero Autographa californica e contén un xenoma de 134-kbp con 154 marcos abertos de lectura (ORF). A principal proteína da cápside é a VP39, que xunto con algunhas proteínas menores forma a nucleocápside (21 nm x 260 nm) que encerra o ADN coa proteína p6.9.
O BV adquire a súa envoltura da membrana da célula hóspede e necesita a glicoproteína gp64 para poder espallarse e xerar unha infección sistémica. Esta proteína forma estruturas chamadas peplómeros nun dos extremos da partícula do virus evaxinado pero non se encontra no ODV (aínda que outras proteínas están só asociadas coa forma ODV). Existen tamén algunhas diferenzas na composición lipídica da envoltura viral das dúas formas. Aínda que a envoltura do BV consta de fosfatidilserina, o ODV contén fosfatidilcolina e fosfatidiletanolamina.
Identificouse un elemento esencial para a ensamblaxe da nucleocápside (NAE) no xenoma da AcMNPV. O NAE é un elemento en cis interno dentro do xene ac83. A ensamblaxe da nucleocápside non depende da produción da proteína Ac83.[6]
Durante a evolución, as glicoproteínas da envoltura baculoviral non sufriron cambios. A Ld130, tamén chamada proteína F de baculovirus de Lymantria dispar (LdMNPV) suxeriuse que é unha proteína de fusión de envoltura ancestral que foi substituída por medio dunha substitución de xene non ortólogo por gp64 no AcMNPV, Bombyx mori (BmNPV) e Orgyia pseudotsugata (OpMNPV), mentres que aínda reteñen o xene ld130.
A Gp64 é unha glicoproteína de membrana homotrimérica, que está polarmente presente sobre o virión con forma de bastón. Consta de 512 aminoácidos (aa) con catro sitios de glicosilación en residuos de asparaxina e ten unha secuencia sinal N-terminal de (20 aa), a oligomerización e o dominio de fusión e un dominio transmembrana hidrófobo preto do C-terminal (7 aa).
É producido tanto nas fases temperá coma na tardía do ciclo de infección e chega á velocidade máxima de síntese entre as 24 e 26 horas post-infección. A trimerización con enlaces disulfuro de cisteína intermoleculares parece ser un paso crucial para o transporte da proteína á superficie celular, dado que só o 33% da proteína sintetizada chega á superficie celular, xa que a gp64 monomérica é degradada dentro das células.
A gp64 é esencial para a evaxinación eficiente do virión e para a transmisión de célula a célula durante o ciclo de infección e para a entrada viral, é dicir, causando un trofismo viral e a captación pola célula mediada por endosomas. A función principal da proteína da envoltura gp64 é causar a fusión mediada polo pH da envoltura e o endosoma. Aínda que o gp64 ten varias funcións esenciais, informouse que nos baculovirus nulos para a gp64 poden substituírses esta por outras glicoproteínas virais como a Ld130, a proteína G do virus da estomatite vesicular. Estas substitucións teñen como resultado a formación de virións funcionais.
A expresión dos baculovirus en células de insectos representa un método robusto para producir glicoproteínas recombinantes ou proteínas de membrana.[7][8][9] As proteinas producidas por baculovirus están a ser estudados actualmente como vacinas terapéuticas contra o cancro con varias vantaxes inmunolóxicas sobre as proteínas derivadas de fontes de mamíferos.[10]
Os baculovirus non poden replicarse dentro de células de mamíferos ou plantas.[11] Teñen un rango de hóspedes restrinxido aos que poden infectar normalmenre limitado a un número limitado de especies de insectos estreitamente relacionadas. Como os baculoviruses non son nocivos para os humanos, son considerados unha opción segura para o seu uso nas aplicacións de investigación. Tamén se usan como axentes biolóxicos como no caso da loita contra a pirálida Plodia interpunctella, unha praga dos cereais.[12]
O nome desta familia deriva do latín baculus, que significa bastón. A familia foi dividida en catro xéneros: Alphabaculovirus (nucleopoliedrovirus específicos de lepidópteros), Betabaculovirus (granulovirus específicos de lepidópteros), Gammabaculovirus (nucleopoliedrovirus específicos de himenópteros) e Deltabaculovirus (nucleopoliedrovirus específicos de dípteros).[13] Ver a seguinte lista despregable:
Grupo: dsDNA (virus de ADN bicatenario)
Os baculovirus crese que evolucionaron a partir de virus da familia dos nudivirus (Nudiviridae) hai 310 millóns de anos.[15]
Baculoviridae é unha familiade virus comunmente chamados baculovirus, que teñen como hóspedes naturais artrópodos, como insectos lepidópteros, himenópteros, dípteros e crustáceos decápodos. Actualmente existen nesta familia 66 especies, repartidas en 4 xéneros.
Os baculovirus infectan invertebrados, e describíronse unhas 600 especies de hóspedes. As formas inmaduras (larvarias) de especies de avelaíñas son os hóspedes máis comúns, pero estes virus tamén infectan sínfitos, mosquitos, e gambas. Aínda que os baculovirus poden entrar en células de mamíferos en cultivo non poden replicarse en células de vertebrados.
A partir da década de 1940 foron estudados e utilizados como biopesticidas en agricultura. Os baculovirus conteñen un xenoma de ADN bicatenario de entre 80 e 180 kbp.
I Baculoviridae sono una famiglia di virus. Sono particelle bastoncellari provviste di involucro e di un nucleocapside di 300 nanometri x 30-60 nanometri. Ci sono 85 specie conosciute in questa famiglia, assegnate a quattro generi.
I generi di Baculoviridae sono: Alphabaculovirus, Betabaculovirus, Deltabaculovirus e Gammabaculovirus.
È noto che i baculovirus infettano gli insetti, con oltre 600 specie ospiti descritte. Le forme immature (larvali) di specie di lepidotteri (falene e farfalle) sono gli ospiti più comuni, ma sono stati trovati anche questi virus che infettano le symphyta e le zanzare. Sebbene i baculovirus siano in grado di entrare in cellule di mammifero in coltura, non sono noti per essere in grado di replicarsi in cellule di mammiferi o altri animali vertebrati .
A partire dagli anni '40 sono stati ampiamente utilizzati e studiati come biopesticidi nei campi coltivati. I baculovirus contengono un genoma circolare di DNA a doppia elica (dsDNA) che va da 80 a 180 kbp.
Le prime registrazioni di baculovirus possono essere trovate in letteratura fin dal XVI secolo nei rapporti di "malattia dell'appassimento" che infettano le larve del baco da seta. Dagli anni '90 sono stati impiegati per produrre proteine eucariotiche complesse in colture di cellule di insetto. Queste proteine ricombinanti sono state utilizzate nella ricerca e come vaccini in trattamenti medici sia umani che veterinari (ad esempio, il vaccino più utilizzato per la prevenzione, nei polli, dell'influenza aviaria causata da H5N1 è stato prodotto in un vettore di espressione del baculovirus). Più recentemente è stato scoperto che i baculovirus possono trasdurre cellule di mammifero con un promotore adatto.
Il ciclo di vita del baculovirus coinvolge due forme distinte di virus. Il virus derivato dall'occlusione (ODV) è presente in una matrice proteica (poliedrina o granulina) ed è responsabile dell'infezione primaria dell'ospite mentre il virus gemmato (BV) viene rilasciato dalle cellule ospiti infette successivamente durante l' infezione secondaria.
I baculovirus hanno tropismi specifici tra gli invertebrati con oltre 700 specie ospiti descritte. Le forme immature (larvali) delle specie di lepidotteri sono gli ospiti più comuni, ma sono stati trovati anche questi virus che infettano le symphyta e le zanzare. Segnalazioni di infezioni da baculovirus di gamberetti e coleotteri (ad esempio Oryctes rhinocerus) sono risultate essere nudiviridae, una stirpe strettamente correlata ai baculovirus.
In genere, l'infezione iniziale si verifica quando un insetto ospite suscettibile si nutre di piante contaminate dalla forma occlusa del virus. La matrice proteica si dissolve nell'ambiente alcalino dell'intestino medio dell'ospite (stomaco), rilasciando ODV che poi si fondono alla membrana delle cellule epiteliali colonnari dell'intestino dell'ospite e vengono assorbiti nella cellula negli endosomi. I nucleocapsidi sfuggono dagli endosomi e vengono trasportati al nucleo. Questo passaggio è probabilmente mediato da filamenti di actina. La trascrizione e la replicazione virale avvengono nel nucleo cellulare e nuove particelle di BV vengono espulse per diffondere l'infezione a livello sistemico. Durante il germogliamento, BV acquisisce una membrana della cellula ospite che si adatta liberamente con glicoproteine virali.
L'infezione da baculovirus può essere suddivisa in tre fasi distinte:
Mentre la BV viene prodotta nella fase tardiva, la forma ODV viene prodotta nella fase molto tardiva acquisendo l'involucro dal nucleo della cellula ospite ed incorporata nella matrice della proteina del corpo di occlusione. Questi corpi di occlusione vengono rilasciati quando le cellule si lisano per diffondere ulteriormente l'infezione da baculovirus all'ospite successivo. L'estesa lisi delle cellule spesso fa sì che l'insetto ospite si disintegra letteralmente, da qui il motivo del nome storico "malattia dell'appassimento". Le particelle complete di ODV-poliedrina sono resistenti al calore e all'inattivazione della luce, mentre il virione BV nudo è più sensibile all'ambiente.
Quando si infetta un bruco, gli stadi avanzati dell'infezione fanno sì che l'ospite si nutra senza riposarsi, per poi arrampicarsi sulle parti più alte degli alberi, compresi i luoghi esposti che normalmente eviterebbe a causa del rischio di predatori. Questo è un vantaggio per il virus se (quando l'ospite si dissolve) può gocciolare sulle foglie che verranno consumate dai nuovi ospiti.
I Baculoviridae sono una famiglia di virus. Sono particelle bastoncellari provviste di involucro e di un nucleocapside di 300 nanometri x 30-60 nanometri. Ci sono 85 specie conosciute in questa famiglia, assegnate a quattro generi.
I generi di Baculoviridae sono: Alphabaculovirus, Betabaculovirus, Deltabaculovirus e Gammabaculovirus.
È noto che i baculovirus infettano gli insetti, con oltre 600 specie ospiti descritte. Le forme immature (larvali) di specie di lepidotteri (falene e farfalle) sono gli ospiti più comuni, ma sono stati trovati anche questi virus che infettano le symphyta e le zanzare. Sebbene i baculovirus siano in grado di entrare in cellule di mammifero in coltura, non sono noti per essere in grado di replicarsi in cellule di mammiferi o altri animali vertebrati .
A partire dagli anni '40 sono stati ampiamente utilizzati e studiati come biopesticidi nei campi coltivati. I baculovirus contengono un genoma circolare di DNA a doppia elica (dsDNA) che va da 80 a 180 kbp.
Bakulowirusy mogą być wykorzystywane jako biologiczne środki do zwalczania owadzich szkodników[5]. Wiele zostało genetycznie udoskonalonych przez wprowadzenie dodatkowych genów, aby stały się skutecznymi, przyjaznymi środowisku insektycydami[2]. Wykazują zwykle dużą specyficzność, przez co jeśli ma się do czynienia z wieloma gatunkami szkodników, trzeba stosować kilka preparatów[6].
Niektóre wirusy NPV, w tym wirus wyizolowany z Autographa californica, zostały przerobione na wydajne eukariotyczne wektory ekspresyjne do użytku w inżynierii genetycznej[1]. Pozwalają na uzyskanie wielu potranslacyjnych modyfikacji białek charakterystycznych dla eukariotów, niezbędnych dla uzyskania funkcjonalnego białka, a jednocześnie są bardzo wydajne, umożliwiając produkcję porównywalnych ilości białek z wektorami opartymi na prokariotycznych systemach[8]. Ze względu na niepatogeniczność względem ssaków i nietoksyczność, brak możliwości replikacji w takich komórkach zmodyfikowane bakulowirusy są obiecującym narzędziem do zastosowania w medycynie do terapii genowej[9].
Bakulowirusy mogą być wykorzystywane jako biologiczne środki do zwalczania owadzich szkodników. Wiele zostało genetycznie udoskonalonych przez wprowadzenie dodatkowych genów, aby stały się skutecznymi, przyjaznymi środowisku insektycydami. Wykazują zwykle dużą specyficzność, przez co jeśli ma się do czynienia z wieloma gatunkami szkodników, trzeba stosować kilka preparatów.
Niektóre wirusy NPV, w tym wirus wyizolowany z Autographa californica, zostały przerobione na wydajne eukariotyczne wektory ekspresyjne do użytku w inżynierii genetycznej. Pozwalają na uzyskanie wielu potranslacyjnych modyfikacji białek charakterystycznych dla eukariotów, niezbędnych dla uzyskania funkcjonalnego białka, a jednocześnie są bardzo wydajne, umożliwiając produkcję porównywalnych ilości białek z wektorami opartymi na prokariotycznych systemach. Ze względu na niepatogeniczność względem ssaków i nietoksyczność, brak możliwości replikacji w takich komórkach zmodyfikowane bakulowirusy są obiecującym narzędziem do zastosowania w medycynie do terapii genowej.
Os baculovírus, membros da família Baculoviridae, são vírus que infectam artrópodes, majoritariamente insetos da ordem Lepidoptera, assim como insetos das ordens Diptera e Hymenoptera.[1] Por causarem infecção letal nestes animais, tais vírus são aplicados como agentes de controle biológico de insetos pragas da agricultura. Por meio de técnicas de engenharia genética, os baculovírus também são utilizados como vetores de expressão de proteínas heterólogas, e em terapia gênica.[2][3]
Baculovírus possuem DNA fita dupla, circular, superenrolado, com tamanho variando de 80 a 180 Kpb, o qual codifica entre 90 e 180 genes. O DNA viral encontra-se associado a uma estrutura protéica e a este conjunto dá-se o nome de nucleocapsídeo. O nucleocapsídeo dos baculovírus tem formato de bastão, com 200 a 450 ηm de comprimento e com 30 a 100 ηm de diâmetro. Quando envoltos por um envelope de bicamada lipídica, são denominados vírions. Adicionalmente, estes vírions são envolvidos por uma matriz protéica, formando o corpo de oclusão (OB, do inglês Occlusion Body), estrutura que confere estabilidade física e biológica às partículas virais. Quando a matriz protéica dos OB é formada por poliedrina, estes são denominados poliedros, quando por granulina, grânulos.[4][5]
A família Baculoviridae apresenta dois grupos: Nucleopolyhedrovirus (NPV) e Granulovirus (GV), designações que fazem referência às proteínas que constituem os OB. Os NPV são classificados como SNPV ou MNPV de acordo com o número de nucleocapsídeos presentes nos ODV: Single (S), quando possui apenas um, ou Multiple (M), caso possua múltiplos nucleocapsídeos por envelope.[1]
Os baculovirus apresentam dois fenótipos, Budded virus (BV) e Occlusion-derived virus (ODV), os quais apresentam estrutura de nucleocapsídeo semelhante e possuem a mesma informação genética. Os BV são responsáveis pela infecção sistêmica do hospedeiro (célula a célula), enquanto os ODV são responsáveis pela transmissão dos vírus entre insetos hospedeiros.[1][4]
Atualmente, a família Baculoviridae possui quatro gêneros: Alphabaculovirus (NPVs de Lepidoptera), Betabaculovirus (GVs de Lepidoptera), Gammabaculovirus (NPVs de Hymenoptera) e Deltabaculovirus (NPVs de Diptera).[6]
A infecção das larvas pelos vírus se dá principalmente por via oral, por ingestão da forma viral oclusa. Após a ingestão, os vírions são liberados mediante a dissolução dos corpos de oclusão nas condições alcalinas (pH>7,5) e sob ação de proteases do tubo digestivo do inseto. Em contato com as microvilosidades intestinais, os nucleocapsídeos invadem as células epiteliais, onde se multiplicam no núcleo da célula sem ainda produzirem corpos de oclusão, formando BV e caracterizando o que é conhecido como infecção primária. Os vírus extracelulares (BV), quando alcançam a hemolinfa da larva hospedeira, podem gerar uma infecção sistêmica. A partir de então ocorre a produção de grandes quantidades de nucleocapsídeos e posterior formação dos corpos de oclusão no núcleo das células dos tecidos susceptíveis da lagarta, caracterizando a infecção secundária. Os insetos mortos são importantes fontes de inóculos virais para infecções seguintes, tanto no campo quanto em laboratório.[4][7]
Os baculovírus possuem três principais aplicações tecnológicas, sendo utilizado para: controle biológico de pragas da agricultura; terapia gênica; e como vetor de expressão de proteínas heterólogas.[2]
Os insetos são responsáveis por gerar danos a produção agrícola global, gerando perdas que podem chegar a 15% da produção. A utilização de inseticidas químicos gera sérios efeitos negativos derivados de seu uso continuado: apresentam ação em organismos não alvo, favorecem o aumento da resistência dos insetos a estes produtos, poluem o ambiente, entre outras conseqüências negativas. Como alternativa ao uso dos inseticidas químicos, os baculovírus podem ser empregados como uma forma de manejo racional de pragas dos agrossistemas (principalmente da ordem Lepidoptera), sem, porém, trazer danos ao meio ambiente.[4]
O sistema baculovírus de expressão de proteínas heterólogas apresenta uma série de vantagens, entre as quais se podem destacar:
Os baculovírus, membros da família Baculoviridae, são vírus que infectam artrópodes, majoritariamente insetos da ordem Lepidoptera, assim como insetos das ordens Diptera e Hymenoptera. Por causarem infecção letal nestes animais, tais vírus são aplicados como agentes de controle biológico de insetos pragas da agricultura. Por meio de técnicas de engenharia genética, os baculovírus também são utilizados como vetores de expressão de proteínas heterólogas, e em terapia gênica.
Bakulovirüsler böcek patojeni virüsler arasında en yaygın olan ve üzerinde en çok çalışılan virüs familyasını oluşturmaktadır. Bütün bakulovirüsler aynı temel yapıya sahip olup zarflı, çubuk şekilli nükleokapsidlerin bulunduğu ve virionları içeren occlusion body’lerden meydana gelmiştir. Çubuk şekilli bu nükleokapsidlerin boyutları 40-60 x 200-400 nanometre arasında değişmektedir. Bakulovirüsler dar bir konukçu dizisine sahip olup oldukça patojenik özellik gösterirler ve sahip oldukları "occlusion Body"‘leri sayesinde diğer entomopatojen virüs familyalarına göre çevrede daha kalıcı olabilmektedirler. Bu virüslerin dairesel, çift sarmallı (80-220 kilobaz çifti) DNA'ya sahip olfuğu rapor edilmiştir.
Bakulovirüslerde virüs partikülü (virion) bir nükleik asit ve nükleik asidin çevresini kuşatan bir protein tabakasından (Kapsid) meydana gelir. Kapsid virüsün morfolojik ve fonksiyonel özelliklerini korumasını sağlar. Her bir virüs DNA'dan oluşan nükleik aside sahiptir. Nükleik asit ise çift sarmallıdır. Nükleik asidin kapsidle birlikte oluşturduğu yapıya ise "nükleokapsid" adı verilmektedir. Bakulovirüslerde nükleokapsid lipoprotein yapısında olan bir zarf ile kaplıdır. Bu zarf viral çoğalma esnasında veya virüs hücreyi terkederken ya da hücreye giriş yaparken kazanılır. Zarf virüsün hücreyi penetrasyonunda önemli rol oynar. Virionlar ise "viral occlusion" ya da "occlusion Body" olarak adlandırılan protein yapılarının içinde bulunmaktadır.
Bakulovirüsler böcek patojeni virüsler arasında en yaygın olan ve üzerinde en çok çalışılan virüs familyasını oluşturmaktadır. Bütün bakulovirüsler aynı temel yapıya sahip olup zarflı, çubuk şekilli nükleokapsidlerin bulunduğu ve virionları içeren occlusion body’lerden meydana gelmiştir. Çubuk şekilli bu nükleokapsidlerin boyutları 40-60 x 200-400 nanometre arasında değişmektedir. Bakulovirüsler dar bir konukçu dizisine sahip olup oldukça patojenik özellik gösterirler ve sahip oldukları "occlusion Body"‘leri sayesinde diğer entomopatojen virüs familyalarına göre çevrede daha kalıcı olabilmektedirler. Bu virüslerin dairesel, çift sarmallı (80-220 kilobaz çifti) DNA'ya sahip olfuğu rapor edilmiştir.
Bakulovirüslerde virüs partikülü (virion) bir nükleik asit ve nükleik asidin çevresini kuşatan bir protein tabakasından (Kapsid) meydana gelir. Kapsid virüsün morfolojik ve fonksiyonel özelliklerini korumasını sağlar. Her bir virüs DNA'dan oluşan nükleik aside sahiptir. Nükleik asit ise çift sarmallıdır. Nükleik asidin kapsidle birlikte oluşturduğu yapıya ise "nükleokapsid" adı verilmektedir. Bakulovirüslerde nükleokapsid lipoprotein yapısında olan bir zarf ile kaplıdır. Bu zarf viral çoğalma esnasında veya virüs hücreyi terkederken ya da hücreye giriş yaparken kazanılır. Zarf virüsün hücreyi penetrasyonunda önemli rol oynar. Virionlar ise "viral occlusion" ya da "occlusion Body" olarak adlandırılan protein yapılarının içinde bulunmaktadır.
Важнейшими морфологическими типами являются вирионы палочковидной, изометрической и прямоугольной формы. Дальнейшая классификация вирусов проводится по наличию или отсутствию белковых включений, содержащих вирионы. Вирусы с включениями разделяют по их генетическому материалу и месту происхождения в клетке хозяина на вирусы ядерного полиэдроза, содержащие ДНК, и вирусы цитоплазматического полиэдроза, содержащие РНК.
Вирусы цитоплазменного полиэдроза в преобладающем большинстве являются менее вирулентными и менее специфичными, чем вирусы ядерного полиэдроза и гранулеза. Вирусы ядерного полиэдроза действуют очень избирательно, являются высоковирулентными и годами не теряют своей жизнеспособности вследствие наличия инертной защитной оболочки.
По морфологии включений делят на две группы: возбудители ядерных полиэдрозов и возбудители гранулезов.
Вирионы возбудителей ядерных полиэдрозов заключены в белковые включения типа многогранников (полиэдров) с размером от 0,3 до 15 мкм. Вирионы располагаются в белковом матриксе обычно одиночно или группами. В одной группе может находиться до 20—25 вирионов, включенных в общую мембрану.
Вирусные частицы возбудителей гранулезов заключены по одному (реже по два) в гранулы овальной формы. Размеры гранул значительно меньше, чем размеры полиэдров возбудителей ядерных полиэдрозов.
Вирусные заболевания типа полиэдрозов и гранулезов известны только среди насекомых. Данные вирусные заболевания являются наиболее широко распространенными в популяциях лесных чешуекрылых и пилильщиков. Вирусы семейства Baculoviridae характеризуются крайне высокой специфичностью к своим видовым хозяевам.
Формы взаимодействия вирусов с организмом хозяина могут быть двух типов, в зависимости от продолжительности пребывания вирусных агентов в организме и популяциях насекомых.
Первый тип взаимодействия характеризуется непродолжительным пребыванием вирусов в организме хозяина и, как правило, приводит к острому инфекционному процессу с не продолжительным инкубационным периодом и последующим развитием характерных для конкретного типа возбудителя симптомов заболевания. Острая вирусная инфекция заканчивается гибелью насекомого. Из погибших насекомых вирус выходит в окружающую среду и далее распространяется в популяции вида-хозяина, заражая восприимчивых к себе особей. В биотопах вирус может долго сохраняться на протяжении месяцев и даже лет, пока вновь не попадет в организм вида-хозяина.
Второй тип взаимодействия вируса характеризуется длительным пребыванием последнего в организме и в популяции хозяина — персистенция. В данном случае вирусы находятся в неактивном состоянии — латентной форме, и передаются в популяции от родителей к потомству. Механизм подобной передачи является довольно сложным.
Латентные вирусы могут длительное время циркулировать в популяциях насекомых, переходя из поколения в поколение, пока не будут активированы какими-либо стрессовыми для вида-хозяина факторами. Данным фактором может быть аномальные отклонения погоды, чаще всего засуха, голодание или питание неподходящими кормами, инфекция другой этиологии и т. д. Обычно такое состояние наблюдается во время вспышки массового размножения. При данных ситуациях, латентная форма вируса, существовавшая в клетках хозяина, переходит в активную форму, индуцируя эпизоотический процесс.
Для каждого вида насекомого свойственен свой собственный специфичный видовой вирус. Например Lymantria dispar multiple nucleopolyhedrovirus — специфичен только непарному шелкопряду.
В наибольшей степени чувствительны к вирусу гусеницы младших возрастов. Гусеницы первого возраста заражаются даже небольшими дозами возбудителя. При попадании в кишечник гусениц полиэдры растворяются, высвобождая вирионы, которые в кишечнике распадаются на «провирусы». Они проникают сквозь стенки кишки в полость тела, а затем разносятся гемолимфой по телу и проникают в восприимчивые ткани органов. Обычно поражаются клетки жирового тела, эпителий трахей, нервные клетки, гиподерма, половые железы, придаточные железы к мальпигиевым сосудам, мышечная ткань. На протяжении не более 12 часов после инфицирования происходит внедрение вирусных частиц в клетки — частицы вируса прикрепляются к мембране ядра. После чего вирус проникает в ядро, где и происходит его рост и размножение за счет хроматина и других компонентов ядра. Уже через 24—26 часов после инфицирования отмечается гипертрофия ядер и образование вирусных палочек. Затем вирионы образуют мембраны. Часть вирионов перемещается в вирусную строму, где и происходит формирование полиэдров, в которые происходит включение созревших вирионов. Заполнив ядро, полиэдры разрывают его оболочку, выходят в цитоплазму клеток, а затем по мере их разрушения и в полость тела. Вирионы, не включившиеся в полиэдры, разносятся вместе с гемолимфой и инфицируют другие, ещё здоровые, клетки. Процесс патогенеза обычно длится 3—12 дней, и варьирует в зависимости от возраста гусениц, температуры окружающей среды. Патогенез всегда заканчивается гибелью гусениц. К этому моменту все внутренние органы и ткани лизируются, все инфицированные клетки разрушаются, превращаясь в жидкость, содержащую большое количество вирусных включений.
Внешние симптомы вирусного заболевания связаны со снижением питания, замедленностью движений, изменением окраски и покровов зараженный насекомых. В связи с репродукцией вирусов в клетках гиподермы, вызывающей её разрушение, покровы тела личинок истончаются и при даже небольших механических воздействиях разрываются с выделением гемолимфы.
Некоторые представители бакуловирусов имеют большое практическое значение, поскольку могут вызывать эпизоотии в популяциях насекомых, дающих вспышки массового размножения[5]. Среди таких насекомых можно отметить сибирского (Dendrolimus sibiricus Tschetw.), непарного (Lymantria dispar L.), кольчатого (Malacosoma neustria L.) шелкопрядов, шелкопряда-монашенку (Lymantria monacha L.), американскую белую бабочку (Hyphantria cunea Drury), капустную (Mamestra brassicae L.), сосновую (Panolis flammea Schiff.) и хлопковую (Heliothis armigera L.) совки, некоторых пилильщиков и пядениц. На основе бакуловирусов были созданы и зарегистрированы биопрепараты для контроля численности вышеперечисленных насекомых.
В Восточной Азии из энтомопатогенных вирусов насекомых наибольшее применение нашли вирусы ядерного полиэдроза, например, против гусениц Heliothls armigera, вредящих хлопчатнику. В Китае и Японии используется вирус цитоплазматического полиэдроза против вредителя сосны Dendrolimus spectabilis. В Японии зарегистрирован промышленный препарат на основе данного вируса под названием «матсукемин». Они также успешно применялись, например в борьбе с еловым пилильщиком в Канаде и сосновым шелкопрядом во Франции.
Основное затруднение в использовании вирусных препаратов — это их сложное промышленное изготовление.

Важнейшими морфологическими типами являются вирионы палочковидной, изометрической и прямоугольной формы. Дальнейшая классификация вирусов проводится по наличию или отсутствию белковых включений, содержащих вирионы. Вирусы с включениями разделяют по их генетическому материалу и месту происхождения в клетке хозяина на вирусы ядерного полиэдроза, содержащие ДНК, и вирусы цитоплазматического полиэдроза, содержащие РНК.
Вирусы цитоплазменного полиэдроза в преобладающем большинстве являются менее вирулентными и менее специфичными, чем вирусы ядерного полиэдроза и гранулеза. Вирусы ядерного полиэдроза действуют очень избирательно, являются высоковирулентными и годами не теряют своей жизнеспособности вследствие наличия инертной защитной оболочки.
По морфологии включений делят на две группы: возбудители ядерных полиэдрозов и возбудители гранулезов.
Вирионы возбудителей ядерных полиэдрозов заключены в белковые включения типа многогранников (полиэдров) с размером от 0,3 до 15 мкм. Вирионы располагаются в белковом матриксе обычно одиночно или группами. В одной группе может находиться до 20—25 вирионов, включенных в общую мембрану.
Вирусные частицы возбудителей гранулезов заключены по одному (реже по два) в гранулы овальной формы. Размеры гранул значительно меньше, чем размеры полиэдров возбудителей ядерных полиэдрозов.
棒狀病毒科(Baculoviridae),又名枝狀病毒科,是雙鏈去氧核糖核酸病毒的一種。

本文参照
バキュロウイルス科(Baculoviridae)はDNAウイルスの科。Nucleopolyhedrovirus(NPV:核多角体病ウイルス)とGranulovirus(GV:顆粒病ウイルス)の2属に分けられ、一般にバキュロウイルス(Baculovirus)と呼ばれる。バキュロウイルスは節足動物に感染し、宿主に対する種特異性が高い。大部分はチョウ目の幼虫に感染するが、ハチ、カ、エビに感染するものも知られている。脊椎動物には感染・増殖しない。特定の宿主にしか感染しないが致死性が高く、他の動物には安全なので、生物農薬として利用されるものもある。
ビリオンは大型の棒状。エンベロープを持ち、GVはエンベロープ1個にヌクレオカプシドを1個だけ含むが、NPVにはエンベロープ1個にヌクレオカプシドを複数含むものもある。さらにビリオンはタンパク質(GVではグラニュリン、NPVではポリヘドリン)の結晶(包埋体)に包まれる。GVの包埋体(顆粒体)は小型でビリオンを1個しか含まないが、NPVの包埋体(核多角体)は大型で多数のビリオンを含む。ゲノムは環状二本鎖DNAで、80から180kbpと大きく、遺伝子数も100以上ある。
包埋体は虫に食べられたあと、中腸でアルカリ性のため溶解し、感染する。細胞内で複製されたウイルスはまず細胞表面から出芽し、他の細胞に感染して全身感染に至る。次に各細胞内で包埋体が作られる。包埋体型ウイルスは出芽型ウイルスよりも安定性、感染性ともに高い。虫は死んで虫体内部は液状化し、その後皮膚が破れると内部に充満したウイルスがまき散らされ、他の虫に食べられて感染する。
ウイルスの分類(ボルティモア分類) DNAバキュロウイルス科(Baculoviridae)はDNAウイルスの科。Nucleopolyhedrovirus(NPV:核多角体病ウイルス)とGranulovirus(GV:顆粒病ウイルス)の2属に分けられ、一般にバキュロウイルス(Baculovirus)と呼ばれる。バキュロウイルスは節足動物に感染し、宿主に対する種特異性が高い。大部分はチョウ目の幼虫に感染するが、ハチ、カ、エビに感染するものも知られている。脊椎動物には感染・増殖しない。特定の宿主にしか感染しないが致死性が高く、他の動物には安全なので、生物農薬として利用されるものもある。