nomes no trilho de navegação

Die Trypanosomatida sind eine Gruppe von einzelligen Flagellaten innerhalb der Gruppe der Kinetoplastea, die sich durch eine einzelne Geißel auszeichnen und die durchweg als Parasiten leben. Viele Trypanosomatida vermehren sich ausschließlich in Insekten; manche Gattungen machen einen Wirtswechsel zwischen Insekten als Vektor und einem Wirbeltierwirt oder einem Pflanzenwirt durch.
Zu den Trypanosomatida gehören mit den Leishmanien und den Trypanosomen wichtige Krankheitserreger für die Leishmaniose, die Chagas-Krankheit und die afrikanische Trypanosomiasis beim Menschen sowie für die Tierseuchen Nagana und Surra. Auch einige Pflanzenschädlinge der Gattung Phytomonas zählen zu den Trypanosomatida.
Alle Trypanosomatida haben eine einzelne Geißel, die soweit reduziert sein kann, dass sie im Lichtmikroskop nicht sichtbar ist (amastigote Form). Ferner findet sich in einem einzelnen großen Mitochondrium ein Kinetoplast, eine durch Anfärbung sichtbare Ansammlung von mitochondrialer DNA. Viele Arten zeigen einen Formenwechsel, bei dem sich die Zellform im Laufe des Lebenszyklus deutlich ändert. Das trifft sowohl für die Zellgröße wie auch für die Position der Geißelbasis (trypomastigot, epimastigot, promastigot) relativ zum Zellende zu.
Vertreter der Trypanosomatida sind weltweit anzutreffen. Alle Arten leben als Parasiten. Manche Gattungen parasitieren nur in einer Klasse von Lebewesen, meist Insekten; darunter fallen die Gattungen Blastocrithidia, Crithidia, Herpetomonas, Leptomonas und Rhynchoidomonas. Die Gattung Phytomonas macht einen Wirtswechsel zwischen Insekten als Vektor und Pflanzen durch, während Endotrypanum, Leishmania und Trypanosoma zwischen Insekten (bei aquatischen Trypanosomen auch Egeln) und Wirbeltieren wechseln.
Die erste Beschreibung einer Ordnung noch unter der Bezeichnung Trypanosomata, die auf der Typusgattung Trypanosoma basierte, stammt vom britischen Meeresbiologen William Saville-Kent (1845–1908).[1]
Die Trypanosomatida gehören mit den Neobodonida, den Parabodonida und den Eubodonida zu den Metakinetoplastina, einer aufgrund von Sequenzvergleichen von ribosomaler RNA definierten Gruppe innerhalb der Kinetoplastea.[2] Im Gegensatz zu den anderen Vertretern sind die Trypanosomatida ausschließlich Parasiten mit nur einer einzelnen Geißel. Innerhalb der Trypanosomatida werden derzeit elf Gattungen unterschieden.
Während die Trypanosomatida selbst sowie die Gattungen Leishmania, Phytomonas und Trypanosoma als monophyletisch gelten, sind die anderen Gattungen vermutlich paraphyletisch.[3] Die Vermehrung ausschließlich in Insekten gilt als ursprünglich; der Wirtswechsel zwischen Insekten und Wirbeltieren ist innerhalb der Trypanosomatida vermutlich mehrmals entstanden.
Die Trypanosomatidae als Familie wurden etwas weiter gefasst:
Die Trypanosomatida sind eine Gruppe von einzelligen Flagellaten innerhalb der Gruppe der Kinetoplastea, die sich durch eine einzelne Geißel auszeichnen und die durchweg als Parasiten leben. Viele Trypanosomatida vermehren sich ausschließlich in Insekten; manche Gattungen machen einen Wirtswechsel zwischen Insekten als Vektor und einem Wirbeltierwirt oder einem Pflanzenwirt durch.
Zu den Trypanosomatida gehören mit den Leishmanien und den Trypanosomen wichtige Krankheitserreger für die Leishmaniose, die Chagas-Krankheit und die afrikanische Trypanosomiasis beim Menschen sowie für die Tierseuchen Nagana und Surra. Auch einige Pflanzenschädlinge der Gattung Phytomonas zählen zu den Trypanosomatida.
Trypanosomatida is a group of kinetoplastid unicellular organisms distinguished by having only a single flagellum. The name is derived from the Greek trypano (borer) and soma (body) because of the corkscrew-like motion of some trypanosomatid species. All members are exclusively parasitic, found primarily in insects.[1] A few genera have life-cycles involving a secondary host, which may be a vertebrate, invertebrate or plant. These include several species that cause major diseases in humans.[2] Some trypanosomatida are intracellular parasites, with the important exception of Trypanosoma brucei.
The three major human diseases caused by trypanosomatids are; African trypanosomiasis (sleeping sickness, caused by Trypanosoma brucei and transmitted by tsetse flies), South American trypanosomiasis (Chagas disease, caused by T. cruzi and transmitted by triatomine bugs), and leishmaniasis (a set of trypanosomal diseases caused by various species of Leishmania transmitted by sandflies).
The family is known from fossils of the extinct genus Paleoleishmania preserved in Burmese amber dating to the Albian (100 mya) and Dominican amber from the Burdigalian (20–15 mya) of Hispaniola.[3] The genus Trypanosoma is also represented in Dominican amber in the extinct species T. antiquus.[4]
Three genera are dixenous (two hosts in the life cycle) – Leishmania, Phytomonas and Trypanosoma and the remainder are monoxenous (one host in the life cycle). Paratrypanosoma appears to be the first evolving branch in this order. Fifteen genera are recognised in the Trypanosomatidae and there are three subfamilies – Blechomonadinae, Leishmaniinae and Strigomonadinae. The genera in the subfamily Strigomonadinae are characterised by the presence of obligatory intracellular bacteria of the Kinetoplastibacterium genus.
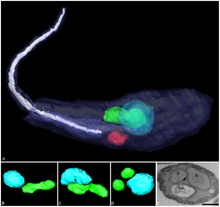
Some trypanosomatids only occupy a single host, while many others are heteroxenous: they live in more than one host species over their life cycle. This heteroxenous life cycle typically includes the intestine of a bloodsucking insect and the blood and/or tissues of a vertebrate. Rarer hosts include other bloodsucking invertebrates, such as leeches,[5] and other organisms such as plants. Different species go through a range of different morphologies at different stages of the life cycle, with most having at least two different morphologies. Typically the promastigote and epimastigote forms are found in insect hosts, trypomastigote forms in the mammalian bloodstream and amastigotes in intracellular environments.
Among commonly studied examples, T. brucei, T. congolense, and T. vivax are extracellular, while T. cruzi and Leishmania spp. are intracellular.[6] Trypanosomatids with intracellular stages express δ-amastin proteins on their surfaces.[6] de Paiva et al., 2015 illuminates δ-amastins' roles in intracellular success.[6]
Trypanosomatids that cause globally known diseases such leishmaniasis (Leishmania species), African trypanosomiasis referred to as sleeping sickness (Trypanosoma brucei), and Chagas disease (Trypanosoma cruzi) were found to be capable of meiosis and genetic exchange.[7] These findings indicate the capability for sexual reproduction in the Trypanosomatida.[7]

A variety of different morphological forms appear in the life cycles of trypanosomatids, distinguished mainly by the position, length and the cell body attachment of the flagellum. The kinetoplast is found closely associated with the basal body at the base of the flagellum and all species of trypanosomatid have a single nucleus. Most of these morphologies can be found as a life cycle stage in all trypanosomatid genera however certain morphologies are particularly common in a particular genus. The various morphologies were originally named from the genus where the morphology was commonly found, although this terminology is now rarely used because of potential confusion between morphologies and genus. Modern terminology is derived from the Greek; "mastig", meaning whip (referring to the flagellum), and a prefix which indicates the location of the flagellum on the cell. For example, the amastigote (prefix "a-", meaning no flagellum) form is also known as the leishmanial form as all Leishmania have an amastigote life cycle stage.

Amastigote: False colour SEM micrograph of amastigote form Leishmania mexicana. The cell body is shown in orange and the flagellum is in red. 219 pixels/μm.

Promastigote: False colour SEM micrograph of promastigote form Leishmania mexicana. The cell body is shown in orange and the flagellum is in red. 119 pixels/μm.

Trypomastigote: False colour SEM micrograph of procyclic form Trypanosoma brucei. The cell body is shown in orange and the flagellum is in red. 84 pixels/μm.
Notable characteristics of trypanosomatids are the ability to perform trans-splicing of RNA and possession of glycosomes, where much of their glycolysis is confined to. The acidocalcisome, another organelle, was first identified in trypanosomes.[10]
A grand total of six species of trypanosomatids carry an additional proteobacterial endosymbioant, termed TPE (trypanosomatid proteobacterial endosymbionts). These trypansomatids (Strigomonas oncopelti, S. culicis, S. galati, Angomonas desouzai, and A. deanei) are in turn known as SHTs, for symbiont-harboring trypanosomatids. All such symbionts have a shared evolutionary origin and are classified in the Candidatus genus "Kinetoplastibacterium".[11]
As with many symbionts, the bacteria have a much reduced genome compared to their free-living relatives of genera Taylorella and Achromobacter. (GTDB finds the genus sister to Proftella, a symbiont of Diaphorina citri.)[12] Reflecting their inability to live alone, they have lost genes dedicated to essential biological functions, relying on the host instead. They have modified their division to become synchronized with the host. In S. culicis at least, the TPE helps the host by synthesizing heme[11] and producing essential enzymes, staying tethered to the kinetoplast.[13]
{{cite journal}}: CS1 maint: multiple names: authors list (link) {{cite journal}}: CS1 maint: multiple names: authors list (link) {{cite journal}}: CS1 maint: multiple names: authors list (link)
{{cite journal}}: CS1 maint: multiple names: authors list (link)
{{cite journal}}: CS1 maint: multiple names: authors list (link)
{{cite journal}}: CS1 maint: multiple names: authors list (link)
Trypanosomatida is a group of kinetoplastid unicellular organisms distinguished by having only a single flagellum. The name is derived from the Greek trypano (borer) and soma (body) because of the corkscrew-like motion of some trypanosomatid species. All members are exclusively parasitic, found primarily in insects. A few genera have life-cycles involving a secondary host, which may be a vertebrate, invertebrate or plant. These include several species that cause major diseases in humans. Some trypanosomatida are intracellular parasites, with the important exception of Trypanosoma brucei.
Los tripanosomátidos (Trypanosomatida) son un orden de protistas del filo Kinetoplastea[2] que se caracterizan por tener un solo flagelo. Todos sus miembros son exclusivamente parásitos, de animales, plantas y otros protozoos. Algunos géneros tienen ciclos vitales que implican un huésped secundario, que puede ser alternativamente un vertebrado o una planta. Se incluyen varias especies que causan enfermedades graves en seres humanos, entre las que destacan:
Los tripanosomátidos presentan un único flagelo que emerge de un bolsillo anterior o bien está insertado lateralmente y adherido a la célula a lo largo de toda su longitud. Son osmotrofos con un citostoma, si es que está presente, se localiza próximo a al bolsillo flagelar y carece de estructuras orales asociadas. El cinetoplasto es de tipo eucinetoplasto, con círculos de ADN encadenados formando una red.[4]
En el ciclo vital de estos organismos se presentan diversas formas que se distinguen principalmente por la posición del flagelo:[5]
Todos las especies de Trypanosomatida tienen por lo menos etapas amastigote y promastigote. Trypanosoma aparece en las cinco formas; la etapa tripanosomial tiene lugar en un huésped vertebrado.[6] Las subespecies de Trypanosoma brucei tienen dos formas en la circulación sanguínea de un huésped vertebrado: la forma larga y delgada que se divide rápidamente y la forma corta que no se divide. La forma corta se adapta al vector de la mosca tse-tse y es no proliferativa en comparación con la forma delgada.
Trypanosoma brucei presenta la peculiaridad única de la expresión de una capa de glicoproteína variante de superficie (VSG) sobre la superficie de la célula, que experimenta una variación constante para evadir los anticuerpos del sistema inmunitario del huésped. Se supone que la recombinación de un repertorio de genes VSG en número mayor que 1000 es la responsable de la gran diversidad del parásito y su eficacia en la evasión al sistema inmunitario.[7]
Los tripanosomátidos (Trypanosomatida) son un orden de protistas del filo Kinetoplastea que se caracterizan por tener un solo flagelo. Todos sus miembros son exclusivamente parásitos, de animales, plantas y otros protozoos. Algunos géneros tienen ciclos vitales que implican un huésped secundario, que puede ser alternativamente un vertebrado o una planta. Se incluyen varias especies que causan enfermedades graves en seres humanos, entre las que destacan:
Tripanosomiasis (enfermedad del sueño en África y enfermedad de Chagas en América del Sur), causada por especies del género Trypanosoma. Leishmaniasis, causada por especies del género Leishmania.В жизненных циклах трипаносоматид происходит смена различных морфологических форм, которые в основном различаются по расположению, длине и месту прикрепления жгутика. Большинство из этих форм можно наблюдать в качестве стадий жизненного цикла у представителей различных родов трипаносоматид, но некоторые особенно характерны для отдельных конкретных родов. Первоначально названия отдельным морфологическим формам давались по имени рода, для которого данные формы были типичны, но современные названия (восходящие к статье К. А. Хоара и Ф. Уоллеса, 1966[7]) унифицированы: в их основе лежит корень мастиг (от греч. μάστιξ — жгутик), дополняемый той или иной приставкой. В указанной статье было выделено 7 морфологических форм трипаносоматид[8][9]:
В жизненных циклах трипаносоматид происходит смена различных морфологических форм, которые в основном различаются по расположению, длине и месту прикрепления жгутика. Большинство из этих форм можно наблюдать в качестве стадий жизненного цикла у представителей различных родов трипаносоматид, но некоторые особенно характерны для отдельных конкретных родов. Первоначально названия отдельным морфологическим формам давались по имени рода, для которого данные формы были типичны, но современные названия (восходящие к статье К. А. Хоара и Ф. Уоллеса, 1966) унифицированы: в их основе лежит корень мастиг (от греч. μάστιξ — жгутик), дополняемый той или иной приставкой. В указанной статье было выделено 7 морфологических форм трипаносоматид:
Амастиготы («лейшманиальные формы») — мелкие клетки округлой или эллипсоидальной формы, лишённые жгутика полностью или имеющие его рудимент, не выходящий за пределы клетки (второй подтип амастигот иногда выделяют в качестве самостоятельной морфологической формы под названием эндомастиготы). Сферомастиготы (округлые «лейшманиальные формы») — мелкие клетки округлой формы с небольшим свободным жгутиком (переходная форма между амастиготами и мастиготами). Промастиготы («лептомонадные формы») — клетки удлинённой формы со жгутиком на переднем конце клетки, у которых кинетопласт лежит перед ядром. Хоаномастиготы — клетки, по строению близкие клеткам промастигот, но отличающиеся кувшинообразной формой, причём основание жгутика заключено в воронкообразный резервуар (редко встречающаяся форма, отмеченная у представителей рода Crithidia). Эпимастиготы («критидиальные формы») — клетки удлинённой формы со жгутиком, отходящим от середины клетки, у которых кинетопласт располагается рядом с ядром, а прилегающая к поверхности клетки часть жгутика образует ундулирующую мембрану (способствующую извивающемуся движению всего организма жгутиконосца). Трипомастиготы («трипаносомные формы») — клетки удлинённой формы со жгутиком, отходящим от заднего конца клетки, у которых кинетопласт располагается за ядром, а прилегающая к поверхности клетки часть жгутика образует длинную ундулирующую мембрану. Опистомастиготы — клетки, по строению близкие клеткам трипомастигот, но отличающиеся тем, что жгутик не образует ундулирующей мембраны, а проходит на большей части своего протяжения под поверхностью клетки (редко встречающаяся форма, отмеченная только у представителей рода Herpetomonas)._Trypanosoma_equiperdum.jpg)