Glucagon-like peptide-1 (GLP-1) is an incretin derived from the transcription product of the proglucagon gene. The major source of GLP-1 in the body is the intestinal L cell that secretes GLP-1 as a gut hormone. Its physiological functions include promoting insulin sensitivity, decreasing food intake by increasing satiety in brain and increasing insulin secretion from the pancreas in a glucose-dependent manner.
GLP-1 (7-36) amide Acetate augments glucose-induced insulin secretion and simultaneously inhibits glucagon secretion and gastric emptying.
GLP-1 (7-36) amide Acetate augments glucose-induced insulin secretion and simultaneously inhibits glucagon secretion and gastric emptying.
Papillomaviridae és una família de virus ADN sense cobertura viral antiga i diversa. Alguns centenars d'espècies de papil·lomavirus, tradicionalment coneguts com a "tipus",[1] s’han identificat com infestants de tots els ocells i mamífers inspeccionats acuradament.[1] i també un petit grup de rèptils, en particular tortugues.[2][3] la infecció per a la majoria dels tipus de papil·lomavirus, és o bé sense símptomes o causa petits tumors benignes coneguts com a papil·loma. Els papil·lomes causat per determinats tipus però poden produir càncer.[4]
Els papil·lomavirus es transmeten rarament entre espècies.[5] Els papil·lomavirus repliquen exclusivament en Stratum germinativum (capa basal) de l’epiteli esquamós estratificat (teixits de la superfície del cos). Tots els papil·lomavirus coneguts infecten una superfície del cos particular,[1] típicament la pell o els genitals (en l’epiteli mucós), anus, boca, o vies aèries.[6] Per exemple el papil·lovirus humà tipus 1 (HPV) type 1 tendeix a infectar la sola dels peus, i el tipus 2 HPV type 2 les palmes de la mà, on pot causar papil·loma.
Els papil·lomavirus van ser identificats el segle XX. El 1935 Francis Peyton Rous, comprovà càncer per papil·lomes en conills. Aquesta va ser la primera demostració que un virus podia produir càncer en animals mamífers.
Els papil·lomavirus (PVs) tenen similar organització genòmica, i cada parell de PVs conté com a mínim cinc gens homòlegs, malgrat que la seqüència de nucleòtids pot divergir en més de 50%. Els estudis filogenètics suggereixen que els PVs normalment evolucionen junt amb el seu hoste (ocell o mamífer), no es recombina, i han mantingut la seva organització genòmica bàsica per més de 100 milions d’anys.
Els papil·lomavirus no tenen coberta cosa que vol dir que la capa exterior de la càpsida del virus no està coberta per una membrana cel·lular lipídica. La càpside presenta simetria icosahèdrica..
El genoma dels papil·lomavirus fa 8.000 parells de base de llargada.
Els papil·lomavirus infecten les cèl·lules per petites ferides anomenades microtraumes, de la pell o mucosa.[7][8] El virus és capaç d’entrar dins la cèl·lula via la interacció amb un receptor específic,[9][10] i transportat a la membrana dins de vesícules.[11][12] La proteïna càpsida L2 trenca la membrana de la vesícula i deixa el genoma viral escapar i anar al nucli de la cèl·lula.[13][14]
Papillomaviridae és una família de virus ADN sense cobertura viral antiga i diversa. Alguns centenars d'espècies de papil·lomavirus, tradicionalment coneguts com a "tipus", s’han identificat com infestants de tots els ocells i mamífers inspeccionats acuradament. i també un petit grup de rèptils, en particular tortugues. la infecció per a la majoria dels tipus de papil·lomavirus, és o bé sense símptomes o causa petits tumors benignes coneguts com a papil·loma. Els papil·lomes causat per determinats tipus però poden produir càncer.
Els papil·lomavirus es transmeten rarament entre espècies. Els papil·lomavirus repliquen exclusivament en Stratum germinativum (capa basal) de l’epiteli esquamós estratificat (teixits de la superfície del cos). Tots els papil·lomavirus coneguts infecten una superfície del cos particular, típicament la pell o els genitals (en l’epiteli mucós), anus, boca, o vies aèries. Per exemple el papil·lovirus humà tipus 1 (HPV) type 1 tendeix a infectar la sola dels peus, i el tipus 2 HPV type 2 les palmes de la mà, on pot causar papil·loma.
Els papil·lomavirus van ser identificats el segle XX. El 1935 Francis Peyton Rous, comprovà càncer per papil·lomes en conills. Aquesta va ser la primera demostració que un virus podia produir càncer en animals mamífers.
Papillomaviridae er en virusfamilie der tilhører gruppen af dobbeltstrenget DNA-virus (gruppe I, dsDNA).
Human papilloma virus (HPV) findes i over hundrede forskellige arter. De kan bl.a være årsag til vorter, kønsvorter og livmoderhalskræft.
Da HPV kan forårsage kræft betegnes den som en oncogen virus eller oncovirus.
Papillomaviridae familien har 16 slægter
Papillomaviridae er en virusfamilie der tilhører gruppen af dobbeltstrenget DNA-virus (gruppe I, dsDNA).
Human papilloma virus (HPV) findes i over hundrede forskellige arter. De kan bl.a være årsag til vorter, kønsvorter og livmoderhalskræft.
Da HPV kan forårsage kræft betegnes den som en oncogen virus eller oncovirus.
Die Virusfamilie Papillomaviridae umfasst 16 Gattungen von unbehüllten Viren mit einer doppelsträngigen, zirkulären DNA als Genom. Bis 2002 bildete die Familie Papillomaviridae als Gattung Papillomavirus zusammen mit der jetzt eigenen Familie Polyomaviridae die Familie Papovaviridae. 1964 hatte Joseph Melnick aufgrund der morphologischen und genetischen Ähnlichkeiten diese heute obsolete Virusfamilie vorgeschlagen. Die Vermehrungsstrategie, das Spektrum der Erkrankungen und die Anordnung der Gene sind jedoch so unterschiedlich, dass man sie in zwei verschiedene Familien unterteilte. Im März 2020 hat das International Committee on Taxonomy of Viruses (ICV) diese ehemaligen Familie Papovaviridae wieder als Klasse Papovaviricetes in die offizielle Taxonomie eingeführt, unter der die beiden Teilfamilien wieder vereint sind.[2]
Papillomviren verursachen bei sehr vielen verschiedenen Wirbeltieren Hautwucherungen im Sinne von Warzen (Papillome). Das erste als Virus erkannte Agens aus dieser Gruppe wurde von Richard Shope 1932 bei erkrankten Kaninchen isoliert (Shopes Kaninchenpapillomvirus). Die Entstehung bösartiger Tumoren aus diesen gutartigen Wucherungen wurde bereits damals beobachtet. Heute ist die Entstehung verschiedener Krebsarten bei Menschen und Tieren mit Vertretern der Papillomaviridae verbunden. Beim Menschen ist dies vorwiegend das Cervixkarzinom durch Infektionen mit Humanen Papillomviren. Die Vermehrung von Papillomviren ist strikt auf Epithelien beschränkt und sie bedürfen zur Vermehrung Zellen, die sich in einer Phase der Entdifferenzierung befinden. Aufgrund dieses strengen Gewebstropismus ist bislang eine vollständige Vermehrung der Papillomaviridae in Zellkulturen nicht möglich.
2013 wurden Hybride beschrieben zwischen Papillomviren und Polyomaviren.[3]
Die etwa 55 nm großen, unbehüllten Virionen (Viruspartikel) der Papillomviren bestehen aus einem ikosaedrischen Kapsid (T=7), das aus 72 Kapsomeren aufgebaut ist.[4] Von den Kapsomeren sind 60 in einer fünfstrahligen Symmetrie (Pentamere) und diese wiederum in 12 sechsstrahligen (Hexamere) angeordnet.[5] Zwischen den Pentameren bestehen Disulfidbrücken. Bei fehlerhafter Reifung und Zusammenbau können auch filamentöse Kapside beobachtet werden. Die Kapsomere werden von zwei Strukturproteinen (L1 und L2) gebildet. Das L1 (55 bis 60 kDa) ist mit über 80 % Gewichtsanteil dominierend. Es besitzt nach außen gerichtete, speziesspezifische Epitope. Das L2 (50 bis 53 kDa) besitzt gruppenspezifische Epitope. Die Virionen besitzen eine hohe Umweltstabilität; milde Detergenzien (Seife), saure pH-Werte und zur Virusinaktivierung üblicherweise verwendete fettlösende Substanzen (beispielsweise 2-Propanol) inaktivieren Papillomviren nicht. Sie sind für 1 Stunde bei 50 °C hitzestabil.
Im Inneren der Kapside befindet sich die zirkuläre DNA des Virusgenoms. Diese ist mehrfach verdrillt („supercoiled“) und bildet zusammen mit zellulären Histonen einen Nukleoproteinkomplex, der den eukaryotischen Nukleosomen strukturell sehr ähnelt. Von den fünf bekannten Histonen findet man die Histone H2a, H2b, H3 und H4.
Das Genom der Papillomviren ist etwa 6800 bis 8400 bp groß und codiert mit 9 bis 10 Offene Leserahmen (ORF) für die frühen Virusproteine (early: E1-E8) und die späten Strukturproteine (late: L1 und L2). Bei einigen Spezies fehlen E3- oder E8-Leserahmen. Im Gegensatz zu den Polyomaviridae ist die Leserichtung für alle ORFs gleich. Vor den Genen für die frühen Virusproteine befindet sich eine regulatorische Region (LCR, long control region), die Steuerungssequenzen (Promotoren, Enhancer und Replikationsursprung) enthalten. Die Leserahmen überlappen sich teilweise und liegen in verschiedenen Leserastern. Die im Zellkern durch zelluläre RNA-Polymerasen transkribierten viralen mRNAs unterliegen weiteren Modifizierungen wie einem Capping, einer Polyadenylierung und einem RNA-Splicing. Zur Vermehrung der genomischen DNA der Papillomaviridae besitzen die Viren keine eigene DNA-Polymerase. Sie sind bei der Virusreplikation darauf angewiesen, dass ausreichende Mengen zellulärer DNA-Polymerasen vorhanden und aktiv sind. Da dies besonders während der Zellteilung der Fall ist, haben die Papillomaviridae verschiedene Mechanismen entwickelt, um die Wirtszelle zur ständigen Teilung anzuregen, durch E6 und E7. Diese Mechanismen sind die Ursache für die typischen Gewebswucherungen bei Infektionen mit Papillomviren und eine mögliche Entartung der Zellen.
Die innere Systematik der Papillomaviridae ist nach Maßgabe des ICTV, Stand November 2018, wie folgt;[6] Gattungen ohne aufgeführte Spezies haben nur eine einzige (d. h. sie sind monotypisch):
An diesem Benennungsschema hat sich auch bis April 2020 (ICTV MSL #35) nichts Wesentlichen geändert, es liegt inzwischen jedoch ein Vorschlag an das ICTV zur Umbenennung vor.[26]
Die Virusfamilie Papillomaviridae umfasst 16 Gattungen von unbehüllten Viren mit einer doppelsträngigen, zirkulären DNA als Genom. Bis 2002 bildete die Familie Papillomaviridae als Gattung Papillomavirus zusammen mit der jetzt eigenen Familie Polyomaviridae die Familie Papovaviridae. 1964 hatte Joseph Melnick aufgrund der morphologischen und genetischen Ähnlichkeiten diese heute obsolete Virusfamilie vorgeschlagen. Die Vermehrungsstrategie, das Spektrum der Erkrankungen und die Anordnung der Gene sind jedoch so unterschiedlich, dass man sie in zwei verschiedene Familien unterteilte. Im März 2020 hat das International Committee on Taxonomy of Viruses (ICV) diese ehemaligen Familie Papovaviridae wieder als Klasse Papovaviricetes in die offizielle Taxonomie eingeführt, unter der die beiden Teilfamilien wieder vereint sind.
Papillomviren verursachen bei sehr vielen verschiedenen Wirbeltieren Hautwucherungen im Sinne von Warzen (Papillome). Das erste als Virus erkannte Agens aus dieser Gruppe wurde von Richard Shope 1932 bei erkrankten Kaninchen isoliert (Shopes Kaninchenpapillomvirus). Die Entstehung bösartiger Tumoren aus diesen gutartigen Wucherungen wurde bereits damals beobachtet. Heute ist die Entstehung verschiedener Krebsarten bei Menschen und Tieren mit Vertretern der Papillomaviridae verbunden. Beim Menschen ist dies vorwiegend das Cervixkarzinom durch Infektionen mit Humanen Papillomviren. Die Vermehrung von Papillomviren ist strikt auf Epithelien beschränkt und sie bedürfen zur Vermehrung Zellen, die sich in einer Phase der Entdifferenzierung befinden. Aufgrund dieses strengen Gewebstropismus ist bislang eine vollständige Vermehrung der Papillomaviridae in Zellkulturen nicht möglich.
2013 wurden Hybride beschrieben zwischen Papillomviren und Polyomaviren.
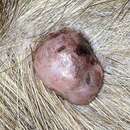
Papillomaviridae is a family of non-enveloped DNA viruses whose members are known as papillomaviruses.[1] Several hundred species of papillomaviruses, traditionally referred to as "types",[2] have been identified infecting all carefully inspected mammals,[2] but also other vertebrates such as birds, snakes, turtles and fish.[3][4][5] Infection by most papillomavirus types, depending on the type, is either asymptomatic (e.g. most Beta-PVs) or causes small benign tumors, known as papillomas or warts (e.g. human papillomavirus 1, HPV6 or HPV11). Papillomas caused by some types, however, such as human papillomaviruses 16 and 18, carry a risk of becoming cancerous.[6]
Papillomaviruses are usually considered as highly host- and tissue-tropic, and are thought to rarely be transmitted between species.[7] Papillomaviruses replicate exclusively in the basal layer of the body surface tissues. All known papillomavirus types infect a particular body surface,[2] typically the skin or mucosal epithelium of the genitals, anus, mouth, or airways.[8] For example, human papillomavirus (HPV) type 1 tends to infect the soles of the feet, and HPV type 2 the palms of the hands, where they may cause warts. Additionally, there are descriptions of the presence of papillomavirus DNA in the blood and in the peripheral blood mononuclear cells.
Papillomaviruses were first identified in the early 20th century, when it was shown that skin warts, or papillomas, could be transmitted between individuals by a filterable infectious agent. In 1935 Francis Peyton Rous, who had previously demonstrated the existence of a cancer-causing sarcoma virus in chickens, went on to show that a papillomavirus could cause skin cancer in infected rabbits. This was the first demonstration that a virus could cause cancer in mammals.
There are over 100 species of papillomavirus recognised,[9] though the ICTV officially recognizes a smaller number, categorized into 53 genera, as of 2019.[10][11][12] All papillomaviruses (PVs) have similar genomic organizations, and any pair of PVs contains at least five homologous genes, although the nucleotide sequence may diverge by more than 50%. Phylogenetic algorithms that permit the comparison of homologies led to phylogenetic trees that have a similar topology, independent of the gene analyzed.[13]
Phylogenetic studies strongly suggest that PVs normally evolve together with their mammalian and bird host species, but adaptive radiations, occasional zoonotic events and recombinations may also impact their diversification.[13] Their basic genomic organization appears maintained for a period exceeding 100 million years, and these sequence comparisons have laid the foundation for a PV taxonomy, which is now officially recognized by the International Committee on Taxonomy of Viruses. All PVs form the family Papillomaviridae, which is distinct from the Polyomaviridae thus eliminating the term Papovaviridae. Major branches of the phylogenetic tree of PVs are considered genera, which are identified by Greek letters. Minor branches are considered species and unite PV types that are genomically distinct without exhibiting known biological differences. This new taxonomic system does not affect the traditional identification and characterization of PV "types" and their independent isolates with minor genomic differences, referred to as "subtypes" and "variants", all of which are taxa below the level of "species".[14] Additionally, phylogenetic groupings at higher taxonomic level have been proposed.[15]
This classification may need revision in the light of the existence of papilloma–polyoma virus recombinants.[16] Additional species have also been described. Sparus aurata papillomavirus 1 has been isolated from fish.[17]
Over 170 human papillomavirus types have been completely sequenced.[18] They have been divided into 5 genera: Alphapapillomavirus, Betapapillomavirus, Gammapapillomavirus, Mupapillomavirus and Nupapillomavirus. At least 200 additional viruses have been identified that await sequencing and classification.
Individual papillomavirus types tend to be highly adapted to replication in a single animal species. In one study, researchers swabbed the forehead skin of a variety of zoo animals and used PCR to amplify any papillomavirus DNA that might be present.[19] Although a wide variety of papillomavirus sequences were identified in the study, the authors found little evidence for inter-species transmission. One zookeeper was found to be transiently positive for a chimpanzee-specific papillomavirus sequence. However, the authors note that the chimpanzee-specific papillomavirus sequence could have been the result of surface contamination of the zookeeper's skin, as opposed to productive infection.
Cottontail rabbit papillomavirus (CRPV) can cause protuberant warts in its native host, the North American rabbit genus Sylvilagus. These horn-like warts may be the original basis for the urban legends of the American antlered rabbit the Jackalope and European Wolpertinger.[20] European domestic rabbits (genus Oryctolagus) can be transiently infected with CRPV in a laboratory setting. However, since European domestic rabbits do not produce infectious progeny virus, they are considered an incidental or "dead-end" host for CRPV.[21]
Inter-species transmission has also been documented for bovine papillomavirus (BPV) type 1.[22] In its natural host (cattle), BPV-1 induces large fibrous skin warts. BPV-1 infection of horses, which are an incidental host for the virus, can lead to the development of benign tumors known as sarcoids. The agricultural significance of BPV-1 spurred a successful effort to develop a vaccine against the virus.
A few reports have identified papillomaviruses in smaller rodents, such as Syrian hamsters, the African multimammate rat and the Eurasian harvest mouse.[23] However, there are no papillomaviruses known to be capable of infecting laboratory mice. The lack of a tractable mouse model for papillomavirus infection has been a major limitation for laboratory investigation of papillomaviruses.
Four papillomaviruses are known to infect birds: Fringilla coelebs papillomavirus 1, Francolinus leucoscepus papillomavirus 1, Psittacus erithacus papillomavirus 1 and Pygoscelis adeliae papillomavirus 1.[24] All these species have a gene (E9) of unknown function, suggesting a common origin.
The evolution of papillomaviruses is thought to be slow compared to many other virus types, but there are no experimental measurements currently available. This is probably because the papillomavirus genome is composed of genetically stable double-stranded DNA that is replicated with high fidelity by the host cell's DNA replication machinery.
It is believed that papillomaviruses generally co-evolve with a particular species of host animal over many years, although there are strong evidences against the hypothesis of coevolution.[13][25] In a particularly speedy example, HPV-16 has evolved slightly as human populations have expanded across the globe and now varies in different geographic regions in a way that probably reflects the history of human migration.[26][27] Cutaneotropic HPV types are occasionally exchanged between family members during the entire lifetime, but other donors should also be considered in viral transmission.[28]
Other HPV types, such as HPV-13, vary relatively little in different human populations. In fact, the sequence of HPV-13 closely resembles a papillomavirus of bonobos (also known as pygmy chimpanzees).[29] It is not clear whether this similarity is due to recent transmission between species or because HPV-13 has simply changed very little in the six or so million years since humans and bonobos diverged.[27]
The most recent common ancestor of this group of viruses has been estimated to have existed 424 million years ago.[30]
There are five main genera infecting humans (Alpha, Beta, Gamma, Mu and Nu). The most recent common ancestor of these genera evolved 49.7 million years ago-58.5 million years ago.[31] The most recent ancestor of the gamma genus was estimated to have evolved between 45.3 million years ago and 67.5 million years ago.
Papillomaviruses are non-enveloped, meaning that the outer shell or capsid of the virus is not covered by a lipid membrane. A single viral protein, known as L1, is necessary and sufficient for formation of a 55–60 nanometer capsid composed of 72 star-shaped capsomers (see figure). Like most non-enveloped viruses, the capsid is geometrically regular and presents icosahedral symmetry. Self-assembled virus-like particles composed of L1 are the basis of a successful group of prophylactic HPV vaccines designed to elicit virus-neutralizing antibodies that protect against initial HPV infection. As such, papillomaviridæ are stable in the environment.
The papillomavirus genome is a double-stranded circular DNA molecule ~8,000 base pairs in length. It is packaged within the L1 shell along with cellular histone proteins, which serve to wrap and condense DNA.
The papillomavirus capsid also contains a viral protein known as L2, which is less abundant. Although not clear how L2 is arranged within the virion, it is known to perform several important functions, including facilitating the packaging of the viral genome into nascent virions as well as the infectious entry of the virus into new host cells. L2 is of interest as a possible target for more broadly protective HPV vaccines.
The viral capsid consists of 72 capsomeres of which 12 are five-coordinated and 60 are six-coordinated capsomeres, arranged on a T = 7d icosahedral surface lattice.[32]
Papillomaviruses replicate exclusively in keratinocytes. Keratinocytes form the outermost layers of the skin, as well as some mucosal surfaces, such as the inside of the cheek or the walls of the vagina. These surface tissues, which are known as stratified squamous epithelia, are composed of stacked layers of flattened cells. The cell layers are formed through a process known as cellular differentiation, in which keratinocytes gradually become specialized, eventually forming a hard, crosslinked surface that prevents moisture loss and acts as a barrier against pathogens. Less-differentiated keratinocyte stem cells, replenished on the surface layer, are thought to be the initial target of productive papillomavirus infections. Subsequent steps in the viral life cycle are strictly dependent on the process of keratinocyte differentiation. As a result, papillomaviruses can only replicate in body surface tissues.
Papillomaviruses gain access to keratinocyte stem cells through small wounds, known as microtraumas, in the skin or mucosal surface. Interactions between L1 and sulfated sugars on the cell surface promote initial attachment of the virus.[33][34] The virus is then able to get inside from the cell surface via interaction with a specific receptor, likely via the alpha-6 beta-4 integrin,[35][36] and transported to membrane-enclosed vesicles called endosomes.[37][38] The capsid protein L2 disrupts the membrane of the endosome through a cationic cell-penetrating peptide, allowing the viral genome to escape and traffic, along with L2, to the cell nucleus.[39][40][41]
After successful infection of a keratinocyte, the virus expresses E1 and E2 proteins, which are for replicating and maintaining the viral DNA as a circular episome. The viral oncogenes E6 and E7 promote cell growth by inactivating the tumor suppressor proteins p53 and pRb. Keratinocyte stem cells in the epithelial basement layer can maintain papillomavirus genomes for decades.[8]
The current understanding is that viral DNA replication likely occurs in the G2 phase of the cell cycle and rely on recombination-dependent replication supported by DNA damage response mechanisms (activated by the E7 protein) to produce progeny viral genomes.[42] Papillomavirus genomes are sometimes integrated into the host genome, especially noticeable with oncogenic HPVs, but is not a normal part of the virus life cycle and a dead-end that eliminates the potential of viral progeny production.[42]
The expression of the viral late genes, L1 and L2, is exclusively restricted to differentiating keratinocytes in the outermost layers of the skin or mucosal surface. The increased expression of L1 and L2 is typically correlated with a dramatic increase in the number of copies of the viral genome. Since the outer layers of stratified squamous epithelia are subject to relatively limited surveillance by cells of the immune system, it is thought that this restriction of viral late gene expression represents a form of immune evasion.
New infectious progeny viruses are assembled in the cell nucleus. Papillomaviruses have evolved a mechanism for releasing virions into the environment. Other kinds of non-enveloped animal viruses utilize an active lytic process to kill the host cell, allowing release of progeny virus particles. Often this lytic process is associated with inflammation, which might trigger immune attack against the virus. Papillomaviruses exploit desquamation as a stealthy, non-inflammatory release mechanism.
Although some papillomavirus types can cause cancer in the epithelial tissues they inhabit, cancer is not a typical outcome of infection. The development of papillomavirus-induced cancers typically occurs over the course of many years. Papillomaviruses have been associated with the development of cervical cancer, penile cancer[43] and oral cancers.[44] An association with vulval cancer and urothelial carcinoma with squamous differentiation in patients with neurogenic bladder has also been noted.[45][46] There are cancer causing papillomavirus genome that encodes two small proteins called E6 and E7 that mimic cancer causing oncogenes. The way they work is that they stimulate unnatural growth of cells and block their natural defenses. Also they act on many signaling proteins that control proliferation and apoptosis.[47]
The fact that the papillomavirus life cycle strictly requires keratinocyte differentiation has posed a substantial barrier to the study of papillomaviruses in the laboratory, since it has precluded the use of conventional cell lines to grow the viruses. Because infectious BPV-1 virions can be extracted from the large warts the virus induces on cattle, it has been a workhorse model papillomavirus type for many years. CRPV, rabbit oral papillomavirus (ROPV) and canine oral papillomavirus (COPV) have also been used extensively for laboratory studies. As soon as researchers discovered that these viruses cause cancer, they worked together to find a vaccine to it. Currently, the most effective way to go about it is to mimic a virus that is composed of L1 protein but lack the DNA. Basically, our immune system builds defenses against infections, but if these infections do not cause disease they can be used as a vaccine. PDB entry 6bt3 shows how antibodies surfaces attack the surface of the virus to disable it.[48]
Some sexually transmitted HPV types have been propagated using a mouse "xenograft" system, in which HPV-infected human cells are implanted into immunodeficient mice. More recently, some groups have succeeded in isolating infectious HPV-16 from human cervical lesions. However, isolation of infectious virions using this technique is arduous and the yield of infectious virus is very low.
The differentiation of keratinocytes can be mimicked in vitro by exposing cultured keratinocytes to an air/liquid interface. The adaptation of such "raft culture" systems to the study of papillomaviruses was a significant breakthrough for in vitro study of the viral life cycle.[49] However, raft culture systems are relatively cumbersome and the yield of infectious HPVs can be low.[50]
The development of a yeast-based system that allows stable episomal HPV replication provides a convenient, rapid and inexpensive means to study several aspects of the HPV lifecycle (Angeletti 2002). For example, E2-dependent transcription, genome amplification and efficient encapsidation of full-length HPV DNAs can be easily recreated in yeast (Angeletti 2005).
Recently, transient high-yield methods for producing HPV pseudoviruses carrying reporter genes has been developed. Although pseudoviruses are not suitable for studying certain aspects of the viral life cycle, initial studies suggest that their structure and initial infectious entry into cells is probably similar in many ways to authentic papillomaviruses.
Human papillomavirus binds to heparin molecules on the surface of the cells that it infects. Studies have shown that the crystal of isolated L1 capsomeres has the heparin chains recognized by lysines lines grooves on the surface of the virus. Also those with the antibodies show that they can block this recognition.[51]
The papillomavirus genome is divided into an early region (E), encoding six open reading frames (ORF) (E1, E2, E4, E5, E6, and E7) that are expressed immediately after initial infection of a host cell, and a late region (L) encoding a major capsid protein L1 and a minor capsid protein L2. All viral ORFs are encoded on one DNA strand (see figure). This represents a dramatic difference between papillomaviruses and polyomaviruses, since the latter virus type expresses its early and late genes by bi-directional transcription of both DNA strands. This difference was a major factor in establishment of the consensus that papillomaviruses and polyomaviruses probably never shared a common ancestor, despite the striking similarities in the structures of their virions.
After the host cell is infected, HPV16 early promoter is activated and a polycistronic primary RNA containing all six early ORFs is transcribed. This polycistronic RNA contains three exons and two introns and undergoes active RNA splicing to generate multiple isoforms of mRNAs.[52] One of the spliced isoform RNAs, E6*I, serves as an E7 mRNA to translate E7 oncoprotein.[53] In contrast, an intron in the E6 ORF that remains intact without splicing is necessary for translation of E6 oncoprotein.[53] However, viral early transcription subjects to viral E2 regulation and high E2 levels repress the transcription. HPV genomes integrate into host genome by disruption of E2 ORF, preventing E2 repression on E6 and E7. Thus, viral genome integration into host DNA genome increases E6 and E7 expression to promote cellular proliferation and the chance of malignancy.
A major viral late promoter in viral early region becomes active only in differentiated cells and its activity can be highly enhanced by viral DNA replication. The late transcript is also a polycistronic RNA which contains two introns and three exons. Alternative RNA Splicing of this late transcript is essential for L1 and L2 expression and can be regulated by RNA cis-elements and host splicing factors.[52][54][55]
Genes within the papillomavirus genome are usually identified after similarity with other previously identified genes. However, some spurious open reading frames might have been mistaken as genes simply after their position in the genome, and might not be true genes. This applies specially to certain E3, E4, E5 and E8 open reading frames.
Encodes a protein that binds to the viral origin of replication in the long control region of the viral genome. E1 uses ATP to exert a helicase activity that forces apart the DNA strands, thus preparing the viral genome for replication by cellular DNA replication factors.
The E2 protein serves as a master transcriptional regulator for viral promoters located primarily in the long control region. The protein has a transactivation domain linked by a relatively unstructured hinge region to a well-characterized DNA binding domain. E2 facilitates the binding of E1 to the viral origin of replication. E2 also utilizes a cellular protein known as Bromodomain-4 (Brd4) to tether the viral genome to cellular chromosomes.[56] This tethering to the cell's nuclear matrix ensures faithful distribution of viral genomes to each daughter cell after cell division. It is thought that E2 serves as a negative regulator of expression for the oncogenes E6 and E7 in latently HPV-infected basal layer keratinocytes. Genetic changes, such as integration of the viral DNA into a host cell chromosome, that inactivate E2 expression tend to increase the expression of the E6 and E7 oncogenes, resulting in cellular transformation and possibly further genetic destabilization.
This small putative gene exists only in a few papillomavirus types. The gene is not known to be expressed as a protein and does not appear to serve any function.
Although E4 proteins are expressed at low levels during the early phase of viral infection, expression of E4 increases dramatically during the late phase of infection. In other words, its "E" appellation may be something of a misnomer. In the case of HPV-1, E4 can account for up to 30% of the total protein at the surface of a wart.[57] The E4 protein of many papillomavirus types is thought to facilitate virion release into the environment by disrupting intermediate filaments of the keratinocyte cytoskeleton. Viral mutants incapable of expressing E4 do not support high-level replication of the viral DNA, but it is not yet clear how E4 facilitates DNA replication. E4 has also been shown to participate in arresting cells in the G2 phase of the cell cycle.
The E5 are small, very hydrophobic proteins that destabilise the function of many membrane proteins in the infected cell.[58] The E5 protein of some animal papillomavirus types (mainly bovine papillomavirus type 1) functions as an oncogene primarily by activating the cell growth-promoting signaling of platelet-derived growth factor receptors. The E5 proteins of human papillomaviruses associated to cancer, however, seem to activate the signal cascade initiated by epidermal growth factor upon ligand binding. HPV16 E5 and HPV2 E5 have also been shown to down-regulate the surface expression of major histocompatibility complex class I proteins, which may prevent the infected cell from being eliminated by killer T cells.

E6 is a 151 amino-acid peptide that incorporates a type 1 motif with a consensus sequence –(T/S)-(X)-(V/I)-COOH.[60][61] It also has two zinc finger motifs.[60]
E6 is of particular interest because it appears to have multiple roles in the cell and to interact with many other proteins. Its major role, however, is to mediate the degradation of p53, a major tumor suppressor protein, reducing the cell's ability to respond to DNA damage.[62][63]
E6 has also been shown to target other cellular proteins, thereby altering several metabolic pathways. One such target is NFX1-91, which normally represses production of telomerase, a protein that allows cells to divide an unlimited number of times. When NFX1-91 is degraded by E6, telomerase levels increase, inactivating a major mechanism keeping cell growth in check.[64] Additionally, E6 can act as a transcriptional cofactor—specifically, a transcription activator—when interacting with the cellular transcription factor, E2F1/DP1.[60]
E6 can also bind to PDZ-domains, short sequences which are often found in signaling proteins. E6's structural motif allows for interaction with PDZ domains on DLG (discs large) and hDLG (Drosophila large) tumor suppressor genes.[61][65] Binding at these locations causes transformation of the DLG protein and disruption of its suppressor function. E6 proteins also interact with the MAGUK (membrane-associated guanylate kinase family) proteins. These proteins, including MAGI-1, MAGI-2, and MAGI-3 are usually structural proteins, and can help with signaling.[61][65] More significantly, they are believed to be involved with DLG's suppression activity. When E6 complexes with the PDZ domains on the MAGI proteins, it distorts their shape and thereby impedes their function. Overall, the E6 protein serves to impede normal protein activity in such a way as to allow a cell to grow and multiply at the increased rate characteristic of cancer.
Since the expression of E6 is strictly required for maintenance of a malignant phenotype in HPV-induced cancers, it is an appealing target of therapeutic HPV vaccines designed to eradicate established cervical cancer tumors.
In most papillomavirus types, the primary function of the E7 protein is to inactivate members of the pRb family of tumor suppressor proteins. Together with E6, E7 serves to prevent cell death (apoptosis) and promote cell cycle progression, thus priming the cell for replication of the viral DNA. E7 also participates in immortalization of infected cells by activating cellular telomerase. Like E6, E7 is the subject of intense research interest and is believed to exert a wide variety of other effects on infected cells. As with E6, the ongoing expression of E7 is required for survival of cancer cell lines, such as HeLa, that are derived from HPV-induced tumors.[66]
Only a few papillomavirus types encode a short protein from the E8 gene. In the case of BPV-4 (papillomavirus genus Xi), the E8 open reading frame may substitute for the E6 open reading frame, which is absent in this papillomavirus genus.[67] These E8 genes are chemically and functionally similar to the E5 genes from some human papillomaviruses, and are also called E5/E8.
L1 spontaneously self-assembles into pentameric capsomers. Purified capsomers can go on to form capsids, which are stabilized by disulfide bonds between neighboring L1 molecules. L1 capsids assembled in vitro are the basis of prophylactic vaccines against several HPV types. Compared to other papillomavirus genes, the amino acid sequences of most portions of L1 are well-conserved between types. However, the surface loops of L1 can differ substantially, even for different members of a particular papillomavirus species. This probably reflects a mechanism for evasion of neutralizing antibody responses elicited by previous papillomavirus infections.[68]
L2 exists in an oxidized state within the papillomavirus virion, with the two conserved cysteine residues forming an intramolecular disulfide bond.[69] In addition to cooperating with L1 to package the viral DNA into the virion, L2 has been shown to interact with a number of cellular proteins during the infectious entry process. After the initial binding of the virion to the cell, L2 must be cleaved by the cellular protease furin.[70] The virion is internalized, probably through a clathrin-mediated process, into an endosome, where acidic conditions are thought to lead to exposure of membrane-destabilizing portions of L2.[39] The cellular proteins beta-actin[71] and syntaxin-18[72] may also participate in L2-mediated entry events. After endosome escape, L2 and the viral genome are imported into the cell nucleus where they traffic to a sub-nuclear domain known as an ND-10 body that is rich in transcription factors.[40] Small portions of L2 are well-conserved between different papillomavirus types, and experimental vaccines targeting these conserved domains may offer protection against a broad range of HPV types.[73]
Papillomaviridae is a family of non-enveloped DNA viruses whose members are known as papillomaviruses. Several hundred species of papillomaviruses, traditionally referred to as "types", have been identified infecting all carefully inspected mammals, but also other vertebrates such as birds, snakes, turtles and fish. Infection by most papillomavirus types, depending on the type, is either asymptomatic (e.g. most Beta-PVs) or causes small benign tumors, known as papillomas or warts (e.g. human papillomavirus 1, HPV6 or HPV11). Papillomas caused by some types, however, such as human papillomaviruses 16 and 18, carry a risk of becoming cancerous.
Papillomaviruses are usually considered as highly host- and tissue-tropic, and are thought to rarely be transmitted between species. Papillomaviruses replicate exclusively in the basal layer of the body surface tissues. All known papillomavirus types infect a particular body surface, typically the skin or mucosal epithelium of the genitals, anus, mouth, or airways. For example, human papillomavirus (HPV) type 1 tends to infect the soles of the feet, and HPV type 2 the palms of the hands, where they may cause warts. Additionally, there are descriptions of the presence of papillomavirus DNA in the blood and in the peripheral blood mononuclear cells.
Papillomaviruses were first identified in the early 20th century, when it was shown that skin warts, or papillomas, could be transmitted between individuals by a filterable infectious agent. In 1935 Francis Peyton Rous, who had previously demonstrated the existence of a cancer-causing sarcoma virus in chickens, went on to show that a papillomavirus could cause skin cancer in infected rabbits. This was the first demonstration that a virus could cause cancer in mammals.
Papillomaviridae es una familia de virus que infectan vertebrados.[1][2][3] Además, se conocen más de 100 diferentes tipos de papilomavirus humanos (VPH). Tienen un genoma ADN bicatenario y por lo tanto pertenecen al Grupo I de la Clasificación de Baltimore. Su estructura se caracteriza por no presentar envoltura, la cápside es de simetría icosaédrica y mide unos 60 nm de diámetro.
El virus del papiloma se identificó por primera vez a principios del siglo XX, cuando se demostró que las verrugas podrían transmitirse entre personas por un agente infeccioso filtrable. En 1935 Francis Peyton Rous, que había demostrado previamente la existencia de un cáncer que causa el sarcoma viral en pollos, demostró que el virus del papiloma puede causar cáncer de piel en conejos infectados. Esta fue la primera demostración de que un virus podía causar cáncer en los mamíferos.
El virus del papiloma se replica exclusivamente en la superficie de los tejidos corporales como la piel, mucosas, genitales, ano, boca o vías respiratorias.[4] La mayoría de los tipos de virus del papiloma se adaptan a la infección a una superficie corporal particular. Por ejemplo, los VPH de tipos 1 y 2 tienden a infectar las plantas de los pies o las palmas de las manos, respectivamente, donde pueden causar verrugas.[5] Los cánceres causados por una docena de tipos de VPH de "alto riesgo" matan varios cientos de miles de personas al año en todo el mundo y son uno de los focos importantes de investigación en salud pública.[6]
Las distintas especies de virus del papiloma tienden a ser muy específicas en la replicación, que suele realizarse en una sola especie animal. En un estudio los investigadores intercambiaron piel de una gran variedad de animales en un zoológico y utilizaron PCR para amplificar el ADN del virus que debería estar presente.[7] A pesar de la amplia variedad de secuencias del virus del papiloma que se identificaron en el estudio, los autores encontraron pocas pruebas de transmisión interespecífica. Curiosamente, un cuidador resultó positivo transitoriamente a la secuencia de un virus del papiloma específico del chimpancé. Sin embargo, los autores señalan que la secuencia detectada podría haber sido el resultado de contaminación superficial en la piel del cuidador, frente a una infección productiva.
El virus del papiloma de la cola de algodón del conejo (CRPV) puede causar verrugas protuberantes en su huésped nativo, el conejo de América del Norte del género Sylvilagus. Los conejos domésticos europeos del género Oryctolagus pueden ser infectados en el laboratorio temporalmente con el CRPV. Sin embargo, puesto que los conejos domésticos Europea no producen progenie infecciosas del virus, se consideran como un "callejón sin salida" para el CRPV.[8]
La transmisión interespecífica ha sido también documentada para el papilomavirus bovino de tipo 1.[9] En su huésped natural, el ganado, BPV-1 produce la piel grandes verrugas fibrosas en la piel. La infección por BPV-1 en el caballo, que es un hospedador accidental para el virus, puede conducir al desarrollo de tumores benignos conocidos como sarcoides. La importancia de BPV-1 en la ganadería ha conducido al éxito en los esfuerzos para desarrollar una vacuna contra el virus.
En algunos estudios se han identificado virus del papiloma en pequeños roedores, tales como hámsteres dorados, ratas africanas y ratones de campo europeos.[10] Sin embargo, no se conocen virus del papiloma capaces de infectar al ratón de laboratorio. La falta de un virus modelo que infecte fácilmente al ratón ha sido una importante limitación en el laboratorio para la investigación de los virus del papiloma.
Los virus del papiloma no presentan envoltura. Una sola proteína viral, denominada L1, es necesaria y suficiente para la formación de un cápside de 60 nm compuesta por 72 capsómeros con forma de estrella. Como la mayoría de los virus sin envoltura, la cápside es geométricamente regular y presentan simetría icosaédrica. Las partículas auto-ensambladas similares al virus compuestas de L1 son la base de un grupo de las vacunas profilácticacontra el virus, diseñados para generar anticuerpos neutralizantes del virus que protegen contra la infección inicial.
El genoma es una molécula circular de ADN bicatenario de aproximadamente 8000 pares de bases de longitud. Es ensamblado dentro de la cubierta L1 junto con las proteínas histonas celulares, que envuelven y condensan el ADN.
La cápside viral también contiene una proteína conocida como L2, que es menos abundante. Aunque no está claro cómo se organiza L2 en el virión, se sabe que realiza varias funciones importantes, en particular, facilita el ensamblado del genoma viral en los viriones nacientes, así como la entrada del virus en nuevas células huésped. L2 es interesante como un posible objetivo para vacunas de protección más amplia contra el virus.
La evolución de los virus del papiloma es lenta en comparación con muchos otros tipos de virus. Esto probablemente se debe a que el genoma del virus del papiloma está compuesto por DNA de doble cadena genéticamente estable que es replicado con alta fidelidad por la maquinaria de la célula huésped.
Se cree que el virus del papiloma generalmente co-evoluciona durante muchos años con una especie particular de huésped animal. Como ejemplo particular, el VPH-16 que afecta a los humanos ha evolucionado ligeramente y en la actualidad varía en diferentes regiones geográficas de una forma que probablemente refleja la historia de la migración humana.[11][12]
Otros tipos de VPH, tales como VPH-13, varían relativamente poco en las diferentes poblaciones humanas. De hecho, la secuencia de VPH-13 se parece a un virus del papiloma de los bonobos (también conocidos como chimpancés pigmeos).[13] No está claro si esta similitud se debe a la reciente transmisión entre especies o porque el VPH-13, simplemente ha cambiado muy poco en los seis o más millones de años desde que los seres humanos y bonobos divergieron.[12]
Los papilomavirus presentan organizaciones genómicas similares, y cualquier par de especies contiene al menos cinco genes homólogos, aunque la secuencia de nucleótidos puede diferir en más de un 50%. Los algoritmos filogenéticos que permitan la comparación de homologías llevado a árboles filogenéticos que tienen una topología similar independiente de los genes analizados. El análisis filogenético sugiere fuertemente que normalmente los virus evolucionan junto a las especies de mamíferos y aves que actúan como huéspedes, no cambian de especie huésped, no se recombinan, y han mantenido su organización base genómica, por un período superior a 100 millones de años.
Esta comparación de secuencias han sentado las bases de una taxonomía de los papilomavirus, que es ahora reconocida oficialmente por el Comité Internacional de Taxonomía de Virus. Todos los papilomavirus se incluyen en la familia Papillomaviridae. Las principales ramas del árbol filogenético son considerados géneros, que se identifican por letras griegas. Las ramas inferiores se consideran especies y unen tipos que son genómicamente distintos pero no exhiben diferencias biológicas conocidas. Este nuevo sistema taxonómico no afecta a la tradicional identificación y caracterización de "tipos" y muestras con diferencias genómicas pequeñas, conocidas como "subtipos" y "variantes", todos los cuales son taxones por debajo del nivel de "especie".[14]
Papillomaviridae es una familia de virus que infectan vertebrados. Además, se conocen más de 100 diferentes tipos de papilomavirus humanos (VPH). Tienen un genoma ADN bicatenario y por lo tanto pertenecen al Grupo I de la Clasificación de Baltimore. Su estructura se caracteriza por no presentar envoltura, la cápside es de simetría icosaédrica y mide unos 60 nm de diámetro.
El virus del papiloma se identificó por primera vez a principios del siglo XX, cuando se demostró que las verrugas podrían transmitirse entre personas por un agente infeccioso filtrable. En 1935 Francis Peyton Rous, que había demostrado previamente la existencia de un cáncer que causa el sarcoma viral en pollos, demostró que el virus del papiloma puede causar cáncer de piel en conejos infectados. Esta fue la primera demostración de que un virus podía causar cáncer en los mamíferos.
Papilloomiviirused (Papillomaviridae) on sugukond väikseid ümbriseta DNA-viirusi.
Sugukonda Papillomaviridae liigitatakse järgmised viiruste perekonnad:
Võrreldes teiste viirustega on papilloomiviiruste evolutsioon olnud aeglane. Aeglane areng tuleneb sellest, et papilloomiviiruse genoomi, mis koosneb geneetiliselt stabiilsest kaheahelalisest DNA-st, sünteesitakse ainult peremeesraku DNA replikatsioonimehhanismi abil. Papilloomiviirused arenevad üldiselt koos oma peremeesorganismiga. HPV-16 arenes inimpopulatsioonide laienemisel üle maakera ning nüüd ta varieerub erinevates geograafilistes piirkondades.[1][2]
Teised HPV tüübid, nagu HPV-13, varieeruvad erinevates inimpopulatsioonides vähe. HPV-13 järjestus on sarnane bonobote papilloomiviiruste omaga.[3] Ei ole selge, kas sarnasuse põhjuseks on hiljutine liikidevaheline ülekandumine või asjaolu, et HPV-13 on alates inimese ja bonobote lahknemisest vähe muutunud.[2]
Papilloomiviirused on ümbriseta viirused, mis tähendab, et viiruse kapsiid ei ole kaetud lipiidse membraaniga. Peamine struktuurvalk on L1, mis on vajalik ja piisav 55–60 nm diameetriga kapsiidi moodustamiseks. Kapsiidi koostisse kuulub 72 tähekujulist kapsomeeri. Sarnaselt suurema osaga ümbriseta viirustest on papilloomiviiruse kapsiidi geomeetria korrapärane ja omab ikosaeedrilist sümmeetriat. In vitro L1 valgust kokku pandud viiruselaadsed partiklid on edukate profülaktiliste HPV vaktsiinide aluseks.
Papilloomiviiruse genoom on kaheahelaline tsirkulaarne DNA molekul. Pikkus on umbes 8000 aluspaari. DNA paikneb virionis koos rakuliste histoonidega, mis moodustab kromatiiniga sarnaneva kompleksi.
Papilloomiviiruse kapsiid sisaldab lisaks L1 valgule ka valku L2. Peale kapsiidi moodustamise täidab E2 ka selliseid funktsioone nagu genoomi pakkimise hõlbustamine virionidesse ja viiruse infektsioonilisele sisenemisele kaasaaitamine. L2 on HPV vaktsiinide üks võimalikke sihtmärke.
Papilloomiviirused replitseeruvad üksnes keratinotsüütides, mis paiknevad naha välimistes kihtides ja limaskestadel. Need epiteelkoed koosnevad lamestunud rakkude kihtidest. Kihid moodustuvad rakulise diferentseerumise tulemusena, milles keratinotsüüdid järk-järgult spetsialiseeruvad. Lõpuks moodustub tugev ja omavaheliste kontaktidega seostunud pind, mis takistab niiskuskadu ja on mehaaniliseks barjääriks patogeenidele. Vähediferentseerunud keratinotsüütide tüvirakud paiknevad basaalkihis ning nad täiendavad pidevalt pindmisi kihte. Kuna nad on ainsana epiteelirakkudest jagunemisvõimelised, siis on nad papilloomiviiruse infektsiooni sihtmärgiks. Edasised sammud viiruse elutsüklis on ranges sõltuvuses keratinotsüütide diferentseerumisest ning seetõttu saavad papilloomiviirused replitseeruda ainult keha pindmistes kudedes.
Nende genoomi moodustab kaheahelaline tsirkulaarne DNA molekul, mille keskmine pikkus on 8000 aluspaari. Põhiliseks struktuurvalguks on L1, mille abil moodustatakse korrapärane ikosaeedrilise sümmeetriaga ja 55–60 nm diameetriga kapsiid.
Teatakse sadu papilloomiviiruste liike, millele viidatakse kui "tüüpidele". Need on väga peremehe- ja koespetsiifilised. Viirused nakatavad epiteelkoe rakke ning replitseeruvad rakutuumas. Seejuures võivad mõned viiruse tüübid vähki tekitada.
Siiani on koostatud rohkem kui saja inimese papilloomiviiruse tüübi genoomid ja PCR-andmete töötlemise tulemiks võib olla veel umbes 100 tüübi olemasolu tuvastamine.[4]
Tuntuim esindaja on inimese papilloomiviirus.
Papilloomiviiruse genoom on jagatud varajaseks regiooniks (early region, E) ja hiliseks regiooniks (late region, L). Varajase ja hilise kodeeriva ala vahele jääb mittekodeeriv regulatoorne ala (URR ehk upstream regulatory region, LCR ehk long control region). Varajane regioon kodeerib geene, mida ekspresseeritakse kohe pärast esmast raku infektsiooni ning hiline regioon kapsiidivalke L1 ja L2. Kõik geenid asuvad ühes DNA ahelas. See on suur erinevus papilloomiviiruste ja polüoomiviiruste vahel, kuna viimases ekspresseeritakse varajased ja hilised geenid bipolaarse transkriptsiooniga mõlemalt DNA ahelalt. See näitab, et papilloomiviirustel ja polüoomiviirustel pole olnud ühist eellast, kuigi nende struktuurides on märkimisväärsed sarnasused.
E1 on papilloomiviiruse peamine replikatsioonivalk. E1 avatud lugemisraam (open reading frame, ORF) kodeerib valku, mis seostub viraalsele replikatsioonioriginile ehk replikatsiooni alguspunktile LCR'is. E1 kasutab ATP energiat, et rakendada helikaasset aktiivsust DNA ahelate lahutamiseks. Nii valmistab ta viiruse genoomi ette DNA replikatsiooniks.
E2 valk täidab viraalsetes promootorites põhiliselt transkriptsioon regulaatori rolli. Valgul on transaktivatsiooni domeen ja DNA-d seondav domeen, mis on omavahel seotud hinge (hinge) domeeniga. Üksinda seondub E1 valk replikatsioonioriginile nõrgalt. E2 aitab kaasa E1 tugevamale seondumisele. E2 valk seondab viiruse genoomi ka raku mitootilistele kromosoomidele ja see on oluline viiruse genoomi segregatsioonil ehk jaotumisel tütarrakkude vahel.[5] Arvatakse, et onkogeenide E6 ja E7 ekspresseerumisel käitub E2 negatiivse regulaatorina. Geneetilised muutused, nagu viiruse DNA integratsioon peremeesraku kromosoomi, inaktiveerivad E2 ekspressiooni ja suureneb onkogeenide E6 ja E7 ekspressioon. Tulemuseks on rakusisesed muutused.
Väikest oletuslikku geeni E3 leidub ainult vähestes papilloomiviiruse tüüpides. Siiani ei ole õnnestunud tuvastada selle geeni ekspresseeritavat valku ja tal ei tundu olevat funktsioone.
E4 valke ekspresseeritakse varajases infektsioonifaasis madalal tasemel, kuigi ta kuulub varajaste geenite hulka. Hilises infektsioonifaasis võib ekspressioon järsult suureneda. HPV-1 tekitatud tüüka puhul moodustab E4 ligi 30% tüüka pinnavalgust.[6] Arvatakse, et mitmetes papilloomiviiruse tüüpides vahendab E4 virionide vabanemist keskkonda, selleks katkestab ta keratinotsüütide tsütoskeleti filamentide vahelised sidemed. E4 mutantidel ei toimub kõrgel tasemel DNA replikatsiooni, kuid ei ole selge, kuidas E4 replikatsioonis osaleb. Samuti on näidatud, et E4 osaleb rakutsükli peatamises G2 faasis.
E5 valgud on väga väikesed ja hüdrofoobsed, mis destabiliseerivad suurt hulka nakatunud raku membraanivalkude funktsioone.[7] Mõnedes papilloomiviiruse tüüpides (enamjaolt BPV-1) toimib E5 onkogeenina. Sellisel juhul ta aktiveerib rakkude kasvu soodustavaid signaale. Inimese papilloomiviirustes on E5 seotud vähitekkega. HPV-16 ja HPV-2 on näidatud, et E5 surub maha peamise koesobivuskompleksi (major histocompatibility complex, MHC), mis hoiab ära nakatunud rakkude elimineerimise tsütotoksiliste T-rakkude poolt.
Valgul E6 on rakus palju erinevaid rolle ja ta omab vastastikmõju paljude teiste valkudega. Peamiseks rolliks on tuumor-supressorvalgu p53 lagundamine. p53 peamine funktsioon on rakutsükli peatamine DNA kahjustuse korral ja raku suunamine apoptoosi, kui kahjustuse ulatus on parandamiseks liiga suur. p53 lagundamine vähendab raku suutlikkust vastata DNA kahjustustele.[8][9] E6 sihtmärgiks on mitmed rakulised valgud ning ta suudab muuta metabolismiradasid. Üheks selliseks valguks on NFX1-91, mis surub maha telomeraasi tootmist. Telomeraas lubab rakkudel jaguneda piiramata arv kordi. Kui NFX1-91 lagundatakse E6 poolt, siis telomeraasi hulk suureneb ja üks peamisi raku kasvu kontrollivaid mehhanisme inaktiveeritakse.[10]
HPV põhjustatud vähkides on E6 ekspressioon vajalik pahaloomulise fenotüübi säilitamiseks. Seetõttu on E6 ka inimese papilloomiviiruste vaktsiinide sihtmärk.
Enamikus papilloomiviirusetüüpides on E7 funktsiooniks tuumor-supressorvalgu pRb mahasurumine. Samuti indutseerib ta DNA sünteesi latentses faasis. Koos valguga E6 hoiab E7 ära raku surma e apoptoosi ning soodustab rakutsükli kulgemist. E7 osaleb telomeraasi aktiveerimises. E6 ja E7 on tähtsad uurimisobjektid ja usutakse, et neil on nakatunud rakkudes palju teisi olulisi efekte. E6 ja E7 on vajalikud vähirakuliinide elushoidmiseks (nt HeLa).[11]
Vähesed papilloomiviiruse tüübid kodeerivad E8 geenist lühikest valku. Näiteks mõnedel veise papilloomiviirustel on E8 geenid funktsionaalselt sarnased HPV E5 geenidega.[12]
L1 on peamine kapsiidivalk ja temast piisab viiruslaadsete partiklite moodustamiseks. L1 valgust in vitro kokku pandud kapsiidid on mitmete HPV-vastaste vaktsiinide aluseks.
L2 on samuti kapsiidivalk, mida leidub papilloomiviiruse virionis oksüdeeritud kujul. L2 osaleb koos valguga L1 viiruse DNA pakkimisel virioni, kuid L2 osaleb ka viiruse sisenemisel rakku.[13] Pärast sisenemist satub virion endosoomi, kus happelised tingimused viivad L2 valgu põhjustatud destabiliseerivate funktsioonideni. Vabanedes endosoomist, liigub viiruse genoom rakutuuma. Kuna L2 on oluline nakatumiseks, on ta huviobjektiks vaktsiinide väljatöötajatele.[14]
Papilloomiviiruse produktiivne infektsioon algab epiteelkoe basaalse rakukihi nakatamisest. Viirus pääseb keratinotsüütidele ligi väikeste haavade kaudu nahas või limaskestas. Esmast viiruse kinnitumist raku pinnale vahendavad kapsiidivalgu L1 ja sulfateeritud suhkrute interaktsioonid.[15][16] Viirus on seejärel võimeline sisenema rakku, spetsiifilisele retseptorile seondudes. Raku pinnal asuvaks retseptoriks on tõenäoliselt alfa-6 beeta-4 integriin.[17][18] Viiruse antiretseptoriks on kapsiidivalk L1. Seostunud viirus transporditakse endosoomidesse.[19][20] Kapsiidivalk L2 katkestab endosoomi membraani, mis lubab viiruse genoomil väljuda. Genoom transporditakse koos L2 valguga rakutuuma.[21][22]
Pärast edukat keratinotsüütide nakatumist hakkab viirus ekspresseerima valke E1 ja E2. Need valgud on vajalikud replitseerumiseks ja viiruse säilitamiseks tsirkulaarse episoomina. Viiruse genoomi koopiaarv on episoomina säilides väike. Viiruslikud onkogeenid E6 ja E7 inaktiveerivad tuumor-supressorgeenid p53 ja pRb, millega stimuleeritakse raku kasvu. Papilloomiviiruse genoomid võivad säilida keratinotsüütide tüvirakkudes episoomina aastakümneid.[23]
Hiliste geenide L1 ja L2 ekspressioon toimub ainult naha või limaskesta välimise kihi keratinotsüütides. L1 ja L2 suurenenud ekspressioon on seotud viiruse genoomi koopianumbri hüppelise kasvuga. Kuna välimiste kihtide rakud on suhteliselt väikse immuunsüsteemirakkude järelevalve all, siis arvatakse, et hiliste geenide ekspressioon on seotud immuunsüsteemist kõrvalehoidmisega.
Uued nakatumisvõimelised viirused pannakse kokku rakutuumas. Papilloomiviirustel on arenenud mehhanism, kuidas vabastada virione keskkonda. Teised ümbriseta imetajate viirused kasutavad lüütilisi protsesse peremeesraku tapmiseks, mis võimaldavad viiruspartiklite vabanemist. Sageli kaasneb selle protsessiga põletik, mis omakorda võib päästa valla immuunsüsteemi rünnaku viiruse vastu. Papilloomiviirused kasutavad ära aga epiteeli pindmise kihi rakkude irdumist kui mittepõletikulist viiruspartiklite vabastamismehhanismi.
Papilloomiviirused tehti kindlaks 20. sajandil, kui leiti, et papilloomid või tüükad võivad kanduda ühelt indiviidilt teisele.
Papilloomiviirused nakatavad linde ja imetajaid,[26] aga ka väikest hulka roomajaid (kilpkonni).[27][28]
Enamiku papilloomiviiruste infektsioon on kas asümptomaatiline (näiteks tüüp 6, tüüp 11) või põhjustab väikseid healoomulisi kasvajaid, mis on tuntud kui papilloomid või tüükad (inglise warts).
Papilloomiviirused on peremehe- ja koespetsiifilised ning seega kanduvad liigilt liigile üle väga harva.[29] Papilloomiviirused replitseeruvad ainult keha epiteelkudede basaalkihis. Kõik tuntud papilloomiviirused nakatavad kindlat epiteeli,[26] tüüpiliselt nahaepiteeli või genitaalide, suu või hingamisteede limaskesta.[23] Limaskesta nakatavate inimese papilloomiviiruste (HPV) tüübid jagunevad kõrgesse ja madalasse riskirühma vastavalt sellele, kas nad on seotud vähitekkega või mitte. Kõrge riskiga on näiteks inimese papilloomiviiruse tüübid 16, 18, 31 ja 45 ning nende põhjustatud genitaalpiirkonna nakkused võivad edasi areneda pahaloomulisteks kasvajateks. Madala riski tüübid on näiteks HPV-6 ja −11, mis kahekesi vastutavad ligi 90% kõigi genitaalpiirkonna healoomuliste papilloomide tekke eest, kuid nende arenemist vähiks pole kirjeldatud.
Papilloomiviiruste tüübid on kõrgelt kohanenud replitseeruma ainult üksikutes loomaliikides. Sooküüliku papilloomiviirus (cottontail rabbit papillomavirus, CRPV) võib oma Põhja-Ameerika peremehes (sooküülikutel – perekond Sylvilagus) põhjustada kühmuna esileulatuvaid tüükaid.[25] Euroopa kodujänestele (perekond Oryctolagus) saab CRPV kanduda üle ainult laboratoorsetes tingimustes. Euroopa kodujänesed ei suuda toota nakatavaid viiruspartikleid, seega peetakse neid CRPV jaoks juhuslikeks või tupikperemeesteks.[30]
Liikidevahelist ülekandumist on näidatud ka veise papilloomiviiruse (BPV) tüübil 1.[31] Veistel põhjustab BPV-1 suuri kiulisi nahatüükaid. BPV nakatab ka hobuseid, kes on viirusele juhuslikud peremehed. Nakatumine võib viia healoomuliste kasvajate tekkeni.
Papilloomiviiruse juhte on olnud ka väiksematel närilistel, nagu Süüria hamster ja Euraasia põlluhiir.[32] Ometi ei ole leitud papilloomiviirust, mis suudaks nakatada laborihiiri ja seetõttu puudub laboratoorseteks uuringuteks hea uurimismudel.
Kuigi mõned papilloomiviiruse tüübid põhjustavad vähki, ei ole vähk infektsiooni tüüpiline tulemus. Tavaliselt toimub papilloomiviiruste indutseeritud vähkide areng mitme aasta jooksul.
Papilloomiviirused (Papillomaviridae) on sugukond väikseid ümbriseta DNA-viirusi.
La famille des Papillomaviridae, dont les membres sont couramment appelés papillomavirus, est un groupe de petits virus à ADN. Ils sont très répandus dans la nature. C’est la seule famille de l’ordre des Zurhausenvirales.
Ils infectent de nombreux mammifères (humain, bovidés, canidés) et ont un tropisme particulier pour les kératinocytes des épithéliums malpighiens qu'ils soient kératinisés ou non[3]. Ils peuvent également affecter d'autres groupes comme les oiseaux, les serpents et les tortues.
Le papillomavirus humain se divise en 2 familles :
Les lapins atteints d'un papillomavirus CRPV (cottontail rabbit papillomavirus) peuvent développer des tumeurs noirâtres et allongées (sur le corps, parfois sur la tête). Cet aspect étrange pourrait être l'une des origines de la légende de l'existence d'un animal mi-lièvre, mi-antilope : le jackalope.
Du latin -papillo (« mamelon ») et oma (« tumeur »), les papillomavirus sont des virus à ADN non enveloppés qui induisent des lésions exophytiques de la peau et des muqueuses. Le premier papillomavirus animal a été décrit en 1933 par Richard Shope, qui a étudié les papillomes chez les lapins de garenne sauvages « verruqueux ». En 1975, Harald zur Hausen a publié l'hypothèse que le virus du papillome humain a joué un rôle dans l'étiologie du cancer du col utérin, un travail pour lequel il a reçu le prix Nobel de physiologie ou médecine en 2008[4].
La famille des Papillomaviridae, dont les membres sont couramment appelés papillomavirus, est un groupe de petits virus à ADN. Ils sont très répandus dans la nature. C’est la seule famille de l’ordre des Zurhausenvirales.
Ils infectent de nombreux mammifères (humain, bovidés, canidés) et ont un tropisme particulier pour les kératinocytes des épithéliums malpighiens qu'ils soient kératinisés ou non. Ils peuvent également affecter d'autres groupes comme les oiseaux, les serpents et les tortues.
Papillomawirusy (łac. Papillomaviridae) - rodzina wirusów, zaliczanych wspólnie z poliomawirusami do grupy papowawirusów. Papillomawirusy charakteryzują się następującymi cechami:
Dotychczas zidentyfikowano ok. 60 odmian ludzkiego papillomawirusa (HPV). Znane są także papillomawirusy innych kręgowców. Pozycja systematyczna papillomawirusów jest następująca:
HPV wywołuje szereg chorób, wiele z nich związanych jest z transformacją nowotworową. Ze zwględu na złożoność zagadnienia, zostały one przedstawione w artykule dotyczącym wirusa brodawczaka ludzkiego. Charakter chorób przez niego wywołanych (brodawczaki - papilloma) przyczynił się do nadania nazwy całej rodzinie.
Papillomawirusy (łac. Papillomaviridae) - rodzina wirusów, zaliczanych wspólnie z poliomawirusami do grupy papowawirusów. Papillomawirusy charakteryzują się następującymi cechami:
Symetria: ikosaedralna Otoczka lipidowa: brak Kwas nukleinowy: kolisty dsDNA Replikacja: zachodzi w jądrze zakażonej komórki Wielkość: ok. 55 nm Gospodarz: kręgowce Cechy dodatkowe: trudne w hodowli ze względu na zdolność do namnażania się jedynie w różnicującym się wielowarstwowym nabłonku płaskim. Komórki te są niezwykle trudne w hodowli, co powoduje, że papillomawirusy są stosunkowo słabo zbadaneDotychczas zidentyfikowano ok. 60 odmian ludzkiego papillomawirusa (HPV). Znane są także papillomawirusy innych kręgowców. Pozycja systematyczna papillomawirusów jest następująca:
Rodzina: Papillomaviridae (Papillomawirusy) Rodzaj: Papillomavirus Human papillomavirus, wirus brodawczaka ludzkiegoHPV wywołuje szereg chorób, wiele z nich związanych jest z transformacją nowotworową. Ze zwględu na złożoność zagadnienia, zostały one przedstawione w artykule dotyczącym wirusa brodawczaka ludzkiego. Charakter chorób przez niego wywołanych (brodawczaki - papilloma) przyczynił się do nadania nazwy całej rodzinie.
Papillomaviridae é uma família de vírus que infectam vertebrados (incluindo humanos). Os papilomavírus possuem genoma de DNA fita dupla circular, que mede de 6,8 a 8,4 kb e codifica entre 8 e 10 ORFs.[1] A partícula viral destes vírus consiste em um capsídeo icosaédrico (formado por 72 capsômeros), não-envelopado, medindo de 52 a 55 nm. A replicação e a montagem dos vírions ocorre no núcleo celular. Alguns papilomavírus são oncogênicos, como o vírus do papiloma humano (HPV). Papillomaviridae integra o grupo I do sistema de classificação de Baltimore.[2]
Esta família viral possui 16 gêneros:[3]
Papillomaviridae é uma família de vírus que infectam vertebrados (incluindo humanos). Os papilomavírus possuem genoma de DNA fita dupla circular, que mede de 6,8 a 8,4 kb e codifica entre 8 e 10 ORFs. A partícula viral destes vírus consiste em um capsídeo icosaédrico (formado por 72 capsômeros), não-envelopado, medindo de 52 a 55 nm. A replicação e a montagem dos vírions ocorre no núcleo celular. Alguns papilomavírus são oncogênicos, como o vírus do papiloma humano (HPV). Papillomaviridae integra o grupo I do sistema de classificação de Baltimore.
Папіломавіруси (Papillomaviridae) — родина вірусів, що були ідентифіковані на початку 20 століття, коли було показано, що бородавки (папіломи) можуть передаватися між інивідуумами за допомогою виділеного інфекційного агенту.
В 1935 році Френсіс Пейтон Раус, який до того продемонстрував існування віруса саркоми у курей, також показав, що папіломавірус може викликати рак шкіри у заражених кролів. Це було першою демонстрацією того, що вірус може викликати рак у ссавців.
Прості віруси. Віріони сферичні, ікосаедрична симетрія (Т=7), 55 нм в діаметрі. Капсид складається з 72 пентамерів. Маса віріонів Mr = 47x106.
Плавуча густина в CsCl = 1.34-1.35 г/см3. Віріони нечутливі до дії кислот, ефірів, нагрівання (50˚С, 1 год.).
Коефіцієнт седиментації S20,w = 300S.
Тип нуклеїнової кислоти: кільцева dsDNA суперспіралізована, формує хроматиноподібний комплекс з гістонами, взятими у клітини-хазяїна. Розмір: 6.8-8.4 kbp. Вміст G+C = 40-60%. Геном більшості видів містить 9-10 ORFs: Е1-Е8 (ранні гени), L1-L2 (пізні гени). Транскрипція і пізніх, і ранніх генів відбувається на одному ланцюгу ДНК та в одному напрямку. Пре-мРНК піддається посттранскрипційному процесингу (приєднання кепу, поліаденілування, сплайсинг). Продукти ранніх генів – неструктурні, регуляторні білки, які необхідні для реплікації, що пригнічують дію клітинних ферментів та стимулюють реплікацію ДНК клітини. Пізні гени кодують структурні білки. Реплікація вірусу поділена на дві чіткі фази, які пов’язані зі ступенем диференціації клітини-хазяїна (епітеліоцита).
Цикл вірусу:
вірус адсорбується до рецепторів клітини-мішені і потрапляє в цитоплазму шляхом ендоцитозу. транспортується в ядро, ДНК роздягається. транскрипція ранніх генів і трансляція ранніх білків. «Спокійна» реплікація вірусної ДНК, відбувається синхронно з клітинною реплікацією, переважно один вірусний геном на клітину.
Вірусний геном кодує 8-10 білків масою 7-73 kDa. Капсид побудований двома білками: L1 (головний капсидний білок, формує пентамери) та L2 (мінорний капсидний білок). До складу віріону входять клітинні гістонові білки (H2a, H2b, H3 і H4), які зв'язуються з вірусною ДНК.
Неструктурні білки є продуктами ранніх генів. Е1 та Е2 задіяні у вірусній реплікації та регуляції генів (Е2). Е5, Е6 та Е7 стимулюють клітинну ДНК-реплікацію. Е4 звязуються зі специфічними структурами клітинного цитоскелету.
Ліпіди та вуглеводи відсутні.
Головними антигенами є капсидні білки. Білок L1 містить видоспецифічний антигенний домен, L2 – антигенний епітоп групової специфічності.
Папіломавіруси є дуже видоспецифічними. Уражують хребетних.
Тропізм – епітелій шкіри і слизових оболонок.
Інфікують лише проліферуючі епітеліоцити, інтенсивна реплікація і вихід вірусу відбувається у диференційованих кератиноцитах, що знаходяться у верхніх шарах тканини.
Передача: контактно, статевим шляхом.
Спричиняють утворення доброякісних пухлин епітеліального походження (бородавки, папіломи). Проте такі утворення, викликані певними видами папілома вірусів, мають високий ризик злоякісного розвитку. На даний час доведено зв'язок багатьох онкологічних захворювань епітеліальних тканин із різними видами HPV. Часто геном цих вірусів вбудовується в ДНК клітини. Продукти вірусних генів Е6 та Е7 претворюють клітини на ракові. Білки Е6 зв'язують та деградують білок р53, стимулюють теломеразну активність, Е7 – активують циклони Е та А.
Специфічна профілактика папіломавірусної інфекції не розроблена. Вакцинація білками вірусного капсиду, отриманими на основі рекомбінантної ДНК технології, була обнадійливою. Складність полягає в тому, що вакцина повинна містити багато типів вірусу, тому перехресний (межтиповий) захист відсутній.
Рішення задачі специфічної профілактики ускладнюється відсутністю типоспецифічних діагностичних серологічних тестів і неможливістю розмноження папіломавірусів в культурі клітин. ПВ ВРХ включають різні типи, що інфікують шкірні покриви (BPV1 і BPV2) або слизові (BPV4). За структурою геному ПВ ВРХ подібний ПВ людини.
Папіломавіруси (Papillomaviridae) — родина вірусів, що були ідентифіковані на початку 20 століття, коли було показано, що бородавки (папіломи) можуть передаватися між інивідуумами за допомогою виділеного інфекційного агенту.
В 1935 році Френсіс Пейтон Раус, який до того продемонстрував існування віруса саркоми у курей, також показав, що папіломавірус може викликати рак шкіри у заражених кролів. Це було першою демонстрацією того, що вірус може викликати рак у ссавців.
Papillomaviridae
ПодсемействаI: дцДНК-вирусы
Папилломавирусы[2] (лат. Papillomaviridae) — семейство вирусов, ранее — род вирусов из семейства паповавирусов. Относится к I группе классификации вирусов по Балтимору.
Размеры вириона около 55 нм. Размножается в ядре клеток-хозяев.
Большинство представителей обладает онкогенными свойствами и вызывает доброкачественные опухоли (папилломы) человека, кроликов, коров, оленей, собак и кошек, лошадей, овец, крыс. Показана этиологическая роль папилломавируса в развитии широко распространённого, передающегося половым путём, рака шейки матки[3].
Папилломавирусы имеют видоспецифические и некоторые типоспецифические антигены. Не культивируются в клеточных системах. Вирус папилломы человека выявляется в кератоцитах в виде эписомы, 50—200 копий на поражённую клетку. Процессы продуктивной инфекции и интеграции не наблюдаются.
Папилломавирус обнаружен пышнохвостого лесного хомяка (en:Neotoma cinerea), жившего в Северной Америке 27 тыс. л. н.[4]
Сначала виды в роде Papillomavirus назывались по животным, у которых они могут вызвать заболевания, например, в 2002 году в состав рода входило всего 7 видов[5]:
В 2003 году произведена ревизия семейства и эти 7 видов разделены на 16 родов с 43 видами. После неё название каждого из родов состоит из двух частей: латинизированной буквы греческого (или другого) алфавита и общей части -papillomavirus[6]. В 2010 году произведено переименование входящих в семейство видов: теперь их научное название состоит из названия рода и порядкового номера, не совпадающего с ранее принятой нумерацией видов. Было увеличено количество родов и видов, и, так как количество родов превысило количество букв греческого алфавита, начиная с 25 рода добавлена приставка dyo-[7], а затем и приставка treis-. В 2017 году семейство разделено на 2 подсемейства: Firstpapillomavirinae, куда вошли все старые и 3 новых рода, и Secondpapillomavirinae с одним новым родом Alefpapillomavirus[8].
На апрель 2018 года в подсемейство Firstpapillomavirinae включают 52 рода[9]:
Самым обширным является род Gammapapillomavirus, в который включают 27 видов: Gammapapillomavirus 1—27. Многие же рода монотипные[9].
Папилломавирусы (лат. Papillomaviridae) — семейство вирусов, ранее — род вирусов из семейства паповавирусов. Относится к I группе классификации вирусов по Балтимору.
Размеры вириона около 55 нм. Размножается в ядре клеток-хозяев.
Большинство представителей обладает онкогенными свойствами и вызывает доброкачественные опухоли (папилломы) человека, кроликов, коров, оленей, собак и кошек, лошадей, овец, крыс. Показана этиологическая роль папилломавируса в развитии широко распространённого, передающегося половым путём, рака шейки матки.
Папилломавирусы имеют видоспецифические и некоторые типоспецифические антигены. Не культивируются в клеточных системах. Вирус папилломы человека выявляется в кератоцитах в виде эписомы, 50—200 копий на поражённую клетку. Процессы продуктивной инфекции и интеграции не наблюдаются.
Папилломавирус обнаружен пышнохвостого лесного хомяка (en:Neotoma cinerea), жившего в Северной Америке 27 тыс. л. н.
乳頭瘤病毒科(Papillomaviridae)是雙鏈DNA病毒的一科,該類病毒會導致腫瘤,但大多情況不會成為癌症,僅有少數種類會導致子宮頸癌。
下有一屬:











