Haptofity (Haptophyta) – typ (gromada) glonów z supergrupy Chromalveolata (Chromista).
Haptofity to grupa glonów o nie do końca ustalonej pozycji systematycznej. W XX w. organizmy te grupowano jako Coccolithinae wśród złotowiciowców i określano polską nazwą wiciowce wapienne[1]. Na początku XXI w. większość badaczy umieszczała je wśród chromistów (bądź chromalweolatów) jako odrębną linię ewolucyjną niezwiązaną ani ze stramenopilami, ani z alweolatami. W podobnej sytuacji znajdują się kryptomonady i czasem te dwie linie są łączone jako Hacrobia[2]. Jedne badania z XXI w. potwierdzają bliskość haptofitów i kryptomonad[3][4], podczas gdy inne wskazują na ich dużą odrębność, potwierdzając bliskość haptofitów i pozostałych chromalweolatów (oraz ameb Rhizaria), a zbliżając kryptomonady do linii rozwojowej obejmującej rośliny[5].
W jednym z systemów zaproponowanych przez Cavaliera-Smitha haptofity mają rangę infrakrólestwa Haptista, które jest monotypowe, zawierając jedynie typ Haptophyta. Jest on tam równoważny infrakrólestwu Heterokonta, z którym tworzy podkrólestwo Chromobiota, które z kolei z podkrólestwem Cryptista (zawierającym szeroko ujmowane kryptomonady) tworzy królestwo Chromista[6].
Od drugiej połowy XX w. haptofity dzieli się na dwie główne grupy:
W rejestrze AlgaeBase do drugiej połowy 2012 r. zgromadzono informacje o 385 gatunkach Coccolithophyceae, 15 gatunkach Pavlovophyceae i 125 gatunkach skupionych w kilku rodzinach nieprzypisanych do żadnej z tych grup[7]. Za najbardziej liczebnego przedstawiciela haptofitów kokolitotwórczych uważana jest Emiliania huxleyi[8].
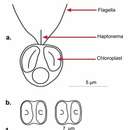
Haptofity mają postać wiciowców, często, ale nie zawsze ruchliwych – jednokomórkowych monad lub ich kolonii, np. w formie nitkowatej lub palmelloidalnej, przy czym u niektórych gatunków stadium kolonijne występuje na przemian z monadalnym. Zdarzają się też postacie pełzakowate. Pod względem rozmiaru niektóre sytuują się wśród organizmów nanoplanktonowych, podczas gdy inne tworzą widoczne gołym okiem kolonie (Phaeocystis). Charakterystycznym, niewystępującym poza tą grupą organellum jest haptonema, czyli wyrostek przypominający wić. Może on przybierać różne postacie i długości (u Pavlovophyceae i niektórych Coccolithophyce raczej krótka, a u Isochrysidales może wręcz być jej brak). Wici u Coccolithophyceae równe lub jedna nieco dłuższa od drugiej, zwykle dwie, umiejscowione mniej lub bardziej szczytowo. Natomiast u Pavlovophyceae jedna wić jest zdecydowanie krótsza od drugiej, a położone są po przeciwnych końcach komórki. Przednia wić Pavlovophyceae pokryta jest wyrostkami. U Coccolithophyce wici mogą zawierać substancję autofluoroscencyjną, czego nie ma u Pavlovophyceae. Z kolei u tych ostatnich mogą występować niezwiązane z wiciami plamki oczne[9]. Retikulum endoplazmatyczne położone pod samą błoną komórkową, wnikając do haptonemy, jednak pozostawiając lukę u nasady wici. Aparat Golgiego położony blisko kinetosomów wici i haptonemy. Jego cysterny leżą prostopadle do dłuższej osi komórki, układając się wachlarzowo. Aparat Golgiego u gatunków wytwarzających pancerzyki jest zaangażowany w to działanie. Liczba chloroplastów od 1 do 4 (u niektórych zanikły)[9], choć podawany jest też zakres 1–2[10]. W chloroplastach znajduje się pirenoid. Zewnętrzna błona chloroplastów przechodzi w zewnętrzną błonę jądra. Barwniki fotosyntetyczne to chlorofile a, c1, c2 i c3 (ostatni u Coccolithophyceae), β-karoten, diadinoksantyna i diatoksantyna. Głównym produktem zapasowym często jest chryzolaminaryna[9][10]. Chloroplasty haptofitów powstały na drodze wtórnej endosymbiozy, pochodząc od krasnorostów. Są spokrewnione z chloroplastami kryptomonad i stramenopili[8]. Realizowane strategie pokarmowe to autotrofizm, miksotrofizm i heterotrofizm[9][10].
Wielu przedstawicieli podklasy Coccolithophyce wytwarza organiczne płytki, wśród nich zaś grupa określana jako kokolitofory, pokrywa je wapiennymi skorupkami zwanymi kokolitami (stąd nazwa)[9][10]. Na podstawie badań filogenetycznych sugeruje się, że jest to historycznie cecha powszechna w tej podklasie, a niezdolność do tworzenia kokolitów jest zjawiskiem wtórnym[11]. Płytki pancerzyka haptofitów mają różną budowę, a często ten sam gatunek wytwarza różne ich typy w zależności od fazy ploidii. Za najpierwotniejszą cechę uważa się brak jakichkolwiek płytek, typowy dla podklasy Pavlovoohyceae. Niektóre Coccolithophyceae (np. Chrysochromulina) wytwarzają jedynie płytki zbudowane z substancji organicznej, bez kokolitów. Inne wytwarzają kokolity w fazie diploidalnej, a w fazie haploidalnej jedynie płyki organiczne. Nie jest jasne, czy jest to cecha pierwotna ewolucyjnie czy też wtórna – rozważane są też obie opcje, np. według jednej z hipotez Emiliania zachowała ją po swoich przodkach, podczas gdy przodkowie Pleurochrysis wytwarzali kokolity w obu fazach, a następnie utracili tę zdolność. Dość często występuje model, w którym w fazie haploidalnej wytwarzane są inne rodzaje kokolitów (nanolity, holokokolity) niż w fazie diploidalnej (heterokokolity). Typowe kokolity są zbudowane z kalcytu, podczas gdy nanolity mogą być aragonitowe[12].
Wśród haptofitów zdarzają się gatunki słodkowodne, jednak zdecydowana większość żyje w wodach słonych, zarówno przybrzeżnych, jak i pełnomorskich[10]. Haptofity nierzadko osiągają bardzo duże zagęszczenia, zajmując istotną pozycję w fitoplanktonie niemal wszystkich środowisk morskich. Czasem powodują zakwity wód. Ze względu na tworzenie kokolitów odgrywają znaczącą rolę w obiegu węgla i wapnia, a ich szczątki od późnego triasu są ważnym elementem wapiennych osadów. Pośrednio, przez wiązanie dwutlenku węgla, ma to wpływ na kształtowanie klimatu. Rozległe zakwity są widoczne na zdjęciach satelitarnych. Mają one wpływ na biocenozy oceaniczne, jako że mogą być toksyczne dla innych organizmów[13]. Pomory ryb wywołane zakwitami wód w słonawych duńskich jeziorach powodowanymi przez Prymnesium parvum opisywano już w pierwszej połowie XX w[14], natomiast jest możliwe, że podobne oddziaływanie na ekosystem od czasu do czasu haptofity wywierały przez ok. 200 milionów lat[13]. Rodzaj Phaeocystis jest kosmopolityczny, a w chłodniejszych rejonach oceanu może dominować w fitoplanktonie[15].
Wpływ haptofitów na klimat, zwłaszcza wytwarzających kokolity, jest złożony. Kokolity otaczające komórki odbijają światło w charakterystyczny sposób, co w przypadku zakwitu daje wrażenie białej wody. Ma to pewien wpływ na albedo obszaru objętego zakwitem. Według modeli rozpraszania światła słonecznego docierającego do oceanu wynika, że istnienie warstwy zajętej przez kokolitofory zwiększa albedo oceanu, sprawiając że najpłytsza jego warstwa (kilka metrów) jest jaśniejsza, a głębsza ciemniejsza niż wynikałoby to z samego pochłaniania przez glony. Może to prowadzić do redukcji strefy eufotycznej, preferowanej przez organizmy fotosyntetyzujące, z ponad 24 m do 8 m w strefie zakwitu. Bardziej naświetlona warstwa przypowierzchniowa ogrzewa się, a mniej naświetlona ochładza, co powoduje powstanie stratyfikacji termicznej i zmniejsza mieszanie się wody. Jednocześnie strefa rozjaśniona przez światło odbijane od kokolitów zwiększa strefę fotoinhibicji. Sam więc wpływ kokolitów (nawet bez wpływu toksycznego innych substancji wytwarzanych przez haptofity) ostatecznie może zmniejszyć produkcję pierwotną strefy objętej zakwitem. Jednocześnie jednak ze względu na skalę przestrzenną i czasową takich zakwitów, ich wpływ na całkowite albedo oceanu uważany jest za nieistotny[16]. Do wytwarzania wapiennych struktur zużywany jest dwutlenek węgla. W związku z tym organizmy je produkujące w pewnych warunkach (np. dostępności substancji odżywczych) mogą zwiększać tę produkcję przy wzroście stężenia CO2. W przypadku organizmów wodnych jednak może zachodzić przeciwny proces, tj. rozpuszczanie struktur wapiennych przez CO2 powodujący zakwaszanie wód. Ze względu na specyficzny sposób wytwarzania kokolitów, zakwaszenie ma słabszy wpływ, przez co w ciągu XIX i XX w. średnia masa kokolitów wzrosła o 40%, a eksperymenty w warunkach laboratoryjnych sugerują, że kokolitofory mogą rozwijać się lepiej przy nieco podwyższonym stężeniu CO2[17]. Badania w różnych warunkach jednak dają zróżnicowane wyniki. Kokolitofory zasadniczo dostosowują się do różnych warunków, o czym świadczy stan kokolitów zdeponowanych w czasach zlodowaceń, gdy stężenie dwutlenku węgla było niskie[18].
Haptofity (Haptophyta) – typ (gromada) glonów z supergrupy Chromalveolata (Chromista).

.png)