Les criptofícies (Cryptophyceae) són una petita classe d'organismes amb 24 gèneres que es troben en aigües marines i continentals. Són organismes unicel·lulars nus, amb dos flagels quasi iguals. Les cèl·lules són ovals i aplanades amb una mida de prop d'uns 10-50 μm. Poden aparèixer estats palmeloides o cocoides. Algunes espècies formen zooxanteles, és dir viuen com a simbionts d'animals.
Presenten un o dos plasts, amb pirenoides o sense. Els plastidis presenten els til·lacoides agrupats en parells, sense lamel·la cenyidora. Amb clorofil·la a i c; presenten α-carotens i xantofil·las, aloxantina dominant, tanmateix falta el β-carotè. Posseeixen pigments ficobilínics (ficocianina i ficoeritrina), però no es presenten en ficobilisomes, sinó a l'espai intratil·lacoidal, s'ha postulat el seu origen en la simbiosi amb cianòfits. com a material de reserva fan servir midó veritable, extraplastidial, associat amb plasts i el nucli.
Presenten dos flagels anteriors quasi iguals que surten d'un reservori entapissat per tricocists. Tots dos són pleuronemàtics. Hi pot haver una paret cel·lular de cel·lulosa, presenten un periplast de tres capes, també pot presentar plaques poligonals externes. Els mitocondris tenen crestes planes, i la mitosi és oberta. La reproducció és per bipartició, mitjançant zoòspores i cists. En algun cas s'ha observat també reproducció sexual.
En altres projectes de Wikimedia:Skrytěnky, též kryptomonády (Cryptophyceae, Cryptomonada), v širším smyslu také Cryptophyta či Cryptista, jsou jednobuněční eukaryotičtí bičíkovci, obvykle dnes řazení do říše Chromista[1] nebo považovaní za eukaryota incertae sedis.[2] Podle některých nových fylogenetických studií jsou blízkými příbuznými kmene Haptista/Haptophyta,[1] jiné molekulární analýzy ukazují na umístění skrytěnek a jim blízce příbuzných katablefarid jako sesterské skupiny archaeplastid.[3] Žijí v sladké i slané vodě, v ČR se objevují již časně zjara, kdy se ještě ostatní fytoplankton nestačil pomnožit. V antarktických jezerech mohou tvořit velkou část biomasy; stejně jako v hlubinných oligotrofních jezerech. Známo je celkem asi 20 rodů s 200 druhy.[4]
Skrytěnky jsou drobní protisté s dvěma bičíky, jedním kratším a druhým delším. Jsou sice obvykle autotrofní – získávají organické látky fotosyntézou – ale na druhou stranu se mnohé druhy vyživují i heterotrofně pomocí fagocytózy a osmotrofie. K fotosyntéze jim slouží chloroplasty vzniklé sekundární endosymbiózou (pohlcením) jisté ruduchy. Uvnitř chloroplastů bývá pyrenoid a někdy i stigma; navíc jako zbytek po endosymbióze i nukleomorf, čily zakrnělý genom oné červené řasy. Obsahují fotosyntetická barviva chlorofyl a a c2, karoteny, xanthofyl alloxanthin a fykobiliny (fykoerythrin nebo fykocyanin). Zásobní látkou je škrob a oleje uložené v cytoplazmě. Rozmnožují se hlavně nepohlavně (dělením buněk), bylo však popsáno i pohlavní rozmnožování.[4]
Některé skrytěnky produkují toxiny, byly zaznamenány i otravy ryb přičítané druhu Cyanomonas americana.[4]
Do dříve monotypického kmene Cryptophyta s jedinou třídou Cryptophyceae či do jeho těsné blízkosti byly v poslední době na základě fylogenetických analýz přiřazeny nové skupiny eukaryotních organismů (goniomonády, katablefaridy, palpitomonády, pikomonády, telonemidi a část slunivek (Endohelea)), což si vyžádalo několikerou změnu systému. Protože takto rozšířený kmen již neobsahuje pouze fotoautotrofní organismy, nedoporučuje Thomas Cavalier-Smith používat dosavadní jméno Cryptophyta a navrhuje namísto toho název Cryptista.[1]
Aktuální (2015) klasifikace:[1]
Skrytěnky, též kryptomonády (Cryptophyceae, Cryptomonada), v širším smyslu také Cryptophyta či Cryptista, jsou jednobuněční eukaryotičtí bičíkovci, obvykle dnes řazení do říše Chromista nebo považovaní za eukaryota incertae sedis. Podle některých nových fylogenetických studií jsou blízkými příbuznými kmene Haptista/Haptophyta, jiné molekulární analýzy ukazují na umístění skrytěnek a jim blízce příbuzných katablefarid jako sesterské skupiny archaeplastid. Žijí v sladké i slané vodě, v ČR se objevují již časně zjara, kdy se ještě ostatní fytoplankton nestačil pomnožit. V antarktických jezerech mohou tvořit velkou část biomasy; stejně jako v hlubinných oligotrofních jezerech. Známo je celkem asi 20 rodů s 200 druhy.
The cryptomonads (or cryptophytes)[1] are a group of algae,[2] most of which have plastids. They are common in freshwater, and also occur in marine and brackish habitats. Each cell is around 10–50 μm in size and flattened in shape, with an anterior groove or pocket. At the edge of the pocket there are typically two slightly unequal flagella.
Some may exhibit mixotrophy.[3]

Cryptomonads are distinguished by the presence of characteristic extrusomes called ejectosomes, which consist of two connected spiral ribbons held under tension.[4] If the cells are irritated either by mechanical, chemical or light stress, they discharge, propelling the cell in a zig-zag course away from the disturbance. Large ejectosomes, visible under the light microscope, are associated with the pocket; smaller ones occur underneath the periplast, the cryptophyte-specific cell surrounding.[5][6]
Except for the class Goniomonadea, which lacks plastids entirely,[7] and Cryptomonas paramecium (previously called Chilomonas paramecium), which has leucoplasts, cryptomonads have one or two chloroplasts. These contain chlorophylls a and c, together with phycobiliproteins and other pigments, and vary in color (brown, red to blueish-green). Each is surrounded by four membranes, and there is a reduced cell nucleus called a nucleomorph between the middle two. This indicates that the plastid was derived from a eukaryotic symbiont, shown by genetic studies to have been a red alga.[8] However, the plastids are very different from red algal plastids: phycobiliproteins are present but only in the thylakoid lumen and are present only as phycoerythrin or phycocyanin. In the case of Rhodomonas, the crystal structure has been determined to 1.63 Å;[9] and it has been shown that the alpha subunit bears no relation to any other known phycobiliprotein.
A few cryptomonads, such as Cryptomonas, can form palmelloid stages, but readily escape the surrounding mucus to become free-living flagellates again. Some Cryptomonas species may also form immotile microbial cysts—resting stages with rigid cell walls to survive unfavorable conditions. Cryptomonad flagella are inserted parallel to one another, and are covered by bipartite hairs called mastigonemes, formed within the endoplasmic reticulum and transported to the cell surface. Small scales may also be present on the flagella and cell body. The mitochondria have flat cristae, and mitosis is open; sexual reproduction has also been reported.


The first mention of cryptomonads appears to have been made by Christian Gottfried Ehrenberg in 1831,[10] while studying Infusoria. Later, botanists treated them as a separate algae group, class Cryptophyceae or division Cryptophyta, while zoologists treated them as the flagellate protozoa order Cryptomonadina. In some classifications, the cryptomonads were considered close relatives of the dinoflagellates because of their (seemingly) similar pigmentation, being grouped as the Pyrrhophyta. Cryptomonad chloroplasts are closely related to those of the heterokonts and haptophytes, and the three groups were united by Cavalier-Smith as the Chromista. However, the case that the organisms themselves are closely related was counter-indicated by the major differences in cell organization (ultrastructural identity), suggesting that the three major lineages assigned to the chromists had acquired plastids independently, and that chromists are polyphyletic. The perspective that cryptomonads are primitively heterotrophic and secondarily acquired chloroplasts, is supported by molecular evidence [11] Parfrey et al. and Burki et al. placed Cryptophyceae as a sister clade to the Green Algae.[12][13] The sister group to the cryptomonads is likely the kathablepharids (also referred to as katablepahrids), a group of flagellates that also have ejectisomes.[14]
One suggested grouping is as follows: (1) Cryptomonas, (2) Chroomonas/Komma and Hemiselmis, (3) Rhodomonas/Rhinomonas/Storeatula, (4) Guillardia/Hanusia, (5) Geminigera/Plagioselmis/Teleaulax, (6) Proteomonas sulcata, (7) Falcomonas daucoides.[15]
The cryptomonads (or cryptophytes) are a group of algae, most of which have plastids. They are common in freshwater, and also occur in marine and brackish habitats. Each cell is around 10–50 μm in size and flattened in shape, with an anterior groove or pocket. At the edge of the pocket there are typically two slightly unequal flagella.
Some may exhibit mixotrophy.
Cryptophyta, Cryptomonada o criptofitas es un grupo pequeño de algas unicelulares con unas 200 especies que viven en aguas marinas y continentales.[3] Son miembros importantes del fitoplancton y se pueden encontrar en aguas estancadas, soportando moderados niveles de contaminación. Son especialmente abundantes en aguas frías tales como los lagos de alta montaña y en aguas árticas y antárticas. En general, las criptofitas son mixótrofas, es decir, capaces tanto de la fotosíntesis como de la fagotrofia. Presentan una cubierta rígida, una invaginación ventral (de ahí el nombre de crypto, que significa oculto) de la que salen dos flagelos y varias filas de eyectosomas, de función defensiva. Algunas especies forman zooxantelas, es decir, viven como simbióticas de animales.
Las criptofitas se pueden encontrar en todos los mares, en agua dulce y en aguas estancadas, incluyendo las charcas intersticiales de las playas de arena. Aproximadamente la mitad de las especies son de agua dulce y la otra mitad marinas. Son especialmente abundantes en aguas frías y parecen ser muy sensibles a la cantidad de luz que reciben, a menudo viviendo en las zonas más profundas de los claros lagos oligotróficos más septentrionales y de alta montaña.
Son capaces de sobrevivir durante el invierno en las aguas cubiertas de nieve o hielo, en este caso, desplazándose a la zona más superficial con el objeto de recibir la suficiente cantidad de luz. La supervivencia en estas condiciones de escasez de luz requiere tanto de un sistema fotosintético altamente eficiente como unas reducidas tasas de respiración celular en las bajas temperaturas. Cuando el hielo se funde y la cantidad de luz aumenta considerablemente, las criptofitas se desplazan a las aguas más profundas para evitar el exceso de luz. Son también las especies dominantes en los lagos de agua dulce de la Antártida.[4]
Las criptofitas son organismos principalmente mixotrofos, capaces de realizar la fotosíntesis, si bien algunos miembros son totalmente heterótrofos. La ingestión fagotrófica de bacterias les proporciona la fuente de fósforo y nitrógeno que necesitan en condiciones de escasez de nutrientes.
Las criptofitas son unicelulares y biflageladas, con células ovales y aplanadas y con un tamaño de alrededor de 10-50 μm. Algunas se presentan en formas cocoides o palmeloides (grupos de células rodeadas de una cubierta de mucílago) y existe un género filamentoso. Por ejemplo, Cryptomonas puede presentar un estado palmeloide del que puede liberarse fácilmente para volver a ser una forma móvil. También pueden formar quistes, rodeados de una pared celular y en animación suspendida, que les permiten sobrevivir en condiciones desfavorables.
El cuerpo celular de las criptofitas es asimétrico, organizado dorsiventralmente. Las células tienen una invaginación ventral de la que salen dos flagelos anteriores casi iguales. Los flagelos presentan fibrillas o mastigonemas, que en el más corto están organizadas en una fila, pero en el más largo lo están en dos hileras opuestas. Los flagelos se insertan de forma asimétrica en el lado derecho de la invaginación, lo que provoca una rotación alrededor del eje longitudinal cuando nadan.
Las criptofitas no presentan una verdadera pared celular. La cubierta específica que rodea a la criptofita es un periplasto de dos capas, una interna y otra externa a la membrana plasmática. La capa externa es fibrosa, mientras que la interna presenta placas proteináceas. Adicionalmente pueden presentar pequeñas placas poligonales externas tanto sobre la superficie celular como sobre los flagelos.[5][6]
Presentan eyectosomas que emplean para la defensa, pues pueden ser disparados al exterior. Consisten en dos filamentos espirales mantenidos bajo tensión.[7] Si la célula recibe un estímulo, ya sea de tipo mecánico, químico o luminoso, los eyectosomas son descargados, propulsando a la célula lejos de la perturbación en una trayectoria en zig-zag. Los eyectosomas grandes, visibles al microscopio óptico, están asociados a la invaginación; los más pequeños se sitúan por debajo de la periplasto.
Algunas criptofitas presentan una mancha ocular, un orgánulo sensible a luz, constituido por una sola hilera de gránulos y que puede estar situado dentro de la membrana que rodea al cloroplasto o en otros lugares de la célula. En todas las especies de agua dulce está presente una vacuola contráctil para la regulación de la presión osmótica localizada en el vértice de la célula. Las mitocondrias tienen crestas planas.
La reproducción es asexual, por simple división longitudinal y la mitosis es abierta. Las especies sésiles o coloniales producen zoosporas. La reproducción sexual ha sido observada recientemente.
Para las características de los plastos, véase la clase Cryptophyceae.
Se distinguen tres órdenes en dos clases.[3]
Cavalier-Smith las incluye en el propuesto reino Chromista en el grupo Cryptista, integrado por criptomonadas, goniomonadas, katablefáridas y otros organismos heterótrofos relacionados.[8][9] Sin embargo varios especialistas sugieren que son necesarios más estudios filogenéticos para definir la relación entre Cryptophyta y Chromista.
|número-autores= (ayuda) Cryptophyta, Cryptomonada o criptofitas es un grupo pequeño de algas unicelulares con unas 200 especies que viven en aguas marinas y continentales. Son miembros importantes del fitoplancton y se pueden encontrar en aguas estancadas, soportando moderados niveles de contaminación. Son especialmente abundantes en aguas frías tales como los lagos de alta montaña y en aguas árticas y antárticas. En general, las criptofitas son mixótrofas, es decir, capaces tanto de la fotosíntesis como de la fagotrofia. Presentan una cubierta rígida, una invaginación ventral (de ahí el nombre de crypto, que significa oculto) de la que salen dos flagelos y varias filas de eyectosomas, de función defensiva. Algunas especies forman zooxantelas, es decir, viven como simbióticas de animales.
Nielulevät (Cryptophyta) on yksisoluisten, merten ja sisävesien levien ryhmä, joka nykyisin luokitellaan Chromalveolata-kuntaan kuuluvaksi kaareksi. Nielulevälajien määräksi on arvioitu 60-200. Niiden nimitys tulee solun etupäässä olevasta ontelosta, jota kutsutaan nieluksi.
Nielulevien kehityshistoria on ollut pitkään epäselvä. Solun hienorakenteen perusteella otaksutaan nielulevien kehityslinjan syntyneen, kun varhainen Chromalveolata-kuntaan kuulunut uintisiimallinen eliö otti soluunsa endosymbiontiksi punalevien kehityslinjaan kuuluneen yksisoluisen eliön. Punalevä oli puolestaan aiemmin evoluution kuluessa ottanut soluunsa endosymbionttisen syanobakteerin, minkä vuoksi se kykeni yhteyttämään. Sittemmin symbionttina ollut punalevä surkastui ja muuttui nielulevän kloroplasteiksi eli viherhiukkasiksi. Yhdessä nieluleväsolussa on yleensä vain yksi hyvin suuri viherhiukkanen. Nykyään muinaisesta punalevästä on enää havaittavissa viherhiukkasen ympärillä oleva ylimääräinen kalvopari ja tämän uloimman kalvoparin alla havaittavissa oleva surkastunut punalevän tumarakenne eli nukleomorfi. Nieluleväkloroplasteilla on tämän vuoksi ympärillään neljä kalvoa.
Vaikka useimmilla nielulevillä on viherhiukkasia, ne kykenevät myös syömään ravinnokseen bakteereja, joten ne ovat sekä autotrofeja että heterotrofeja. Solut ovat 10–50 mikrometrin kokoisia, muodoltaan litteitä ja soikeita, ja niiltä puuttuu jäykkä soluseinä. Solussa on kaksi uintisiimaa. Nielulevillä on erityisominaisuutena ulkopinnalla olevia spiraaliin muotoon kiertyneitä rihmoja eli ejektiosomeja, jotka toimivat jännitetyn jousen tavoin. Jos levää ärsytetään, ejektiosomit oikenevat ja levä sinkoaa toisaalle.
Nielulevät tuottavat yhteyttämistuotteenaan tärkkelystä ja samankaltaisia aineita. Ne ovat ravintoa muun muassa eläinplanktonille. Vaikka nieluleviä esiintyy niin makeissa kuin suolaisissakin vesissä, ovat ne tavallisimpia viileissä ja tummissa humuspitoisissa vesissä, joissa ne pystyvät yhteyttämään vähässäkin valossa viherhiukkasten sisältämän fykosyaniinin ja fykoerytriinin avulla. Kyseisiä pigmenttejä tavataan edelleenkin punalevillä ja syanobakteereilla. Chilomonas-suvun lajeilla ei ole viherhiukkasia, vaan ne käyttävät esimerkiksi bakteereja ravintonaan; tarvittaessa ne voivat tulla toimeen pelkillä mineraaleilla.
Nielulevät (Cryptophyta) on yksisoluisten, merten ja sisävesien levien ryhmä, joka nykyisin luokitellaan Chromalveolata-kuntaan kuuluvaksi kaareksi. Nielulevälajien määräksi on arvioitu 60-200. Niiden nimitys tulee solun etupäässä olevasta ontelosta, jota kutsutaan nieluksi.
Nielulevien kehityshistoria on ollut pitkään epäselvä. Solun hienorakenteen perusteella otaksutaan nielulevien kehityslinjan syntyneen, kun varhainen Chromalveolata-kuntaan kuulunut uintisiimallinen eliö otti soluunsa endosymbiontiksi punalevien kehityslinjaan kuuluneen yksisoluisen eliön. Punalevä oli puolestaan aiemmin evoluution kuluessa ottanut soluunsa endosymbionttisen syanobakteerin, minkä vuoksi se kykeni yhteyttämään. Sittemmin symbionttina ollut punalevä surkastui ja muuttui nielulevän kloroplasteiksi eli viherhiukkasiksi. Yhdessä nieluleväsolussa on yleensä vain yksi hyvin suuri viherhiukkanen. Nykyään muinaisesta punalevästä on enää havaittavissa viherhiukkasen ympärillä oleva ylimääräinen kalvopari ja tämän uloimman kalvoparin alla havaittavissa oleva surkastunut punalevän tumarakenne eli nukleomorfi. Nieluleväkloroplasteilla on tämän vuoksi ympärillään neljä kalvoa.
Vaikka useimmilla nielulevillä on viherhiukkasia, ne kykenevät myös syömään ravinnokseen bakteereja, joten ne ovat sekä autotrofeja että heterotrofeja. Solut ovat 10–50 mikrometrin kokoisia, muodoltaan litteitä ja soikeita, ja niiltä puuttuu jäykkä soluseinä. Solussa on kaksi uintisiimaa. Nielulevillä on erityisominaisuutena ulkopinnalla olevia spiraaliin muotoon kiertyneitä rihmoja eli ejektiosomeja, jotka toimivat jännitetyn jousen tavoin. Jos levää ärsytetään, ejektiosomit oikenevat ja levä sinkoaa toisaalle.
Nielulevät tuottavat yhteyttämistuotteenaan tärkkelystä ja samankaltaisia aineita. Ne ovat ravintoa muun muassa eläinplanktonille. Vaikka nieluleviä esiintyy niin makeissa kuin suolaisissakin vesissä, ovat ne tavallisimpia viileissä ja tummissa humuspitoisissa vesissä, joissa ne pystyvät yhteyttämään vähässäkin valossa viherhiukkasten sisältämän fykosyaniinin ja fykoerytriinin avulla. Kyseisiä pigmenttejä tavataan edelleenkin punalevillä ja syanobakteereilla. Chilomonas-suvun lajeilla ei ole viherhiukkasia, vaan ne käyttävät esimerkiksi bakteereja ravintonaan; tarvittaessa ne voivat tulla toimeen pelkillä mineraaleilla.
Les Cryptophytes (Embranchement Cryptophyta, Classe Cryptophyceae) sont des organismes vivants unicellulaires, photosynthétiques pour la plupart. Ce petit taxon (200 espèces) est relativement homogène et de nombreux caractères lui sont propres.
Le mot est composé de deux mots grecs : de κρυπτός, kruptós (« caché ») et de φυτόν, phyton (« végétal »).
Les Cryptophytes se rencontrent dans tous les types de milieux aquatiques : des milieux océaniques aux eaux douces en passant par les eaux interstitielles des milieux terrestres humides.
Certaines espèces sont devenues parasites intestinaux de métazoaires. Quelques-unes sont des endosymbiotes de Dinophytes. Certaines vivent de manière symbiotique dans l'ectoplasme des Radiolaires.
Ordre Goniomonadales
Les Cryptophytes (Embranchement Cryptophyta, Classe Cryptophyceae) sont des organismes vivants unicellulaires, photosynthétiques pour la plupart. Ce petit taxon (200 espèces) est relativement homogène et de nombreux caractères lui sont propres.
Le criptoficee (Cryptophyceae Fritsch, 1927) sono alghe unicellulari che fanno parte del nanoplancton; sono l'unica classe della divisione Cryptophyta.[1][2]
Possiedono due flagelli lunghi all'incirca come la cellula stessa. Al Guiry, M.D. & Guiry, G.M. 2019di sotto della membrana esterna vi sono delle placche il cui numero è un importante carattere tassonomico.
I pigmenti fotosintetici che contengono sono: clorofilla a, c2, ficobiline, ficoeritrina, ficocianina.
In acqua dolce si trovano i generi Cryptomonas e Rhodomonas.
Le criptoficee (Cryptophyceae Fritsch, 1927) sono alghe unicellulari che fanno parte del nanoplancton; sono l'unica classe della divisione Cryptophyta.
Kryptomonady, kryptomonadowe, kryptofity (Cryptophyta, Cryptista, Cryptomonadea, Cryptophyceae) – glony zaliczane niegdyś do gromady tobołków, jedna z grup wiciowców. Większość kryptomonad to organizmy jednokomórkowe, choć czasami można spotkać je w postaci palmelloidalnych kolonii. W różnych systemach taksonomicznych ta sama grupa organizmów może mieć różną rangę taksonomiczną, a kilka kolejnych taksonów może być monotypowe.
Komórki płaskie, nagie, z dwiema wiciami (rzadziej jedną lub czterema). Okryte mało elastycznym peryplastem o różnej grubości. Dominuje postać wiciowca (monady) o kształcie asymetrycznym i mało zmiennym – jajowatym, gruszkowatym, cylindrycznym[1]. Mogą tworzyć się palmelloidalne kolonie[2]. Komórki mniej lub bardziej spłaszczone grzbietobrzusznie. Po stronie brzusznej zagłębienie zwane gardzielą. Wici osadzone w bruździe gardzieli. W gardzieli znajdują się ułożone w regularnych rzędach lub pierścieniach trychocysty[1]. Na wiciach mastygonemy[2]. Chromatofory od brązowych, przez czerwonobrunatne do niebieskozielonych, otoczone czterema błonami, między którymi znajduje się nukleomorf[3]. Zwykle duże, przyścienne, od jednego do kilku. Barwniki fotosyntetyczne to chlorofile a i c oraz fikobiliny oraz β-karoten i ksantofile.
Dodatkowe błony oraz obecność nukleomorfu w chloroplastach świadczą o tym, że powstały one na drodze wtórnej endosymbiozy, gdy przodek kryptomonad (a być może wszystkich Chromalveolata) wchłonął pierwotną komórkę krasnorostu[3].
Produktem fotosyntezy jest skrobia gromadzona wokół pirenoidów lub w cytoplazmie. Rozmnażanie przez podział komórki[1].
Występują w różnych typach wód, jako składnik fitoplanktonu, fitobentosu, w metafitonie. Niektóre (np. Cryptomonas erosa) mogą powodować zakwity[1].
W polskiej florze stwierdzono ok. 60 gatunków[4].
W systemie przyjętym przez serwis AlgaeBase (stan wiosną 2014 r.[a]) systematyka kryptofitów przedstawiona jest następująco[5]:
Oznacza to, że w systemie tym gromada Cryptophyta jest monotypowa, jako że klasa Cryptophyceae jest jej jedynym taksonem niższego rzędu. Przyjęto w nim również, że autorem tak zdefiniowanego taksonu Cryptophyta jest Thomas Cavalier-Smith, który przedefiniował takson opisany wcześniej przez Paschera.
Kryptomonady, kryptomonadowe, kryptofity (Cryptophyta, Cryptista, Cryptomonadea, Cryptophyceae) – glony zaliczane niegdyś do gromady tobołków, jedna z grup wiciowców. Większość kryptomonad to organizmy jednokomórkowe, choć czasami można spotkać je w postaci palmelloidalnych kolonii. W różnych systemach taksonomicznych ta sama grupa organizmów może mieć różną rangę taksonomiczną, a kilka kolejnych taksonów może być monotypowe.
Cryptophyta é um grupo pequeno de organismos com 24 géneros que se encontram em águas marinhas e continentais. São organismos unicelulares, com dois flagelos quase iguais que apresentam bárbulas. No mais curto estão organizadas numa fila, mas no maior estão em duas. As células são ovais e planas com um tamanho de cerca de 10-50 μm. Podem aparecer estados palmeloides ou cocoides. Algumas espécies formam zooxantelas, quer dizer, vivem como simbióticas de animais.
Apresentam um ou dois plastos, com ou sem pirenoides. Os plastídeos apresentam os tilacoides agrupados em pares, sem lamela. Com clorofila a e c; apresentam α-caroteno e xantofilas, aloxantina dominante, no entanto falta o β-caroteno. Possuem pigmentos ficobilínicos (ficocianina ou ficoeritrina), mas não se apresentan em ficobilissomas. Utilizam como material de reserva, amido verdadeiro, extraplastidial, associado com plastos e núcleo.
Apresentam dois flagelos anteriores quase iguais que saem de um reservatório revestido por tricocistos, ambos são pleuronemáticos. Podem ter uma parede celular de celulose, apresentam um periplasto de três camadas, também podem apresentar placas poligonais externas. As mitocôndrias têm cristas planas, e a mitose é aberta. A reprodução é por bipartição, mediante zoósporos e cistos. Em alguns casos observou-se também reprodução sexual.
Cavalier-Smith, agrupa-os dentro do proposto reino Chromista como subreino Cryptista. No entanto, vários especialistas sugerem que são necessários mais estudos filogenéticos para definir a relação entre Cryptophyta e Chromista ou Chromalveolata.
A partir de evidências moleculares foi proposto que o grupo Cryptophyta é resultado de endossimbiose secundária de um ser eucarionte heterotrófico unicelular que teria fagocitado uma Rhodophyta (alga vermelha). [1] [2]
Cryptophyta é um grupo pequeno de organismos com 24 géneros que se encontram em águas marinhas e continentais. São organismos unicelulares, com dois flagelos quase iguais que apresentam bárbulas. No mais curto estão organizadas numa fila, mas no maior estão em duas. As células são ovais e planas com um tamanho de cerca de 10-50 μm. Podem aparecer estados palmeloides ou cocoides. Algumas espécies formam zooxantelas, quer dizer, vivem como simbióticas de animais.
Apresentam um ou dois plastos, com ou sem pirenoides. Os plastídeos apresentam os tilacoides agrupados em pares, sem lamela. Com clorofila a e c; apresentam α-caroteno e xantofilas, aloxantina dominante, no entanto falta o β-caroteno. Possuem pigmentos ficobilínicos (ficocianina ou ficoeritrina), mas não se apresentan em ficobilissomas. Utilizam como material de reserva, amido verdadeiro, extraplastidial, associado com plastos e núcleo.
Apresentam dois flagelos anteriores quase iguais que saem de um reservatório revestido por tricocistos, ambos são pleuronemáticos. Podem ter uma parede celular de celulose, apresentam um periplasto de três camadas, também podem apresentar placas poligonais externas. As mitocôndrias têm cristas planas, e a mitose é aberta. A reprodução é por bipartição, mediante zoósporos e cistos. Em alguns casos observou-se também reprodução sexual.
Cavalier-Smith, agrupa-os dentro do proposto reino Chromista como subreino Cryptista. No entanto, vários especialistas sugerem que são necessários mais estudos filogenéticos para definir a relação entre Cryptophyta e Chromista ou Chromalveolata.
Rekylalgerna (Cryptophyta) är en division encelliga alger som finns i både marina och limniska miljöer, inte minst i kalla områden. De flesta medlemmar är fotosyntetiserande men ändå mixotrofa. Fotosyntetiserande medlemmar har klorofyll a, c samt fykobiliner. Kloroplasten härrör ursprungligen från en upptagen rödalg (sekundär endosymbios). Kloroplasten omges av 4 membran. Totalt finns omkring 200 arter fördelade på tre ordningar.[1][2][3][4] Gruppen är även känd som kryptomonader.
Rekyalgerna indeles i tre ordningar. Cryptomonadales och Chroomonadeles innehåller många fotosyntetiserande medlemmar (se nedan). Goniomonadales innehåller endast det färglösa släktet Goniomonas.[2]
En del molekylärbiologiska data samt närvaro av rödalg som endosymbiont stödjer en placering i gruppen Chromalveolata.[5][6] Rekylalgerna verkar vara närmast släkt med fästalgerna (Haptophyta).[5] Rekylalger och fästalger räknas därför ibland tillsammans som gruppen Hacrobia.[7] Å andra sidan har rekylalgerna fler "primitiva" drag jämfört med fästalger och heterokonter såsom närvaro nukleomorf och fykobiliner (se nedan), vilket skulle kunna tyda på att de är en mycket tidigt divergerande grupp inom Chromalveolata.[6]
Plastiden (kloroplasten) härrör av allt att döma från en rödalg som tagits upp genom sekundär endosymbios. Istället för att smältas av den ursprungliga heterotrofa värdcellen behölls rödalgen inuti en särskild "vesikel", troligen gynnad av den sura miljön (sur miljö gör att koldioxid/bikarbonat framförallt föreligger som koldioxid vilket gynnar Rubiscos koldioxidfixering). Som ett arv från den sekundära endosymbiosen omges plastiden av inte mindre än fyra membran. De två membranen närmast kloroplasten härrör från denna. Membran nummer tre är endosymbiontens (rödalgens) plasmamembran medan det yttersta membranet ursprungligen är membranet från fagocytvesikeln. Just mellan membran 2 och 3 återfinns resten av rödalgens gamla cellkärna vilken reducerats till en nukleomorf, vars återstående mycket begränsade genom nu sekvenserats. De allra flesta gener har överflyttats till värdcellens genom. Det yttersta membranet är kontinuerligt med värdcellens endoplasmatiska nätverk (ER).[1][2][6]
Fotosyntetiserande rekylalger besitter klorofyll a och c2[8] samt fykobiliner (fykobiliner förekommer även hos rödalger och cyanobakterier). Hos rekylalgerna är fykobilin-proteinen utspridda över plastidernas lumen.[8] I ordningen Chroomonades utgörs fykobilinerna av en typ av fykocyanin medan hos ordningen Cryptomonadales förekommer istället fykobilinet fykoerytrin.[2] Inga rekylalger har både fykocyanin och fykoerytrin samtidigt.[2][8] Färgen hos kryptomonaderna skiftar beroende på vilka fykobiliner som finns. Fykocyanin ger algerna ett blåaktigt intryck medan fykoerytrin ger en rödaktig nyans. Även alfakaroten, diatoxantin och alloxantin förekommer. Det finns även rekylalger som saknar pigment och är således färglösa och heterotrofa. Näring lagras som stärkelse.[2][8] Hos Chliomonas har innehåller plastiden inte längre fotosyntetiska pigment, utan har blivit en leukoplast.[2]
Vissa rekylalger har ögonfläckar.[2] Enligt vissa forskare är det två rhodopsiner som fungerar som ljusreceptorer hos rekylalger, likt hos vissa grönalger.[9]
Rekylalgerna har utskjutbara projektiler kallade ejektosomer. När dessa skjuts ut rör sig kryptomonaden åt motsatt håll (därav det svenska namnet)[3]. Det är tänkbart att ejektosomerna används som försvarsmekanism, men de kan också vara en flyktmekanism.[2][4]
Rekylalgerna är viktiga fotosyntetisernade alger i framförallt kalla vatten. Exempelvis kan de leva i vegetativt tillstånd (jmf vilospor) under isen i nordliga och/eller högt belägna sjöar och på Antarktis. De har således mycket god förmåga att även ta vara på mycket sparsamt ljus.[2] Energitransfern från fykocyaninet till fotosystem I och II har också visats vara mycket effektiv.[8]
Alla fotosyntetiserande medlemmar är mixotrofa: de har alltså både fotosyntes och fagocytos. Fagocytos av bakterier tros spela en stor roll för upptag och fosfor och kväve i näringsfattiga miljöer/tider, till exempel under sjöis på vinterhalvåret.[2][1] Man har exempelvis observerat fagocytos hos Cryptomonas.[4] Rekylalgerna använder sin förmåga till rörelse genom att aktivt söka näring och/eller solljus, men samtidigt ändå undgå det starkaste ljuset mitt på dagen.[2]
Rekylalgerna är en mycket viktig näringskälla för zooplankton framförallt i kallare vatten. De är också mycket rika på omättade fettsyror.[1]
Det förekommer att rekylalger i sin tur utgör endosymbionter (tertiär endosymbios). Exempelvis lever rekylalger av släktena Chroomonas eller Hemiselmis som endosymbiont i dinoflagellaten Gymnodinium myriopyrenoides.[10] Även ciliaten Mesodinium rubrum har en rekylalg som endosymbiont.[2]
Rekylalgerna (Cryptophyta) är en division encelliga alger som finns i både marina och limniska miljöer, inte minst i kalla områden. De flesta medlemmar är fotosyntetiserande men ändå mixotrofa. Fotosyntetiserande medlemmar har klorofyll a, c samt fykobiliner. Kloroplasten härrör ursprungligen från en upptagen rödalg (sekundär endosymbios). Kloroplasten omges av 4 membran. Totalt finns omkring 200 arter fördelade på tre ordningar. Gruppen är även känd som kryptomonader.
Для фотоавтотрофних криптофітових характерні хлорофіли а та с, α- та ε-каротини, фікобілінові пігменти — фікоціанін та фікоеритрин, ксантофіли лютеїнового ряду (лютеїн, віолаксантин, зеаксантин) та специфічні ксантофіли (алоксантин, крококсантин, монадоксантин). Кількісні співвідношення пігментів у різних родів варіюють у широкому діапазоні. Це обумовлює велику різноманітність забарвлення криптофітових водоростей. Так, хлоропласти можуть бути червоними, блакитними, жовтими, коричневими, зеленими. Вторинно гетеротрофні представники безбарвні. Основний продукт асиміляції — крохмаль, який відкладається у перипластидному просторі, часто — поблизу піреноїда. Додатковий асимілят — олія.
Клітини криптофітових звичайно дорзовентральні, на передньому кінці косо зрізані, мають глибоку чи невелику глотку, два гетероконтні джгутики, що відходять від дорзального боку глоткового отвору. При основі глотки у прісноводних видів є одна-дві, рідше п'ять-вісім скоротливих вакуолей. На вентральному боці розташовується вузька поздовжня борозенка. Клітинні покриви представлені перипластом. Він складається з плазмалеми, яка з внутрішнього боку несе прямокутні чи полігональні білкові платівки, що прикріплюються до плазмалеми за допомогою білкових шипів. На зовнішньому боці плазмалеми зазвичай є шар зовнішніх білкових платівок, що за формою повторюють внутрішні. Платівки утворюють правильні ряди, які надають клітинним покривам криптофітових характерної поздовжньої посмугованості. Під перипластом знаходяться трихоцисти. Великі трихоцисти звичайно вистилають глотку, дрібні розсіяні під усією поверхнею клітини. Ядро розташовується в задній частині клітини, має досить велике ядерце. Зовнішня ядерна мембрана безпосередньо переходить у зовнішню мембрану хлоропласта. Центріолі відсутні, їх функцію виконують базальні тіла джгутиків та ризопласти. Хромосоми в інтерфазі знаходяться у конденсованому стані. Мітоз відкритий. Зазвичай у клітині є один-два великі пристінні хлоропласти, вкриті чотиримембранною оболонкою. Дві зовнішні мембрани утворюють хлоропластну ендоплазматичну сітку. Зовнішня мембрана хлоропластного ендоплазматичного ретикулюму безпосередньо переходить у зовнішню мембрану ядра. Перипластидний простір між двома зовнішніми та двома внутрішніми мембранами хлоропласта містить крохмаль та особливу органелу — нуклеоморф. Від перипластидного простору нуклеоморф відмежований подвійною мембраною, яка, подібно до ядерної, має пори та містить ДНК та РНК. Тилакоїди у хлоропласті зібрані парами. На внутрішніх боках кожної пари локалізовано фікобілінові пігменти, які, проте, не організовані у фікобілісоми. Більшість криптофітових мають великий піреноїд напівзануреного типу. У зоні перипластидного простору, що безпосередньо прилягає до піреноїда, відкладається крохмаль. У деяких криптофітових у хлоропласті є вічко, положення якого не зкоординовано із джгутиковим апаратом. Вторинні гетеротрофи безбарвні, але зберігають редуковану пластиду — лейкопласт. Мітохондріальний апарат представлений двома великими мітохондріями — дорзальною та вентральною. Обидві мають пластинчасті кристи. Монадні клітини дводжгутикові, джгутики злегка гетероконтні, спрямовані вперед. На довшому джгутику мастигонеми розташовуються у два ряди, на коротшому вони однорядні. У перехідній зоні джгутики мають по дві поперечні платівки. Специфічними органелами криптофітових є тільця Мопа — великі, добре помітні одномембранні структури, що знаходяться поблизу глотки і містять переважно залишки трихоцист. Припускають, що тільця Мопа є гомологами лізосом і їх функція полягає в перетравленні надлишкових трихоцист. Звичайно між глоткою та тільцями Мопа розташовується один комплекс Гольджі.
Майже всі криптофітові водорості мають монадний тип структури тіла. Переважна більшість видів здатна втрачати джгутики, одягатися шаром слизу і переходити у пальмелевидний стан.
Розмноження відбувається поділом клітин надвоє у поздовжньому напрямку в рухливому стані. У одного виду (Proteomonas sulcata) описаний гаплодиплофазний життєвий цикл з гетероморфною зміною поколінь. У Chroomonas acuta спостерігався хологамний статевий процес.
Всі криптофітові належать до одного класу — Cryptophyceae та одного порядку — Cryptomonadales. Характерними представниками є роди Cryptomonas, Rhodomonas та Chilomonas.
Большинство криптофитовых — одноклеточные подвижные организмы монадной формы с дорсовентральным строением и жгутиками. Своё название группа получила за очень маленькие размеры (от 3 до 50 мкм), которые делают их малозаметными (др.-греч. κρυπτός — скрытый и др.-греч. μόνος — одинокий)[8]. У некоторых представителей в жизненном цикле имеется пальмеллоидная стадия (например, у Cryptomonas[en] и Chroomonas[en][9]). Лишь один вид — Bjornbergiella hawaiiensis — формирует простые нитчатые талломы. Среди криптофит известны неподвижные в вегетативном состоянии колониальные и одноклеточные коккоидные организмы[10] (однако существование таких форм оспаривается)[11].
Клетка типичной криптофитовой водоросли уплощённая, с выпуклой спинной и вогнутой брюшной сторонами. Форма клеток разнообразна: яйцевидная, эллипсоидная, грушевидная, бобовидная, веретеновидная. Передний конец клетки скошен, от него отходит продольная борозда, не доходящая до заднего конца. Наличие борозды и её длина используются в качестве признаков для определения родов[12].
У некоторых представителей на переднем конце тела имеется мешковидная глотка, или вестибулум[13], или крипта[14]. У разных форм глотка имеет различную длину, она может проходить вдоль или поперёк тела. Иногда она разветвляется; у некоторых видов криптофит глотка укреплена цитоскелетными образованиями — электронно-плотными пластинами. Функциональное значение глотки до сих пор неясно; во всяком случае, у большинства фототрофных форм никогда не наблюдался захват ею каких-либо частиц[11][15].
Покровы клеток криптофит представлены перипластом, который состоит из плазмалеммы и двух дополнительных слоёв белкового материала, расположенных над и под плазмалеммой. Внутриклеточная часть перипласта может иметь форму сплошного чехла, как у Chilomonas[en], или состоять из отдельных пластинок: шестиугольных, как у Cryptomonas[en], или прямоугольных, как у Chroomonas[en]. Прикрепление белковых пластинок к плазмалемме осуществляется за счёт округлых или многоугольных внутримембранных частиц. Наружная часть перипласта может состоять из пластинок, чешуек, слизи или их комбинации. В области глотки перипласта нет[16], однако закладка новых пластинок перипласта начинается в области рядом с глоткой. Перипласт обеспечивает постоянство формы клетки[13] и предопределяет вид её поверхности: ровный или структурированный (так, у Chroomonas поверхность клетки состоит из прямоугольников, у Rhinomonas[en] — из шестигранников)[15]. Клетка может секретировать наружу сульфатированные полисахариды, богатые фукозой[17].
На переднем конце клетки у монадных форм криптофит имеются два неравных жгутика[10], по длине сопоставимые с самой клеткой. Оба жгутика отходят от возвышения на краю глотки[18]. Различия в длине между жгутиками невелики: отношения их длин колеблются от 3:5 до 9:10. Оба жгутика заканчиваются более узкой частью — акронемой, в которую заходят лишь две центральные трубочки аксонемы[19].
Когда клетка движется, то оба жгутика направлены вперёд или же один направлен вперёд, а второй — назад. На жгутиках имеются особые мастигонемы[en] уникального строения, которые могут располагаться в два противолежащих ряда на длинном жгутике (длина мастигонем до 2 мкм), в один ряд на коротком жгутике или же на обоих жгутиках. Мастигонемы криптофит двухчастные и состоят из трубчатой части и тонкого терминального волоска. У некоторых представителей короткий жгутик не несёт мастигонем. У основания длинного жгутика может находиться дополнительный пучок, включающий около 45 волосков. Жгутики могут нести небольшие органические чешуйки в виде семиугольных розеток диаметром 140—170 нм[20]. Известны формы с единственным перистым жгутиком[10].
Ультраструктура жгутика также имеет свои особенности. В переходной зоне жгутиков имеются две или более пластинчатые структуры (септы) ниже точки, где заканчиваются две центральные микротрубочки. Строение корешкового аппарата варьирует у представителей различных родов, но в общем случае корешковая система представлена ризостилем, который идёт в глубь клетки и состоит из 6—10 микротрубочек, соединённых сократительными фибриллами, трёх микротрубочковых корешков и фибриллярного корешка, прилегающего к одному из микротрубочковых[20].
Асимметричное расположение жгутиков вызывает несбалансированное вращение клетки вдоль продольной оси при плавании. Чем крупнее клетка и чем более изогнутую форму она имеет, тем более заметным становится такое «раскачивание»[21].
В клетке криптофитовой водоросли имеется один или два хлоропласта (реже больше), окрашенных в разнообразные цвета: от сине-зелёного и оливково-зелёного до жёлто-бурого, коричневого и тёмно-красного[20]. Хлоропласты могут быть глубоко разделены на две доли или иметь Н-образную форму[21]. Имеются бесцветные формы с деградировавшими пластидами, они ведут гетеротрофный образ жизни[13].
Хлоропласты криптофит содержат следующие пигменты: хлорофилл а, хлорофилл c2 и фикобилины: фикоцианин и фикоэритрин (вместе эти пигменты никогда не встречаются, хлоропласт может содержать лишь один из них). За передачу энергии света от фикобилинов к хлорофиллу а отвечает хлорофилл c2, а аллофикоцианин, выполняющий эту функцию в клетках красных и синезелёных водорослей, отсутствует. Фикобилины криптофит расположены не в фикобилисомах, а во внутритилакоидных пространствах хлоропластов, поэтому тилакоиды криптофит толще, чем у других водорослей[22]. Выявлено несколько типов фикоцианина и фикоэритрина, отличающихся различным положением максимумов поглощения света; так, в клетках Cryptomonas[en] содержится фикоэритрин 566, Hemiselmis[en] — фикоэритрин 555, Pyrenomonas[en] — фикоэритрин 545; у Komma[en] клетки содержат фикоцианин 645, у Chroomonas[en] — фикоцианин 630 (цифры задают длину в нанометрах волны, на которой поглощение максимально)[23].
У криптофит имеется несколько уникальных ксантофиллов, в частности, аллоксантин, который можно использовать для выявления их наличия в смеси планктонных водорослей, однако главным ксантофиллом является диатоксантин[22]. Имеются также α-[en] и β-каротин, крококсантин, зеаксантин и монадоксантин[24]. Соотношение пигментов видоспецифично[13]. Кроме того, оно зависит от интенсивности освещения: чем меньше интенсивность освещения, тем больше фикоэритрина содержится в хлоропластах и тем толще тилакоиды[25].
Криптофиты характеризуются уникальным типом расположения тилакоидов в хлоропласте. Они собраны в пары или в тройки, причём соседние тилакоиды не имеют соединений[22]. Опоясывающей ламеллы нет. ДНК хлоропласта представлена небольшими нуклеоидами, которые рассеяны в строме хлоропласта[1].
В хлоропласте может присутствовать пиреноид, в который могут заходить ламеллы из 1—2 тилакоидов[24]. У видов с двудольными хлоропластами пиреноид располагается в месте соединения двух долей[21]. Таким образом, в отличие от хлоропластов красных и зелёных водорослей, хлоропласты криптофит не лежат свободно в цитоплазме, а связаны с другими органеллами (пиреноидом, вакуолями)[26].
Хлоропласт криптофитовой водоросли окружён 4 мембранами[en], наружная несёт на своей поверхности рибосомы и продолжается в ядерную мембрану и эндоплазматический ретикулум (ЭПР)[21]. Происхождение хлоропластов криптофит рассматривают как пример вторичного эндосимбиоза, когда бесцветный фаготрофный жгутиконосец, сходный с современной криптофитовой водорослью Goniomonas[en][17], «съел» красную водоросль. Поэтому две внутренние мембраны хлоропласта криптофит интерпретируют как мембраны хлоропласта красной водоросли, третью мембрану — как плазмалемму клетки красной водоросли, а четвёртую мембрану — как мембрану пищеварительной вакуоли клетки-хозяина. Между двумя парами мембран расположено перипластидное пространство, в котором располагаются 80S рибосомы, содержащие, однако, рРНК эукариотического типа, зёрна крахмала, а также нуклеоморф — сильно редуцированное ядро красной водоросли. Происхождение хлоропластов криптофит от красных водорослей подтверждается анализом 16S рРНК нуклеоморфа и ядер красных водорослей[24][27]. Две наружные мембраны иногда называют пластидным эндоплазматическим ретикулумом[26].
Процесс доставки в хлоропласты белков, кодируемых ядерной ДНК, у криптофит организован гораздо более сложно, чем у растений, чьи пластиды представляют собой результат первичного, а не вторичного эндосимбиоза. У криптофит и других организмов, чьи хлоропласты появились в результате вторичного эндосимбиоза, белки, кодируемые ядром, но предназначенные для пластид, содержат на N-конце лидерную последовательность, состоящую из сигнального пептида и переходного пептида. Два этих сигнала совместно обеспечивают прохождение белком всех четырёх мембран для доставки к месту своего функционирования. Было высказано предположение, что в переносе белков в перипластидное пространство принимает участие кодируемая нуклеоморфом ЭПР-связанная система деградации. Поскольку эндосимбионт утратил свой ЭПР, то эта система могла приобрести новую функцию в транспорте белков[21].
Как уже отмечалось выше, нуклеоморф представляет собой рудиментарное ядро эукариотического фототрофного эндосимбионта, инкорпорированного предками криптофитов в ходе вторичного эндосимбиоза (кроме криптофит, нуклеоморф также имеется у водорослей группы Chlorarachniophyta; однако в последнем случае происходила инкорпорация не красной, а зелёной водоросли[28]). У криптофит нуклеоморф обычно имеет округлую или грушевидную форму; он окружён двойной мембраной, содержащей поры, сходные с ядерными поровыми комплексами, и имеет ядрышко с типичными эукариотическими РНК. Нуклеоморф может быть ассоциирован с пиреноидом; иногда он располагается внутри самого пиреноида, иногда — в выросте хлоропласта[27][29].
Размер ДНК нуклеоморфа Rhodomonas salina[en] составляет только 660 тыс. пар нуклеотидов в трёх хромосомах (240, 225 и 195 тыс. пар нуклеотидов). Таким образом, геном нуклеоморфа — мельчайший из описанных ядерных геномов эукариот[30]. Интересно, что нуклеоморфы Chlorarachniophyta также имеют 3 хромосомы. У различных видов криптофит ДНК нуклеоморфа содержит от 450 до 850 тыс. пар нуклеотидов. Нуклеоморф имеет 531 ген, которые кодируют 30 белков, функционирующих в хлоропластах. Плотность генов в ДНК нуклеоморфов криптофит довольно высока. Гены нуклеоморфа в большинстве своём являются генами домашнего хозяйства, их продукты участвуют в таких процессах, как фолдинг и разрушение белков, транскрипция и трансляция. В 2001 году был секвенирован геном нуклеоморфа Guillardia theta, а в 2007 году — Hemiselmis andersenii[17][21][24].
Деление нуклеоморфа происходит в препрофазе деления главного ядра вслед за репликацией базального тела, но до деления хлоропласта. Деление амитотическое, и, хотя геном нуклеоморфа содержит гены α-, β- и γ-тубулина, микротрубочки не принимают участия в образовании веретена деления; вместо этого имеются фибриллярные структуры, появляющиеся только во время деления. Дочерние нуклеоморфы мигрируют в противоположные концы хлоропласта, поэтому каждый дочерний хлоропласт наследует один нуклеоморф. Единственная известная криптофитовая водоросль, не имеющая нуклеоморфа — Goniomonas, которая утратила пластиды. У другой бесцветной криптофитовой водоросли, Chilomonas[en], имеются лейкопласт и нуклеоморф[24][31].
У ряда видов криптофит имеется глазок (стигма). У криптофит он связан с хлоропластом, но не связан со жгутиковым аппаратом, а потому может располагаться и в центре клетки, как, например, у Chroomonas mesostigmatica. Стигма состоит из большого количества пигментных липидных глобул, расположенных параллельными рядами под оболочкой хлоропласта. Для некоторых криптофит показан положительный фототаксис. В частности, бесцветный Chilomonas наиболее чувствителен к синему свету при длине волны 366 нм[20][32].
Ядра криптофит, как правило, невелики и не превышают в диаметре 3 мкм. Клетка имеет единственное ядро, расположенное в задней части клетки. Для ядер криптофит характерен сложный хромоцентрический тип организации. Хроматиновый ободок (скопление хроматина по краю ядра) может быть сплошным, почти сплошным или же узким и едва различимым, сильно разорванным. Остальное пространство ядра заполнено многочисленными глыбками хроматина, только у Rhinomonas panca они единичны[33]. У различных криптофит имеется от 40 до 210 хромосом[34]. В 2012 году было завершено секвенирование ядерного генома криптофитовой водоросли Guillardia theta[en], её ядерный геном оказался гаплоидным[35].
Митоз у исследованных криптофит отличается поразительным сходством и относится к открытому центрическому типу. Процесс деления клеток начинается с удвоения базальных тел. Ядра перемещаются в переднюю часть клетки и иногда вплотную подходят к базальным телам. Базальные тела не меняют своего положения в течение всего митоза и во время метафазы располагаются по обе стороны метафазной пластинки. Они не становятся центриолями, но выполняют их функции, в частности, около них формируются многочисленные микротрубочки, участвующие в образовании веретена[33].
В профазе митоза ядерная оболочка фрагментируется и постепенно растворяется, лишь малая часть её фрагментов сохраняется до конца митоза. В случае открытого митоза у криптофит происходит формирование дополнительной перинуклеарной оболочки из хлоропластного ретикулума[36][37]. При митозе ядрышко не исчезает[34].
Веретено деления криптофит также необычно. Оно состоит из параллельных неконвергирующих микротрубочек. Область полюсов веретена уплощена и ограничена цистернами ЭПР[34]. На его концах, кроме ризостилей, которые отходят от каждой пары базальных тел и не контактируют с микротрубочками веретена, располагаются нуклеоморфы, специально переместившиеся сюда. В метафазе вместо типичной экваториальной пластинки из морфологически дифференцированных хромосом образуется очень компактное скопление хроматина, в котором отдельные хромосомы не различимы, однако образуются особые проходы для микротрубочек веретена. Одно время считали, что истинных хромосом у криптофит нет, а компактную массу хроматина можно рассматривать как одну сборную хромосому. Однако сейчас считают, что хромосомы криптофит очень мелкие, а их кинетохоры морфологически очень слабо дифференцированы. Роль кинетохоров, по-видимому, выполняют выступы хроматина в области коридоров, которые контактируют с микротрубочками веретена[38]. При делении клеток также происходит формирование новой глотки, которая достаётся одной из дочерних клеток[14].
Клетки криптофит нередко несут особые стрекательные структуры — эжектосомы, или трихоцисты. Их строение отличается от стрекающих структур динофлагеллят, однако они, возможно, родственны R-тельцам каппа-частиц инфузорий. Крупные эжектосомы располагаются под плазмалеммой в области глотки, а более мелкие разбросаны по всей поверхности клетки. Каждая эжектосома окружена одной мембраной и содержит внутри 2 ленты, скрученные в цилиндры. Более крупный цилиндр состоит из намотанного на спирали лентовидного мембранного материала. В его верхней части находится V-образное углубление, в котором залегает второй цилиндр. По-видимому, эти два цилиндра связаны друг с другом при помощи волокон. При химическом и физическом раздражении скрученные ленты распрямляются и выстреливают, разрывая мембрану эжектосомы и перипласт. Сначала выбрасывается маленький цилиндр, он вытягивает за собой ленту большого цилиндра. Выстрелившая эжектосома с периферии клетки имеет длину 4 мкм, а крупная эжектосома, расположенная около глотки, после срабатывания имеет длину 20 мкм. Выстреливание эжектосом заставляет клетку двигаться в противоположном направлении. Кроме того, при выстреливании из эжектосом высвобождаются вещества, убивающие бактерий, которыми питаются криптофиты[14]. Эжектосомы формируются в пузырьках аппарата Гольджи, причём количество оборотов в цилиндре увеличивается по мере созревания эжектосомы[12][39].
С наличием эжектосом связаны трудности по переводу криптофит на лабораторные среды. Их клетки чрезвычайно чувствительны к изменению окружающих условий (рН, осмотические условия, температура) и так сильно отстреливают содержимое эжектосом, что клетки быстро разрушаются[12].
В цитоплазме клеток криптофит имеется одна сетчатая митохондрия с пластинчатыми кристами[40]. Она образует две сети: периферическую (лежащую под плазмалеммой) и центральную (в центре клетки)[27].
На переднем конце тела у пресноводных криптофит располагаются сократительные вакуоли, изливающие своё содержимое в глотку или борозду[12]. Они служат для регуляции осмотического давления внутри клетки[21].
В клетках криптофит имеются два тельца Мопа. Тельце Мопа представляет собой крупную везикулярную структуру, расположенную в передней части клетки. Точные функции телец Мопа неизвестны, но, так как они содержат много мембран и фибрилл, они могут принимать участие в разрушении клеточных органелл, особенно отработанных эжектосом, и других инородных частиц в цитоплазме[12][39].
Среди криптофит имеются автотрофы, гетеротрофы (сапротрофы и фаготрофы) и миксотрофы. Большинство криптофит нуждаются в витамине B12 и тиамине, а некоторые — в биотине. В качестве источников азота криптофиты могут использовать аммоний и органические азотсодержащие соединения, однако морские формы менее способны к превращениям нитрата в нитрит по сравнению с пресноводными представителями. Органические вещества стимулируют рост криптофит. Различные бесцветные формы, а также окрашенный Cryptomonas ovata способны к фагоцитозу, а сине-зелёный Chroomonas pochmannii является миксотрофом. Для захвата и удержания бактериальных клеток используются особые вакуоли. Бактерии втягиваются в них через маленькую пору в глотке, где нет пластинок перипласта, и в вакуолях же перевариваются[34]. Некоторые криптофиты (например, Goniomonas truncata) могут поедать вирусные частицы[41]. При фаготрофном питании криптофиты демонстрируют хемотаксис в направлении наибольшей концентрации питательных веществ[42].
Основное запасное вещество у криптофит — крахмал, он откладывается между второй и третьей мембранами хлоропласта; из всех водорослей только криптофиты откладывают крахмал в этом месте. Как и у красных водорослей, он содержит больше амилопектина, чем амилозы, поэтому с йодом даёт красное окрашивание. В цитоплазме могут иметься липидные капли[43].
Криптофиты размножаются, главным образом, вегетативно — путём деления клетки надвое с помощью борозды деления, причём впячивание плазмалеммы начинается с заднего конца клетки. В большинстве случаев делящаяся клетка сохраняет подвижность. Наибольшая скорость роста для многих криптофит — одно деление в день при температуре около 20 °С[34].
Половой процесс описан у немногих видов, может протекать в форме изогамии. У Chroomonas acuta[en] описана хологамия, то есть вегетативные клетки функционируют как гаметы. Слияние клеток происходит на брюшной стороне, причём задний конец одной клетки прикрепляется к середине другой. После слияния клеток формируется четырёхжгутиковая зигота, которая делится, вероятно, мейозом. У Proteomonas sulcata описан гетероморфный гаплодиплонтный жизненный цикл, в котором имеются как гаплоидная, так и диплоидная стадии. Обе стадии различаются по размеру, структуре перипласта и строению жгутикового аппарата (остальные криптофиты — гаплоидные организмы[37]). Однако у этого вида до сих пор не открыты оплодотворение и мейоз[44].
При неблагоприятных условиях (дефицит азота, избыточное освещение) внутри клеток формируются толстостенные цисты[34][45].
Криптофиты — планктонные организмы, иногда они встречаются в иле солёных озёр и детрите в пресных водоёмах. Большая часть из них может жить в широком температурном диапазоне, однако некоторые виды Rhodomonas могут жить только при строго определённых температурах. Виды рода Cryptomonas криофильны, они способны жить и размножаться на поверхности льда и снега[46]. Chroomonas africana не утрачивает жизнеспособности при перенесении из морской воды в пресную. Криптофиты устойчивы к загрязнению, они встречаются даже в сточных водах и навозной жиже[37].
По сравнению с другими водорослями, криптофиты чрезвычайно чувствительны к избыточному освещению, поэтому они часто формируют глубоководные популяции в чистых олиготрофных озёрах. В высокогорных водоёмах и северных озёрах криптофиты встречаются в воде в течение всей зимы. Поскольку лёд и снег пропускают очень мало света, они собираются в поверхностных слоях воды, чтобы получить достаточное для фотосинтеза количество света. Выживание в условиях такой низкой освещённости обеспечивается не только высокой эффективностью фотосинтетической системы, но также низкой скоростью клеточного дыхания при низких температурах и снижением поедания криптофит зоопланктоном в условиях зимы. Весной, когда снег тает и освещённость арктических и горных водоёмов повышается, криптофиты страдают от повышенной освещённости, и большая часть их биомассы уходит в более глубокие слои воды[42].
Криптофиты нередко совершают суточные миграции в вертикальном направлении с амплитудой менее 5 м. Cryptomonas, обитающие в небольших лесных озёрах, утром демонстрируют положительный фототаксис и перемещаются в верхний слой воды, бедный фосфором. Далее криптофиты уходят из самых верхних слоёв, избегая избыточного освещения, и перемещаются в богатый фосфором нижний слой холодной воды (гиполимнион). Этот цикл миграций также помогает криптофитам избежать поедания зоопланктоном[42].
Криптофиты — доминирующая группа водорослей в покрытых льдами пресноводных водоёмах Антарктики, где они могут составлять до 70 % биомассы фитопланктона. В таких водоёмах наблюдается значительное расслоение воды, поскольку факторы, способные вызывать движение воды (ветер, изменение температуры воды), отсутствуют. Криптофиты доминируют в самых нижних слоях, где в течение зимы они ведут гетеротрофный образ жизни и поглощают одну бактерию в час путём фагоцитоза. Летом криптофиты миксотрофны. Залогом выживания криптофитов в таких условиях является то, что они всё время находятся в вегетативном состоянии, а не уходят в покоящиеся формы. Находясь в вегетативной форме, популяция может быстро отреагировать на улучшение условий и приход короткого антарктического лета[47]. Летом, когда начинается приток воды от таяния ледников, криптофиты вызывают локализованные цветения в антарктических водах[46]. Цветение воды, вызванное криптофитовыми водорослями, было зафиксировано и в Белом море[48].
Очень редко криптофиты живут в океанических водах при температуре 22 °С и выше, они отсутствуют в горячих источниках и гиперсолёных водоёмах. Пресноводные криптофиты населяют искусственные и естественные водоёмы со стоячей водой, где также избегают сильно освещённых слоёв воды. Бесцветные формы распространены в водах, загрязнённых органикой, и могут служить показателями загрязнения воды органическими веществами[49].
Среди криптофит имеются виды, населяющие болота с низкими значениями рН, однако ряд видов не обладает избирательностью по отношению к рН: так, Chilomonas живёт при рН 4,1—8,4[50].
Некоторые криптомонады обитают в пищеварительном тракте домашних животных[14].
Криптофиты живут в качестве эндосимбионтов в клетках инфузории Myrionecta rubra. Эта инфузория может вызывать нетоксичное красное цветение воды в зоне апвеллинга. Цвет обусловлен красновато-коричневыми хлоропластами эндосимбионта. Криптофитовая водоросль, находящаяся внутри инфузории, окружена двойной мембраной (плазмалемма водоросли и пищеварительная вакуоль клетки-хозяина) и функционирует как полноценный автотроф. Ей присущи все особенности строения криптофитовых водорослей, но она лишена перипласта, жгутикового аппарата и эжектосом. Инфузория, содержащая эндосимбионтов-криптофит, целиком полагается на них в своём питании и утрачивает клеточный рот[50][51].
Некоторые криптофиты являются эндосимбионтами некоторых динофлагеллят (например, Dinophysis[en]), причём в некоторых случаях у них может сохраняться ядро криптофитовой водоросли, а в других случаях сохраняются только пластиды[50].
Криптофиты имеют большое значение в круговороте кислорода, углерода, азота и фосфора в водоёмах, в синтезе органики из минеральных веществ. Криптофиты служат пищей для различных представителей зоопланктона, которым, в свою очередь, питаются рыбы. Ветвистоусые рачки интенсивно питаются криптофитами в эвтрофных прудах в зимнее время. Однако цветение воды в прудах, обусловленное Cyanomonas, приводило к массовой гибели сомов[50].
В настоящее время криптофиты находят применение в исследовательской сфере. Так, недавно открытые родопсины водоросли Guillardia theta, представляющие собой анионные каналы, могут использоваться в качестве очень эффективного оптогенетического метода для подавления нейронов[52].
На протяжении почти всего XX века классификация криптофитов разрабатывалась протозоологами и ботаниками-альгологами практически независимо. Протозоологи трактовали таксон как отряд Cryptomonadida Calkins, 1926 в составе типа Protozoa[53]; альгологи — либо как класс Cryptophyceae Pascher, 1914 в составе отдела Pyrrophyta (включавшего также класс Dinophyceae, а иногда — ещё и класс Chloromonadophyceae[en])[54], либо как самостоятельный отдел Cryptophyta Silva, 1962[19]. При этом внутреннюю классификацию криптофитовых водорослей ботаники строили на основе типа организации таллома, выделяя порядки Cryptomonadales (монадные формы) и Cryptococcales (коккоидные формы)[19]. В конце XX века эти ранние классификации полностью устарели.
Исследования в области филогении криптофитов и родственных им групп эукариот, развернувшиеся в конце XX — начале XXI века, нашли отражение в системе типа Cryptysta (тип Cryptophyta в широком понимании), которая была предложена в 2015 году Майклом Руджеро с соавторами и входит как составная часть в представленную ими макросистему живущих организмов. Здесь данный тип рассматривается в составе подцарства Hacrobia[en] (включающего также гаптофитов, центрохелидных солнечников и несколько мелких групп с неясными родственными связями: отряды Heliomonadida[en], Microhelida, Picomonadida и Telonemida[en]). Система доведена до уровня порядков и отрядов (по отношению к классу Cryptophyceae авторы системы следуют правилам ботанической номенклатуры, по отношению к другим классам криптистов — правилам зоологической номенклатуры) и выглядит следующим образом[55][56]:
Тип Cryptista Cavalier-Smith, 1989 — криптисты[57]
Внутри криптофит эволюционно наиболее древними считаются формы, близкие к роду Goniomonas. Представители этого рода лишены пластид и нуклеоморфа, и наличие у них продольной борозды в отсутствие глотки считается наиболее примитивной особенностью. Формирование глотки происходило за счёт смыкания краёв борозды, начиная с заднего конца; этот процесс завершился у Chroomonas, имеющего глотку, но лишённого борозды. В случае Chilomonas произошёл вторичный переход к гетеротрофности: у этого организма редуцирована борозда, развита глотка и имеются лейкопласты. Такая схема эволюционных взаимоотношений подтверждается анализом 18S рРНК[37][50].
Статус таксономической группы Hacrobia (а следовательно, и внешние связи криптофитов) до сих пор окончательно неясен. В одних исследованиях группа предстаёт как клада, сестринская по отношению к группе SAR (страменопилы, альвеоляты и ризарии)[58]. Но есть и работы, в которых единство группы Hacrobia отвергается; в одной из них центрохелидные солнечники, гаптофиты и телонемиды предстают как последовательные ответвления у основания группы SAR, а криптисты не связаны с ними и оказываются наиболее ранним ответвлением группы Archaeplastida[59].
Большинство криптофитовых — одноклеточные подвижные организмы монадной формы с дорсовентральным строением и жгутиками. Своё название группа получила за очень маленькие размеры (от 3 до 50 мкм), которые делают их малозаметными (др.-греч. κρυπτός — скрытый и др.-греч. μόνος — одинокий). У некоторых представителей в жизненном цикле имеется пальмеллоидная стадия (например, у Cryptomonas[en] и Chroomonas[en]). Лишь один вид — Bjornbergiella hawaiiensis — формирует простые нитчатые талломы. Среди криптофит известны неподвижные в вегетативном состоянии колониальные и одноклеточные коккоидные организмы (однако существование таких форм оспаривается).
Клетка типичной криптофитовой водоросли уплощённая, с выпуклой спинной и вогнутой брюшной сторонами. Форма клеток разнообразна: яйцевидная, эллипсоидная, грушевидная, бобовидная, веретеновидная. Передний конец клетки скошен, от него отходит продольная борозда, не доходящая до заднего конца. Наличие борозды и её длина используются в качестве признаков для определения родов.
У некоторых представителей на переднем конце тела имеется мешковидная глотка, или вестибулум, или крипта. У разных форм глотка имеет различную длину, она может проходить вдоль или поперёк тела. Иногда она разветвляется; у некоторых видов криптофит глотка укреплена цитоскелетными образованиями — электронно-плотными пластинами. Функциональное значение глотки до сих пор неясно; во всяком случае, у большинства фототрофных форм никогда не наблюдался захват ею каких-либо частиц.
Клеточные покровыПокровы клеток криптофит представлены перипластом, который состоит из плазмалеммы и двух дополнительных слоёв белкового материала, расположенных над и под плазмалеммой. Внутриклеточная часть перипласта может иметь форму сплошного чехла, как у Chilomonas[en], или состоять из отдельных пластинок: шестиугольных, как у Cryptomonas[en], или прямоугольных, как у Chroomonas[en]. Прикрепление белковых пластинок к плазмалемме осуществляется за счёт округлых или многоугольных внутримембранных частиц. Наружная часть перипласта может состоять из пластинок, чешуек, слизи или их комбинации. В области глотки перипласта нет, однако закладка новых пластинок перипласта начинается в области рядом с глоткой. Перипласт обеспечивает постоянство формы клетки и предопределяет вид её поверхности: ровный или структурированный (так, у Chroomonas поверхность клетки состоит из прямоугольников, у Rhinomonas[en] — из шестигранников). Клетка может секретировать наружу сульфатированные полисахариды, богатые фукозой.
ЖгутикиНа переднем конце клетки у монадных форм криптофит имеются два неравных жгутика, по длине сопоставимые с самой клеткой. Оба жгутика отходят от возвышения на краю глотки. Различия в длине между жгутиками невелики: отношения их длин колеблются от 3:5 до 9:10. Оба жгутика заканчиваются более узкой частью — акронемой, в которую заходят лишь две центральные трубочки аксонемы.
Когда клетка движется, то оба жгутика направлены вперёд или же один направлен вперёд, а второй — назад. На жгутиках имеются особые мастигонемы[en] уникального строения, которые могут располагаться в два противолежащих ряда на длинном жгутике (длина мастигонем до 2 мкм), в один ряд на коротком жгутике или же на обоих жгутиках. Мастигонемы криптофит двухчастные и состоят из трубчатой части и тонкого терминального волоска. У некоторых представителей короткий жгутик не несёт мастигонем. У основания длинного жгутика может находиться дополнительный пучок, включающий около 45 волосков. Жгутики могут нести небольшие органические чешуйки в виде семиугольных розеток диаметром 140—170 нм. Известны формы с единственным перистым жгутиком.
Ультраструктура жгутика также имеет свои особенности. В переходной зоне жгутиков имеются две или более пластинчатые структуры (септы) ниже точки, где заканчиваются две центральные микротрубочки. Строение корешкового аппарата варьирует у представителей различных родов, но в общем случае корешковая система представлена ризостилем, который идёт в глубь клетки и состоит из 6—10 микротрубочек, соединённых сократительными фибриллами, трёх микротрубочковых корешков и фибриллярного корешка, прилегающего к одному из микротрубочковых.
Асимметричное расположение жгутиков вызывает несбалансированное вращение клетки вдоль продольной оси при плавании. Чем крупнее клетка и чем более изогнутую форму она имеет, тем более заметным становится такое «раскачивание».
ХлоропластыВ клетке криптофитовой водоросли имеется один или два хлоропласта (реже больше), окрашенных в разнообразные цвета: от сине-зелёного и оливково-зелёного до жёлто-бурого, коричневого и тёмно-красного. Хлоропласты могут быть глубоко разделены на две доли или иметь Н-образную форму. Имеются бесцветные формы с деградировавшими пластидами, они ведут гетеротрофный образ жизни.
Хлоропласты криптофит содержат следующие пигменты: хлорофилл а, хлорофилл c2 и фикобилины: фикоцианин и фикоэритрин (вместе эти пигменты никогда не встречаются, хлоропласт может содержать лишь один из них). За передачу энергии света от фикобилинов к хлорофиллу а отвечает хлорофилл c2, а аллофикоцианин, выполняющий эту функцию в клетках красных и синезелёных водорослей, отсутствует. Фикобилины криптофит расположены не в фикобилисомах, а во внутритилакоидных пространствах хлоропластов, поэтому тилакоиды криптофит толще, чем у других водорослей. Выявлено несколько типов фикоцианина и фикоэритрина, отличающихся различным положением максимумов поглощения света; так, в клетках Cryptomonas[en] содержится фикоэритрин 566, Hemiselmis[en] — фикоэритрин 555, Pyrenomonas[en] — фикоэритрин 545; у Komma[en] клетки содержат фикоцианин 645, у Chroomonas[en] — фикоцианин 630 (цифры задают длину в нанометрах волны, на которой поглощение максимально).
У криптофит имеется несколько уникальных ксантофиллов, в частности, аллоксантин, который можно использовать для выявления их наличия в смеси планктонных водорослей, однако главным ксантофиллом является диатоксантин. Имеются также α-[en] и β-каротин, крококсантин, зеаксантин и монадоксантин. Соотношение пигментов видоспецифично. Кроме того, оно зависит от интенсивности освещения: чем меньше интенсивность освещения, тем больше фикоэритрина содержится в хлоропластах и тем толще тилакоиды.
Криптофиты характеризуются уникальным типом расположения тилакоидов в хлоропласте. Они собраны в пары или в тройки, причём соседние тилакоиды не имеют соединений. Опоясывающей ламеллы нет. ДНК хлоропласта представлена небольшими нуклеоидами, которые рассеяны в строме хлоропласта.
В хлоропласте может присутствовать пиреноид, в который могут заходить ламеллы из 1—2 тилакоидов. У видов с двудольными хлоропластами пиреноид располагается в месте соединения двух долей. Таким образом, в отличие от хлоропластов красных и зелёных водорослей, хлоропласты криптофит не лежат свободно в цитоплазме, а связаны с другими органеллами (пиреноидом, вакуолями).
Хлоропласт криптофитовой водоросли окружён 4 мембранами[en], наружная несёт на своей поверхности рибосомы и продолжается в ядерную мембрану и эндоплазматический ретикулум (ЭПР). Происхождение хлоропластов криптофит рассматривают как пример вторичного эндосимбиоза, когда бесцветный фаготрофный жгутиконосец, сходный с современной криптофитовой водорослью Goniomonas[en], «съел» красную водоросль. Поэтому две внутренние мембраны хлоропласта криптофит интерпретируют как мембраны хлоропласта красной водоросли, третью мембрану — как плазмалемму клетки красной водоросли, а четвёртую мембрану — как мембрану пищеварительной вакуоли клетки-хозяина. Между двумя парами мембран расположено перипластидное пространство, в котором располагаются 80S рибосомы, содержащие, однако, рРНК эукариотического типа, зёрна крахмала, а также нуклеоморф — сильно редуцированное ядро красной водоросли. Происхождение хлоропластов криптофит от красных водорослей подтверждается анализом 16S рРНК нуклеоморфа и ядер красных водорослей. Две наружные мембраны иногда называют пластидным эндоплазматическим ретикулумом.
Процесс доставки в хлоропласты белков, кодируемых ядерной ДНК, у криптофит организован гораздо более сложно, чем у растений, чьи пластиды представляют собой результат первичного, а не вторичного эндосимбиоза. У криптофит и других организмов, чьи хлоропласты появились в результате вторичного эндосимбиоза, белки, кодируемые ядром, но предназначенные для пластид, содержат на N-конце лидерную последовательность, состоящую из сигнального пептида и переходного пептида. Два этих сигнала совместно обеспечивают прохождение белком всех четырёх мембран для доставки к месту своего функционирования. Было высказано предположение, что в переносе белков в перипластидное пространство принимает участие кодируемая нуклеоморфом ЭПР-связанная система деградации. Поскольку эндосимбионт утратил свой ЭПР, то эта система могла приобрести новую функцию в транспорте белков.
НуклеоморфКак уже отмечалось выше, нуклеоморф представляет собой рудиментарное ядро эукариотического фототрофного эндосимбионта, инкорпорированного предками криптофитов в ходе вторичного эндосимбиоза (кроме криптофит, нуклеоморф также имеется у водорослей группы Chlorarachniophyta; однако в последнем случае происходила инкорпорация не красной, а зелёной водоросли). У криптофит нуклеоморф обычно имеет округлую или грушевидную форму; он окружён двойной мембраной, содержащей поры, сходные с ядерными поровыми комплексами, и имеет ядрышко с типичными эукариотическими РНК. Нуклеоморф может быть ассоциирован с пиреноидом; иногда он располагается внутри самого пиреноида, иногда — в выросте хлоропласта.
Размер ДНК нуклеоморфа Rhodomonas salina[en] составляет только 660 тыс. пар нуклеотидов в трёх хромосомах (240, 225 и 195 тыс. пар нуклеотидов). Таким образом, геном нуклеоморфа — мельчайший из описанных ядерных геномов эукариот. Интересно, что нуклеоморфы Chlorarachniophyta также имеют 3 хромосомы. У различных видов криптофит ДНК нуклеоморфа содержит от 450 до 850 тыс. пар нуклеотидов. Нуклеоморф имеет 531 ген, которые кодируют 30 белков, функционирующих в хлоропластах. Плотность генов в ДНК нуклеоморфов криптофит довольно высока. Гены нуклеоморфа в большинстве своём являются генами домашнего хозяйства, их продукты участвуют в таких процессах, как фолдинг и разрушение белков, транскрипция и трансляция. В 2001 году был секвенирован геном нуклеоморфа Guillardia theta, а в 2007 году — Hemiselmis andersenii.
Деление нуклеоморфа происходит в препрофазе деления главного ядра вслед за репликацией базального тела, но до деления хлоропласта. Деление амитотическое, и, хотя геном нуклеоморфа содержит гены α-, β- и γ-тубулина, микротрубочки не принимают участия в образовании веретена деления; вместо этого имеются фибриллярные структуры, появляющиеся только во время деления. Дочерние нуклеоморфы мигрируют в противоположные концы хлоропласта, поэтому каждый дочерний хлоропласт наследует один нуклеоморф. Единственная известная криптофитовая водоросль, не имеющая нуклеоморфа — Goniomonas, которая утратила пластиды. У другой бесцветной криптофитовой водоросли, Chilomonas[en], имеются лейкопласт и нуклеоморф.
СтигмаУ ряда видов криптофит имеется глазок (стигма). У криптофит он связан с хлоропластом, но не связан со жгутиковым аппаратом, а потому может располагаться и в центре клетки, как, например, у Chroomonas mesostigmatica. Стигма состоит из большого количества пигментных липидных глобул, расположенных параллельными рядами под оболочкой хлоропласта. Для некоторых криптофит показан положительный фототаксис. В частности, бесцветный Chilomonas наиболее чувствителен к синему свету при длине волны 366 нм.
ЯдроЯдра криптофит, как правило, невелики и не превышают в диаметре 3 мкм. Клетка имеет единственное ядро, расположенное в задней части клетки. Для ядер криптофит характерен сложный хромоцентрический тип организации. Хроматиновый ободок (скопление хроматина по краю ядра) может быть сплошным, почти сплошным или же узким и едва различимым, сильно разорванным. Остальное пространство ядра заполнено многочисленными глыбками хроматина, только у Rhinomonas panca они единичны. У различных криптофит имеется от 40 до 210 хромосом. В 2012 году было завершено секвенирование ядерного генома криптофитовой водоросли Guillardia theta[en], её ядерный геном оказался гаплоидным.
МитозМитоз у исследованных криптофит отличается поразительным сходством и относится к открытому центрическому типу. Процесс деления клеток начинается с удвоения базальных тел. Ядра перемещаются в переднюю часть клетки и иногда вплотную подходят к базальным телам. Базальные тела не меняют своего положения в течение всего митоза и во время метафазы располагаются по обе стороны метафазной пластинки. Они не становятся центриолями, но выполняют их функции, в частности, около них формируются многочисленные микротрубочки, участвующие в образовании веретена.
В профазе митоза ядерная оболочка фрагментируется и постепенно растворяется, лишь малая часть её фрагментов сохраняется до конца митоза. В случае открытого митоза у криптофит происходит формирование дополнительной перинуклеарной оболочки из хлоропластного ретикулума. При митозе ядрышко не исчезает.
Веретено деления криптофит также необычно. Оно состоит из параллельных неконвергирующих микротрубочек. Область полюсов веретена уплощена и ограничена цистернами ЭПР. На его концах, кроме ризостилей, которые отходят от каждой пары базальных тел и не контактируют с микротрубочками веретена, располагаются нуклеоморфы, специально переместившиеся сюда. В метафазе вместо типичной экваториальной пластинки из морфологически дифференцированных хромосом образуется очень компактное скопление хроматина, в котором отдельные хромосомы не различимы, однако образуются особые проходы для микротрубочек веретена. Одно время считали, что истинных хромосом у криптофит нет, а компактную массу хроматина можно рассматривать как одну сборную хромосому. Однако сейчас считают, что хромосомы криптофит очень мелкие, а их кинетохоры морфологически очень слабо дифференцированы. Роль кинетохоров, по-видимому, выполняют выступы хроматина в области коридоров, которые контактируют с микротрубочками веретена. При делении клеток также происходит формирование новой глотки, которая достаётся одной из дочерних клеток.
ЭжектосомыКлетки криптофит нередко несут особые стрекательные структуры — эжектосомы, или трихоцисты. Их строение отличается от стрекающих структур динофлагеллят, однако они, возможно, родственны R-тельцам каппа-частиц инфузорий. Крупные эжектосомы располагаются под плазмалеммой в области глотки, а более мелкие разбросаны по всей поверхности клетки. Каждая эжектосома окружена одной мембраной и содержит внутри 2 ленты, скрученные в цилиндры. Более крупный цилиндр состоит из намотанного на спирали лентовидного мембранного материала. В его верхней части находится V-образное углубление, в котором залегает второй цилиндр. По-видимому, эти два цилиндра связаны друг с другом при помощи волокон. При химическом и физическом раздражении скрученные ленты распрямляются и выстреливают, разрывая мембрану эжектосомы и перипласт. Сначала выбрасывается маленький цилиндр, он вытягивает за собой ленту большого цилиндра. Выстрелившая эжектосома с периферии клетки имеет длину 4 мкм, а крупная эжектосома, расположенная около глотки, после срабатывания имеет длину 20 мкм. Выстреливание эжектосом заставляет клетку двигаться в противоположном направлении. Кроме того, при выстреливании из эжектосом высвобождаются вещества, убивающие бактерий, которыми питаются криптофиты. Эжектосомы формируются в пузырьках аппарата Гольджи, причём количество оборотов в цилиндре увеличивается по мере созревания эжектосомы.
С наличием эжектосом связаны трудности по переводу криптофит на лабораторные среды. Их клетки чрезвычайно чувствительны к изменению окружающих условий (рН, осмотические условия, температура) и так сильно отстреливают содержимое эжектосом, что клетки быстро разрушаются.
Другие органеллыВ цитоплазме клеток криптофит имеется одна сетчатая митохондрия с пластинчатыми кристами. Она образует две сети: периферическую (лежащую под плазмалеммой) и центральную (в центре клетки).
На переднем конце тела у пресноводных криптофит располагаются сократительные вакуоли, изливающие своё содержимое в глотку или борозду. Они служат для регуляции осмотического давления внутри клетки.
В клетках криптофит имеются два тельца Мопа. Тельце Мопа представляет собой крупную везикулярную структуру, расположенную в передней части клетки. Точные функции телец Мопа неизвестны, но, так как они содержат много мембран и фибрилл, они могут принимать участие в разрушении клеточных органелл, особенно отработанных эжектосом, и других инородных частиц в цитоплазме.
МетаболизмСреди криптофит имеются автотрофы, гетеротрофы (сапротрофы и фаготрофы) и миксотрофы. Большинство криптофит нуждаются в витамине B12 и тиамине, а некоторые — в биотине. В качестве источников азота криптофиты могут использовать аммоний и органические азотсодержащие соединения, однако морские формы менее способны к превращениям нитрата в нитрит по сравнению с пресноводными представителями. Органические вещества стимулируют рост криптофит. Различные бесцветные формы, а также окрашенный Cryptomonas ovata способны к фагоцитозу, а сине-зелёный Chroomonas pochmannii является миксотрофом. Для захвата и удержания бактериальных клеток используются особые вакуоли. Бактерии втягиваются в них через маленькую пору в глотке, где нет пластинок перипласта, и в вакуолях же перевариваются. Некоторые криптофиты (например, Goniomonas truncata) могут поедать вирусные частицы. При фаготрофном питании криптофиты демонстрируют хемотаксис в направлении наибольшей концентрации питательных веществ.
Основное запасное вещество у криптофит — крахмал, он откладывается между второй и третьей мембранами хлоропласта; из всех водорослей только криптофиты откладывают крахмал в этом месте. Как и у красных водорослей, он содержит больше амилопектина, чем амилозы, поэтому с йодом даёт красное окрашивание. В цитоплазме могут иметься липидные капли.
隐藻门是一大类的藻类,[2] 大都具有色素体,淡水中常见。细胞大小约为10-50微米,形状扁平,有两个稍微不等长的鞭毛。一个著名特征是有红藻寄生于其细胞中,形成一种内共生关系,并把藻胆素带给宿主。
原本一般认为其与定鞭藻門关系更为密切,但2015年牛津大学卡瓦利埃 - 史密斯等人的研究[3]及2016年加拿大不列颠哥伦比亚大学帕特里克 · 基林等人的研究[4]显示其实际上是原始色素體生物的一部分,至于其与其他植物的系统发生关系则仍不明确。[3][4]
隐藻门是一大类的藻类, 大都具有色素体,淡水中常见。细胞大小约为10-50微米,形状扁平,有两个稍微不等长的鞭毛。一个著名特征是有红藻寄生于其细胞中,形成一种内共生关系,并把藻胆素带给宿主。
原本一般认为其与定鞭藻門关系更为密切,但2015年牛津大学卡瓦利埃 - 史密斯等人的研究及2016年加拿大不列颠哥伦比亚大学帕特里克 · 基林等人的研究显示其实际上是原始色素體生物的一部分,至于其与其他植物的系统发生关系则仍不明确。

クリプト藻は遊泳性の単細胞藻類で、淡水及び海水に広く分布する。人目に触れる機会は少ないが、一般的な池や湖沼の水を観察すればほぼ確実に目にする事ができる生物である。藻類の中でも小型の部類に入り、体長は3-50μm前後。およそ200種が知られているが、生活環の各ステージの細胞に別個の学名が与えられている、つまり命名が重複している可能性のある種が含まれている。
細胞はおおよそ扁平な米粒型で、米で言えば胚芽のあたりに2本の鞭毛を持つ。鞭毛基部の近傍にはガレット(咽喉部)と呼ばれる陥入部があり、ガレットの内壁にはこれを取り巻くようにトリコシスト(trichocyst; 毛胞)と呼ばれる射出装置が配されている。このトリコシストはちょうどトイレットペーパーが巻かれたような状態で収納されており、射出時にはこれが螺旋状に解けて展開する。ガレットの周囲には、他に眼点や反射体(refractile body)といった光学系の器官を持つ種もある。
クリプト藻の鞭毛は不等長〜亜等長で、種によって異なる。修飾としては鞭毛小毛や鱗片などが知られている。鱗片を持つ場合、この鱗片はロゼット状の特徴的な形態を示す。鞭毛小毛は鞭毛の軸に対して左右対称に生えているもの(両羽型)や片方にのみ生えているもの(片羽型)があり、2本の鞭毛における両羽/片羽の組み合わせも様々である。小毛は不等毛藻のそれとは異なり、軸部と先端毛から成る2部構成である(不等毛藻では基部を加えた3部構成)。クリプト藻の鞭毛小毛も不等毛藻と同様に推進力逆転の効果があると言われるが、前述のように片羽型の鞭毛や組み合わせの問題もあり、力学的な裏付けは取られていない。
クリプト藻の葉緑体はふつう細胞内に1つで、核やその他の細胞小器官を取り巻くように細胞表面に沿った形で配置されている。光合成色素はクロロフィルa/cとカロテノイド類、そしてフィコビリンタンパクとしてフィコシアニンやフィコエリスリン(フィコエリトリン)を含む。フィコビリンタンパクはチラコイドの内腔に詰まっている。その為クリプト藻のチラコイドは他の藻類よりも厚い。フィコシアニンを持つ Chroomonas 属では、細胞は藍藻のような青緑色を呈する。
クリプト藻の葉緑体は紅藻由来であるとされており、取り込まれた紅藻の核はヌクレオモルフとして葉緑体内に残存している。葉緑体膜は4重膜で、不等毛藻やハプト藻と同様に、最外膜は核膜と連絡している。ヌクレオモルフは外側の2枚(葉緑体ER)と内側の2枚との間の領域(periplastidal compartment)に位置しており、種によってはピレノイドに埋没する形になっている。この領域にはヌクレオモルフの他、真核型の80S rRNAやデンプン粒なども見られる。
クリプト藻の外被はペリプラスト(periplast)と呼ばれる。これは細胞膜とそれを裏打ちする板状の構造から成る。板状構造はタンパク質でできており、その規則的なパターンは分類形質としても有用である。
ヌクレオモルフを持つがゆえに、細胞内共生説との関係で研究者の注目を集めてきたクリプト藻であるが、さらにクリプト藻を細胞内に取り込んで保持する藻類や原生動物が報告されている。クリプト藻を葉緑体として活用する生物は、渦鞭毛藻や繊毛虫といったアルベオラータに多い。
淡水及び海水に広く分布する。北海や大西洋ではブルームの報告もある。
通常は無性的に二分裂で増殖する。少数ながら同形配偶による有性生殖も知られている。
古くから、クリプト藻はその生活環の中で異なる形態の鞭毛虫として生活する事が報告されていた。例えばProteomonas sulcata では複相世代が大型の鞭毛虫、単相世代が小型の鞭毛虫であり、それぞれのステージではペリプラストのパターンも異なる(Hill and Wetherbee 1986)。このような例はクリプト藻のごく一部だと考えられてきたが、実は大部分のクリプト藻に適用されるべき現象である事が近年報告された(Hoef-Emden and Melkonian 2003)。これにより、単相世代と複数世代における命名の重複が明らかになると共に、分類群の整理が要求される事となった。これはハプト藻と同様の問題であり、やはり培養技術や分子系統解析の発展に伴い明らかとなった問題である。
古くは渦鞭毛植物門と合わせて炎色植物門(Division Pyrrophyta)にまとめられた事もあったが、後に同植物門は解体されている。近年では独立のクリプト植物門(Division Cryptophyta, Cavalier-Smith 1986)として扱うか、クリプト藻綱(Class Cryptophyceae, Pascher 1913, emend. Schoenichen, 1925) として綱レベルでの集団として扱うのが主流である。なお、下記の分類も、前述の生活環の問題を受けて再編中のものである点に留意されたい。
クリプト藻は遊泳性の単細胞藻類で、淡水及び海水に広く分布する。人目に触れる機会は少ないが、一般的な池や湖沼の水を観察すればほぼ確実に目にする事ができる生物である。藻類の中でも小型の部類に入り、体長は3-50μm前後。およそ200種が知られているが、生活環の各ステージの細胞に別個の学名が与えられている、つまり命名が重複している可能性のある種が含まれている。
은편모조류(隱鞭毛藻類) 또는 갈색편모조류[1](褐色鞭毛藻類, Cryptophyceae), 암색편모충류(暗色鞭毛蟲類, cryptomonads 또는 cryptophytes)는 조류의 생물군으로[2] 대부분 엽록체를 가지고 있다. 담수에서 흔히 발견되며, 해양과 염분이 있는 서식 환경에서도 서식한다. 각 세포의 크기는 약 10-50 μm이고, 모양이 평평하며, 앞면에 홈 또는 주머니가 있다. 그 주머니의 끝단에 일반적으로 2개의 약간 일정하지 않은 편모가 있다.
2016년 실라(Silar)에 의한 하크로비아의 계통 분류는 다음과 같다.[4][5]
하크로비아 넓은 의미의 크립티스타 코르비헬리아 엔도헬레아상강 코르비스토마상강 좁은 의미의 크립티스타 팔피토아문 롤로모나스아문 레우크립토스상강 은편모조류 합티스타 육질태양충류 착편모조아문 RappemonadaRappephyceae
Haptomonada은편모조류(隱鞭毛藻類) 또는 갈색편모조류(褐色鞭毛藻類, Cryptophyceae), 암색편모충류(暗色鞭毛蟲類, cryptomonads 또는 cryptophytes)는 조류의 생물군으로 대부분 엽록체를 가지고 있다. 담수에서 흔히 발견되며, 해양과 염분이 있는 서식 환경에서도 서식한다. 각 세포의 크기는 약 10-50 μm이고, 모양이 평평하며, 앞면에 홈 또는 주머니가 있다. 그 주머니의 끝단에 일반적으로 2개의 약간 일정하지 않은 편모가 있다.
일부는 혼합영양을 하는 것으로 보인다.
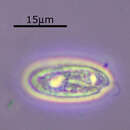


.jpg)




