Els Reoviridae són una família de virus que n'incloue alguns que afecten el sistema gastrointestinal (com el Rotavirus), i alguns que causen infeccions respiratòries. El material genètic dels virus d'aquesta família és la doble cadena d'ARN. El nom Reoviridae deriva de virus entèrics respiratoris orfes.
Els Reoviridae són una família de virus que n'incloue alguns que afecten el sistema gastrointestinal (com el Rotavirus), i alguns que causen infeccions respiratòries. El material genètic dels virus d'aquesta família és la doble cadena d'ARN. El nom Reoviridae deriva de virus entèrics respiratoris orfes.
Die Reoviridae bilden die größte Familie von unbehüllten Viren mit doppelsträngiger RNA (dsRNA). Zusammen mit den Birnaviridae und Partitiviridae sind sie die einzigen Viren, deren dsRNA-Genom segmentiert vorliegt. Zur Familie gehören so wichtige Erreger wie das Blauzungenvirus, die humanen Rotaviren oder das Colorado-Zeckenfieber-Virus. Der Name der Familie ist ein Akronym aus „respiratory, enteric, orphan“, da man der Ansicht war, dass Mitglieder dieser taxonomischen Gruppe nur Darmerkrankungen (enteric), Atemwegserkrankungen (respiratory) oder keine Erkrankung (orphan) hervorrufen würden. Tatsächlich wurden die Reoviridae bei vielen Säugetieren gefunden, ebenso bei Reptilien, Fischen, Krustentieren und Insekten. Drei Gattungen der Familie (Fijivirus, Phytoreovirus und Oryzavirus) umfassen Erreger bei Pflanzen (Pflanzenviren) und Pilzen (Mykoviren).
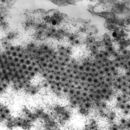
Die reifen Virusteilchenen (Virionen) der Reoviridae sind unbehüllte, ikosaedrische Kapside mit einem Durchmesser von etwa 60–80 nm. Charakteristisch für die meisten Gattungen der Familie (mit Ausnahme der Gattung Cypovirus) ist der Aufbau des Virions aus zwei ineinander geschachtelte und eng wechselwirkende Kapside (T=13), die wiederum eine innere, regelmäßige Corestruktur umgeben. Die so aus drei Lagen von Proteinen aufgebauten Virionen zeigen daher im Elektronenmikroskop eine sehr typische, dickwandige Struktur mit regelmäßigen, speichenförmigen Verdickungen. Von dieser Speichenstruktur abgeleitet erhielt die Gattung Rotavirus (lat. rota: Rad) ihren Namen. Diese Speichenstruktur entsteht durch 132 Öffnungen des äußeren Kapsids, die in ebenso viele Kanäle des inneren Kapsids münden. Diese Kanäle, die bis an das innere Core reichen, lassen eine Diffusion von Ionen in das Virion zu.
Dadurch, dass das innere Core und das innere Kapsid für die korrekte Verpackung der RNA-Segmente verantwortlich sind, kann das äußere Kapsid eine relativ große Variabilität der Oberflächenstrukturen aufweisen ohne die Stabilität des Virions und die Vollständigkeit des Genoms zu beeinträchtigen. Daher zeigen sich bei vielen Reoviren zahlreiche Spezies und Serotypen.
In der inneren Coreschale befindet sich ein Molekül der viralen RNA-Polymerase und Guanyltransferase. Bei einigen Gattungen sind Myristyl-Reste (Tetradecyl-Reste) kovalent an das Kapsid gebunden. Bei den Gattungen Rotavirus, Orbivirus und Coltivirus kommen unreife Virionen vor, die Reste einer Lipidhülle besitzen. Bei diesen Gattungen geschieht das Verlassen der Zelle durch Knospung an der Zellmembran (Orbivirus, Coltivirus) oder der Membran des Endoplasmatischen Retikulums (ER) (Rotavirus). Durch einen enzymatischen Reifungsschritt verlieren die Virionen die Lipidhülle wieder. Bei einigen Gattungen wird das äußere Kapsid durch wirtseigene Proteasen (z. B. Trypsin, Chymotrypsin) verändert, wodurch sogenannte „infectious subviral particles“ (ISVPs) entstehen. Nur die ISVPs sind infektiös und der Übergang von unreifen Virionen zu ISVPs spielt bei diesen Viren eine entscheidende Rolle in der Krankheitsentstehung und der unterschiedlichen Erscheinungsform der Virionen im Darm, Speichel oder Blutserum.
Das Genom besteht aus einer doppelsträngigen RNA, die je nach Gattung in 10 bis 12 Segmente unterteilt ist. Die Molmasse der Segmente reicht von 0,2 bis 3 × 106 Da. Die virale mRNA der Reoviridae besitzt keinen Poly-A-Schwanz. Bei einigen Virusspezies findet man kurze, einzelsträngige RNA-Oligonukleotide in der inneren Coreschale.
Mit Stand November 2018 gliedern sich nach Maßgabe des ICTV die Reoviriridae wie folgt (neben den Typusspezies ist nur eine Auswahl weiterer Spezies angegeben):[3]
Das folgende Kladogramm gibt vereinfacht die Verwandtschaftsbeziehungen wieder, wie sie im 9. Report des International Committee on Taxonomy of Viruses (ICTV, 2011) und von MacLachlan/Dubovi (2016);[4][11] sowie fast übereinstimmend von Belhouchet et al. (2010)[12] vorgeschlagen wurde:
Reoviridae SedoreovirinaeMimoreovirus (MPrV)
Orbivirus (SCRV, CHUV, AHSV, BTV, KEMV, TRBV, EHDV)
Phytoreovirus (RDV)
Seadornavirus (LNV, KDV, BAV)
Cardoreovirus (ESRV)
Rotavirus (RVA, RVB, RVC)
Orthoreovirus (MRV)
Oryzavirus (RRSV)
Dinovernavirus (APRV)
Coltivirus (EYAV, CTFV)
Für einige der Gattungen sind die abgekürzten Speziesnamen angegeben, diese umfassen jeweils einen oder mehrere der untersuchten zugehörigen Stämme (englisch strains).
In älteren Arbeiten gibt es noch gewisse Abweichungen (z. B. bei Attoui et al. (2006)[13] bei Quito-Avila (2011)[14] und bei Ke et al. (2010).[15]
Das ICTV hat mit der Master Species List #35 vom März 2020 die Reoviridae dem neu geschaffenen Phylum Duplornaviricota zugeordnet.[16] Eine Kladogramm findet sich bei Picornavirales §ICTV Master Species List #35.
Die Reoviridae bilden die größte Familie von unbehüllten Viren mit doppelsträngiger RNA (dsRNA). Zusammen mit den Birnaviridae und Partitiviridae sind sie die einzigen Viren, deren dsRNA-Genom segmentiert vorliegt. Zur Familie gehören so wichtige Erreger wie das Blauzungenvirus, die humanen Rotaviren oder das Colorado-Zeckenfieber-Virus. Der Name der Familie ist ein Akronym aus „respiratory, enteric, orphan“, da man der Ansicht war, dass Mitglieder dieser taxonomischen Gruppe nur Darmerkrankungen (enteric), Atemwegserkrankungen (respiratory) oder keine Erkrankung (orphan) hervorrufen würden. Tatsächlich wurden die Reoviridae bei vielen Säugetieren gefunden, ebenso bei Reptilien, Fischen, Krustentieren und Insekten. Drei Gattungen der Familie (Fijivirus, Phytoreovirus und Oryzavirus) umfassen Erreger bei Pflanzen (Pflanzenviren) und Pilzen (Mykoviren).
Sedoreoviridae (formerly Reoviridae) is a family of double-stranded RNA viruses. Member viruses have a wide host range, including vertebrates, invertebrates, plants, protists and fungi.[1] They lack lipid envelopes and package their segmented genome within multi-layered capsids. Lack of a lipid envelope has allowed three-dimensional structures of these large complex viruses (diameter ∼60–100 nm) to be obtained, revealing a structural and likely evolutionary relationship to the cystovirus family of bacteriophage.[2] There are currently 97 species in this family, divided among 15 genera in two subfamilies.[3] Reoviruses can affect the gastrointestinal system (such as rotaviruses) and respiratory tract.[4] The name "reo-" is an acronym for "respiratory enteric orphan" viruses.[5] The term "orphan virus" refers to the fact that some of these viruses have been observed not associated with any known disease. Even though viruses in the family Reoviridae have more recently been identified with various diseases, the original name is still used.
Reovirus infections occur often in humans, but most cases are mild or subclinical. Rotaviruses, however, can cause severe diarrhea and intestinal distress in children, and lab studies in mice have implicated orthoreoviruses in the expression of coeliac disease in pre-disposed individuals.[6] The virus can be readily detected in feces, and may also be recovered from pharyngeal or nasal secretions, urine, cerebrospinal fluid, and blood. Despite the ease of finding reoviruses in clinical specimens, their role in human disease or treatment is still uncertain.
Some viruses of this family, such as phytoreoviruses and oryzaviruses, infect plants. Most of the plant-infecting reoviruses are transmitted between plants by insect vectors. The viruses replicate in both the plant and the insect, generally causing disease in the plant, but little or no harm to the infected insect.[7]: 148
Reoviruses are non-enveloped and have an icosahedral capsid composed of an outer (T=13) and inner (T=2) protein shell.[1][8] Ultrastructure studies show that virion capsids are composed of two or three separate layers which depends on species type. The innermost layer (core) has T=1 icosahedral symmetry and is composed of 60 different types of structural proteins. The core contains the genome segments, each of them encode a variety enzyme structure which is required for transcription. The core is covered by capsid layer T=13 icosahedral symmetry. Reoviruses have a unique structure which is contains a glycolisated spike protein on the surface.[9]
The genomes of viruses in family Reoviridae contain 9–12 segments which are grouped into three categories corresponding to their size: L (large), M (medium) and S (small). Segments range from about 0.2 to 3 kbp and each segment encodes 1–3 proteins (10–14 proteins in total[1]). Proteins of viruses in the family Reoviridae are denoted by the Greek character corresponding to the segment it was translated from (the L segment encodes for λ proteins, the M segment encodes for μ proteins and the S segment encodes for σ proteins).[8]
Viruses in the family Reoviridae have genomes consisting of segmented, double-stranded RNA (dsRNA).[4] Because of this, replication occurs exclusively in the cytoplasm, and the virus encodes several proteins which are needed for replication and conversion of the dsRNA genome into positive-sense RNAs.[10]
The virus can enter the host cell via a receptor on the cell surface. The receptor is not known but is thought to include sialic acid and junctional adhesion molecules (JAMs).[10] The virus is partially uncoated by proteases in the endolysosome, where the capsid is partially digested to allow further cell entry. The core particle then enters the cytoplasm by a yet unknown process where the genome is transcribed conservatively causing an excess of positive-sense strands, which are used as messenger RNA templates to synthesize negative-sense strands.[10]
The genome of the rotavirus is divided into 11 segments. These segments are associated with the VP1 molecule which is responsible for RNA synthesizes. In early events, the selection process occurs so that the entry of the 11 different RNA segments go in the cell. This procedure is performed by newly synthesized RNAs. This event ensures that one each of the 11 different RNA segments is received. In late events, the transcription process occurs again but this time is not capped unlike the early events. For virus different amounts of RNAs are required therefore during the translation step there is a control machinery. There are the same quantities of RNA segments but different quantities of proteins. The reason for this is that the RNA segments are not translated at the same rate.[7]
Viral particles begin to assemble in the cytoplasm 6–7 hours after infection. Translation takes place by leaky scanning, suppression of termination, and ribosomal skipping. The virus exits the host cell by monopartite non-tubule guided viral movement, cell to cell movement, and existing in occlusion bodies after cell death and remaining infectious until finding another host.[1]
Multiplicity reactivation (MR) is the process by which two or more virus genomes, each containing inactivating genome damage, can interact within an infected cell to form a viable virus genome. McClain and Spendlove[11] demonstrated MR for three types of reovirus after exposure to ultraviolet irradiation. In their experiments, reovirus particles were exposed to doses of UV-light that would be lethal in single infections. However, when two or more inactivated viruses were allowed to infect individual host cells MR occurred and viable progeny were produced. As they stated, multiplicity reactivation by definition involves some type of repair. Michod et al.[12] reviewed numerous examples of MR in different viruses, and suggested that MR is a common form of sexual interaction in viruses that provides the benefit of recombinational repair of genome damages.
The family Reoviridae is divided into two subfamilies[13] based on the presence of a "turret" protein on the inner capsid.[14][15] From ICTV communications: "The name Spinareovirinae will be used to identify the subfamily containing the spiked or turreted viruses and is derived from 'reovirus' and the Latin word 'spina' as a prefix, which means spike, denoting the presence of spikes or turrets on the surface of the core particles. The term 'spiked' is an alternative to 'turreted', that was used in early research to describe the structure of the particle, particularly with the cypoviruses. The name Sedoreovirinae will be used to identify the subfamily containing the non-turreted virus genera and is derived from 'reovirus' and the Latin word 'sedo', which means smooth, denoting the absence of spikes or turrets from the core particles of these viruses, which have a relatively smooth morphology."[16]

The family Reoviridae is divided into the following subfamilies and genera:
Although reoviruses are mostly nonpathogenic in humans, these viruses have served as very productive experimental models for studies of viral pathogenesis.[17] Newborn mice are extremely sensitive to reovirus infections and have been used as the preferred experimental system for studies of reovirus pathogenesis.[2]
Reoviruses have been demonstrated to have oncolytic (cancer-killing) properties, encouraging the development of reovirus-based therapies for cancer treatment.[18][19][20]
Reolysin is a formulation of reovirus (Mammalian orthoreovirus serotype 3-dearing strain[21]) that is currently in clinical trials for the treatment of various cancers,[22] including studies currently developed to investigate the role of Reolysin combined with other immunotherapies.[21]
Sedoreoviridae (formerly Reoviridae) is a family of double-stranded RNA viruses. Member viruses have a wide host range, including vertebrates, invertebrates, plants, protists and fungi. They lack lipid envelopes and package their segmented genome within multi-layered capsids. Lack of a lipid envelope has allowed three-dimensional structures of these large complex viruses (diameter ∼60–100 nm) to be obtained, revealing a structural and likely evolutionary relationship to the cystovirus family of bacteriophage. There are currently 97 species in this family, divided among 15 genera in two subfamilies. Reoviruses can affect the gastrointestinal system (such as rotaviruses) and respiratory tract. The name "reo-" is an acronym for "respiratory enteric orphan" viruses. The term "orphan virus" refers to the fact that some of these viruses have been observed not associated with any known disease. Even though viruses in the family Reoviridae have more recently been identified with various diseases, the original name is still used.
Reovirus infections occur often in humans, but most cases are mild or subclinical. Rotaviruses, however, can cause severe diarrhea and intestinal distress in children, and lab studies in mice have implicated orthoreoviruses in the expression of coeliac disease in pre-disposed individuals. The virus can be readily detected in feces, and may also be recovered from pharyngeal or nasal secretions, urine, cerebrospinal fluid, and blood. Despite the ease of finding reoviruses in clinical specimens, their role in human disease or treatment is still uncertain.
Some viruses of this family, such as phytoreoviruses and oryzaviruses, infect plants. Most of the plant-infecting reoviruses are transmitted between plants by insect vectors. The viruses replicate in both the plant and the insect, generally causing disease in the plant, but little or no harm to the infected insect.: 148
Reoviridae es una familia de virus de ARN que infectan animales, protistas, hongos y plantas.[1] El genoma es ARN de cadena doble y por lo tanto se incluyen en el Grupo III de la Clasificación de Baltimore. El nombre de "Reoviridae" se deriva de "virus respiratorio entérico huérfano", en donde el término "virus huérfano" hace referencia al desconocimiento de alguna enfermedad a la que poder asociar al virus.
Los viriones no presentan envoltura. Sus cápsides, formadas por múltiples proteínas, tienen simetría icosaédrica (T-13) y están generalmente organizadas en dos capas concéntricas, una exterior y otra interior. Los genomas de estos virus consisten en 10-12 segmentos de ARN bicatenario que se agrupan en tres categorías de acuerdo a su tamaño: L (grande), M (mediano) y S (pequeño). Los segmentos tienen una longitud comprendida entre 3,9 y 1 kpb,[2] y cada segmento codifica 1-3 proteínas. Las proteínas se denotan por la letra griega correspondiente al segmento del que fue traducido (proteínas λ para el segmento L, μ para M, y σ para S).
Dado que estos virus tienen genomas ARN bicatenarios, la replicación se produce exclusivamente en el citoplasma. El virus codifica varias proteínas que son necesarias para la replicación y conversión del genoma ARN bicatenario en ARN monocatenario de sentido positivo o negativo. El virus puede entrar en la célula huésped a través de un receptor en la superficie de la célula. El receptor es desconocido pero se cree que incluye ácido siálico y moléculas de adhesión celular (JAMs). El virus presenta parte del endolisosoma sin revestir por proteasas, donde la cápsida es parcialmente digerida para permitir la entrada en la célula. El núcleo del virus entra entonces en el citoplasma mediante un proceso todavía desconocido y allí el genoma es transcrito conservativamente, produciendo un exceso de cadenas de sentido positivo, que se utilizan como plantillas para sintetizar ARNm de sentido negativo. Las partículas virales comienzan a ensamblarse en el citoplasma 6-7 horas después de la infección.
Una característica que distingue a los virus ARN bicatenarios, independentemente de la familia a la que pertenezcan, es su capacidad para llevar a cabo la transcripción de los segmentos de ARN bicatenarios bajo las condiciones apropiadas dentro de la cápsida. En todos estos virus, las enzimas requeridas para la transcripción endógena son, por tanto, parte de la estructura del virión.[3]
Las partículas virales comienzan a ensamblarse en el citoplasma 6 a 7 horas después de la infección. La traducción tiene lugar mediante exploración con fugas, supresión de la terminación y omisión de ribosomas. El virus sale de la célula huésped mediante un movimiento viral monopartito no guiado por túbulos, movimiento de célula a célula y existiendo en cuerpos de oclusión después de la muerte celular y permaneciendo infeccioso hasta encontrar otro huésped.
En los animales afectan al sistema gastrointestinal (como los Rotavirus) y a las vías respiratorias del huésped. Aunque recientemente han sido identificadas diversas enfermedades causadas por los virus de la familia Reoviridae, el nombre original aún se utiliza.
La infección se produce a menudo en seres humanos, pero la mayoría de los casos son leves o subclínicos. El virus puede ser fácilmente detectado en heces y también en secreciones nasales o faríngeas, orina, líquido cefalorraquídeo y sangre. A pesar de la facilidad con la que se detecta, el papel del virus en las enfermedades o el tratamiento es aún incierto. Actualmente se está investigando el uso de estos virus en la lucha contra el cáncer.
La taxonomía actualizada del ICTV (2021) clasifica la familia de la siguiente manera:[4]
Reoviridae es una familia de virus de ARN que infectan animales, protistas, hongos y plantas. El genoma es ARN de cadena doble y por lo tanto se incluyen en el Grupo III de la Clasificación de Baltimore. El nombre de "Reoviridae" se deriva de "virus respiratorio entérico huérfano", en donde el término "virus huérfano" hace referencia al desconocimiento de alguna enfermedad a la que poder asociar al virus.
Reobirusak Reoviridae familian sailkatzen diren birusak dira. Familia horretan genero desberdinak daude; Rotavirus, Cypivirus eta Orbivirus dira aipagarrienak.
Familiaren izena ingelesezko sigletatik dator: Reoviridae (Respiratory Enteric Orphan Virus), birus hauek arnas traktuan eta digestio-aparatuan sortzen baitituzte gaitzak; Orphan edo Umezurtz terminoa ere gehitu zitzaion, hasieran isolatu ziren lehenengo birusak ez baitzeuden inongo gaitzekin lotuak.
Reobirusak tamaina ertaineko birusak dira (60-80 nm). Material genetikoari dagokionez, harizpi bikoitzeko RNA dute. Hain zuzen ere, animalia-birusen artean reobirusak dira kate bikoitzeko RNA duten bakarrak. Genomaren beste ezaugarri bereziren bat zatituta dagoela izaten da (10-12 zatitan, generoaren arabera), ortomixobirusen antzera. Kapsidea ikosaedrikoa da, kanpoko bilgarririk gabekoa (birus "biluziak" baitira).
Harizpi bikoitzeko RNA-k ezin du zuzenean itzulpen genetikoa garatu. Hori dela eta, RNA horrek RNA-m sortzen du lehen transkripzioaren bitartez, birusak berak dituen entzimei esker.
Reobirusak giza-patogenoak dira. Batzuek digestio-aparatuari eragiten diote, gastroenteritis sortuz (errotabirusak, esaterako); beste batzuek arnas aparatuari, arnas patologiak sorraraziz.
Reobirusak Reoviridae familian sailkatzen diren birusak dira. Familia horretan genero desberdinak daude; Rotavirus, Cypivirus eta Orbivirus dira aipagarrienak.
Familiaren izena ingelesezko sigletatik dator: Reoviridae (Respiratory Enteric Orphan Virus), birus hauek arnas traktuan eta digestio-aparatuan sortzen baitituzte gaitzak; Orphan edo Umezurtz terminoa ere gehitu zitzaion, hasieran isolatu ziren lehenengo birusak ez baitzeuden inongo gaitzekin lotuak.
Reobirusak tamaina ertaineko birusak dira (60-80 nm). Material genetikoari dagokionez, harizpi bikoitzeko RNA dute. Hain zuzen ere, animalia-birusen artean reobirusak dira kate bikoitzeko RNA duten bakarrak. Genomaren beste ezaugarri bereziren bat zatituta dagoela izaten da (10-12 zatitan, generoaren arabera), ortomixobirusen antzera. Kapsidea ikosaedrikoa da, kanpoko bilgarririk gabekoa (birus "biluziak" baitira).
Harizpi bikoitzeko RNA-k ezin du zuzenean itzulpen genetikoa garatu. Hori dela eta, RNA horrek RNA-m sortzen du lehen transkripzioaren bitartez, birusak berak dituen entzimei esker.
Reobirusak giza-patogenoak dira. Batzuek digestio-aparatuari eragiten diote, gastroenteritis sortuz (errotabirusak, esaterako); beste batzuek arnas aparatuari, arnas patologiak sorraraziz.
(RLQ=window.RLQ||[]).push(function(){mw.log.warn("Gadget "ErrefAurrebista" was not loaded. Please migrate it to use ResourceLoader. See u003Chttps://eu.wikipedia.org/wiki/Berezi:Gadgetaku003E.");});Reoviridae eli Reovirukset on kaksijuosteisten RNA-virusten heimo, joka voi vaikuttaa ruoansulatuskanavaan ja hengityselimistöön. Reoviruksilla on segmentoitunutta kaksijuosteista RNA:ta (dsRNA) sisältävät genomit.[1] Heimon nimi tulee englanninkielisestä fraasista respiratory enteric orphan viruses.[2] Englanninkielinen termi "orphan virus" tarkoittaa virusta, johon ei ole yhdistetty yksikään tunnettu tauti. Alkuperäinen nimi on vieläkin käytössä, vaikka nykyään jotkin reovirukset on yhdistetty tiettyin tauteihin, esimerkiksi rotavirukseen.
Vaikka reovirustartunnat ovat vähäoireisia, ne ovat ihmisillä kuitenkin aika yleisiä. Virus on helposti löydettävissä ulosteista. Sen lisäksi sitä voi löytää nielu- tai nenäerityksistä, virtsasta, aivo-selkäydinnesteestä tai verestä. Siitä huolimatta, että reovirukset ovat helposti löydettävissä kliinisistä näytteistä, niiden rooli ihmisten taudeissa tai hoidoissa ei ole varma.
Reoviridae eli Reovirukset on kaksijuosteisten RNA-virusten heimo, joka voi vaikuttaa ruoansulatuskanavaan ja hengityselimistöön. Reoviruksilla on segmentoitunutta kaksijuosteista RNA:ta (dsRNA) sisältävät genomit. Heimon nimi tulee englanninkielisestä fraasista respiratory enteric orphan viruses. Englanninkielinen termi "orphan virus" tarkoittaa virusta, johon ei ole yhdistetty yksikään tunnettu tauti. Alkuperäinen nimi on vieläkin käytössä, vaikka nykyään jotkin reovirukset on yhdistetty tiettyin tauteihin, esimerkiksi rotavirukseen.
Vaikka reovirustartunnat ovat vähäoireisia, ne ovat ihmisillä kuitenkin aika yleisiä. Virus on helposti löydettävissä ulosteista. Sen lisäksi sitä voi löytää nielu- tai nenäerityksistä, virtsasta, aivo-selkäydinnesteestä tai verestä. Siitä huolimatta, että reovirukset ovat helposti löydettävissä kliinisistä näytteistä, niiden rooli ihmisten taudeissa tai hoidoissa ei ole varma.
La famille des Reoviridae comprend onze genres et inclut certains virus affectant, entre autres, le système digestif (tels que les Rotavirus), ou le système respiratoire. C’est la seule famille de l’ordre des Reovirales et de la classe des Resentoviricetes.
Certains virus de cette famille infectent les plantes, par exemple ceux des genres Fijivirus, Oryzavirus et Phytoreovirus.
Elle appartient au Groupe III des virus à ARN à double brin.
Leur taille varie de 60 à 80 nm.
Ce sont des virus non enveloppés possédant une capside de forme icosaédrique constituée de 2 ou 3 couches protéiques.
Ces virus peuvent être détectés dans les selles, mais également, selon les genres, dans les sécrétions nasales et pharyngées, dans l'urine, le liquide céphalo-rachidien et le sang.
Ces virus pourraient être source de maladies émergentes et ont pour certains au moins un potentiel zoonotique, au moins pour certains virus de cette famille détectés chez des chauve-souris[4].
Un nouveau réovirus dit Réovirus Piscine (Piscine signifiant qu'il affecte les poissons), a été découvert en 2010 dans des élevages salmonicoles norvégiens. Son génome a été séquencé[5]. Il est la cause d'épidémies chez les saumons d'élevage en Norvège depuis quelques années (il affecte le cœur, en provoquant des myocardites[6]) et le squelette des saumons d'élevage[7]. Il a été détecté récemment loin de la Norvège, au Chili, dans des saumons atlantiques d'élevage[8],[9].
Une épidémie d'orthoreovirus a été signalée (2000) chez des babouins, cause de méningoencéphalomyélites[10].
Des virus de cette famille ont aussi été détectés chez des serpents Elaphe (de la famille des Colubridae)[11].
La famille des Reoviridae comprend onze genres et inclut certains virus affectant, entre autres, le système digestif (tels que les Rotavirus), ou le système respiratoire. C’est la seule famille de l’ordre des Reovirales et de la classe des Resentoviricetes.
Certains virus de cette famille infectent les plantes, par exemple ceux des genres Fijivirus, Oryzavirus et Phytoreovirus.
Reoviridae è una famiglia di virus a doppio filamento di RNA che possono colpire il sistema gastrointestinale (come Rotavirus) e il tratto respiratorio.
De Reoviridae betreft een virusfamilie van het geslacht Orbivirus.
De naam "Reoviridae" is afgeleid van "respiratory enteric orphan viruses" – respiratoir enterisch weesvirus. De uitdrukking "wees" verwijst naar het gegeven dat, toen het virus voor het eerst werd beschreven, er geen associatie was met enige ziekte. Hoewel nu diverse ziektebeelden toe te wijzen zijn aan reovirussen, is de naam niet meer veranderd.
Reovirusinfecties komen vaak voor bij de mens, maar deze verlopen vaak mild of subklinisch.
Er zijn 12 genera beschreven, waarvan het rotavirus klinisch het meest bekend is: een veel voorkomende veroorzaker van diarree. Bij dierlijke infecties zijn het blauwtongvirus en het Afrikaanse-paardenpestvirus berucht.
Het reovirus heeft dubbelstrengs RNA als drager van genetische informatie. Het infecteert en lyseert voornamelijk cellen waarin het ras (eiwit) geactiveerd is.
Omdat het Reovirus selectief cellen vernietigt met een geactiveerd ras eiwit, kan het virus heel goed gebruikt worden bij kankerbestrijding. Ongeveer 30% van alle kankersoorten ontstaan doordat het gen voor Ras gemuteerd is waardoor het eiwit permanent geactiveerd is.
De efficiëntie van het Reovirus om tumoren te vernietigen werd bekeken in uitzaaiingen van alvleesklierkanker in de lever van hamsters. Een behandeling met het virus reduceerde de grootte van de tumoren en het gezonde omliggende weefsel werd niet aangetast.[1]
De Reoviridae betreft een virusfamilie van het geslacht Orbivirus.
De naam "Reoviridae" is afgeleid van "respiratory enteric orphan viruses" – respiratoir enterisch weesvirus. De uitdrukking "wees" verwijst naar het gegeven dat, toen het virus voor het eerst werd beschreven, er geen associatie was met enige ziekte. Hoewel nu diverse ziektebeelden toe te wijzen zijn aan reovirussen, is de naam niet meer veranderd.
Reovirusinfecties komen vaak voor bij de mens, maar deze verlopen vaak mild of subklinisch.
Er zijn 12 genera beschreven, waarvan het rotavirus klinisch het meest bekend is: een veel voorkomende veroorzaker van diarree. Bij dierlijke infecties zijn het blauwtongvirus en het Afrikaanse-paardenpestvirus berucht.
Het reovirus heeft dubbelstrengs RNA als drager van genetische informatie. Het infecteert en lyseert voornamelijk cellen waarin het ras (eiwit) geactiveerd is.
Reowirusy (łac. Reoviridae, od ang. respiratory enteric orphan - sieroce[a] wirusy oddechowe i jelitowe) – rodzina wirusów, charakteryzujących się następującymi cechami:
Systematyka reowirusów przedstawia się następująco:
Dla człowieka chorobotwórcze są orbiwirusy, wirus gorączki kleszczowej Kolorado oraz rotawirusy. Pierwsze dwa wywołują choroby gorączkowe przenoszone przez stawonogi, natomiast rotawirusy powodują choroby biegunkowe.
Reowirusy (łac. Reoviridae, od ang. respiratory enteric orphan - sieroce wirusy oddechowe i jelitowe) – rodzina wirusów, charakteryzujących się następującymi cechami:
Symetria: ikosaedralna Otoczka lipidowa: brak, choć wiriony mogą mieć bardzo skomplikowaną budowę Kwas nukleinowy: dsRNA, segmentowany (10-12 cząsteczek) Replikacja: zachodzi w cytoplazmie Wielkość: 75–80 nm Gospodarz: kręgowce, bezkręgowce i rośliny Cechy dodatkowe: ze względu na segmentację można wyróżnić tzw. elektroferotypy, które cechują się, ze względu na różnice w budowie segmentów RNA, odmienną prędkością ruchu w żelu podczas elektroforezy. Elektroferotypy nie są tożsame z serotypami.Systematyka reowirusów przedstawia się następująco:
Rodzina: Reoviridae (reowirusy) Rodzaj Orthoreovirus Rodzaj Orbivirus (orbiwirusy) Rodzaj Rotavirus (rotawirusy) Rotavirus A (RV-A) Rotavirus B (RV-B) Rodzaj Coltivirus wirus gorączki kleszczowej Kolorado (CTFV, z ang. Colorado tick fever virus) Rodzaj Aquareovirus Rodzaj Cypovirus Rodzaj Fijivirus – wyłącznie roślinne Rodzaj Phytoreovirus – wyłącznie roślinne Rodzaj Oryzavirus – wyłącznie roślinneDla człowieka chorobotwórcze są orbiwirusy, wirus gorączki kleszczowej Kolorado oraz rotawirusy. Pierwsze dwa wywołują choroby gorączkowe przenoszone przez stawonogi, natomiast rotawirusy powodują choroby biegunkowe.
Reoviridae é uma família de vírus ARN de vertebrados que podem afetar o sistema gastrointestinal (como os Rotavirus) e as vias respiratórias do hospedeiro.[1] Atingem um ampla variedade de hospedeiros, incluindo vertebrados, invertebrados, plantas e fungos.[2] O genoma é ARN de dupla hélice e portanto está incluída no Grupo III da Classificação de Baltimore. O nome "Reoviridae" deriva de "vírus respiratório entérico órfão", em que o termo "vírus órfão" alude à ausência de qualquer doença à qual poder o vírus pode estar associado. Embora recentemente foram identificadas diversas doenças causadas pelos vírus da família Reoviridae, o nome original é ainda utilizado.
Uma característica que distingue os vírus de RNA bicatenários, independentemente da família a que pertençam, é a sua capacidade para levar a cabo a transcrição dos segmentos de RNA bicatenários sob condições apropriadas dentro da cápside. Em todos estes vírus, as enzimas requeridas para a transcrição endógena são, portanto, parte da estrutura do virião.[3]
Reoviridae é uma família de vírus ARN de vertebrados que podem afetar o sistema gastrointestinal (como os Rotavirus) e as vias respiratórias do hospedeiro. Atingem um ampla variedade de hospedeiros, incluindo vertebrados, invertebrados, plantas e fungos. O genoma é ARN de dupla hélice e portanto está incluída no Grupo III da Classificação de Baltimore. O nome "Reoviridae" deriva de "vírus respiratório entérico órfão", em que o termo "vírus órfão" alude à ausência de qualquer doença à qual poder o vírus pode estar associado. Embora recentemente foram identificadas diversas doenças causadas pelos vírus da família Reoviridae, o nome original é ainda utilizado.
Uma característica que distingue os vírus de RNA bicatenários, independentemente da família a que pertençam, é a sua capacidade para levar a cabo a transcrição dos segmentos de RNA bicatenários sob condições apropriadas dentro da cápside. Em todos estes vírus, as enzimas requeridas para a transcrição endógena são, portanto, parte da estrutura do virião.
Familia Reoviridae (Respiratory Enteritic Orphan virus) cuprinde specii de virusuri ce produc boli ale aparatului digestiv-afecțiuni gastrointestinale- (Rotavirus) iar unele pot determina afecțiuni respiratorii.
Fac parte din categoria virusurilor cu ARN bicatenar.
Familia Reoviridae (Respiratory Enteritic Orphan virus) cuprinde specii de virusuri ce produc boli ale aparatului digestiv-afecțiuni gastrointestinale- (Rotavirus) iar unele pot determina afecțiuni respiratorii.
Віруси в лабораторних умовах вирощують на культурах клітин нирок мавп, легенях та шкірно-м'язовій тканині ембріона людини, клітинах HeLa тощо. Розмножується повільно, цитопатична дія проявляється приблизно на 10 добу. Віруси характеризуються термостабільністю, зберігають життєздатність після 2-годинного нагрівання при температурі 56°С. Інфекційні властивості вірусів зберігаються при рН 3,0 більше доби.
Віріон має форму ікосаедра розміром 70 — 80 нм, зовнішньої оболонки немає, капсид поділяють на зовнішній та внутрішній, зовнішній має невизначену кількість та розміри капсомерів та може бути видалений трипсином, внутрішній має капсомери розміром близько 4 нм.
Реовіруси поділяють на підродини і роди. Орбі-, рота-, ортореовіруси мають внутрішню серцевину приблизно однакових розмірів 50-60 нм, з ікосаедричною симетрією та з субодиницями (капсомерами). У випадку ортореовірусів серцевину в основному складають три білка λ1, λ2, σ2; в невеликій кількості присутні також три інших білка λ3, μ1, μ2. Білок λ2 утворює «шипи» на 12 вершинах серцевини. Серцевину ротавірусів на 80 % складає білок VP6, майже всю частку, яка залишилася, складає білок VP2. В невеликій кількості в серцевині міститься ще один білок VP1. Серцевина орбівірусів складається з п'яти білків P1, P2, P4, P6, P7. Різні представники родини мають суттєву різницю в структурі зовнішнього капсиду. У ортореовірусів зовнішній капсид чітко виражений і складається з гекса- і пентагональних субодиниць, які складають в основному два білки — μ1С і σ3. У кожній з 12 вершин ікосаедра розміщено по дві молекули білка σ1. Нуклеокапсиди орбівіруси оточені дифузним шаром, який складають два білки — Р1, Р2. Цей зовнішній капсид не містить виражених морфологічних структур, як і капсид ротавірусів. Але останній має більш чіткі рельєфні особливості, його складають два білки — VP3, VP7.
Інфекційні субвірусні частинки прикріплюються до сіалофільних глікопротеїнвих рецепторів клітин за допомогою відростків, розташованих на 12 вершинах ікосаедричного віріона. Тканевий тропізм кожного виду реовірусів визначають варіації білка, який утворює відростки. Після адсорбції на клітині, віруси шляхом ендоцитозу потрапляють у цитоплазму, віріони руйнуються з утворенням серцевинних частинок. Кожна така частинка містить РНК-полімеразу (транскриптазу) та ферменти для синтезу мРНК, яка виходить у цитоплазму клітини через канали серцевинних частинок. РНК-полімераза використовує негативні ланцюги кожного РНК-сегменту як матрицю. Після початку синтезу мРНК, починається синтез геномної РНК для нових генерацій субвірусних частинок. Утворюються дволанцюгові РНК, які є матрицями для синтезу нових мРНК, а ті в свою чергу — матрицями для синтезу вірусних білків.
Різні представники родини Reoviridae мають приблизно однакову плавучу щільність у градієнті щільності CsCl (1,36 г/см3). При 70°С вони залишаються стабільними як завгодно тривало й навіть при звичайній температурі втрачають інфекційність досить повільно. Орто- і ротавіруси стійкі до хлороформу та інших органічних розчинників і стабільні в широкому діапазоні pH. Орбівіруси менш стійкі, звичайно їх інфекційність зменшується після обробки детергентами й органічними розчинниками в стандартних умовах. При низьких рН орбівіруси втрачають свою інфекційність, різко відрізняючись цим від інших представників родини Reoviridae, стабільних в інтервалі рН від 3 до 9.
Коефіцієнт седиментації 510 S20w
Усі представники родини Reoviridae містять сегментований геном, якого представляє дволанцюгова РНК. В орторео- і орбівірусів таких сегментів 10, у ротавірусів — 11. Найбільшу молекулярну масу має геном ортореовірусів (15*106), а найменшу — геном орбівірусів (12*106); молекулярна маса генома ротавірусів становить (11-14)*106. Розмір геному у орторео-, орбі-, рота-, колті- й аквареовірусів відповідно становить 23, 18, 16-21, 27, 15 тпн. Чітка відповідність між різними класами сегментів РНК різних груп реовірусів відсутня. В ортореовірусів молекулярна маса найбільшого сегмента дорівнює приблизно 2,7*106, в орбівірусів 2,5*106, у ротавірусів 1,1*106. Самі низькомолекулярні сегменти рота- і орбівірусів менші, ніж у реовірусів: їх молекулярна маса становить 2*105, Ці величини відповідають розмірам від 4 500 пар нуклеотидов (сегмент L-реовірусів) до 680 (малий сегмент ротавірусів).
Стратегія реплікації цитоплазматична:
Ліпіди відсутні.
Вуглеводи входять до складу глікопротеїдів, що знаходяться в складі зовнішнього капсиду.
Вісім поліпептидів є структурними білками і їх позначають грецькими літерами: λ1, λ2, λ3 (1400—1500 амінокислотних залишків); μ1, μ2 (700—800 амінокислотних залишків); δ1, δ2, δ3 (350—400 амінокислотні залишки). П'ять поліпептидів входять до складу серцевини, три — μ1С, δ1, δ3 — до складу зовнішнього капсиду. Білок δ1 забезпечує специфічне зв'язування з клітинними рецепторами і визначає тропізм вірусу до певних клітин. Цей же білок є вірусним гемаглютиніном. Він визначає собою основний типоспецифічний антиген. Тим більше, кількість цього білка на поверхні віріонів невелика — 24 молекули на вірусну частку і 1 % від сухої маси всіх білків. Білок μ1С забезпечує проникнення вірусу в клітину і його поширення в організмі. Цей білок утворюється шляхом протеолітичного нарізання білка μ1. У найбільшій кількості на поверхні вірусу представлено білок δ3. У серцевині віріонів знаходиться РНК-залежна РНК-транскриптаза, яка стає активною після видалення зовнішнього капсиду. Також є фосфогідролаза, гуанілтрансфераза і метилтрансфераза.
Білок δ1 являє собою основний типоспецифічний антиген.
Ортореовіруси, вочевидь, не є збудниками яких-небудь захворювань людини. Вважають, що ротавіруси є одними з основних збудників ентериту немовлят. Діарея є однією з основних причин дитячої смертності, особливо в слаборозвинених країнах. Ротавіруси спричиняють також діарею в корів і свиней. Орбівіруси, за винятком вірусу колорадської кліщової гарячки, спричинюють в основному захворювання у тварин. Вони відрізняються від інших представників родини Reoviridae, оскільки їхній життєвий цикл обов'язково включає стадію, коли вони персистують в організмі переносника — комахи. Орбівірусі зумовлюють ряд захворювань у тварин, включаючи африканську хворобу коней, энцефалоз коней, геморагічну хворобу оленів і корів, хворобу синього язика овець. Вірус колорадської кліщової гарячки спричинює в людей тяжке захворювання, подібне гарячці денге, а також енцефаліт. Ця хвороба розвивається після укусу кліща Dermacentor andersoni, вона зустрічається у Скелястих горах США.
Механізми передачі в природі — основним є повітряно-крапельний, однак можливий і фекально-оральний. Реовіруси у зв'язку з їх високою контагіозністю і стійкістю в навколишньому середовищі часто є причиною епідеміологічних спалахів. у циркуляції вірусу важливу роль відіграє безсимптомне носійство вірусу.
Вірус виділяють з слизу дихальних шляхів або з фекалій хворих шляхом посіву на культуру клітин. Серологічний діагноз ставлять по результатах реакцій зв'язування комплементу, гальмування гемаглютинації та нейтралізації.
Реовіруси (рос.)
Reoviridae
Подсемейства Группа по БалтиморуIII: дцРНК-вирусы
Реови́русы[2] (лат. Reoviridae, от англ. respiratory enteric orphan viruses) — семейство сферических вирусов, геном которых состоит из сегментированной двуцепочечной РНК.
Обнаружены в 1959 году[3]. Вирусные частицы имеют диаметр 60—80 нм, просто устроенные, так как нет суперкапсида, содержат белки формирующие шипы на поверхности VP4, VP7, в сердцевине — VP1, 2, 3, 6, каждый из которых играет важную роль в взаимодействии с клеткой. Размножаются в цитоплазме. Реовирусы необычайно устойчивы. Сохраняют инфекционные свойства при −20 °C около 2 лет, 4 °C — более 70 дней, 37 °C — 15—20 дней. Устойчивы к эфиру, кислотам, формалину. Реовирусы поражают молодняк животных.
По данным Международного комитета по таксономии вирусов (ICTV), на март 2017 года в семейство включают 15 родов, объединённых в 2 подсемейства[4]:
Реови́русы (лат. Reoviridae, от англ. respiratory enteric orphan viruses) — семейство сферических вирусов, геном которых состоит из сегментированной двуцепочечной РНК.
正呼腸孤病毒屬 Orthoreovirus
環狀病毒屬 Orbivirus
輪狀病毒 Rotavirus
科羅拉多壁蝨熱病毒 Coltivirus
水產呼腸孤病毒 Aquareovirus
質型多角體病毒 Cypovirus
斐濟病毒屬 Fijivirus]
植物呼腸孤病毒屬 Phytoreovirus
水稻病毒屬 Oryzavirus
Idnoreovirus
Mycoreovirus
呼腸孤病毒科(Reoviridae),是由respiratory enteric orphan virus各取其第一個字母而來,包括了許多影響胃腸系統的病毒,像是輪狀病毒Rotavirus,一部分的呼腸孤病毒會導致呼吸系統感染。
呼腸孤病毒科中的病毒,其遺傳物質為雙股RNA。
呼腸孤病毒科(Reoviridae),是由respiratory enteric orphan virus各取其第一個字母而來,包括了許多影響胃腸系統的病毒,像是輪狀病毒Rotavirus,一部分的呼腸孤病毒會導致呼吸系統感染。
呼腸孤病毒科中的病毒,其遺傳物質為雙股RNA。
 分類(ウイルス) 群 : 第3群(2本鎖RNA) 目 : 未割当 科 : レオウイルス科 Reoviridae 亜科
分類(ウイルス) 群 : 第3群(2本鎖RNA) 目 : 未割当 科 : レオウイルス科 Reoviridae 亜科 レオウイルス科(Reoviridae)はウイルスの分類における一科であり、RNAウイルスの一群の総称。そのビリオンは直径60 - 80nmの正二十面体構造を示し、10 - 12本の線状の二本鎖RNAをゲノムに持つ。エンベロープを持たないため、エーテルに対しては抵抗性である。細胞質内で増殖し、核周辺部に好酸性の細胞質内封入体を形成する。レオウイルス科のウイルスは同属のウイルス種間で遺伝子再集合を起こすことがある。オルビウイルス属およびコルチウイルス属のウイルスはアルボウイルスである。
レオウイルス科(Reoviridae)はウイルスの分類における一科であり、RNAウイルスの一群の総称。そのビリオンは直径60 - 80nmの正二十面体構造を示し、10 - 12本の線状の二本鎖RNAをゲノムに持つ。エンベロープを持たないため、エーテルに対しては抵抗性である。細胞質内で増殖し、核周辺部に好酸性の細胞質内封入体を形成する。レオウイルス科のウイルスは同属のウイルス種間で遺伝子再集合を起こすことがある。オルビウイルス属およびコルチウイルス属のウイルスはアルボウイルスである。
레오바이러스과(Reoviridae), 레오바이러스(영어: Reovirus, 문화어: 레오비루스)는 바이러스가 소화계통(로타바이러스처럼) 또는 기도(호흡기계통)에 영향을 주는 과이다. 이 레노바이러스과에 속하는 바이러스들은 겹가닥 RNA(dsRNA)를 이루는 게놈이다.[1]
레오바이러스과에는 크게 Sedoreovirinae, Spinareovirinae라는 아과로 나뉜다.








