Schizosaccharomyces pombe ist eine Spalthefe, d. h. ein Hefepilz, der sich nicht durch Sprossung (Knospung), sondern durch Teilung der Zelle in zwei Hälften („Spaltung“) vermehrt. Es handelt sich um einen stäbchenförmigen einzelligen Eukaryoten, der in der Molekular- und Zellbiologie häufig als Modellorganismus verwendet wird.[1]
Aus ostafrikanischem Hirsebier isolierte Paul Lindner 1893 am Institut für Gärungsgewerbe diese Spalthefe. Der Name S. pombe stammt vom Swahili-Wort für Bier (Pombe).
Als ein Modellorganismus in der Zellbiologie wurde es von Murdoch Mitchison in den 1950er Jahren eingeführt. Der britische Biochemiker Paul Nurse erhielt für seine Arbeiten über die Zellzyklusregulation in der Spalthefe im Jahr 2001 zusammen mit Leland H. Hartwell und Tim Hunt den Nobelpreis für Physiologie oder Medizin.
Die Genom-DNA-Sequenz von S. pombe wurde 2002 publiziert.[2]
2006 publizierten Akihisa Matsuyama und Kollegen die subzelluläre Lokalisation aller Proteine.[3]
Etwa 160 natürliche Stämme von Schizosaccharomyces pombe sind identifiziert worden, die unter anderem aus Europa, Asien und Amerika stammen. Die Mehrzahl dieser Varianten wurden auf Früchten wie Äpfel und Weintrauben oder in alkoholischen Getränken, wie dem brasilianischen Cachaça, gefunden. Schizosaccharomyces pombe ist auch in fermentiertem Kombucha-Tee enthalten.[4] Es ist nicht bekannt, ob die Hefe maßgeblich an der Fermentation beteiligt ist oder lediglich als Begleiter von aktiveren Mikroorganismen auftritt. Schizosaccharomyces-Hefen sind bislang noch nicht ausgiebig untersucht worden.
Schizosaccharomyces pombe ist eine Spalthefe, d. h. ein Hefepilz, der sich nicht durch Sprossung (Knospung), sondern durch Teilung der Zelle in zwei Hälften („Spaltung“) vermehrt. Es handelt sich um einen stäbchenförmigen einzelligen Eukaryoten, der in der Molekular- und Zellbiologie häufig als Modellorganismus verwendet wird.
Schizosaccharomyces pombe, also called "fission yeast", is a species of yeast used in traditional brewing and as a model organism in molecular and cell biology. It is a unicellular eukaryote, whose cells are rod-shaped. Cells typically measure 3 to 4 micrometres in diameter and 7 to 14 micrometres in length. Its genome, which is approximately 14.1 million base pairs, is estimated to contain 4,970 protein-coding genes and at least 450 non-coding RNAs.[2]
These cells maintain their shape by growing exclusively through the cell tips and divide by medial fission to produce two daughter cells of equal size, which makes them a powerful tool in cell cycle research.
Fission yeast was isolated in 1893 by Paul Lindner from East African millet beer. The species name pombe is the Swahili word for beer. It was first developed as an experimental model in the 1950s: by Urs Leupold for studying genetics,[3][4] and by Murdoch Mitchison for studying the cell cycle.[5][6][7]
Paul Nurse, a fission yeast researcher, successfully merged the independent schools of fission yeast genetics and cell cycle research. Together with Lee Hartwell and Tim Hunt, Nurse won the 2001 Nobel Prize in Physiology or Medicine for their work on cell cycle regulation.
The sequence of the S. pombe genome was published in 2002, by a consortium led by the Sanger Institute, becoming the sixth model eukaryotic organism whose genome has been fully sequenced. S. pombe researchers are supported by the PomBase MOD (model organism database). This has fully unlocked the power of this organism, with many genes orthologous to human genes identified - 70% to date,[8][9] including many genes involved in human disease.[10] In 2006, sub-cellular localization of almost all the proteins in S. pombe was published using green fluorescent protein as a molecular tag.[11]
Schizosaccharomyces pombe has also become an important organism in studying the cellular responses to DNA damage and the process of DNA replication.
Approximately 160 natural strains of S. pombe have been isolated. These have been collected from a variety of locations including Europe, North and South America, and Asia. The majority of these strains have been collected from cultivated fruits such as apples and grapes, or from the various alcoholic beverages, such as Brazilian Cachaça. S. pombe is also known to be present in fermented tea, kombucha.[12] It is not clear at present whether S. pombe is the major fermenter or a contaminant in such brews. The natural ecology of Schizosaccharomyces yeasts is not well-studied.
Schizosaccharomyces pombe was first discovered in 1893 when a group working in a Brewery Association Laboratory in Germany was looking at sediment found in millet beer imported from East Africa that gave it an acidic taste. The term schizo, meaning "split" or "fission", had previously been used to describe other Schizosaccharomycetes. The addition of the word pombe was due to its isolation from East African beer, as pombe means "beer" in Swahili. The standard S. pombe strains were isolated by Urs Leupold in 1946 and 1947 from a culture that he obtained from the yeast collection in Delft, The Netherlands. It was deposited there by A. Osterwalder under the name S. pombe var. liquefaciens, after he isolated it in 1924 from French wine (most probably rancid) at the Federal Experimental Station of Vini- and Horticulture in Wädenswil, Switzerland. The culture used by Urs Leupold contained (besides others) cells with the mating types h90 (strain 968), h- (strain 972), and h+ (strain 975). Subsequent to this, there have been two large efforts to isolate S. pombe from fruit, nectar, or fermentations: one by Florenzano et al.[13] in the vineyards of western Sicily, and the other by Gomes et al. (2002) in four regions of southeast Brazil.[14]
The fission yeast S. pombe belongs to the divisio Ascomycota, which represents the largest and most diverse group of fungi. Free-living ascomycetes are commonly found in tree exudates, on plant roots and in surrounding soil, on ripe and rotting fruits, and in association with insect vectors that transport them between substrates. Many of these associations are symbiotic or saprophytic, although numerous ascomycetes (and their basidiomycete cousins) represent important plant pathogens that target myriad plant species, including commercial crops. Among the ascomycetous yeast genera, the fission yeast Schizosaccharomyces is unique because of the deposition of α-(1,3)-glucan or pseudonigeran in the cell wall in addition to the better known β-glucans and the virtual lack of chitin. Species of this genus also differ in mannan composition, which shows terminal d-galactose sugars in the side-chains of their mannans. S. pombe undergo aerobic fermentation in the presence of excess sugar.[15] S. pombe can degrade L-malic acid, one of the dominant organic acids in wine, which makes them diverse among other Saccharomyces strains.
The yeast species Schizosaccharomyces pombe and Saccharomyces cerevisiae are both extensively studied; these two species diverged approximately 300 to 600 million years before present,[16] and are significant tools in molecular and cellular biology. Some of the technical discriminants between these two species are:
S. pombe gene products (proteins and RNAs) participate in many cellular processes common across all life. The fission yeast GO slim provides a categorical high level overview of the biological role of all S. pombe gene products.[8]

The fission yeast is a single-celled fungus with simple, fully characterized genome and a rapid growth rate. It has long been used in brewing, baking, and molecular genetics. S. pombe is a rod-shaped cell, approximately 3 μm in diameter, that grows entirely by elongation at the ends. After mitosis, division occurs by the formation of a septum, or cell plate, that cleaves the cell at its midpoint.
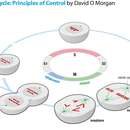
The central events of cell reproduction are chromosome duplication, which takes place in S (Synthetic) phase, followed by chromosome segregation and nuclear division (mitosis) and cell division (cytokinesis), which are collectively called M (Mitotic) phase. G1 is the gap between M and S phases, and G2 is the gap between S and M phases. In the fission yeast, the G2 phase is particularly extended, and cytokinesis (daughter-cell segregation) does not happen until a new S (Synthetic) phase is launched.
Fission yeast governs mitosis by mechanisms that are similar to those in multicellular animals. It normally proliferates in a haploid state. When starved, cells of opposite mating types (P and M) fuse to form a diploid zygote that immediately enters meiosis to generate four haploid spores. When conditions improve, these spores germinate to produce proliferating haploid cells.[19]

General features of the cell cycle.
The particular cell cycle of a fission yeast.

Division stages of Schizosaccharomyces in bright and dark field light microscopy

The general features of cytokinesis are shown here. The site of cell division is determined before anaphase. The anaphase spindle (in green on the figure) is then positioned so that the segregated chromosomes are on opposite sides of the predetermined cleavage plane.

In fission yeast, where growth governs progression through G2/M, a wee1 mutation causes entry into mitosis at an abnormally small size, resulting in a shorter G2. G1 is lengthened, suggesting that progression through Start (beginning of cell cycle) is responsive to growth when the G2/M control is lost. Furthermore, cells in poor nutrient conditions grow slowly and therefore take longer to double in size and divide. Low nutrient levels also reset the growth threshold so that cell progresses through the cell cycle at a smaller size. Upon exposure to stressful conditions [heat (40 °C) or the oxidizing agent hydrogen peroxide] S. pombe cells undergo aging as measured by increased cell division time and increased probability of cell death.[20] Finally, wee1 mutant fission yeast cells are smaller than wild-type cells, but take just as long to go through the cell cycle. This is possible because small yeast cells grow slower, that is, their added total mass per unit time is smaller than that of normal cells.
A spatial gradient is thought to coordinate cell size and mitotic entry in fission yeast.[21][22][23] The Pom1 protein kinase (green) is localized to the cell cortex, with the highest concentration at the cell tips. The cell-cycle regulators Cdr2, Cdr1 and Wee1 are present in cortical nodes in the middle of the cell (blue and red dots). a, In small cells, the Pom1 gradient reaches most of the cortical nodes (blue dots). Pom1 inhibits Cdr2, preventing Cdr2 and Cdr1 from inhibiting Wee1, and allowing Wee1 to phosphorylate Cdk1, thus inactivating cyclin-dependent kinase (CDK) activity and preventing entry into mitosis. b, In long cells, the Pom1 gradient does not reach the cortical nodes (red dots), and therefore Cdr2 and Cdr1 remain active in the nodes. Cdr2 and Cdr1 inhibit Wee1, preventing phosphorylation of Cdk1 and thereby leading to activation of CDK and mitotic entry. (This simplified diagram omits several other regulators of CDK activity.)
Fission yeast switches mating type by a replication-coupled recombination event, which takes place during S phase of the cell cycle. Fission yeast uses intrinsic asymmetry of the DNA replication process to switch the mating type; it was the first system where the direction of replication was shown to be required for the change of the cell type. Studies of the mating-type switching system lead to a discovery and characterization of a site-specific replication termination site RTS1, a site-specific replication pause site MPS1, and a novel type of chromosomal imprint, marking one of the sister chromatids at the mating-type locus mat1. In addition, work on the silenced donor region has led to great advances in understanding formation and maintenance of heterochromatin.[24]
Schizosaccharomyces pombe is a facultative sexual microorganism that can undergo mating when nutrients are limiting.[25] Exposure of S. pombe to hydrogen peroxide, an agent that causes oxidative stress leading to oxidative DNA damage, strongly induces mating and formation of meiotic spores.[26] This finding suggests that meiosis, and particularly meiotic recombination, may be an adaptation for repairing DNA damage.[26] Supporting this view is the finding that single base lesions of the type dU:dG in the DNA of S. pombe stimulate meiotic recombination.[27] This recombination requires uracil-DNA glycosylase, an enzyme that removes uracil from the DNA backbone and initiates base excision repair. On the basis of this finding, it was proposed that base excision repair of either a uracil base, an abasic site, or a single-strand nick is sufficient to initiate recombination in S. pombe.[27] Other experiments with S. pombe indicated that faulty processing of DNA replication intermediates, i.e. Okazaki fragments, causes DNA damages such as single-strand nicks or gaps, and that these stimulate meiotic recombination.[28]
Fission yeast has become a notable model system to study basic principles of a cell that can be used to understand more complex organisms like mammals and in particular humans.[29][30] This single cell eukaryote is nonpathogenic and easily grown and manipulated in the lab.[31][32] Fission yeast contains one of the smallest numbers of genes of a known genome sequence for a eukaryote, and has only three chromosomes in its genome.[33] Many of the genes responsible for cell division and cellular organization in fission yeast cell are also found in the human's genome.[31][32][34] Cell cycle regulation and division are crucial for growth and development of any cell. Fission yeast's conserved genes has been heavily studied and the reason for many recent biomedical developments.[35][36] Fission yeast is also a practical model system to observe cell division because fission yeast's are cylindrically shaped single celled eukaryotes that divide and reproduce by medial fission.[31] This can easily be seen using microscopy. Fission yeast also have an extremely short generation time, 2 to 4 hours, which also makes it an easy model system to observe and grow in the laboratory[32] Fission yeast's simplicity in genomic structure yet similarities with mammalian genome, ease of ability to manipulate, and ability to be used for drug analysis is why fission yeast is making many contributions to biomedicine and cellular biology research, and a model system for genetic analysis.[32][25][30][37][38]
Schizosaccharomyces pombe is often used to study cell division and growth because of conserved genomic regions also seen in humans including: heterochromatin proteins, large origins of replication, large centromeres, conserved cellular checkpoints, telomere function, gene splicing, and many other cellular processes.[33][39][40] S. pombe's genome was fully sequenced in 2002, the sixth eukaryotic genome to be sequenced as part of the Genome Project. An estimated 4,979 genes were discovered within three chromosomes containing about 14Mb of DNA. This DNA is contained within 3 different chromosomes in the nucleus with gaps in the centromeric (40kb) and telomeric (260kb) regions.[33] After the initial sequencing of the fission yeast's genome, other previous non-sequenced regions of the genes have been sequenced. Structural and functional analysis of these gene regions can be found on large scale fission yeast databases such as PomBase.
Forty-three percent of the genes in the Genome Project were found to contain introns in 4,739 genes. Fission yeast does not have as many duplicated genes compared to budding yeast, only containing 5%, making fission yeast a great model genome to observe and gives researchers the ability to create more functional research approaches. S. pombe's having a large number of introns gives opportunities for an increase of range of protein types produced from alternative splicing and genes that code for comparable genes in human.[33] 81% of the three centromeres in fission yeast have been sequenced. The lengths of the three centromeres were found to be 34, 65, and 110 kb. This is 300–100 times longer than the centromeres of budding yeast. An extremely high level of conservation (97%) is also seen over 1,780-bp region in the DGS regions of the centromere. This elongation of centromeres and its conservative sequences makes fission yeast a practical model system to use to observe cell division and in humans because of their likeness.[33][41][42]
PomBase[8][43] reports over 69% of protein coding genes have human orthologs and over 500 of these are associated with human disease . This makes S. pombe a great system to use to study human genes and disease pathways, especially cell cycle and DNA checkpoint systems.[42][44][45][46]
Biodiversity and evolutionary study of fission yeast was carried out on 161 strains of Schizosaccharomyces pombe collected from 20 countries.[47] Modeling of the evolutionary rate showed that all strains derived from a common ancestor that has lived since ~2,300 years ago. The study also identified a set of 57 strains of fission yeast that each differed by ≥1,900 SNPs;,[47] and all detected 57 strains of fission yeast were prototrophic (able to grow on the same minimal medium as the reference strain).[47] A number of studies on S.pombe genome support the idea that the genetic diversity of fission yeast strains is slightly less than budding yeast.[47] Indeed, only limited variations of S.pombe occur in proliferation in different environments. In addition, the amount of phenotypic variation segregating in fission yeast is less than that seen, in S. cerevisiae.[48] Since most strains of fission yeast were isolated from brewed beverages, there is no ecological or historical context to this dispersal.
DNA replication in yeast has been increasingly studied by many researchers. Further understanding of DNA replication, gene expression, and conserved mechanisms in yeast can provide researchers with information on how these systems operate in mammalian cells in general and human cells in particular.[40][49][50][51] Other stages, such as cellular growth and aging, are also observed in yeast in order to understand these mechanisms in more complex systems.[34][52][53][54]
S. pombe stationary phase cells undergo chronological aging due to production of reactive oxygen species that cause DNA damages. Most such damages can ordinarily be repaired by DNA base excision repair and nucleotide excision repair.[55] Defects in these repair processes lead to reduced survival.
Cytokinesis is one of the components of cell division that is often observed in fission yeast. Well-conserved components of cytokinesis are observed in fission yeast and allow us to look at various genomic scenarios and pinpoint mutations.[45][56][57] Cytokinesis is a permanent step and very crucial to the wellbeing of the cell.[58] Contractile ring formation in particular is heavily studied by researchers using S. pombe as a model system. The contractile ring is highly conserved in both fission yeast and human cytokinesis.[45] Mutations in cytokinesis can result in many malfunctions of the cell including cell death and development of cancerous cells.[45] This is a complex process in human cell division, but in S. pombe simpler experiments can yield results that can then be applied for research in higher-order model systems such as humans.
One of the safety precautions that the cell takes to ensure precise cell division occurs is the cell-cycle checkpoint.[59][60] These checkpoints ensure any mutagens are eliminated.[61] This is done often by relay signals that stimulate ubiquitination of the targets and delay cytokinesis.[33] Without mitotic check points such as these, mutagens are created and replicated, resulting in multitudes of cellular issues including cell death or tumorigenesis seen in cancerous cells. Paul Nurse, Leland Hartwell, and Tim Hunt were awarded the Nobel Prize in Physiology or Medicine in 2001. They discovered key conserved checkpoints that are crucial for a cell to divide properly. These findings have been linked to cancer and diseased cells and are a notable finding for biomedicine.[62]
Researchers using fission yeast as a model system also look at organelle dynamics and responses and the possible correlations between yeast cells and mammalian cells.[63][64] Mitochondria diseases, and various organelle systems such as the Golgi apparatus and endoplasmic reticulum, can be further understood, by observing fission yeast's chromosome dynamics and protein expression levels and regulation.[46][50][65][66][67][68]
However, there are limitations with using fission yeast as a model system: its multidrug resistance. "The MDR response involves overexpression of two types of drug efflux pumps, the ATP-binding cassette (ABC) family... and the major facilitator superfamily".[35] Paul Nurse and some of his colleagues have recently created S. pombe strains sensitive to chemical inhibitors and common probes to see whether it is possible to use fission yeast as a model system of chemical drug research.[35]
For example, Doxorubicin, a very common chemotherapeutic antibiotic, has many adverse side-effects. Researchers are looking for ways to further understand how doxorubicin works by observing the genes linked to resistance by using fission yeast as a model system. Links between doxorubicin adverse side-effects and chromosome metabolism and membrane transport were seen. Metabolic models for drug targeting are now being used in biotechnology, and further advances are expected in the future using the fission yeast model system.[36]
Fission yeast is easily accessible, easily grown and manipulated to make mutants, and able to be maintained at either a haploid or diploid state. S. pombe is normally a haploid cell but, when put under stressful conditions, usually nitrogen deficiency, two cells will conjugate to form a diploid that later form four spores within a tetrad ascus.[32] This process is easily visible and observable under any microscope and allows us to look at meiosis in a simpler model system to see how this phenomenon operates.
Virtually any genetics experiment or technique can, therefore, be applied to this model system such as: tetrad dissection, mutagens analysis, transformations, and microscopy techniques such as FRAP and FRET. New models, such as Tug-Of-War (gTOW), are also being used to analyze yeast robustness and observe gene expression. Making knock-in and knock-out genes is fairly easy and with the fission yeast's genome being sequenced this task is very accessible and well known.[69][70]
{{cite journal}}: Cite journal requires |journal= (help) {{cite book}}: CS1 maint: multiple names: authors list (link) Schizosaccharomyces pombe, also called "fission yeast", is a species of yeast used in traditional brewing and as a model organism in molecular and cell biology. It is a unicellular eukaryote, whose cells are rod-shaped. Cells typically measure 3 to 4 micrometres in diameter and 7 to 14 micrometres in length. Its genome, which is approximately 14.1 million base pairs, is estimated to contain 4,970 protein-coding genes and at least 450 non-coding RNAs.
These cells maintain their shape by growing exclusively through the cell tips and divide by medial fission to produce two daughter cells of equal size, which makes them a powerful tool in cell cycle research.
Fission yeast was isolated in 1893 by Paul Lindner from East African millet beer. The species name pombe is the Swahili word for beer. It was first developed as an experimental model in the 1950s: by Urs Leupold for studying genetics, and by Murdoch Mitchison for studying the cell cycle.
Paul Nurse, a fission yeast researcher, successfully merged the independent schools of fission yeast genetics and cell cycle research. Together with Lee Hartwell and Tim Hunt, Nurse won the 2001 Nobel Prize in Physiology or Medicine for their work on cell cycle regulation.
The sequence of the S. pombe genome was published in 2002, by a consortium led by the Sanger Institute, becoming the sixth model eukaryotic organism whose genome has been fully sequenced. S. pombe researchers are supported by the PomBase MOD (model organism database). This has fully unlocked the power of this organism, with many genes orthologous to human genes identified - 70% to date, including many genes involved in human disease. In 2006, sub-cellular localization of almost all the proteins in S. pombe was published using green fluorescent protein as a molecular tag.
Schizosaccharomyces pombe has also become an important organism in studying the cellular responses to DNA damage and the process of DNA replication.
Approximately 160 natural strains of S. pombe have been isolated. These have been collected from a variety of locations including Europe, North and South America, and Asia. The majority of these strains have been collected from cultivated fruits such as apples and grapes, or from the various alcoholic beverages, such as Brazilian Cachaça. S. pombe is also known to be present in fermented tea, kombucha. It is not clear at present whether S. pombe is the major fermenter or a contaminant in such brews. The natural ecology of Schizosaccharomyces yeasts is not well-studied.
Schizosaccharomyces pombe o S. pombe, también llamada "fission yeast" en inglés (levadura de fisión, haciendo referencia a su mecanismo característico de división celular), es una especie de levadura. Es un hongo unicelular eucariota usado como organismo modelo en biología molecular y biología celular. Tiene forma de bastón y mide normalmente de 3 a 4 micrómetros de diámetro y de 7 a 14 micrómetros de longitud. Su genoma, aproximadamente tiene 14,1 millones de pares de bases, conteniendo 4.970 genes codificadores de proteína, y al menos 450 ARN no codificante.[1]
S. pombe se divide por fisión binaria y produce dos células de igual tamaño. Es muy usada como organismo modelo en el estudio del ciclo celular, ARNi, entre otros.
Esta levadura fue aislada por primera vez de una cerveza africana en 1893 por Lindner. Su nombre, "Pombe" significa cerveza en el idioma swahili.[2][3]
Schizosaccharomyces pombe o S. pombe, también llamada "fission yeast" en inglés (levadura de fisión, haciendo referencia a su mecanismo característico de división celular), es una especie de levadura. Es un hongo unicelular eucariota usado como organismo modelo en biología molecular y biología celular. Tiene forma de bastón y mide normalmente de 3 a 4 micrómetros de diámetro y de 7 a 14 micrómetros de longitud. Su genoma, aproximadamente tiene 14,1 millones de pares de bases, conteniendo 4.970 genes codificadores de proteína, y al menos 450 ARN no codificante.
S. pombe se divide por fisión binaria y produce dos células de igual tamaño. Es muy usada como organismo modelo en el estudio del ciclo celular, ARNi, entre otros.
Esta levadura fue aislada por primera vez de una cerveza africana en 1893 por Lindner. Su nombre, "Pombe" significa cerveza en el idioma swahili.
Schizosaccharomyces pombe, fisioaren legamia ere deitua, deskribatutako lehenengo fisio legamia da; hau da, fisio edo erdibitze bidez ugaltzen dena, bi zelula alaba berdin sortuz.
Organismo eukarioto eta unizelularra da. Legamia izanik, Ascomycota filumeko onddoa da. Hala ere, talde honetatik bereizten dituen zenbait ezaugarri ditu; hala nola, bizimodu unizelularra eta azukreak hartzitzeko gaitasuna. Morfologiari dagokionez, zilindrikoa da, kanabera formakoa eta ertz borobilduna. 7-14 µm luzetara eta 3-4 µm diametroko lodiera neurtu ohi du. [1]
S. pombe, Saccharomyces cerevisiae legamiarekin batera, biologia molekularreko eredu oso garrantzitsua da. Bere genomak, bizi-zikloak eta ugaztunen zelulekin dituen antzekotasunak, mikroorganismo hobeezina bihurtzen dute gizakien zenbait prozesu zelular aztertzeko.
S. pombe, erdibitze bidez ugaltzen den aurkitu zuten lehen legamia izan zen, hau da, lehen fisio legamia. P. Lindner-ek aurkitu zuen 1893.urtean, eta Archaeascomycete filumean sailkatu zuen, Afrikako ekialdetik etorritako “Artatxiki garagardo” izeneko garagardo berezia dastatzean. Hori dela eta, pombe du izen espezifikotzat, garagardoa Swahili hizkuntzan “pombe” izena duelarik. Bestalde Schizosaccharomyces generoa bere aurkikuntzarekin sortu zen: “-saccharomyces” legami bat dela adierazten du, eta “Schizo-”, aldiz, fisioz banatzen dela.[2]
XX. mendearen erdialdean, U. Leupold-ek S. pombe-ren azterketa genetikoa hasi zuen. Horrezkero egin diren ikerketetan, U. Leupold hazitako anduiaren eratorriak diren anduiak erabili dira; horregatik, legami honen genoma nahiko uniformea da. U. Leopold-ek hiru andui desberdin deskribatu zituen, bakoitza gurutzatze-forma desberdin batekin: lehenengoa autoernaltzeko ahalmena du, nitrogeno kontzentrazioa mugatzailea denean esporak %90-an sortzeko gai dena (h90); eta beste bi andui, bata bestearekin ugaldu daitezke: h+ eta h-. [2]
2002an, S. pombe-ren genoma osoa sekuentziatu zen, horren aurretik bakarrik bost genoma eukariotiko sekuentziatu zirelarik: Saccharomyces cerevisiae, Caenorhabditis elegans, Drosophila melanogaster, Arabidopsis thaliana eta Homo sapiens-ena. [2]
S. pombe legamia Ascomycota dibisioan sailkatuta dago, Fungi erreinuaren talde dibertsoenean. Oraindik ez dago argi espezie honek duen banaketa geografikoa, izan ere, herrialde askotan aurkitu egin da, normalean azukre kontzentrazio altuko edari alkoholetan edota melazan. Esaterako, eztian, Kombuchan, ardoan, tuban (palmondoaren izerdia hartzitzean lortutako edari alkoholikoa), eta fruitu batzuen hartzidura naturaletan (Coffea arabica eta Theobroma cacao landareen fruituetan gertatzen dena).[3]
S. pombe mikroorganismo kimioorganotrofoa da, beraz, konposatu organikoak erabiltzen ditu energia lortzeko. Oxigeno beharraren aldetik, aukerazko anaerobioak dira; hau da, ingurune aerobiko eta anaerobikoetan haz daitezke: oxigenoaren presentzian arnasketa aerobikoa burutzen dute, eta oxigenorik ez dagoenean, arnasketa anaerobioa edo hartzidura burutu dezakete. Gainera, S. pombe-k hartzidura egiteko gai da oxigenoaren presentzian, azukre kontzentrazio handia dagoenean; hain zuzen ere hartzidura alkoholikoa.[4]Aipatu beharra dago, S. pombe-k inositolarekiko auxotrofoa dela; hau da, kultibo medio batean hazteko, inusitola gaineratu behar dela legami hau haz dadin. Auxotrofoa izatearen arrazoia inositol-1-fosfato sintasa kodetzen duen genea ez izatea da. Inusitola hazkuntzarako sustantzia beharrezkoa da organismo eukariotoetan, mintz plasmatikoaren fosfolipidoen, esfingolipidoen eta glukofosfatidil-inositolaren prekurtsorea delako. [5]
Eredu bezala, S. pombe aurrerapen garrantzitsuak ekarri ditu hainbat ezagumenduetan: ziklo zelularrean eta bere erregulazioan, mikrotubuluen sintesian, desberdintzazio meiotikoan, morfogenesian, polaritatean, estresaren aurreko mekanismoetan edota genomaren kalteen erreparazioan. Bere eskuragarritasuna, kultibatzeko erreztasuna, hazkuntza tasa handia, genoma txikia eta bizimodu haploidea, ikerketan erabiltzeko mikroorganismo oso bideragarria egiten du. [2]
Laburbiltzeko, S. pombe laborategietan erabiltzeko abaintailak bost dira.
Esaterako, organuluen biogenesia, zitoeskeletoaren antolaketa, DNA erreplikazioa, transkripzioa, itzulpena eta erregulazioa.
Hala eta guztiz ere, badaude zenbait muga S. pombe erabateko eredutzat erabiltzeko: medikamentuekiko erresistentzia. Hauek ABC ponpen (transmintz proteinen talde bat, energia gastuaren bidezko garraioa bideratzen dutenak) bidez medikamentuak zelulatik kanpo botatzen dituzte, eta beraz, zenbait medikamenduen efektuak edo funtzioak ikertzea zaila da. Baina hainbat ikertzaile S. pombe andui bereziak sortu dituzte ponpatze mekanismoak baztertuz, legamia hau medikamentu kimikoen ikerketa modelo bezala erabil daitezen.[6]
S. pombe interes handiko organismoa da bere genoma kontuan hartuta; izan ere, 2002an bere genoma osoa sekuentziatu zen, Genoma Projektuan sekuentziaturiko seigarren genoma eukariotikoa izanik. Momentu horretan genoma txikiena zuen ikertutako eukariotoa izan zen. Bere genomaren 13,8 Mb-ak hiru kromosometan antolatzen dira, genoma mitokondialarekin batera. Eukarioten artean, proteina kodetzaile gene gutxien dituena da, genoman soilik 4824 gene baitauka. [2]
Gainera introi kopuru oso altua du (geneen %43 intronak dituzte, 4730 gutxi gora behera)[2], honek aukera ematen du splicing bidez aldi berean mota eta funtzio desberdineko proteina kopurua handitzeko, zelulan proteina barietatea handituz. [1] Gene errepikakorrak genoma eukariotikoaren berezitasun esanguratsua da, baina legami honetan soilik geneen %5 errepikakorrak dira, eta beraz, ikerlariak ikerketa funtzionalagoak egitera baimentzen du, bere azterketa eta sekuentziazioa askoz errazagoa baita. [7] Bestalde kromosometako zentromeroak nahiko luzeak dira, 35 eta 110kb artekoak. [2] Luzera handiko eta oso eskualde kontserbakorrak direla kontuan harturik gizakien zentromeroen antza handia dute eta bihurtzen du S. pombe oso eredu ona gizakien zatiketa zelularra behatzeko. Beste organismo eukariotikoen genomekin konparaturik, proteinak kodetzen dituzten geneen %69 gizakietan sekuentzia ortologoak dira [7]. Gainera S. pombe genoman, 289 genek erlazio oso estua dute gizakien hainbat gaixotasunekin, askok batez ere minbiziarekin. [1] Legamia eta gizakiaren arteko antzekotasun genetiko hauek interes handikoak dira biomedikuntzan. Gizakien gaixotasun genetiko askoren oinarri molekularra ulertzeko eta ikertzeko erabiltzeko aukera dago. Izan ere, oso erabilgarria izan daiteke hauen kontrako tratamenduak edo prebentzio metodoak aurkitzeko.
Hala ere, badaude beste hainbat faktore egiten dutela oraindik eredu hobeagoa: batik bat, herraminta molekularrak. Mekanismo molekularren artean, geneak mutatzeko ahalmena daukgu, fenotipoak eratuko dituzten proteinak determinatzeko eta konplemetatzeko erabili daitekeena; geneen expresioa erregulatzeko promotoreak; geneen fusioa; eta adibidez PCR bidezko delekziorako edo mutagenesirako casetteak. [1]
S. pombe-ren bizi-zikloak bi fase ditu: fase asexuala edo begetatiboa eta fase sexuala. Bere bizi-zikloan zehar, fase haploide eta diploideak tartekatzen dira, ingurumen faktoreen eta gurutzatze-forma osagarria duten zelulen arteko elkarrekintzen arabera.[8]
Fase begetatiboa, fase nagusia eta haploidea da. Honetan zehar, zatitu berri diren zelulak etengabe hazten dira luzeran, baina zabalera aldaketarik gabe. Ezaugarri honi esker, zelula baten luzera jakinda, bere adina estima daiteke. Zelulen luzera 15 μm-tara izatera heltzen denean, mitosia hasten dute. Nukleoa zatitu ondoren, zeharkako septu edo trenkada bat agertzen da zelularen erdialdean, zelula ama erdibituko duena, tamaina bereko bi zelula alaba emateko. Bi zelula alaben arteko trenkada sortzen den arren, zelulak ez dira banantzen DNA erreplikatu den arte.[8] Fase begetatiboan, 4 fase bereiz daitezke: G1 fasean DNA erreplikaziorako beharrezkoa den makinaria prestatzen du; S fasean DNA erreplikatzen da, bi kromatida ahizpez osaturiko kromosomak emateko; G2 fasean mitosia hasteko prestatzen da zelula, eta M fasean zatiketa zelular mitotikoa ematen da, DNA modu baliokidean banatzen da bi zelula alabetan. [1] Fusio legamiaren G1 fasea nahiko laburra da; mitosia eta zelulen banaketa ematen den bitartean gertatzen da. Hau da, septua dagoen bitartean G1 eta S faseak gertatzen dira, S fasea septua edo trenkadaren presentziarekin zuzenki erlazionatuta dagoelarik. Septua desagertzen denean S fasea amaitzen da eta beraz, bi zelulak guztiz bananduta daudenean, bizi-zikloaren G2 fasean sartu dira. Fase hau S. pombe-aren bizi-ziklo osoaren ¾ betetzen du, horregatik, ia zelula haploide guztiak kromosoma erreplikatuak dituzte. [1][8]
Inguruneko baldintza desegokiak direnean (esaterako, nitrogenoa mugatzailea denean) fase sexuala gertatzen da. Fase honetan, aurkako gurutzatze-forma duten bi zelula haploide elkartu eta konjugazio bidez ugaltzen dira; lehenik bi mintzak fusionatuz, eta ondoren bi nukleo parentalak fusionatuz, zigoto bat emateko. Zigoto hau diploidea da, eta meiosia pairatuko du lau nukleo haploide sortuz . Orduan, nukleo bakoitzaren inguruan espora pareta bat sortuko da (esporulazioa). Prozesu hau amaitutakoan, zelulari asko deritzo, eta bere barnean daukan nukleo bakoitzari, pareta batez inguratuta daudena, askoespora deritzo. Baldintza egokiak direnean, askosporak askatzen dira. Espora bakoitzetik zelula begetatibo haploide bat sortzen da, eta hauek fase begetatiboan sartzen dira berriro.[8] Hala nola, askoak zelula begetatibo diploideetatik ere sortu daitezke, zigoto batetik eratorri beharrean. Jatorri desberdineko bi asko horiek morfologia desberdina dute; zelula begetatibo diploideetatik eratu direnak motzagoak eta linelak dira; kontran, zigotoetatik eratorriko askoak tolestuta daude. Osatzen duen angelua normalean zelula diploideak elkartzean osatzen zuten angelua adierazten du. [8] Lehenik aipatutako zigotoen eraketa bi zelula haploideek sortzen duten zelula diploidearen gurutzatze-formaren arabera ematen da. h- eta h+ diren zelula haploideak ugaltzen direnean, h-/h+ zelula diploidea eratuko dute, meiosia eta esporulazioa egiteko gai dena. Baina gurutzatze-forma berdina duten bi zelula haploide ugaltzen direnean, zelula homozigotiko diploide bat emateko, zelula diploide hori ez da gai meiosia burutzeko. Bestalde, h90 kultiboko zelula haploideak eta h+ edo h- kultibokoak ugaltzen direnean, sortuko den zelula diploidea meiosia egiteko gai izango da baita. [1][8]
Fisio legamiaren bizi-zikloan eta geneen erregulazioan inguruko zein organismo barruko faktoreek eragiten dute. Bizi-zikloa erregulatzen duen faktore garrantzitsuen artean, zelulen tamaina daukagu. Izan ere, mitosian sartzeko, zelulek tamaina jakin bat izan behar dute, 15 μm gutxi gora behera.[8] Beste alde batetik, proteina erregulatzaileak ditugu. Azaldutako moduan, S.pombe-ren bizi-zikloaren fase nagusia, G2 fasea da. Fase honetan zelulek DNA-ren sintesian eta erreplikazioan akatsik egon den kontrolatzeko makinaria daukate. Beraz, hau ikusita eta G1 fasearen iraupen laburra kontuan hartuta, pentsa dezakegu G1 fasean ez dagoela kontrol punturik. Baina ikertu egin da G1 fasean ere bizi zikloaren kontrol puntu garrantzitsua dagoela. Bi kontrol puntu hauek funtsezko osagai komun bat dute: cdc2 genearen produktua (kinasa bat). Cdc2 genea gizakien CDK1 kinasa kodetzen duen genearen homologoa da. CDK1 proteinak honek mitosia erreegulatzen du eta beraz, S. pombe-ren cdc2 genea interes handikoa da zelulen bizi-zikloa, zatiketa eta hauen erregulazioa ikertzeko. [8]
Kaltetutako kromosoma bat erreparatzeko era eraginkor bat, kaltetutako sekuentziari dagokion informazioa beste DNA molekula batetik kopiatzea da. Metodo hau erraza da zelula diploideentzat, beti baitituzte informazio genetikoaren bi kopia. Hau ez da gertatzen ordea G1 fasean dagoen zelula haploide batean. [8]Aurreko atalean aipatu bezala, S. pombe-k fase haploide zein diploidea ditu. Hala ere, fase diploide edo sexualean bakarrik sartzen da kanpoko baldintzak ezegokiak direnean eta beraz, bizi-ziklo gehiena fase begetatibo edo haploidean igaroko du. Organismoak beraz, kromosoma homologorik ez dituenez fase gehienetan, DNA erreparaziorako beste mekanismo batzuk erabiltzen ditu. Izan ere, fase begetatiboko (haploide) G2 fasea bizi-zikloaren parte oso handia hartzen du eta bertan zelula haploide gehienak DNA erreplikatuta daukate; hau da, kromosoma bakoitzean bi kromatida ahizpa dituzte. Ezaugarri horri esker, DNAren apurketari aurre egin diezaioke, kromatida baten apurketa gertatzekotan, bestea erabili dezaketelako DNA molde moduan. [8]
Bestalde, DNA, peroxidoen aurre oso mekanismo eraginkorrak ditu. Peroxidoak DNA kaltetzen dutelarik, honen kontaktuan, S. pombe legamiak ahalik eta azkarren meiosporak eratzen ditu. Meiosiaren bidez kromatiden arteko errekonbinazio genetikoa gertatzen da, DNA-ren kontserbaketa baimenduz.
Schizosaccharomyces pombe, fisioaren legamia ere deitua, deskribatutako lehenengo fisio legamia da; hau da, fisio edo erdibitze bidez ugaltzen dena, bi zelula alaba berdin sortuz.
Organismo eukarioto eta unizelularra da. Legamia izanik, Ascomycota filumeko onddoa da. Hala ere, talde honetatik bereizten dituen zenbait ezaugarri ditu; hala nola, bizimodu unizelularra eta azukreak hartzitzeko gaitasuna. Morfologiari dagokionez, zilindrikoa da, kanabera formakoa eta ertz borobilduna. 7-14 µm luzetara eta 3-4 µm diametroko lodiera neurtu ohi du.
S. pombe, Saccharomyces cerevisiae legamiarekin batera, biologia molekularreko eredu oso garrantzitsua da. Bere genomak, bizi-zikloak eta ugaztunen zelulekin dituen antzekotasunak, mikroorganismo hobeezina bihurtzen dute gizakien zenbait prozesu zelular aztertzeko.
Schizosaccharomyces pombe appartient à la classe des champignons ascomycètes. Il s'agit d'une une espèce de levure particulière, aussi appelée "levure à fission", qui présente la particularité de se reproduire en se devisant, à partir de la cellule mère, en deux cellules filles de taille identique[1]. Elle est utilisée comme un organisme modèle en biologie cellulaire et moléculaire qui permet de comprendre des organismes plus complexes[2]. Elle est partiellement domestiquée, notamment pour faire des bières (pombe signifie bière en Swahili)[3]. Elle fut isolée en 1890 d'une bière de l'Est de l'Afrique à base de millet et décrite par Paul Lindner en 1893[4].
C'est un organisme unicellulaire eucaryote, dont les cellules sont cylindriques. Ces cellules mesurent généralement 7 à 14 micromètres de longueur et 4 de largeur. On la trouve sur les raisins et dans les mélanges de fermentation.
Son nom, contenant le préfixe schizo qui signifie "fendre" ou "couper en deux"[5], vient du fait qu'elle pratique une division par fission médiane. Deux types de cycle reproduction existent chez Schizosaccharomyces pombe[6] :
Les cellules maintiennent leur forme allongée en grandissant uniquement par leurs extrémités et se divisent par une fission médiane, produisant ainsi deux cellules filles de taille égale, ce qui fait d'elles un outil puissant pour l'étude du cycle cellulaire et sa régulation.
Sa division cellulaire est régulée par la protéine kinase pom1[8].
S. pombe possède 4 800 ORF, trois chromosomes, soit l'un des plus petits génomes recensés chez un organisme eucaryote[9], et une phase haploïde dominante dans son cycle biologique (et de façon caractéristique une phase G2 très longue). La séquence de son génome a été publiée en 2002, par un consortium dirigé par l'Institut de Sanger (c'est le sixième organisme modèle eucaryote séquencé).
Schizosaccharomyces pombe appartient à la classe des champignons ascomycètes. Il s'agit d'une une espèce de levure particulière, aussi appelée "levure à fission", qui présente la particularité de se reproduire en se devisant, à partir de la cellule mère, en deux cellules filles de taille identique. Elle est utilisée comme un organisme modèle en biologie cellulaire et moléculaire qui permet de comprendre des organismes plus complexes. Elle est partiellement domestiquée, notamment pour faire des bières (pombe signifie bière en Swahili). Elle fut isolée en 1890 d'une bière de l'Est de l'Afrique à base de millet et décrite par Paul Lindner en 1893.
Schizosaccharomyces pombe, tamén chamado "lévedo de fisión", é unha especie de lévedo utilizado na fabricación de certas cervexas tradicionais e como organismo modelo en investigacións de bioloxía molecular e bioloxía celular. É un eucariota do reino dos fungos formado por células cilíndricas con forma de bastón (parecen bacilos, pero máis grandes). As células miden xeralmente de 3 a 4 micrómetros de diámetro e de 7 a 14 micrómetros de lonxitude. O seu xenoma, que é de aproximadamente 14,1 millóns de pares de bases, estímase que contén uns 4.970 xenes codificantes de proteínas e polo menos 450 ARNs non codificantes.[1]
Estas células manteñen a súa forma ao creceren exclusivamente polos extremos da célula e dividirse por fisión binaria medial para producir dúas células fillas de igual tamaño, o que o fai un poderoso modelo para estudar o ciclo celular.
O lévedo de fisión foi illado en 1893 por Paul Lindner da cervexa de millo miúdo producida no leste de África. O nome da especie pombe é unha palabra da lingua swahili que significa cervexa. Na década de 1950 empezou a utilizarse como modelo experimental por Urs Leupold para estudos de xenética,[2][3] e por Murdoch Mitchison para estudar o ciclo celular.[4][5] O investigador do lévedo de fisión Paul Nurse conseguiu unir as dúas escolas ata entón independentes dos estudos xenéticos e do ciclo celular. Xunto con Lee Hartwell e Tim Hunt, Nurse gañou o Premio Nobel de Medicina de 2001 polos seus traballos sobre a regulación dio ciclo celular.
A secuencia do xenoma de S. pombe foi publicada en 2002 por un consorcio liderado polo Sanger Institute, converténdose no sexto modelo de organismo eucariótico cuxo xenoma foi secuenciado completamente. Isto fixo del un poderoso organismo modelo, xa que se identificaron nel moitos xenes homólogos de enfermidades humanas. En 2006, foi publicada a localización subcelular de case todas as proteínas de S. pombe utilizando a proteína fluorescente verde como etiqueta molecular.[6]
S. pombe é tamén un organismo importante para estudar as respostas celulares aos danos no ADN e o proceso da replicación do ADN.
Illáronse aproximadamente 160 cepas naturais de S. pombe. Foron recollidas en varias localizacións de diversas partes do mundo como Europa, Norte e Sudamérica, e Asia. A maioría destas cepas recolléronse de froitas cultivadas como mazás e uvas, ou de diversas bebidas alcohólicas, como por exemplo a cachaza brasileira. S. pombe tamén está presente no té fermentado kombucha.[7] Porén, non está claro se en ditas bebidas S. pombe é o principal fermentador ou é máis ben un contaminante. A ecoloxía natural dos lévedos Schizosaccharomyces aínda non está ben estudada.
S. pombe descubriuse en 1893 cando un grupo que traballaba no Laboratorio da Asociación de Cervexeiros de Alemaña examinou uns pousos sedimentados en cervexa de millo miúdo importada de África Oriental, que lle daba á cervexa un sabor ácido e desagradable. O termo schizo, significa "división" ou "fisión", e xa fora usado previamente no nome Schizosaccharomycetes. A palabra pombe elixida para nomear a especie significa cervexa en swahili, e débese á procedencia desa cervexa. As cepas estándar de S. pombe foron illadas bastante máis tarde por Urs Leupold en 1946 e 1947 a partir dun cultivo que obtivo da colección de lévedos de Delft, nos Países Baixos. Fora depositada alí por A. Osterwalder co nome S. pombe var. liquefaciens, despois de que a illara en 1924 dun viño francés (moi probablemente picado) na Estación Experimental de Vinicultura e Horticultura de Wädenswil, Suíza. O cultivo utilizado por Urs Leupold contiña (entre outras) células cos tipos de apareamento h90 (cepa 968), h- (cepa 972), e h+ (cepa 975). Despois disto, fixéronse dous grandes esforzos para illar S. pombe de froitas, néctar, ou fermentacións: un feito por Florenzano et al.[8] nos viñedos do oeste de Sicilia, e o outro realizado por Gomes et al. (2002) en catro rexións do sueste de Brasil.[9]
O lévedo de fisión S. pombe pertence ao filo Ascomycota, que comprende o grupo máis grande e diverso dos fungos. Os ascomicetos de vida libre atópanse comunmente en exsudados de árbores, raíces de plantas e no solo que as rodea, en froitos podres e maduros, e en asociación con insectos vectores que os transportan entre distintos substratos. Moitas destas asociacións son simbióticas ou saprofíticas, aínda que numerosos ascomicetos (e os seus parentes basidiomicetos) son importantes patóxenos que atacan a moitas especies de plantas, incluíndo plantas agrícolas comerciais. Entre os xéneros de lévedos ascomicetos, os lévedos de fisión Schizosaccharomyces son os únicos que depositan a-(1,3)- glucano ou pseudonixerano na parede celular xunto cos mellor coñecidos b-glucanos e presentan unha carencia virtual de quitina. As especies deste xénero tamén difiren na composición de mananos, que presentan azucres terminais de D-galactosa nas cadeas laterais dos seus mananos. S. pombe realiza a fermentación aerobia en presenza de exceso de azucre.[10] S. pombe pode degradar ácido L-málico, un dos ácidos orgánicos dominantes no viño, o que os diferencia doutras cepas de lévedos do xénero Saccharomyces.
As especies de lévedos Schizosaccharomyces pombe e Saccharomyces cerevisiae son ambas intensamente estudadas, e son importantes ferramentas na bioloxía molecular e celular. Algunhas características xenómicas e do ciclo celular que as diferencian son:
O lévedo de fisión é un fungo unicelular cun xenoma simple e unha rápida taxa de crecemento. S. pombe ten forma de bastonciños de aproximadamente 3 µm de diámetro, e crece só por prolongación dos seus extremos. Despois da mitose, ten lugar a división pola formación dun septo ou placa celular que divide a célula no seu punto medio.
Os procesos principais da reprodución celular son a duplicación dos cromosomas, que ten lugar na fase S da interfase, seguida da segregación dos cromosomas e a división do núcleo (mitose) e a división do citoplasma (citocinese), que en conxunto forman a fase M (mitótica) do ciclo celular. G1 é o período entre as fases M e S, e G2 é o período entre as fases S e M. S. pombe permanece na fase G2 do ciclo celular durante un extenso período de tempo (a diferenza de Saccharomyces cerevisiae, que permanece moito tempo na fase G1).
A mitose dos lévedos de fisión está controlada por mecanismos similares aos dos organismos multicelulares animais. Normalmente prolifera nun estado haploide. Cando ten escaseza de alimentos, as células de tipos de apareamento opostos (P e M) fusiónanse formando un cigoto diploide, que inmediatamente entra en meiose, xerando catro esporas haploides nun asco. Cando melloran as condicións no seu medio, estas esporas xerminan e producen células haploides proliferantes.[12]
Características xerais do ciclo vital.
O ciclo celular dun lévedo de fisión.
Estadios da división de Schizosaccharomyces vistos con microscopia óptica de campo escuro e brillante.

As características xerais da citocinese móstranse aquí. O sitio onde se vai producir a división celular determínase antes da anafase. O fuso da anafase (en verde na fuigura) colócase despois de modo que os cromosomas segregados estean en lados opostos do plano de clivaxe predeterminado.

No lévedo de fisión, no cal o crecemento da célula controla a progresión da fase G2 á M, unha mutación chamada wee1 causa a entrada en mitose e un tamaño anormalmente pequeno, o que orixina unha fase G2 máis curta. A G1 está prolongada, o que suxire que a progresión desde o comezo do ciclo responde ao crecemento cando se perde o control G2/M. Ademais, as células que están en condicións pobres en nutrientes crecen lentamente e lévalles máis tempo duplicar o seu tamaño e dividirse. Os niveis baixos de nutrientes tamén cambian o limiar de crecemento da célula para que a célula avance polo ciclo celular con tamaños menores. Despois dunha exposición a condicións estresantes (calor a 40 °C ou axentes oxidantes como o peróxido de hidróxeno) as células de S. pombe sofren un envellecemento que se mide polo incremento do tempo de división celular e o aumento da probabilidade de morte celular.[13] Finalmente, as células coa mutación wee1 son menores que as de tipo silvestre, pero lévalles o dobre de tempo completar o ciclo celular. Isto é posible porque as células máis pequenas crecen máis lentamente, é dicir, a súa masa sumada total por unidade de tempo é menor que a das células normais.
Crese que un gradiente espacial coordina o tamaño celular e a entrada na mitose.[14][15][16] A proteína quinase Pom1 está localizada no córtex celular, e ten a súa maior concentración nos extremos da célula. Os reguladores do ciclo celular Cdr2, Cdr1 e Wee1 están presentes en nodos corticais da parte media da célula. Nas células pequenas, o gradiente Pom1 chega á maioría dos nodos corticais. Pom1 inhibe Cdr2, impedindo que Cdr2 e Cdr1 inhiban Wee1, e permitindo que Wee1 fosforile Cdk1, o que inactiva a actividade de quinase dependente de ciclina (CDK) e impide a entrada na mitose. En células longas, o gradiente de Pom1 non chega aos nodos corticais, e, por tanto, Cdr2 e Cdr1 permanecen activas nos nodos. Cdr2 e Cdr1 inhiben Wee1, impedindo a fosforilación de Cdk1, o que leva á activación de CDK e a entrada na mitose. (Hai ademais varios outros reguladores da actividade de CDK.)
Os lévedos de fisión cambian os tipos de apareamento por un evento de recombinación acoplado á replicación, que ten lugar durante a fase S do ciclo celular. Os lévedos de fisión usan asimetría intrínseca no proceso de replicación do ADN para cambiar o tipo de apareamento; foi o primeiro sistema no que se viu que se requiría unha dirección da replicación para o cambio de tipo celular. Os estudos do sistema de cambio do tipo de apareamento levaron ao descubrimento e caracterización do sitio RTS1 de terminación da replicación específico de sitio, e a un sitio MPS1 de pausa de replicación específico de sitio MPS1, e tamén a un novo tipo de impronta cromosómica, que marca unha das cromátides irmás no locus de tipo de apareamento mat1. Ademais, os traballos sobre a rexión doante silenciada levaron a grandes avances na comprensión da formación e mantemento da heterocromatina.[17]
S. pombe é un microorganismo sexual facultativo que pode aparearse cando os nutrientes son limitados.[18] A exposición de S. pombe ao peróxido de hidróxeno, que é un axente que causa estrés oxidativo, orixina danos oxidativos no ADN, o que induce fortemente o apareamento entre os lévedos de fisión e a formación de esporas meióticas.[19] Este descubrimento suxire que a meiose, e especialmente a recombinación meiótica, poden ser unha adaptación para reparar o ADN danado.[20][21] Un descubrimento que apoia esta idea é que as lesións que afectan a unha soa base do ADN do tipo dU:dG en S. pombe estimulan a recombinación meiótica.[22] Esta recombinación require a actuación da uracilo-ADN glicosilase, un encima que quita o uracilo do ADN e inicia a reparación de excisión de base. Baseándose neste descubrimento, propúxose que a reparación de excisión de base dunha base uracilo, ou dun sitio abásico, ou dunha rotura ou amosega dunha soa febra do ADN é suficiente para iniciar a recombinación en S. pombe.[22] Outros experimentos feitos con S. pombe indicaron que o procesamento defectuoso dos intermediarios da replicación do ADN, é dicir, os fragmentos de Okazaki, causan danos no ADN, como roturas ou amosegas dunha soa febra ou ocos (gaps), e que isto estimula a recombinación meiótica.[23]
O lévedo de fisión converteuse nun notable sistema modelo para estudar procesos básicos da célula que poden utilizarse para comprender o funcionamento de organismos máis complexos como os mamíferos e especialmente o home.[24][25] Este eucariota unicelular non é patóxeno e é facilmente cultivable e manipulable no laboratorio.[26] O lévedo de fisión é un dos eucariotas secuenciados que contén un menor número de xenes no seu xenoma, e ten só tres cromosomas.[27] Moitos deses xenes que son responsables do control da división celular e da organización da célula no lévedo de fisión, teñen xenes homólogos no xenoma humano.[26][28][29] A regulación do ciclo celular e da división son cruciais para o crecemento e desenvolvemento das células. Os xenes conservados do lévedo de fisión foron moi estudados e isto influíu en moitos desenvolvementos biomédicos recentes.[30][31] O lévedo de fisión é tamén un sistema modelo práctico para observar a división celular, que se pode seguir facilmente polo microscopio.[26] Ademais, ten un tempo de xeración extremadamente curto de só 2 a 4 horas, o que facilita a súa observación e cultivo no laboratorio.[28] A simplicidade da estrutura xenómica do lévedo de fisión pero con semellanzas cos xenomas de mamíferos, a facilidade para manipulalo, e a capacidade de usalo para análises de fármacos son as principais razóns polas que se usa tanto en biomedicina, bioloxía celular e en análises xenéticas.[28][32][33][34]
S. pombe utilízase para o estudo da división e crecemento celular debido ás rexións xenómicas conservadas que tamén se encontran en humanos, como son: proteínas da heterocromatina, orixes de replicación grandes, centrómeros grandes, puntos de control celular conservados, función dos telómeros, splicing de xenes, e outros procesos celulares.[27][35][36] O xenoma de S. pombe foi completamente secuenciado en 2002, e foi o sexto xenoma eucariótico secuenciado como parte do Proxecto Xenoma. Descubríronse uns 4.979 xenes nos seus tres cromosomas, que contiñan uns 14 Mb de ADN, con ocos nas rexións centroméricas (40kb) e teloméricas (260kb).[27] Despois da secuenciación inicial do seu xenoma, secuenciáronse outras rexións previamente non secuenciadas. A análise estrutural e funcional de todas estas rexións xénicas pode atoparse en bases de datos como PomBase.[37]
O lévedo de fisión non ten tantos xenes duplicados coma o lévedo de xemación, xa que estes supoñen só o 5%, o que fai deste organismo un gran modelo xenómico para investigar. O 40% dos xenes contiñan intróns, o que é un número elevado e dá a oportunidade de incrementar o número de tipos de proteínas producidos por medio de empalme alternativo.[27] Secuenciouse tamén o 81% dos seus tres centrómeros. As lonxitudes dos tres centrómeros son 34, 65, e 110 kb. Isto é de 300 a 100 veces meirandes que os do lévedo de xemación. Tamén se observou un extraordinariamente alto nivel de conservación (97%) nunha rexión de 1.780 bp nas rexións DGS do centrómero. Este alongamento dos centrómeros e as súas secuencias conservadas fano moi axeitado para o estudo da división celular, polo seu parecido coa humana.[27][38][39]
Durante o proxecto xenómico de S. pombe atopáronse 50 xenes que codifican proteínas ou mutacións que están ligadas con doenzas humanas, e case a metade (23) están relacionados co cancro.[27] Por tanto, estase usando para estudar xenes humanos, vías metabólicas de doenzas, a división celular e sistemas de comprobacion ou puntos de control (checkpoints) do ADN.[39][40][41][42]
A replicación do ADN neste lévedo está sendo crecementemente estudada. Unha maior comprensión da replicación do ADN, expresión xénica e mecanismos conservados neste lévedo pode proporcionar información sobre eses procesos en mamíferos e no home.[36][43][44][45] Outros procesos, como o crecemento celular e o envellecemento son tamén estudados para comprender a partir deles outros sistemas máis complexos.[29][46][47][48]
A citocinese é un dos compoñentes da división celular que tamén se estuda no lévedo de fisión. Nel obsérvanse compoñentes ben conservados da citocinese que nos permiten estudar mutacións puntuais noutros escenarios xenómicos.[41][49][50][50][51] A citocinese é unha etapa crucial para a célula.[52] A formación do anel contráctil durante a citocinese está sendo moi estudada en S. pombe, xa que está moi conservado con respecto aos humanos.[41] As mutacións na citocinese poden orixinar un mal funcionamento da célula ou mesmo a morte celular e o desenvolvemento de células cancerosas.[41] Este é un proceso complexo na división das células humanas, pero é máis doado de estudar en S. pombe.
Unha das precaucións que toma a célula para asegurarse de que está tendo lugar unha división celular correcta é a existencia dunha serie de puntos de control ou checkpoints,[53][54] os cales aseguran que os mutantes serán eliminados.[55] Isto faise xeralmente por sinais que estimulan a ubiquitinación das dianas e atrasan a citocinese.[27] Sen estes puntos de control mitóticos, créanse e replícanse os mutantes, o que dá lugar a morte celular ou xeración de tumores. Paul Nurse, Leland Hartwell, e Tim Hunt recibiron o Premio Nobel de Fisioloxía e Medicina de 2001 polos seus descubrimentos de puntos de control claves conservados esenciais para que as células se dividan correctamente. Isto está ligado co cancro e células enfermas e foi moi importante en biomedicina.[56]
Tamén se estudou a dinámica de orgánulos e as respostas e posibles correlacións entre as células do lévedo e as de mamíferos.[57][58] As enfermidades mitocondriais e varios sistemas de orgánulos como o aparato de Golgi e o retículo endoplasmático poden comprenderse mellor estudando neste organismo modelo a dinámica dos cromosomas, os niveis de expresión de proteínas e a súa regulación.[42][44][59][60][60][61][62]
Malia todo, hai tamén limitacións á hora de usar o lévedo de fisión como sistema modelo, debido á súa resistencia a múltiples fármacos. A resposta de resistencia a múltiples fármacos débese entre outras razóns á sobreexpresión de bombas de efluxo de drogas.[30] Paul Nurse e colegas crearon recentemente cepas de S. pombe sensibles a inhibidores químicos e a sondas de uso común para ver se é posible utilizar o lévedo de fisión como sistema modelo na investigación de fármacos.[30]
Por exemplo a doxorrubicina, un antibiótico quimioterapéutico moi común, ten moitos efectos adversos. Estanse buscando maneiras de comprender mellor como actúa a doxorrubicina observando os xenes ligados á resistencia usando o lévedo de fisión como sistema modelo. Atopátonse ligazóns entre os efectos adversos da doxorrubicina e o metabolismo de cromosomas e transporte de membrana. Os modelos metabólicos para a dirixir os fármacos ao punto desexado (drug targeting) están utilizándose agora en biotecnoloxía, e agárdanse máis avances no futuro usando como sistema modelo S. pombe.[31]
O lévedo de fisión é facilmente accesible, fácil de cultivar e manipular para crear mutantes, e poden manterse nun estado haploide ou diploide. Normalmente é unha célula haploide, pero posto en condicións de estrés, xeralmente deficiencia de nitróxeno, dúas células conxúganse para formar unha diploide que posteriormente forma catro esporas nun asco de tétrade.[63] Este proceso é doadamente visible e observable con microscopio e permite estudar o funcionamento da meiose.
Virtualmente pode aplicarse calquera experimento xenético ou técnica pode a este sistema modelo, como as seguintes: disección de tétrades, análise de mutáxenos, transformacións, e técnicas de microscopia como FRAT e FREP. Estánse usando tamén novos modelos como Tug-Of-War (gTOW) para analizar a robustez do lévedo e observar a súa expresión xenética. É doado preparar nel experimentos de knock-in e knockout de xenes, e como o xenoma do lévedo foi secuenciado esta tarefa é moi accesible e ben coñecida.[64][65]
Schizosaccharomyces pombe, tamén chamado "lévedo de fisión", é unha especie de lévedo utilizado na fabricación de certas cervexas tradicionais e como organismo modelo en investigacións de bioloxía molecular e bioloxía celular. É un eucariota do reino dos fungos formado por células cilíndricas con forma de bastón (parecen bacilos, pero máis grandes). As células miden xeralmente de 3 a 4 micrómetros de diámetro e de 7 a 14 micrómetros de lonxitude. O seu xenoma, que é de aproximadamente 14,1 millóns de pares de bases, estímase que contén uns 4.970 xenes codificantes de proteínas e polo menos 450 ARNs non codificantes.
Estas células manteñen a súa forma ao creceren exclusivamente polos extremos da célula e dividirse por fisión binaria medial para producir dúas células fillas de igual tamaño, o que o fai un poderoso modelo para estudar o ciclo celular.
O lévedo de fisión foi illado en 1893 por Paul Lindner da cervexa de millo miúdo producida no leste de África. O nome da especie pombe é unha palabra da lingua swahili que significa cervexa. Na década de 1950 empezou a utilizarse como modelo experimental por Urs Leupold para estudos de xenética, e por Murdoch Mitchison para estudar o ciclo celular. O investigador do lévedo de fisión Paul Nurse conseguiu unir as dúas escolas ata entón independentes dos estudos xenéticos e do ciclo celular. Xunto con Lee Hartwell e Tim Hunt, Nurse gañou o Premio Nobel de Medicina de 2001 polos seus traballos sobre a regulación dio ciclo celular.
A secuencia do xenoma de S. pombe foi publicada en 2002 por un consorcio liderado polo Sanger Institute, converténdose no sexto modelo de organismo eucariótico cuxo xenoma foi secuenciado completamente. Isto fixo del un poderoso organismo modelo, xa que se identificaron nel moitos xenes homólogos de enfermidades humanas. En 2006, foi publicada a localización subcelular de case todas as proteínas de S. pombe utilizando a proteína fluorescente verde como etiqueta molecular.
S. pombe é tamén un organismo importante para estudar as respostas celulares aos danos no ADN e o proceso da replicación do ADN.
Illáronse aproximadamente 160 cepas naturais de S. pombe. Foron recollidas en varias localizacións de diversas partes do mundo como Europa, Norte e Sudamérica, e Asia. A maioría destas cepas recolléronse de froitas cultivadas como mazás e uvas, ou de diversas bebidas alcohólicas, como por exemplo a cachaza brasileira. S. pombe tamén está presente no té fermentado kombucha. Porén, non está claro se en ditas bebidas S. pombe é o principal fermentador ou é máis ben un contaminante. A ecoloxía natural dos lévedos Schizosaccharomyces aínda non está ben estudada.
Schizosaccharomyces pombe, juga disebut "khamir fisi", adalah spesies khamir yang digunakan dalam pembuatan bir tradisional dan sebagai organisme model dalam biologi molekuler dan biologi sel. S. pombe adalah eukariota uniseluler, yang selnya berbentuk batang. Sel biasanya berdiameter 3 sampai 4 mikrometer dan panjang 7 sampai 14 mikrometer. Genomnya, yang kira-kira 14,1 juta pasangan basa, diperkirakan mengandung 4.970 gen pengkode protein dan setidaknya 450 RNA bukan pengode.[1]
Sel-sel S. pombe mempertahankan bentuknya dengan tumbuh secara eksklusif melalui ujung sel dan membelah dengan fisi medial untuk menghasilkan dua sel anak dengan ukuran yang sama, yang membuat S. pombe menjadi alat yang hebat dalam penelitian siklus sel.
Khamir fisi diisolasi pada tahun 1893 oleh Paul Lindner dari bir millet Afrika Timur. Nama spesies pombe adalah bahasa Swahili untuk bir. S. pombe pertama kali dikembangkan sebagai model eksperimental pada tahun 1950an: oleh Urs Leupold untuk mempelajari genetika,[2][3] dan oleh Murdoch Mitchison untuk mempelajari siklus sel.[4][5][6]
Paul Nurse, seorang peneliti khamir fisi, berhasil menggabungkan sekolah independen genetika khamir fisi dan penelitian siklus sel. Bersama dengan Lee Hartwell dan Tim Hunt, Nurse memenangkan Penghargaan Nobel Fisiologi atau Kedokteran tahun 2001 untuk karya mereka mengenai regulasi siklus sel.
Urutan genom S. pombe diterbitkan pada tahun 2002, oleh sebuah konsorsium yang dipimpin oleh Sanger Institute, menjadi organisme model eukariotik keenam yang genomnya telah sepenuhnya diurutkan. Ini telah sepenuhnya membuka kekuatan organisme ini, dengan banyak gen yang homolog dengan gen penyakit manusia diidentifikasi. Pada tahun 2006, lokalisasi sub-seluler hampir semua protein di S. pombe diterbitkan dengan menggunakan protein fluoresen hijau sebagai label molekuler.[7]
Schizosaccharomyces pombe juga telah menjadi organisme penting dalam mempelajari respons seluler terhadap kerusakan DNA dan proses replikasi DNA.
Sekitar 160 strain alami S. pombe telah diisolasi. Strain-strain ini telah dikumpulkan dari berbagai lokasi termasuk Eropa, Amerika Utara dan Selatan, dan Asia. Mayoritas strain ini dikumpulkan dari buah-buahan yang dibudidayakan seperti apel dan anggur, atau dari berbagai minuman beralkohol, seperti Cachaça Brasil. S. pombe juga ada dalam teh fermentasi, kombucha.[8] Tidak jelas sekarang apakah S. pombe adalah pemfermentasi utama atau kontaminan di minuman tersebut. Ekologi alami khamir Schizosaccharomyces tidak diketahui dengan baik.
Schizosaccharomyces pombe, juga disebut "khamir fisi", adalah spesies khamir yang digunakan dalam pembuatan bir tradisional dan sebagai organisme model dalam biologi molekuler dan biologi sel. S. pombe adalah eukariota uniseluler, yang selnya berbentuk batang. Sel biasanya berdiameter 3 sampai 4 mikrometer dan panjang 7 sampai 14 mikrometer. Genomnya, yang kira-kira 14,1 juta pasangan basa, diperkirakan mengandung 4.970 gen pengkode protein dan setidaknya 450 RNA bukan pengode.
Sel-sel S. pombe mempertahankan bentuknya dengan tumbuh secara eksklusif melalui ujung sel dan membelah dengan fisi medial untuk menghasilkan dua sel anak dengan ukuran yang sama, yang membuat S. pombe menjadi alat yang hebat dalam penelitian siklus sel.
Khamir fisi diisolasi pada tahun 1893 oleh Paul Lindner dari bir millet Afrika Timur. Nama spesies pombe adalah bahasa Swahili untuk bir. S. pombe pertama kali dikembangkan sebagai model eksperimental pada tahun 1950an: oleh Urs Leupold untuk mempelajari genetika, dan oleh Murdoch Mitchison untuk mempelajari siklus sel.
Paul Nurse, seorang peneliti khamir fisi, berhasil menggabungkan sekolah independen genetika khamir fisi dan penelitian siklus sel. Bersama dengan Lee Hartwell dan Tim Hunt, Nurse memenangkan Penghargaan Nobel Fisiologi atau Kedokteran tahun 2001 untuk karya mereka mengenai regulasi siklus sel.
Urutan genom S. pombe diterbitkan pada tahun 2002, oleh sebuah konsorsium yang dipimpin oleh Sanger Institute, menjadi organisme model eukariotik keenam yang genomnya telah sepenuhnya diurutkan. Ini telah sepenuhnya membuka kekuatan organisme ini, dengan banyak gen yang homolog dengan gen penyakit manusia diidentifikasi. Pada tahun 2006, lokalisasi sub-seluler hampir semua protein di S. pombe diterbitkan dengan menggunakan protein fluoresen hijau sebagai label molekuler.
Schizosaccharomyces pombe juga telah menjadi organisme penting dalam mempelajari respons seluler terhadap kerusakan DNA dan proses replikasi DNA.
Sekitar 160 strain alami S. pombe telah diisolasi. Strain-strain ini telah dikumpulkan dari berbagai lokasi termasuk Eropa, Amerika Utara dan Selatan, dan Asia. Mayoritas strain ini dikumpulkan dari buah-buahan yang dibudidayakan seperti apel dan anggur, atau dari berbagai minuman beralkohol, seperti Cachaça Brasil. S. pombe juga ada dalam teh fermentasi, kombucha. Tidak jelas sekarang apakah S. pombe adalah pemfermentasi utama atau kontaminan di minuman tersebut. Ekologi alami khamir Schizosaccharomyces tidak diketahui dengan baik.
Schizosaccharomyces pombe è una specie di lievito. Si tratta di un organismo che, a differenza del più noto Saccharomyces cerevisiae, si replica per scissione. È utilizzato come organismo modello in biologia molecolare e cellulare. Si tratta di un organismo unicellulare eucariote, dalla forma allungata, e misura tipicamente tra i 2 e i 3 μm di diametro e tra i 7 e 14 di lunghezza.
Queste cellule si riproducono per scissione binaria: da una cellula se ne formano due di dimensioni identiche tra loro. Tale aspetto è di notevole importanza negli studi sul ciclo cellulare. Tali studi sono infatti fondamentali per comprendere come le cellule crescono e si differenziano: comprendendo tali meccanismi è possibile indagare con più efficacia i processi molecolari che sconvolgono il ciclo cellulare e producono proliferazioni incontrollate come nel cancro.
Questo lievito fu identificato per la prima volta da Lindner nel 1893, dalle birre di miglio dell'Africa orientale. Il nome della specie deriva infatti dal termine che, nella lingua swahili, si usa per indicare la birra (Pombe). Il primo modello sperimentale fu sviluppato negli anni cinquanta da Murdoch Mitchison, proprio per lo studio del ciclo cellulare.
Proprio l'ampio studio sulla regolazione del ciclo cellulare, attraverso i lieviti a scissione binaria come S.pombe, ha consentito a Paul Nurse, insieme a Lee Hartwell e a Tim Hunt, di vincere il Premio Nobel per la Medicina e la Fisiologia nel 2001.
La sequenza genomica di Schizosaccharomyces pombe è stata pubblicata nel 2002 da un consorzio facente capo al Sanger Institute. È stato il sesto genoma di organismo eucariote ad essere sequenziato. Il sequenziamento ha permesso di individuare numerosi geni omologhi a quelli di geni umani responsabili di malattie, aprendo nuove strade alla ricerca su malattie come il diabete e la fibrosi cistica.
S.pombe è oggi ampiamente studiato anche per far luce sui meccanismi di risposta del ciclo cellulare ai danni al DNA e ad errori di replicazione del DNA.
Dal punto di vista enologico S. pombe è un microorganismo importante: è dotato di intensa attività fermentativa, ha capacità di dare intensa fermentazione malo-alcolica, è stato proposto il suo impiego per la disacidificazione biologica dei vini.
Schizosaccharomyces pombe è una specie di lievito. Si tratta di un organismo che, a differenza del più noto Saccharomyces cerevisiae, si replica per scissione. È utilizzato come organismo modello in biologia molecolare e cellulare. Si tratta di un organismo unicellulare eucariote, dalla forma allungata, e misura tipicamente tra i 2 e i 3 μm di diametro e tra i 7 e 14 di lunghezza.
Queste cellule si riproducono per scissione binaria: da una cellula se ne formano due di dimensioni identiche tra loro. Tale aspetto è di notevole importanza negli studi sul ciclo cellulare. Tali studi sono infatti fondamentali per comprendere come le cellule crescono e si differenziano: comprendendo tali meccanismi è possibile indagare con più efficacia i processi molecolari che sconvolgono il ciclo cellulare e producono proliferazioni incontrollate come nel cancro.
Schizosaccharomyces pombe is een splijtingsgist die behoort tot de orde Schizosaccharomycetales van de ascomyceten. De gist leeft van suikerhoudende uitscheidingsstoffen van planten. De gist komt voornamelijk voor in de haploïde vorm in de G2-fase van de celcyclus.
Schizosaccharomyces pombe wordt gebruikt bij het traditioneel brouwen van bier en als modelorganisme bij de moleculaire biologie en de celbiologie.
De gistcel is staafvormig en 3 -4 µm breed en 7 - 14 µm lang. Het genoom heeft drie chromosomen en is ongeveer 14,1 miljoen basenparen groot, heeft 4970 eiwitcoderende genen en ten minste 450 nietcoderende RNA 's.[1] Er zijn ongeveer 4.970 open leesramen. Van Schizosaccharomyces pombe zijn ongeveer 160 natuurlijke stammen geïsoleerd. De meeste stammen zijn geïsoleerd uit appels, druiven en alcohol houdende dranken zoals het Braziliaanse Cachaça. De gist komt ook voor in de gefermenteerde thee komboecha.[2].
De vegetatieve cellen zijn cilindrisch, groeien aan de top en vermeerderen zich ongeslachtelijk door een enkelvoudige celdeling (binaire deling), waarbij twee even grote dochtercellen ontstaan.
Door voedselgebrek begint de geslachtelijke voortplanting met de versmelting van twee vegetatieve cellen tot een ascus. Er wordt geen ascocarp of weefsel tussen de asci gevormd. Na de versmelting van twee vegetatieve cellen volgt karyogamie en meiose. De vier celkernen delen zich nog een keer, waardoor er vier ascosporen gevormd worden.
Celdelingstadia bij vegetatieve vermeerdering van Schizosaccharomyces pombe
Schizosaccharomyces pombe werd voor het eerst geïsoleerd door Paul Lindner uit Oost-Afrikaans gierstbier. Pombe is het Swahilisch woord voor bier. In de jaren 1950 werd door Urs Leupold begonnen met experimenteel onderzoek aan de genetische achtergronden van de gist.[3][4] Murdoch Mitchison bestudeerde de celcyclus.[5][6][7]
In 2002 werd het gesequencde genoom van Schizosaccharomyces pombe gepubliceerd[8].
In 2006 werd het proteoom gepubliceerd. De open reading frames werden gelocaliseerd met behulp van het groen fluorescent proteïne[9].
Schizosaccharomyces pombe is een splijtingsgist die behoort tot de orde Schizosaccharomycetales van de ascomyceten. De gist leeft van suikerhoudende uitscheidingsstoffen van planten. De gist komt voornamelijk voor in de haploïde vorm in de G2-fase van de celcyclus.
Schizosaccharomyces pombe wordt gebruikt bij het traditioneel brouwen van bier en als modelorganisme bij de moleculaire biologie en de celbiologie.
De gistcel is staafvormig en 3 -4 µm breed en 7 - 14 µm lang. Het genoom heeft drie chromosomen en is ongeveer 14,1 miljoen basenparen groot, heeft 4970 eiwitcoderende genen en ten minste 450 nietcoderende RNA 's. Er zijn ongeveer 4.970 open leesramen. Van Schizosaccharomyces pombe zijn ongeveer 160 natuurlijke stammen geïsoleerd. De meeste stammen zijn geïsoleerd uit appels, druiven en alcohol houdende dranken zoals het Braziliaanse Cachaça. De gist komt ook voor in de gefermenteerde thee komboecha..
De vegetatieve cellen zijn cilindrisch, groeien aan de top en vermeerderen zich ongeslachtelijk door een enkelvoudige celdeling (binaire deling), waarbij twee even grote dochtercellen ontstaan.
Door voedselgebrek begint de geslachtelijke voortplanting met de versmelting van twee vegetatieve cellen tot een ascus. Er wordt geen ascocarp of weefsel tussen de asci gevormd. Na de versmelting van twee vegetatieve cellen volgt karyogamie en meiose. De vier celkernen delen zich nog een keer, waardoor er vier ascosporen gevormd worden.
Celdelingstadia bij vegetatieve vermeerdering van Schizosaccharomyces pombe
Schizosaccharomyces pombe er ein gjærsopp som økslar seg ved fisjon.
S. pombe var den sjette eukaryoten som fekk heile genomet sitt sekvensert, etter mellom anna mennesket. Sekvenseringa var på 13,8 megabasar og vart publisert i 2002. S. pombe har tre kromosom og eit mitokondriegenom på 20 kilobasar.
Schizosaccharomyces pombe er ein gjærsopp som økslar seg ved fisjon.
S. pombe var den sjette eukaryoten som fekk heile genomet sitt sekvensert, etter mellom anna mennesket. Sekvenseringa var på 13,8 megabasar og vart publisert i 2002. S. pombe har tre kromosom og eit mitokondriegenom på 20 kilobasar.
Schizosaccharomyces pombe – gatunek grzybów należący do rodziny Schizosaccharomycetaceae[1].
Pozycja w klasyfikacji według Index Fungorum: Schizosaccharomyces, Schizosaccharomycetaceae, Schizosaccharomycetales, Schizosaccharomycetidae, Schizosaccharomycetes, Taphrinomycotina, Ascomycota, Fungi[1].
Niektóre synonimy nazwy naukowej[2]:
Jest to, podobnie jak Saccharomyces cerevisiae, organizm modelowy w biologii molekularnej i komórki, komórek eukariotycznych.
Organizm ten to jednokomórkowy eukariota o komórkach wydłużonego, pałeczkowatego kształtu. Wielkość komórek waha się w granicach 2-3 mikrometrów średnicy i 7-14 mikrometrów długości.
Komórki utrzymują swój wydłużony kształt poprzez spolaryzowany wzrost oraz następujący później podział wzdłuż krótkiej osi (podobnie jak większość bakterii), co owocuje dwoma komórkami o jednakowej wielkości (w przeciwieństwie do pączkowania drożdży piekarniczych). Właściwość ta, przy jednoczesnym zachowaniu klasycznych cech organizmu eukariotycznego, czyni z S. pombe doskonałe narzędzie do badania cyklu komórkowego, mechanizmów spolaryzowanego wzrostu oraz mechanizmów naprawy DNA oraz powstawania mutacji.
S. pombe zostały wyizolowane w 1893 przez Lindera z afrykańskiego piwa warzonego z prosa. Nazwa gatunkowa wywodzi się z języka Suahili w którym pombe oznacza właśnie piwo. Jako model do badań cyklu komórkowego zostały po raz pierwszy zastosowane przez Mitchisona w latach 1950.
Badacz S. pombe Paul Nurse razem z Lee Hartwellem i Timothym Huntem w roku 2001 zostali uhonorowani Nagrodą Nobla w dziedzinie fizjologii i medycyny za ich badania nad regulacją cyklu komórkowego.
Pełna sekwencja genomu S.pombe została opublikowana w 2002 przez konsorcjum pod przewodnictwem Instytutu Sangera. Był to szósty organizm eukariotyczny, którego genom zsekwencjonowano w całości i opublikowano. Stało się to impulsem do bardzo szerokich badań z użyciem S. pombe jako analoga komórek organizmów wyższych.
Schizosaccharomyces pombe – gatunek grzybów należący do rodziny Schizosaccharomycetaceae.
Schizosaccharomyces pombe, também chamada "levedura de fissão", é uma espécie de levedura. É usada como organismo modelo em biologia molecular e celular. Trata-se de um eucariota unicelular, cujas células têm forma de bastão, medindo tipicamente entre 3 e 4 micrómetros de diâmetro e 7 a 14 micrómetros de comprimento. Estima-se que o seu genoma, com cerca de 14,1 milhões de pares de bases, contenha 4970 genes codificadores de proteínas e pelo menos 450 ARNs não-codificantes[1].
As células mantêm a sua forma crescendo exclusivamente a partir das extremidades e dividindo-se por fissão para produzirem duas células-filhas de tamanhos iguais, o que as torna uma ferramenta poderosa no estudo do ciclo celular.
Foi isolada pela primeira vez em 1893 por Paul Lindner a partir da cerveja de milhete da África Oriental. O nome da espécie deriva da palavra suaíli para cerveja, pombe. Foi desenvolvida como modelo experimental na década de 1950 por Urs Leupold, para o estudo da genética[2][3], e por Murdoch Mitchison para o estudo do ciclo celular[4][5].
O investigador da levedura de fissão Paul Nurse fundiu com sucesso as escolas independentes da genética da levedura de fissão e da investigação do ciclo celular. Juntamente com Lee Hartwell e Tim Hunt, Nurse ganhou o Prémio Nobel de Fisiologia ou Medicina pelo seu trabalho sobre a regulação do ciclo celular.
A sequência do genoma de S. pombe foi publicada em 2002, por um consórcio liderado pelo Wellcome Trust Sanger Institute, tornando-se o sexto organismo modelo eucariota cujo genoma foi totalmente sequenciado.
Em 2006, foi publicada a localização de todas as proteínas em S. pombe, feita utilizando proteína verde fluorescente como marcador molecular.
Schizosaccharomyces pombe, também chamada "levedura de fissão", é uma espécie de levedura. É usada como organismo modelo em biologia molecular e celular. Trata-se de um eucariota unicelular, cujas células têm forma de bastão, medindo tipicamente entre 3 e 4 micrómetros de diâmetro e 7 a 14 micrómetros de comprimento. Estima-se que o seu genoma, com cerca de 14,1 milhões de pares de bases, contenha 4970 genes codificadores de proteínas e pelo menos 450 ARNs não-codificantes.
As células mantêm a sua forma crescendo exclusivamente a partir das extremidades e dividindo-se por fissão para produzirem duas células-filhas de tamanhos iguais, o que as torna uma ferramenta poderosa no estudo do ciclo celular.
Foi isolada pela primeira vez em 1893 por Paul Lindner a partir da cerveja de milhete da África Oriental. O nome da espécie deriva da palavra suaíli para cerveja, pombe. Foi desenvolvida como modelo experimental na década de 1950 por Urs Leupold, para o estudo da genética, e por Murdoch Mitchison para o estudo do ciclo celular.
O investigador da levedura de fissão Paul Nurse fundiu com sucesso as escolas independentes da genética da levedura de fissão e da investigação do ciclo celular. Juntamente com Lee Hartwell e Tim Hunt, Nurse ganhou o Prémio Nobel de Fisiologia ou Medicina pelo seu trabalho sobre a regulação do ciclo celular.
A sequência do genoma de S. pombe foi publicada em 2002, por um consórcio liderado pelo Wellcome Trust Sanger Institute, tornando-se o sexto organismo modelo eucariota cujo genoma foi totalmente sequenciado.
Em 2006, foi publicada a localização de todas as proteínas em S. pombe, feita utilizando proteína verde fluorescente como marcador molecular.
Este artigo foi inicialmente traduzido, total ou parcialmente, do artigo da Wikipédia em inglês, cujo título é «Schizosaccharomyces pombe», especificamente .Schizosaccharomyces pombe je grzib[2], co go ôpisoł Lindner 1893. Schizosaccharomyces pombe nŏleży do zorty Schizosaccharomyces i familije Schizosaccharomycetaceae.[3][4] Żŏdne podgatōnki niy sōm wymianowane we Catalogue of Life.[3]



Schizosaccharomyces pombe je grzib, co go ôpisoł Lindner 1893. Schizosaccharomyces pombe nŏleży do zorty Schizosaccharomyces i familije Schizosaccharomycetaceae. Żŏdne podgatōnki niy sōm wymianowane we Catalogue of Life.
Schizosaccharomyces pombe, veya "fisyon mayası", tek hücreli bir maya türüdür. Biyologlar tarafından ökaryotik canlıların moleküler biyoloji ve hücre biyolojisini çalışmak için bir model organizma olarak kullanılır. Hücreler çubuk şeklinde, 2-3 mikrometre çapında, 7-14 mikrometre uzunluğundadır. Bu hücreler hücre uçlarında uzayarak şekillerini korurlar, hücre ortasından bölünürek eşit büyüklükte iki yavru hücre oluştururlar.
S. pombe 1893'te Paul Lindner tarafından Doğu Afrika'da darı birasından izole edilmiştir. Pombe ismi Svahili dilinde bira demektir. 1950'li yıllarda Murdoch Mitchison tarafından ilk defa hücre bölünmesini araştırmak için deneysel bir organizma olarak kullanılmıştır.
Fisyon mayası araştırmacılarından Paul Nurse, (başka organizmalarda çalışmış Lee Hartwell ve Tim Hunt ile beraber) hücre döngüsü üzerindeki çalışmalarından dolayı 2001 yılında Nobel Fizyoloji veya Tıp Ödülünü almıştır.
S.pombe'nin genom dizini Sanger Institute önderliğindeki bir konsorsiyum tarafından yayımlanmış, genomu tamamen okunmuş altıncı model organizma olmuştur.
Hücre bölünmesi insan dahil diğer ökaryotlarda da benzer mekanizmalarla gerçekleştiğinden bu süreçle ilgili S. pombe'de yapılan keşifler insana kolaylıkla uygulanabilmektedir. Kanser, kontrolsuz hücre bölünmesi olduğu için bu maya türünden elde edilen bilgilerin pek çok tıbbi uygulaması olmuştur. Ayrıca, genom dizinine bakıldığı zaman insanda diyabet ve sistik fibroz gibi hastalıklarda bozukluğu gösterilmiş genlerin S. pombe'de de olduğu bulunduğundan bu mayanın başka alanlarda da bilime önemli katkıları olacaktır.
Schizosaccharomyces pombe, veya "fisyon mayası", tek hücreli bir maya türüdür. Biyologlar tarafından ökaryotik canlıların moleküler biyoloji ve hücre biyolojisini çalışmak için bir model organizma olarak kullanılır. Hücreler çubuk şeklinde, 2-3 mikrometre çapında, 7-14 mikrometre uzunluğundadır. Bu hücreler hücre uçlarında uzayarak şekillerini korurlar, hücre ortasından bölünürek eşit büyüklükte iki yavru hücre oluştururlar.
S. pombe 1893'te Paul Lindner tarafından Doğu Afrika'da darı birasından izole edilmiştir. Pombe ismi Svahili dilinde bira demektir. 1950'li yıllarda Murdoch Mitchison tarafından ilk defa hücre bölünmesini araştırmak için deneysel bir organizma olarak kullanılmıştır.
Fisyon mayası araştırmacılarından Paul Nurse, (başka organizmalarda çalışmış Lee Hartwell ve Tim Hunt ile beraber) hücre döngüsü üzerindeki çalışmalarından dolayı 2001 yılında Nobel Fizyoloji veya Tıp Ödülünü almıştır.
S.pombe'nin genom dizini Sanger Institute önderliğindeki bir konsorsiyum tarafından yayımlanmış, genomu tamamen okunmuş altıncı model organizma olmuştur.
Ekmek mayası (Saccharomyces cerevisiae) ile karşılaştırılması S. cerevisiae nın 5600 açık okuma çerçevesi, Sch. pombe nin 4970 açık okuma çerçevesi vardır. Gen sayılarının benzer olmasına rağmen, S. cerevisiae 250 intron bulundururken Sch. pombe'de bu sayı yaklaşık 5000'dir. S. cerevisiae nin 16 kromozomu, Sch. pombe nin 3 kromozomu vardır. S. cerevisiae genelde diploiddir Sch. pombe ise genelde haploiddir. S. cerevisiae hücre döngüsünde başlıca G1 evresindedir (dolayısıyla G1-S geçişi sıkı kontrol altındadır) , Sch. pombe ise genelde G2 evresindedir (dolayısıyla G2-M geçişi sıkı kontrol altındadır)Schizosaccharomyces pombe — одноклітинний еукаріот, паличкоподібної форми. За розміром має приблизно від 2 до 3 мікрон в діаметрі і від 7 до 14 мікрон в довжину. S. pombe зазвичай зустрічається при спиртному бродіння цукру в субтропічних регіонах[1].
Організм був знайдений та описаний в 1893 році, коли група Лабораторії Асоціації пивоварів в Німеччині досліджувала осад від виготовлення просяного пива, імпортованого із Східної Африки, що мало сумнівний кислий смак[2]. П. Лінднер був першим, хто описав Schizosaccharomyces pombe. За назву він вибрав слово «пиво» мовою суахілі, pombe. Цей організм був ідентифікований як дріжджі, і став відомим як дріжджі, що діляться, тому що це він відтворюється за допомогою симетричного поділу на відміну від брунькування у більшості інших дріжджів, зокрема пивних дріжджів Saccharomyces cerevisiae. Назва Schizosaccharomyces, від лат. Schizo- — «інший», «різний», вже використовувалося для опису інших видів, що діляться симетрично[2].
Дослідження цього організму, включаючи секвенування генома, були важливими через те, що S. pombe — простий одноклітинний організм, який, проте, розділяє багато спільних рис з клітинами складніших еукаріотів[3]. Дослідники ідентифікували багато генів S. pombe, пов'язаних з хворобами людини, зокрема кістозний фіброз, спадкову глухоту і діабет[3]. Найбільша група генів, пов'язана із хворобами людини — 23 гена, залучені в утворення раку. Ці гени беруть участь в ремонті ДНК і регуляції клітинниго циклу — всі ці процеси впливають на підтримку стабільності генома[3]. Ці відкриття дуже важливі, тому що дозволяють дослідникам більше дізнатися про еволюцію як одноклітинних, так багатоклітинних еукаріотичних організмів.
На підтвердження важливості досліджень цього організму свідчить той факт, що Пол Нерс, отримав в 2001 році Нобелівську премію або фізіології і медицини (разом із Лі Гартвеллом і Тімом Гантом) за дослідження регулювання клітинного циклу та відкриття циклін-залежних кіназ, працюючи саме з S. pombe.[4]
S. pombe був шостим еукаріотичним організмом, чий геном був повністю секвенований. S. pombe має дуже малий і компактний для еукаріотів геном, що містить 13,8 мін. пар основ, 4824 гени, та розділений на 3 хромосоми: I (5,7 Mbp), II (4,6 Mbp) і III (3,5 Mpb). Це найменше число кодуючих білки генів серед усіх відомих еукаріотів[3]. Всі 3 хромосоми — лінійні, але кругові хромосоми спостерігаються в деяких мутантах. Це відбувається, якщо S. pombe втрачає прості повтори на самих кінцях хромосом або субтеломені елементи, у цьому випадку хромосоми зливаються своїми кінцями, формуючі круглі хромосоми[5]. Три центромери займають 35, 65 і 110 kbp відповідно на кожній хромосомі, залишаючи 12,5 унікальної послідовності[3].
4730 інтронів розподілені серед 43 % генів, максимум 15 інтронів в межах єдиного гена, 614 мають два або більше інтронів. Дослідження інтронів S. pombe важливі для розуміння їх виникнення та розповсюдження. Велике число інтронів забезпечує можливості для альтернативного сплайсінга, що дає кілька варіантів тих же самих білків, що важливо як у регуляторних шляхах, так і забезпечує збільшення кількості типів білків у клітині.
Відома цінність S. pombe для дослідження функцій генів, пов'язаних з хворобами людини. Гени цього організму кодують білки, що виробляють продукти схожі до 289 відомих білків, мутації або вилучення яких приводять до хвороб людини[3]. 172 білків S. pombe мають схожість цими білками людини. Серед них 23 гени залучені у виникнення раку. Інші — залучені в хвороби обміну речовин (12 генів), неврологічі (13 генів), серцеві хвороби (1 ген) і хвороби нирок (1 ген)[3].
S. pombe — хемоорганотроф, тобто використовує органічні сполуки як джерело енергії і не вимагає світла для росту. Ці організми — факультативні аероби, тобто можуть рости як за аеробними, так і за анаеробними умовами, крім того, фони проявляють бродіння за аеробними умовами за наявністю додаткового цукру[6]. Спиртова гідрогеназа (ADH) каталізує редукування ацетальдегіду до етанолу на останньому кроці спиртного бродіння. Ця реакція пов'язана з окисленням NADH і забезпечує NAD+ для окислення гліцеральдегід-3-фосфата в процесі глуколізу. Тому, виробництво етанолу важливе для підтримання окислювально-відновлювального балансу в цитоплазмі.
S. pombe є природним ауксотрофом інозитолу. Інозитол важливий для росту всіх клітин еукаріотів, тому що він використовується для синтезу важливих мембранних фосфоліпідів, фсіногліпіду і глікозилфосфатиділінозитолу. Ці речовини грають важливу роль в шляхах передічі сигналів. Вважається, що S. pombe як ауксотроф інозитолу, тому що його природне оточення зазвичай містить велику кількість фітієвої кислоти, яка може використовуватися як джерело інозитолу за певними умовами.
S. pombe знаходять переважно в алкогольних напоях, і таким чином він не крає важливою ролі у природі. Проте, це — один з видів, що грають важливу роль в бродінні чайного гриба (Kombucha). Чайний гриб — це традиційний метод бродіння підсолоджуваного чаю, що залучає симбіоз дріжджів і оцтовокислих бактерій. Вивчення екології дріжджів в бродінні чайного грибу показує, що S. pombe легко вступає в симбіоз з іншими видами дріжджів (наприклад B. bruxellensis, C. stellata, T. delbrueckii і Z. bailii). S. pombe і інші дріжджі перероблюють цукор в чай на етанол, а етанол потім окисляють оцтовокислі бактерії, виробляючи оцтову кислоту. S. pombe важливий у суспільстві через його здатність терпіти високі концентрації цукру. Ізольовані S. pombe, T. delbrueckki і Z. bailii витримують концентрацію клюкози до 60 % і зазвичай використовуються в бродінні солодких і шампанських вин. Із течінням бродіння, види з низькою кислотною толерантністю поступово відмирають, S. pombe, з помірною толерантністю до кислих умов, через 10 день. Загалом, осмотолерантний S. pombe та інші починають бродіння чайного гриба, але процес закінчується іншими, толерантноми до кислоти видами[7].
MicrobeWiki: Schizosaccharomyces pombe
Основні модельні організми в генетиці Фаг лямбда | Кишкова паличка | Хламідомонада | Tetrahymena | Дріжджі (Пивні дріжджі • Schizosaccharomyces pombe) | Neurospora | Кукурудза | Arabidopsis | C. elegans | Дрозофіла | Даніо-реріо | Пацюк сірий | Миша хатняSchizosaccharomyces pombe Lindner, 1893
Schizosaccharomyces pombe (лат.) — вид дрожжей. Одноклеточный представитель царства грибов, относится к классу Schizosaccharomycetes (делящиеся дрожжи). Клетки имеют палочковидную форму и размеры 3—4 мкм в диаметре и 7—14 мкм в длину. Клетки растут только вдоль свой длинной оси и делятся пополам, сохраняя таким образом свою форму. Используется как модельный организм в молекулярной биологии и цитологии.
S. pombe были выделены в чистую культуру в 1893 году Линднером из восточноафриканской бузы. Название происходит от слова «пиво» на языке Суахили — Pombe. Впервые были использованы как модельный организм для изучения клеточного цикла Мердоком Мичизоном в 1950-х. Пол Нерс, Леланд Хартвелл и Тимоти Хант получили в 2001 году Нобелевскую премию по физиологии и медицине за работы по регуляции клеточного цикла, выполненную на S. pombe.
Последовательность нуклеотидов в геноме S. pombe была расшифрована в 2002 году консорциумом во главе с институтом Sanger. S. pombe стали шестым эукариотическим организмом, геном которого был полностью секвенирован. Это позволило идентифицировать многие гомологичные человеческим гены, ответственные за болезни (включая диабет и муковисцидоз). В 2006 году при помощи зелёного флуоресцентного белка как молекулярной метки была установлена внутриклеточная локализация всех белков в S. pombe. На S. pombe также ведётся изучение изменений клеточного цикла, связанных с повреждением ДНК.
Schizosaccharomyces pombe (лат.) — вид дрожжей. Одноклеточный представитель царства грибов, относится к классу Schizosaccharomycetes (делящиеся дрожжи). Клетки имеют палочковидную форму и размеры 3—4 мкм в диаметре и 7—14 мкм в длину. Клетки растут только вдоль свой длинной оси и делятся пополам, сохраняя таким образом свою форму. Используется как модельный организм в молекулярной биологии и цитологии.
S. pombe были выделены в чистую культуру в 1893 году Линднером из восточноафриканской бузы. Название происходит от слова «пиво» на языке Суахили — Pombe. Впервые были использованы как модельный организм для изучения клеточного цикла Мердоком Мичизоном в 1950-х. Пол Нерс, Леланд Хартвелл и Тимоти Хант получили в 2001 году Нобелевскую премию по физиологии и медицине за работы по регуляции клеточного цикла, выполненную на S. pombe.
Последовательность нуклеотидов в геноме S. pombe была расшифрована в 2002 году консорциумом во главе с институтом Sanger. S. pombe стали шестым эукариотическим организмом, геном которого был полностью секвенирован. Это позволило идентифицировать многие гомологичные человеческим гены, ответственные за болезни (включая диабет и муковисцидоз). В 2006 году при помощи зелёного флуоресцентного белка как молекулярной метки была установлена внутриклеточная локализация всех белков в S. pombe. На S. pombe также ведётся изучение изменений клеточного цикла, связанных с повреждением ДНК.

分裂酵母 (ぶんれつこうぼ) は子嚢菌に属する二分裂によって増殖する酵母の総称。分類学上は、シゾサッカロミケス属(Schizosaccharomyces)に対応し、本属のみをもってシゾサッカロミケス科、シゾサッカロミケス目、シゾサッカロミケス綱を構成する。学名のうちSchizo-が「分裂」を意味する。
出芽酵母とは3から4億年以上前に分化したと考えられ、これは分裂酵母と動物との違いに比べられるほど大きな差と言われる。
2010年時点で4種から構成されている。
1893年に P. Lindner によって東アフリカのミレットビール(雑穀を原料とした醸造酒)から単離された最初の分裂酵母。pombe はスワヒリ語でシコクビエなどの雑穀を用いて作られた伝統的なビールの意。 通常一倍体で生活し環境条件等によって接合、減数分裂、胞子形成を行う。分裂の周期は約3時間。 一倍体は長さ13 μm、直径3 μm程度、二倍体はそれぞれ長さ22μm、直径4 μm程度の筒状の形をし、その中央に核を持つ。DNA染色を行った細胞では偏った核小体のため核が三日月型に観察できることがあり、これは間期を区別する特徴として使われる。三本の染色体を持ち、第一染色体は5.7 Mb、第二染色体は4.6 Mb、第三染色体は3.5 Mbで、全体で13.8 Mbのゲノムを持つ。 2002年真核生物としては6番目になるゲノム配列決定が行われた。4割の遺伝子がイントロンを持つことや、タンパク質のアミノ酸配列のヒトとの類似度も出芽酵母より高いことなどから、出芽酵母よりヒトに近いモデル生物とされることもある。また、この生物を材料として細胞周期の理解が大きく前進した。
分裂酵母 (ぶんれつこうぼ) は子嚢菌に属する二分裂によって増殖する酵母の総称。分類学上は、シゾサッカロミケス属(Schizosaccharomyces)に対応し、本属のみをもってシゾサッカロミケス科、シゾサッカロミケス目、シゾサッカロミケス綱を構成する。学名のうちSchizo-が「分裂」を意味する。








