noms dans le fil d’Ariane

Marine Arthropods: A Successful Design Video and Lesson Plans
More Resources About Marine Arthropods
Terrestrial Arthropods: The Conquerors Video and Lesson Plans
More Resources About Terrestrial Arthropods
About Shape of Life
Shape of Life is a series of FREE classroom videos based on an original PBS Series. Explore the beautiful evolution of the animal kingdom on planet earth. The series is NGSS aligned with exquisite focus on diversity, biodiversity, adaptability, body structure, design, behaviors, and the innovative scientists who explore these creatures.
Die Geleedpotiges of artropodes (filum of superfilum Arthropoda) is 'n groot groep ongewerweldes (diere sonder 'n ruggraat).[1] Diere soos insekte, spinagtiges (spinnekoppe), skaaldiere (krappe en krewe), garnale en veelpotiges (duisendpote) is almal geleedpotiges.[1] Geleedpotiges het gelede pote, 'n gesegmenteerde liggaam en 'n eksoskelet (harde dop om die liggaam).[1]
Geleedpotiges het verreweg die grootste aantal spesies van enige groep diere,[1] en moontlik ook van enige filum van enige lewende organisme op aarde. 'n Enkele orde van insekte – die kewers – oortref byvoorbeeld al die ander in die aantal spesies.
Sommige geleedpotiges, veral die insekte, lewe op land. Hulle harde eksoskelette beskerm hulle teen uitdroging. Hulle is ook die eerste filum wat ware vlug bemeester het.
Die skaaldiere (Crustacea) is ook 'n klas van die Arthropoda, maar lewe meestal in die water of is van klam toestande afhanklik vir oorlewing.
Geleedpotiges is 'n bron van voedsel vir baie diere, insluitend die mens.
Die Geleedpotiges of artropodes (filum of superfilum Arthropoda) is 'n groot groep ongewerweldes (diere sonder 'n ruggraat). Diere soos insekte, spinagtiges (spinnekoppe), skaaldiere (krappe en krewe), garnale en veelpotiges (duisendpote) is almal geleedpotiges. Geleedpotiges het gelede pote, 'n gesegmenteerde liggaam en 'n eksoskelet (harde dop om die liggaam).
Geleedpotiges het verreweg die grootste aantal spesies van enige groep diere, en moontlik ook van enige filum van enige lewende organisme op aarde. 'n Enkele orde van insekte – die kewers – oortref byvoorbeeld al die ander in die aantal spesies.
Sommige geleedpotiges, veral die insekte, lewe op land. Hulle harde eksoskelette beskerm hulle teen uitdroging. Hulle is ook die eerste filum wat ware vlug bemeester het.
Die skaaldiere (Crustacea) is ook 'n klas van die Arthropoda, maar lewe meestal in die water of is van klam toestande afhanklik vir oorlewing.
Geleedpotiges is 'n bron van voedsel vir baie diere, insluitend die mens.
Los artrópodos (Arthropoda, del griegu ἄρθρον, árthron, «articulación» y πούς, poús, «pie») constitúin el filu más numberosu y diversu del reinu animal (Animalia). El términu inclúi a animales invertebraos dotaos d'una cadarma esterna y apéndices articulaos, inclúi, ente otros, inseutos, arañes, crustáceos y miriápodos.
Hai más de 1.200.000 especies descrites, na so mayoría inseutos (un millón), que representen aproximadamente'l 80% de toles especies animales conocíes. Dellos grupos d'artrópodos tan perfectamente afechos a la vida nel aire, igual que los vertebraos amniotes, a diferencia de tolos demás filos d'animales, que son acuáticos o riquen ambientes húmedos. La so anatomía, la so fisioloxía y el so comportamientu revelen un diseñu simple pero almirablemente eficaz.
Pue ser que los primeros artrópodos fueren similares a los sos presuntos antecesores, los anélidos. El so cuerpu yera llargu y blandiu y tenía munchos segmentos, toos ellos bien similares y forníos con un par de pates. Darréu la superficie del cuerpu endureciose hasta formar una cadarma esterna (exoesqueletu) o cutícula que contién quitina, proteínes, lípidos y sales de calciu.
Los artrópodos (Arthropoda, del griegu ἄρθρον, árthron, «articulación» y πούς, poús, «pie») constitúin el filu más numberosu y diversu del reinu animal (Animalia). El términu inclúi a animales invertebraos dotaos d'una cadarma esterna y apéndices articulaos, inclúi, ente otros, inseutos, arañes, crustáceos y miriápodos.
Hai más de 1.200.000 especies descrites, na so mayoría inseutos (un millón), que representen aproximadamente'l 80% de toles especies animales conocíes. Dellos grupos d'artrópodos tan perfectamente afechos a la vida nel aire, igual que los vertebraos amniotes, a diferencia de tolos demás filos d'animales, que son acuáticos o riquen ambientes húmedos. La so anatomía, la so fisioloxía y el so comportamientu revelen un diseñu simple pero almirablemente eficaz.
Buğumayaqlılar (lat. Arthropoda) - İlkağızlılar (lat. Protostomia) bölməsinə aid heyvan tipidir.
Buğumayaqlılar tipinin nümayəndələri istər növlərinin çoxluğuna, istərsə də müxtəlifliyinə görə daha çox diqqəti daha çox cəlb edir. Yaşayış mümkün olan elə bir yer yoxdur ki, orada buğumayaqlılara rast gəlinməsin. Tipin 1500000-dən artıq növü məlumdur. Onlar dəniz və okeanlarda, şirin su hövzələrində, quru mühitin müxtəlif sahələrində geniş yayılıblar. Bir çox növləri parazit həyata uyğunlaşmışdır. Parazitliklə əlaqədar olaraq, bəzi növlər öz formalarını elə dəyişmişdir ki, onları buğumayaqlılara oxşatmaq elə də asan olmur.
Buğumayaqlıların bədəninin üzəri xitinləşmiş kutikula ilə örtülüdür. Kutikula dəri epitelisi və ya hipodermadan əmələ gəlir. Tipin ayrı-ayrı nümayəndələrində kutikula qalın və ya nazik örtük şəklində olur. Kutikula qatının tərkibində xitindən başqa, digər üzvi və qeyri-üzvi maddələr də vardır. Ali xərçənglərin və bir çox hörümçəkkimilərin kutikulasında müəyyən miqdarda kalsium-karbonat vardır. Xitinli kutikula elastikdir. Buğumayaqlılarda böyümə qabıqdəyişməklə tamamlanır. Bu zaman bədən örtüyü soyulur, hipoderma daha yumşaq və zərif kutikula əmələ gətirir. Sonra köhnə örtük atılır və yeni kutikula qatı bərkiyəndən sonra heyvanın böyüməsi növbəti qabıqdəyişməyə kimi dayanır. Buğumayaqlılar, adətən inkişaf zamanı daha çox qabıq dəyişirlər, yetkin heyvanlar isə çox nadir hallarda qabıq dəyişir, ya da tamamilə dəyişmirlər. Buğumayaqlıların bədəni buğumludur. Buğumlar heteronomdur. Daha sadə quruluşlu formalarda homonom buğumluluq da müşahidə olunur. Bədəndə olan bir qrup buğumlar birləşərək ayrı-ayrı hissələri əmələ gətirmişdir. Buğumayaqlılarda bir-birindən fərqlənən üç hissə nəzərə çarpır: baş, döş və qarıncıq Başda buğumların sayı sabitdir. Döş və qarıncıqda olan buğumlarının sayı ayrı-ayrı nümayəndələrində müxtəlifdir. Buğumayaqlıların bədən səthini xitinləşmiş kutikula örtür. O, bəzi heyvanlarda, məsələn, çay xərçənglərində və yengəclərdə qalın olub, zireh əmələ gətirir, ağcaqanadlarda, milçəklərdə və başqa cücülərdə isə nazikdir.
Buğumayaqlılarda dəri-əzələ kisəsi inkişaf etməmişdir. Bununla əlaqədar olaraq, onlarda eninəzolaqlı əzələlər yaxşı inkişaf etmişdir. Bu əzələlər mürəkkəb quruluşlu olub, hərəkət zamanı daha tez yığılmaq qabiliyyətinə malikdirlər. Buğumayaqlılar bədən boşluğuna malikdirlər. Bədən boşluğu qarışıq bədən boşluğu (miksosel) adlanır.
Həzm sistemi üç hissədən-ön, orta və arxa başırsaqdan ibarətdir. Buğumayaqlılar olduqca müxtəlif qida ilə qidalanmağa uyğunlaşmışlar. Bununla əlaqədar olaraq, onlarda həzm sistemində müəyyən dərəcədə dəyişikliklər baş vermişdir. Bağırsaqda həzmə xidmət edən vəzilər də vardır.
Tənəffüs orqanları müxtəlifdir. Suda yaşayan nümayəndələri qəlsəmələrlə, quruda yaşayan nümayəndələri isə ağciyər va traxeyalar vasitəsilə tənəffüs edir. İfrazat funksiyasını Malpigi boruları və bəzi vəzilər (koksal, yaşıl vəzilər) yerinə yetirir.
Qan-damar sistemi açıqdır. Qan boşluq mayesi ilə qarışdığına görə, buğumayaqlıların qarışıq bədən boşluğunda hemolimfa dövr edir. Bel qan damarının nəbzli hissəsi ürək adlanır. Ürək boruşəkillidir və yanlarda yerləşən ostiy adlanan dəliklərə malikdir. Qan ürəyə bu dəliklərdən daxil olur. Ürəkdən çıxan qan, damarlarla axaraq bədən boşluğuna tökülür.
Sinir sistemi baş beyin adlanan cüt udlaqüstü sinir düyünündən, udlaqətrafı sinir həlqəsindən və qarın sinir zəncirindən ibarətdir. Buğumayaqlıların bir qrupunda baş-beyin nahiyə daha yüksək quruluşlu olmaqla, bir çox mürəkkəb davranışların yerinə yetirilməsində mühüm rol oynayır. Buğumayaqlıların hiss orqanları da yüksək quruluşludur. Müxtəlif formalarında mürəkkəb fasetli göz, toxunma və kimyəvi hiss orqanları, eşitmə orqanları və s. inkişaf etmişdir.
Müxtəlif cinsli heyvanlardır, əksəriyyəti cinsi yolla çoxalır[1]. İnkişafları ya mürəkkəb çevrilmə ilə (metamorfozla), ya da düzünədir.
Buğumayaqlılar tipi 6 yarımtipə ayrılır:
Buğumayaqlılar (lat. Arthropoda) - İlkağızlılar (lat. Protostomia) bölməsinə aid heyvan tipidir.
An artropoded eo al loened a ya d'ober ar skourrad Arthropoda. Loened divellkeinek ez int, da lavaret eo n'o deus mell-kein ebet. Emañ an amprevaned, ar c'hevnid, ar c'hresteneged, an derederezed e-touez an atropoded, da skouer. An trilobited, loened aet da get bremañ, a oa artropoded ivez.
a vo kavet e Wikimedia Commons.
Els artròpodes (Arthropoda) són invertebrats dotats d'un exoesquelet (esquelet extern), un cos segmentat, i extremitats articulades anomenades apèndixs.[1] Els artròpodes inclouen els insectes, els aràcnids, els crustacis els miriàpodes i altres. La paraula prové del grec ἄρθρον, «articulació», i de la paraula πούς «peu» (genitiu ποδός).[2]
Els artròpodes es caracteritzen per les seves extremitats articulades i la seva cutícula, composta principalment d'alfa-quitina; també està biomineralitzada amb carbonat de calci. La rigidesa de la cutícula impedeix el creixement, de manera que els artròpodes muden periòdicament. El pla corporal dels artròpodes consisteix de segments repetits, cadascun amb un parell d'apèndixs. El seu pla corporal és tan versàtil que s'ha comparat els artròpodes amb les navalles suïsses, i els ha permès que esdevinguin el gremi ecològic més ric en espècies de la majoria d'ambients. Amb més d'un milió d'espècies descrites representen més del 80% de les espècies vivents conegudes. Són un dels dos únics grups que han conegut un gran èxit a ambients secs –els altres són els amniotes–. La seva mida varia, del plàncton microscòpic a crustacis que mesuren metres. La cavitat interna principal dels artròpodes és un hemocel, que acomoda els òrgans interns i a través del qual flueix la seva sang; tenen un sistema circulatori obert.[3] Com el seu exterior, els òrgans interns dels artròpodes es componen generalment de segments repetits. El seu sistema nerviós s'assembla a una escala, amb cordons nerviosos ventrals aparellats que travessen tots els segments i formen ganglis aparellats a cada segment. El seu cap està format per la fusió d'un nombre variable de segments, i el seu cervell està format per la fusió dels ganglis d'aquests segments i encercla l'esòfag. Els sistemes respiratori i excretor dels artròpodes varien, depenent tant del medi en què viuen com del subfílum al qual pertanyen.
La seva visió es basa en diverses combinacions d'ulls compostos i ocels; en moltes espècies els ocels només poden detectar de quina direcció ve la llum, i els ulls compostos són la font principal d'informació, però els ulls principals de les aranyes són ocels que poden formar imatges i, en alguns casos, poden moure's per seguir les preses.[4] Els artròpodes també tenen una àmplia varietat de sensors químics i mecànics, basats en gran part en modificacions de les nombroses setes (cerres) que es projecten a través de la seva cutícula.
Els mètodes de reproducció i desenvolupament dels artròpodes són diversos; totes les espècies terrestres utilitzen fertilització interna, però sovint es fa per transferència indirecta de l'esperma mitjançant un apèndix o el terra, en lloc d'injecció directa. Les espècies aquàtiques utilitzen o bé fertilització interna o bé fertilització externa. Gairebé tots els artròpodes ponen ous, excepte els escorpins, que donen a llum cries vives després que els ous es descloguin dins la mare. Els nounats varien, d'adults en miniatura a larves i erugues que manquen d'extremitats articulades i que eventualment sofreixen una metamorfosi total per assolir la forma adulta. El grau de cura maternal de les cries varia de zero fins a la cura perllongada que ofereixen els escorpins.
La versatilitat del pla corporal modular dels artròpodes ha fet que sigui difícil pels zoòlegs i paleontòlegs classificar-los i esbrinar-ne l'ascendència evolutiva, que es remunta al període Cambrià i possiblement abans. Des dels anys 1950 i al final dels anys 1970 certs pèrits creien que els artròpodes eren polifilètics, és a dir, que no existí cap avantpassat únic per tots els artròpodes. Avui en dia generalment se'ls considera monofilètics. Tradicionalment es considerava que els parents evolutius més propers dels artròpodes eren els cucs anèl·lids, car els dos grups tenen cossos segmentats. Actualment s'accepta generalment que els artròpodes pertanyen al superfílum dels ecdisozous («animals que muden») mentre que els anèl·lids pertanyen a un altre, el dels lofotrocozous. Les relacions entre els diversos grups d'artròpodes encara són objecte de debat.
Tot i que els artròpodes contribueixen al subministrament d'aliment pels humans directament com a menjar, i de manera més important com a pol·linitzadors dels conreus, també poden estendre algunes de les malalties més greus i causen danys considerables al bestiar i els conreus.
Malgrat aquesta gran diversitat els artròpodes presenten una estructura general força homogènia. Es caracteritzen per:[5][6]
L'èxit dels artròpodes es relaciona amb aquest tipus d'estructura i amb l'eficiència de l'exoesquelet, i han pogut colonitzar hàbitats prohibits per a altres invertebrats i molts cordats.
La rigidesa del cos ha empès l'organisme a desenvolupar els apèndixs articulats, que utilitzen principalment per a la locomoció, la funció primària, però que s'han especialitzat per a realitzar altres funcions com l'alimentació, la percepció sensorial i la defensa. La pressió hidràulica originada per la pressió arterial creada pel cor ajuda en part al sistema muscular.[9] Aquest sistema hidràulic està especialment ben desenvolupat en les aranyes.
La seva mida va des del nivell microscòpic (~0,2 mm) en exemplars planctònics, fins a alguns metres; l'artròpode vivent més gran és un cranc, Macrocheira kaempferi, que té una amplitud de cames que arriba als 3 metres i mig, i alguns artròpodes prehistòrics eren fins i tot més grans, com Pterygotus i Arthropleura.
La cutícula, d'origen epidèrmic, forma un exoesquelet rígid, compost principalment de quitina. Periòdicament, per poder créixer, necessita fer la muda i renovar aquesta cobertura. Està formada per:
L'exoesquelet pren la forma de plaques en els segments (on s'anomenen esclerits) i anells en els apèndixs.[10] Serveix de protecció davant l'atac dels predadors i és impermeable. Per créixer, un artròpode ha de desprendre's del seu exoesquelet vell i secretar-ne un de nou. Aquest procés, l'ecdisi, és car en termes d'energia, i durant aquest període de la muda un artròpode és molt vulnerable.
El cos d'artròpodes es divideix en una sèrie de segments o metàmers que, alhora, s'agrupen formant diverses regions o tagmes. D'aquesta manera la segmentació queda alterada pel fenomen de la tagmatització.
La regió anterior o cefàlica és una constant en tots els artròpodes, on ha desaparegut la metamerització primitiva i s'ha produït una modificació dels apèndixs de manera que exerceixen altres funcions a més de la locomoció: tàctil (antenes), mastegadora (mandíbules, maxil·les) i prènsil (pedipalps, quelícers, pinces, maxil·lípedes).[11]
A més de la regió cefàlica, segons els grups, la resta del cos s'organitza de diferents maneres. Per exemple:
La part més anterior de la regió cefàlica s'anomena àcron i la més posterior del cos, tèlson, i no són considerades metàmers perquè no presenten esbossos mesodèrmics en les fases embrionàries.
Hi ha diferents organitzacions dels aparell respiratori en dels artròpodes. Els aquàtics utilitzen les brànquies per intercanviar gasos. Aquestes brànquies tenen una àrea de superfície extensa en contacte amb l'aigua circumdant. Els artròpodes terrestres han incrementat les superfícies internes especialitzades en l'intercanvi de gasos.
Els insectes i la majoria de les altres espècies terrestres tenen sistemes traqueals, que porten l'aire que va des d'uns porus anomenats espiracles situats a la cutícula de l'epidermis, fins a uns sacs aeris en l'interior del cos. Altres artròpodes utilitzen pulmons de llibre, unes brànquies modificades per a poder respirar aire.
En els crancs terrestres les cambres de les brànquies, a vegades, tenen dues estructures diferents: una que utilitzen per respirar sota l'aigua, i un altre adaptada de manera especial per captar l'oxigen de l'aire, una mena de pseudopulmó.
Excepcionalment, alguns artròpodes realitzen la respiració cutània. Algunes àrees de les potes del cranc Birgus latro es cobreixen amb oxigen que poden absorbir per la membrana.
Els artròpodes tenen un aparell digestiu força variat però de manera general és rectilini i se'n poden destacar tres parts:
La digestió de l'aliment generalment és interna però en molts aràcnids és externa; aboquen els sucs digestius damunt de les preses i posteriorment absorbeixen el producte resultant, que acaben de digerir en el mesodeu.
El sistema circulatori és de tipus obert i el líquid circulatori o hemolimfa banya directament els teixits i òrgans. Presenten un cor, l'òrgan propulsor de l'hemolimfa, que està en posició dorsal, i uns vasos per on circula el líquid.
L'hemolimfa conté hemocianina, una proteïna que transporta l'oxigen basant-se en el coure. El coure és el responsable dels color blau de l'hemolimfa dels artròpodes, i és diferent de la sang dels vertebrats que és vermella perquè utilitzen hemoglobina, i conté ferro. L'hemolimfa és propulsada per una sèrie de cors, disposats dorsalment, i està en contacte directe amb els teixits.
Els artròpodes són protòstoms. Hi ha un celoma, però queda reduït a una cavitat minúscula al voltant dels òrgans reproductors i excretors, i la cavitat dominant del cos és un hemocel ple de l'hemolimfa, que banya els òrgans directament.[12]
El sistema nerviós és de tipus ganglionar i està en posició ventral, a excepció de la massa ganglionar que forma el cervell que està per damunt de l'esòfag. Presenten una estructura que s'assembla a la dels anèlids, però amb més complexitat.
A més del cervell, producte de la fusió de tres parells de ganglis, existeix una cadena ventral de ganglis, que conformen el sistema nerviós central. A més, diferenciat, hi ha un sistema nerviós simpàtic, que estan relacionats amb el sistema central, i que innerva les vísceres.
Les sensílies són les unitats funcionals dels òrgans sensorials dels artròpodes;[5] tenen origen ectodèrmic i normalment tenen forma d'espina, placa, vareta, con, o clavilla; es compon d'una o unes poques cèl·lules amb una connexió nerviosa.
La rígida cutícula dels artròpodes bloquejaria la informació sobre el món exterior si no fos pel fet que és penetrada per molts sensors o connexions de sensors al sistema nerviós. De fet, els artròpodes han modificat la seva cutícula en elaborats conjunts de sensors. Diversos mecanoreceptors, principalment setes, responen a diferents nivells de força, des d'un contacte molt fort fins a corrents d'aire molt febles. Els quimioreceptors detecten el que equivaldria als sentits del gust i l'olfacte, sovint també mitjançant setes. Els receptors de pressió sovint prenen la forma de membranes que funcionen com timpans, però estan connectats directament als nervis en lloc d'a ossicles auditius. Les antenes de la majoria d'hexàpodes inclouen paquets de sensors que determinen la humitat i la temperatura.[13]
En general són dioics, és a dir, hi ha mascles i femelles, sovint amb una clara diferenciació o dimorfisme sexual. Hi ha, però, exemples d'hermafroditisme. La reproducció és sexual però hi ha casos de partenogènesi. Són ovípars, però existeixen exemples d'espècies ovovivípars i vivípars.
El desenvolupament es realitza a través d'una sèrie de fases juvenils o larvàries, la metamorfosi, que en alguns casos implica grans diferències morfològiques.
Els artròpodes es classifiquen típicament en cinc subfílums, dels quals un està extingit:[14][15][16]
Cal destacar que, des que el Codi Internacional de Nomenclatura Zoològica no reconeix cap prioritat per sobre del rang de família dels animals, segons les fonts, els grans grups es poden trobar categoritzats de maneres molt diferents.[17]
A banda d'aquests grups essencials, hi ha també un cert nombre de formes fòssils, principalment del Cambrià inferior, incloent-hi els anomalocàrids, Euthycarcinoidea[18] i Arthrogyrinus que són difícils de situar. També s'observa una manca d'afinitat òbvia amb qualsevol dels grups principals o d'una afinitat clara amb uns quants d'ells.
Subfílums i classes:
Durant moltes dècades, les relacions filogenètiques dels celomats es van basar en la concepció dels articulats de Cuvier,[19] un clade format per anèl·lids i artròpodes. Nombroses anàlisi morfològiques moderns basats en principis cladistes han corroborat l'existència del clade articulats, per exemple, Brusca & Brusca,[20] Nielsen[21] o Nielsen et al.,[22] entre d'altres.
No obstant això, diverses anàlisi cladístiques, como el de dades combinades de Zrzavý et al (1998)[23] estan arribant a la conclusió que anèl·lids i artròpodes no estan directament ralcionats. La presència de metamerizació en anèl·lids i artròpodes hauria de considerar-se, doncs, como una convergècia evolutiva. En canvi, aquests estudis proposen el clade Ecdysozoa en el qual els artròpodes mostren estretes relacions filogenètiques amb grups pseudocelomats, com els nematodes, nematomorfs, priapúlids i cinorincs, per la presència compartida d'una cutícula quitinosa que és mudada (ècdisi) periòdicament.
La filogènia interna dels artròpodes ha estat molt controvertida, amb una enfrontada polèmica entre los partidaris del monofiletisme i els del polifiletisme. Snodgrass[24] y Cisne[25] han defensat el monofiletisme, malgrat que el primer contempla els Artròpodes dividits en aracnats + mandibulats, i el segon els considera dividits en esquizoramis i atelocerats. Tiegs & Manton[26] van defensar el difiletisme, amb els artròpodes dividits en esquizoramis + uniramis i els onicòfors como grupo germà de miriàpodes + hexàpodes. Posteriorment, Manton[27] i Anderson[28] van defensar el polifiletisme del grup (veure Uniramia).
Amb l'aparició dels primers estudis basats en dades moleculars i anàlisi combinades de dades morfològiques i moleculars, sembla que l'antiga polèmica sobre monofília i polifília ha quedat superada, ja que tots ells corroboren que els artròpodes són un grup monofilético en el qual inclouen també els onicòfors i tardígrads (el clade s'ha denominat panartròpodes); la majoria també proposen l'existència del clade mandibulats. No obstant això, han sorgit noves controvèrsies, sobretot al voltant de dues hipòtesis alternatives mútuament excloents que són motiu de dabat en nombrosos articles sobre filogènia i evolució dels artròpodes: atelocerats (miriàpodes + hexàpodes) (Wheeler)[29] (cladograma A) versus pancrustacis (crustacis + hexàpodes) (Giribert & Ribera)[30] (cladograma B):
Els estudis més recents tendeixen a suggerir que els crustacis podrien ser un grup polifilètic per la sorprenent posició dels col·lèmbols, considerats fins ara un grup d'hexàpodes.[31][32] Els miriàpodes i el quelicerats anirien dins el clade Paradoxopoda i els insectes dins els Myriochelata al costat de certs grups de crustacis.
El següent cladograma mostra les esmentades relacions filogenètiques entre els principals grups d'artròpodes existents, obtingudes amb les seqüències d'ADN mitocondrial.[31] S'han destacat en rosa els taxons que són considerats crustacis en les classificacions tradicionals per tal d'emfatitzar les grans discrepàncies entre les filogènies tradicionals i les moleculars
Arthropoda Paradoxopoda![]()
Els artròpodes (Arthropoda) són invertebrats dotats d'un exoesquelet (esquelet extern), un cos segmentat, i extremitats articulades anomenades apèndixs. Els artròpodes inclouen els insectes, els aràcnids, els crustacis els miriàpodes i altres. La paraula prové del grec ἄρθρον, «articulació», i de la paraula πούς «peu» (genitiu ποδός).
Els artròpodes es caracteritzen per les seves extremitats articulades i la seva cutícula, composta principalment d'alfa-quitina; també està biomineralitzada amb carbonat de calci. La rigidesa de la cutícula impedeix el creixement, de manera que els artròpodes muden periòdicament. El pla corporal dels artròpodes consisteix de segments repetits, cadascun amb un parell d'apèndixs. El seu pla corporal és tan versàtil que s'ha comparat els artròpodes amb les navalles suïsses, i els ha permès que esdevinguin el gremi ecològic més ric en espècies de la majoria d'ambients. Amb més d'un milió d'espècies descrites representen més del 80% de les espècies vivents conegudes. Són un dels dos únics grups que han conegut un gran èxit a ambients secs –els altres són els amniotes–. La seva mida varia, del plàncton microscòpic a crustacis que mesuren metres. La cavitat interna principal dels artròpodes és un hemocel, que acomoda els òrgans interns i a través del qual flueix la seva sang; tenen un sistema circulatori obert. Com el seu exterior, els òrgans interns dels artròpodes es componen generalment de segments repetits. El seu sistema nerviós s'assembla a una escala, amb cordons nerviosos ventrals aparellats que travessen tots els segments i formen ganglis aparellats a cada segment. El seu cap està format per la fusió d'un nombre variable de segments, i el seu cervell està format per la fusió dels ganglis d'aquests segments i encercla l'esòfag. Els sistemes respiratori i excretor dels artròpodes varien, depenent tant del medi en què viuen com del subfílum al qual pertanyen.
La seva visió es basa en diverses combinacions d'ulls compostos i ocels; en moltes espècies els ocels només poden detectar de quina direcció ve la llum, i els ulls compostos són la font principal d'informació, però els ulls principals de les aranyes són ocels que poden formar imatges i, en alguns casos, poden moure's per seguir les preses. Els artròpodes també tenen una àmplia varietat de sensors químics i mecànics, basats en gran part en modificacions de les nombroses setes (cerres) que es projecten a través de la seva cutícula.
Els mètodes de reproducció i desenvolupament dels artròpodes són diversos; totes les espècies terrestres utilitzen fertilització interna, però sovint es fa per transferència indirecta de l'esperma mitjançant un apèndix o el terra, en lloc d'injecció directa. Les espècies aquàtiques utilitzen o bé fertilització interna o bé fertilització externa. Gairebé tots els artròpodes ponen ous, excepte els escorpins, que donen a llum cries vives després que els ous es descloguin dins la mare. Els nounats varien, d'adults en miniatura a larves i erugues que manquen d'extremitats articulades i que eventualment sofreixen una metamorfosi total per assolir la forma adulta. El grau de cura maternal de les cries varia de zero fins a la cura perllongada que ofereixen els escorpins.
La versatilitat del pla corporal modular dels artròpodes ha fet que sigui difícil pels zoòlegs i paleontòlegs classificar-los i esbrinar-ne l'ascendència evolutiva, que es remunta al període Cambrià i possiblement abans. Des dels anys 1950 i al final dels anys 1970 certs pèrits creien que els artròpodes eren polifilètics, és a dir, que no existí cap avantpassat únic per tots els artròpodes. Avui en dia generalment se'ls considera monofilètics. Tradicionalment es considerava que els parents evolutius més propers dels artròpodes eren els cucs anèl·lids, car els dos grups tenen cossos segmentats. Actualment s'accepta generalment que els artròpodes pertanyen al superfílum dels ecdisozous («animals que muden») mentre que els anèl·lids pertanyen a un altre, el dels lofotrocozous. Les relacions entre els diversos grups d'artròpodes encara són objecte de debat.
Tot i que els artròpodes contribueixen al subministrament d'aliment pels humans directament com a menjar, i de manera més important com a pol·linitzadors dels conreus, també poden estendre algunes de les malalties més greus i causen danys considerables al bestiar i els conreus.
Yr arthropodau yw'r ffylwm mwyaf o anifeiliaid. Mae mwy na miliwn o rywogaethau gan gynnwys pryfed, cramenogion, corynnod, cantroediaid a miltroediaid. Mae gan arthropodau sgerbyd allanol caled, corff cylchrannog a choesau cymalog.
Členovci (Arthropoda) jsou největší kmen živočišné říše. Jsou nejúspěšnějším a druhově nejpestřejším kmenem v evoluci celé živočišné říše a mají značný hospodářský význam. Popsáno je přibližně 1 000 000 druhů. Velikost dospělců se pohybuje od méně než 1 mm (např. roztoči) až po téměř 60 cm (korýši, hrotnatci).
Ve vývojové větvi prvoústých dosáhli členovci nejvyššího stupně dokonalosti, který je srovnatelný s postavením obratlovců mezi druhoústými. Charakteristickým znakem prvoústých je nestejnocenné (heteronomní) článkování. Během fylogeneze u nich docházelo ke splývání sousedních tělních článků ve větší celky, které bylo provázeno odpovídajícími změnami vnitřních orgánů (např. splýváním nervových ganglií).
Každý tělní článek nesl původně jeden pár končetin. Pokud se uvádí, že se končetiny členovců odvozují od parapodií, tedy tělních výběžků mnohoštětinatců,[2][3] je to téměř jistě omyl, neboť mnohoštětinatci a členovci nejsou navzájem příbuzní a končetiny členovců vznikly nezávisle na parapodiích kroužkovců.[4] Domnělá příbuznost členovců a kroužkovců vedla vědce v roce 1795 k vytvoření taxonu Articulata, který tyto kmeny zahrnoval. Dnes se prosazuje zařazení do skupiny Ecdysozoa.
Během vývoje členovců se zřejmě zmenšil počet končetin.[zdroj?] Pohybovou funkci si zanechaly pouze končetiny na hrudní části těla. Ostatní buď zanikly, nebo se přeměnily (např. v tykadla, čelisti, makadla, kusadla).
Členovci jsou polymerní coelomata, u nichž je výrazná tagmatizace těla na tři větší tělní celky (tagmata): hlavu (cephalon, caput), hruď (thorax) a zadeček (abdomen). Další pak na hlavohruď a zadeček. Tato heteronomní metamerie byla i důvodem evoluční úspěšnosti. Končetiny jsou článkované, s klouby, které umožňují připojení k tělu. Původně nesl každý pár končetin jeden článek, později však některé z nich nabyly jiné funkce (hlavové články). Končetiny se skládají z kyčle (coxa), předkyčlí (trochanter), stehna (femur), holeně (tibia) a chodidla (tarsus) zakončeného dvěma drápky či přísavkami (zákožka svrabová).
Tělo členovců je kryto několikavrstevnou kutikulou, která obsahuje polysacharid chitin (polyacetylglukosamin), vylučovaný jednovrstevnou epidermis. Kutikula je složena z epikutikuly a prokutikuly. Epikutikula je velmi tenká, neobsahuje chitin, je však složena z několik vrstev:
Prokutikula navazuje na epidermis. Obsahuje množství kanálků, které spojují epidermis s okolním prostředím, obsahuje chitin. Je složena z tenké, tmavé a tvrdé exokutikuly (sklerotin, někdy i CaCO3) a endokutikuly.
Kutikula většiny členovců je rozdělena na destičky (tzv. sklerity) spojené bílkovinou gumovité povahy (resilin); naproti tomu např. škrkavky mají tělo složené z jedné souvislé kutikuly. Původně patřily ke každému článku těla čtyři sklerity: tergum (tergit), sternum (sternit) a dva pleurity. Někdy však mohou sklerity patřící k jednomu tagmatu splynout a vytvořit jednotný štít (karapax). Na vnitřní straně se kutikula vlní a tvoří četné výběžky (apodémy), na které se upínají svaly. Naopak na vnější straně se na povrchu tvoří drobné útvary podobné chlupům, ostnům a trnům. Tyto výběžky jsou často spojeny s nervovými buňkami – pak se nazývají senzily.
Protože kutikula je pevná schránka, brání živočichům v růstu. Živočich ji musí několikrát za život svléci. Svlékání se označuje jako ekdyse. Před svlékáním je živočich určitou dobu neaktivní – starý exoskelet se odděluje od epidermálních buněk pod ním. Při procesu (tzv. apolýza) dochází také k oddělení svalů od úponů a nervových zakončení od kutikulárních smyslových orgánů. Do prostoru vzniklého mezi epidermálními buňkami a starou kutikulou je vyloučena exuviální tekutina, která obsahuje zatím neaktivní enzymy. Poté začnou epidermální buňky tvořit pod starou kutikulou novou kutikulu. Tím se aktivují enzymy v exuviální tekutině, rozkládají spodní část staré kutikuly a ta je vstřebána. Prasknutí staré kutikuly vyvolá živočich kroutivými pohyby, příp. i zvýšením tlaku hemolymfy na daném místě těla. Ze starého exoskeletu se vysouká. Svlečená kutikula se nazývá svlečka (exuvie). Někteří členovci svlečku požírají.[5] Jednotlivé vývojové fáze mezi ekdysemi jsou instary.
Jednotlivé fáze mezi svlékáními se nazývají instary. Například když se nymfa vylíhne z vajíčka, jedná se o nymfu prvního instaru. Po prvním svlékání je z ní nymfa druhého instaru, po druhém svlékání je ve třetím instaru atd. Pojem instar se často zaměňuje s pojmem stadium. Stadium označuje fáze vývoje nadřazené instarům, a to stadium vajíčka, larvy, kukly, dospělce (u hmyzu s proměnou dokonalou) a stadium vajíčka, nymfy, dospělce (u hmyzu s proměnou nedokonalou). Stadium larvy nebo nymfy se většinou skládá z několika instarů.[5]
Ekdyse u hmyzu je řízena hormonálně, aktivací ekdysonu, steroidního prohormonu hmyzu.[6]
Kutikula je neprostupná pro vzduch, a proto se členovcům vyvinuly různé druhy dýchacích soustav založené na žábrách, plicních vacích a vzdušnicích.
Vývoj členovců je buď přímý (např. pavouci, raci), nebo nepřímý, tj. přes larvu. Vajíčka členovců jsou mozaiková neboli determinační. To umožňuje také vznik tzv. gynandromorfů, jedinců, jejichž každá strana náleží jinému pohlaví. Jelikož jsou vajíčka mezo- nebo polylecitální, dochází k povrchovému (superficiálnímu) rýhování. Některé velmi primitivní formy mají vajíčka oligolecitální a pak dochází k totálnímu rýhování.[7]
Členovci jsou řazeni do nadřazené skupiny Panarthropoda, která patří mezi Ecdysozoa, tedy prvoústé živočichy svlékající chitinovou kutikulu.
Do skupiny Panarthropoda patří dále želvušky (Tardigrada) a drápkovci (Onychophora). Otázka sesterské skupiny recentních členovců dosud není jednoznačně vyřešena. Z molekulárních analýz vycházejí jako nejbližší drápkovci,[8] podle morfologických analýz uvažujících i fosilní záznam jsou sesterskou skupinou želvušky (společná skupina členovců a želvušek se nazývá Tactopoda).[9]
Kdyby byly uvažovány i prozkoumané vyhynulé skupiny, byly by recentním euarthropodům pravděpodobně nejbližší kambrijské rody Chengjiangocaris, Fuxianhuia a Shankouia;[9][10] do nejbližšího příbuzenstva by patřila i Opabinia a anomalokaridi (Anomalocarididae), známí zejména díky kambrijskému predátoru zvanému anomalocaris.
Recentní členovci se tradičně člení na 4 velké skupiny, zpravidla označované jako podkmeny – klepítkatce, stonožkovce, korýše a šestinohé. Dlouho se usuzovalo na užší příbuznost stonožkovců a šestinohých (společný název Tracheata=Atelocerata – vzdušnicovci) a spekulovalo se, zda korýši jsou bližší příbuzní klepítkatců (společný název Schizoramia) nebo vzdušnicovců (společný název Mandibulata – kusadlovci). V 90. letech 20. století se na základě prvních molekulárních analýz objevily další konkurenční hypotézy – užší příbuznost šestinohých a korýšů (společný název Pancrustacea=Tetraconata) a klepítkatců se stonožkovci (společný název Myriochelata=Paradoxopoda).[11] Konečné rozhodnutí snad přinesla komplexní fylogenetická analýza jaderných genů z r. 2010[12]: Byla vyvrácena hypotéza vzdušnicovců[pozn. 1] a paradoxopodů a potvrzena přirozenost mandibulát a pancrustaceí, korýši v užším slova smyslu byli potvrzeni jako nepřirozená skupina, v jejímž rámci došlo k podstatným přeskupením. Studie však nenalezla dostatečnou podporu pro rozhodnutí o vnitřní příbuzenské struktuře pavoukovců jako hlavní skupiny recentních klepítkatců. Závěry studie byly v témž roce podpořeny širokou fylogenetickou studií jaderných genů, mikroRNA i morfologických znaků,[14] i studií z roku 2011, která již konstatovala prokázání dostatečné robustnoti fylogenetického stromu.[15] Novější a obsáhlejší studie pak přinesly jen drobnější změny, např. v postavení lupenonožců.[16] Molekulární analýzy byly často v rozporu s morfologickými analýzami. Teprve studie z r. 2013 potvrdila molekulární výsledky i rozsáhlou analýzou morfologických znaků z fosilního záznamu se správným uvažováním konvergentních znaků.[17][18]
Následující systém recentních členovců vychází (včetně českých názvů) z BioLibu[19], přičemž nejsou uváděny taxonomické ranky (třídy, podtřídy a řády), protože se v různých systémech liší. Byly provedeny některé názvoslovné úpravy při redukci monotypických úrovní[11] a zejména doplněny některé nové skupiny (označené Regier, 2010) a opuštěny všechny skupiny, které se ve fylogenetické analýze[12] ukázaly jako polyfyletické (např. Maxillopoda), u parafyletických je doplněna poznámka o nepřirozenosti.
Poznámka: Systém neuvádí vyhynulé skupiny členovců – trilobiti (Trilobita), kyjonožci (Gigantostraca), Euthycarcinoida apod. – jejichž postavení v systému je odkázáno na morfologii a nemůže být podpořeno molekulární analýzou.
Arthropoda syn. Euarthropoda - členovci
Výše uvedený systém uvádí některé tradiční, ale parafyletické skupiny. Současné (počátek r. 2013, u klepítkatců r. 2019) představy o skutečné vzájemné příbuznosti jednotlivých skupin recentních členovců ukazuje následující fylogenetický strom, zpracovaný podle fylogenetických analýz ("P?" značí skupiny, u kterých není analýzami vyloučena parafyletičnost; ? skupiny s nedostatečně potvrzenou pozicí).[12][14][15][20][16][21][17][22][23]
Parasitiformes včetně Opilioacariformes
Eumalacostraca
Phyllocarida
Diplostraca
Členovci (Arthropoda) jsou největší kmen živočišné říše. Jsou nejúspěšnějším a druhově nejpestřejším kmenem v evoluci celé živočišné říše a mají značný hospodářský význam. Popsáno je přibližně 1 000 000 druhů. Velikost dospělců se pohybuje od méně než 1 mm (např. roztoči) až po téměř 60 cm (korýši, hrotnatci).
Leddyr (Arthropoda) er en række af dyr, der bl.a. er karakteriseret ved at være segmenterede og have et ydre skelet bestående af kitin[1], et såkaldt exoskelet. Insekter, krebsdyr, edderkopper, tusindben o.a. hører til leddyr. Arthropoda betyder "ledben".[1]
Række: Arthropoda
Leddyr (Arthropoda) er en række af dyr, der bl.a. er karakteriseret ved at være segmenterede og have et ydre skelet bestående af kitin, et såkaldt exoskelet. Insekter, krebsdyr, edderkopper, tusindben o.a. hører til leddyr. Arthropoda betyder "ledben".
Die Gliederfüßer (Arthropoda, von altgriechisch ἄρθρον árthron, deutsch ‚Glied, Gelenk‘ und πούς poús, Gen. ποδός podós, deutsch ‚Fuß‘, gelegentlich eingedeutscht als Arthropoden)[1] sind ein Stamm des Tierreichs. Zu ihnen gehören Insekten, Tausendfüßer, Krebstiere (z. B. Krebse, Entenmuscheln), Spinnentiere (z. B. Spinnen, Skorpione, Milben) und die ausgestorbenen Trilobiten.
Gliederfüßer sind ein sehr erfolgreicher Stamm. Rund 80 Prozent aller bekannten rezenten (nicht fossilen) Tierarten sind Gliederfüßer. Sie verteilen sich auf beinahe eine Million Insektenarten, knapp 10.000 sonstige Sechsfüßer, etwa 16.000 Tausendfüßer, gut 100.000 Spinnentiere und knapp 50.000 Krebstiere[2] (nur Größenordnungen, genaue Zahlen schwanken je nach Autor und Jahr der Publikation). Sie entstanden vermutlich während der kambrischen Explosion, als zu Beginn des Kambriums vor etwa 540 Millionen Jahren die mehrzelligen Tierstämme plötzlich in der Fossilüberlieferung auftraten.
Die nächsten Verwandten der Gliederfüßer sind die Bärtierchen (Tardigrada) und Stummelfüßer (Onychophora), mit denen sie zu den Panarthropoda zusammengefasst werden. Einige Taxonomen nennen diese übergeordnete Gruppe Arthropoda, die „eigentlichen“ Arthropoden werden dann als „Euarthropoda“ bezeichnet.
In der traditionellen Systematik wird diese Gruppe aus morphologischen Gründen mit den Ringelwürmern zu den Gliedertieren (Articulata) zusammengefasst. Neuere molekularbiologische Daten sprechen aber für eine enge Verwandtschaft der Gliederfüßer mit den Fadenwürmern (Nematoda) und deren engster Verwandtschaft (Cycloneuralia). Diese gemeinsame Gruppierung wird Häutungstiere (Ecdysozoa) genannt.
Die Körperhülle der Arthropoden bildet ein Außenskelett, das von den Epithelzellen der obersten Zellschicht (Epidermis) nach außen abgeschieden wird; sie wird als Cuticula bezeichnet. Durch die feste Außenhülle ist ein Wachstum nur dadurch möglich, dass diese Hülle periodisch abgestreift und ersetzt wird (Häutung). Die verhärtete Außenhülle an Körper und Extremitäten ist das Merkmal, welches die Arthropoden am besten kennzeichnet, ähnliche Bildungen kommen nur noch bei den verwandten Bärtierchen vor.
Die Körperhülle besteht aus Chitin und zahlreichen Proteinen. Chitin und Proteine werden durch eingelagerte vom Dopamin abgeleitete Verbindungen untereinander netzartig fest verbunden („sklerotisiert“). Sie kann danach flexibel bleiben (z. B. am Hinterleib der meisten Spinnen oder zahlreicher Insektenlarven). Typischerweise sind aber ausgehärtete, stärker sklerotisierte Platten (Sklerite) eingelagert, die von schwächer sklerotisierten Gelenkmembranen unterbrochen, gelegentlich aber zu Hüllen oder Panzern fest verbunden sein können. Bei vielen Krebstieren und Tausendfüßern und wenigen Insekten(larven) ist die Hülle durch Kalkeinlagerung weiter verstärkt.
Die Hülle ist farblos, strohfarben oder gelblich, häufig durch Melanin, Pigmente, gelegentlich auch durch Strukturfarben dunkel oder bunt gefärbt. Bei den Insekten und Spinnen besitzt sie als äußere Lage eine Lipidschicht als Verdunstungsschutz, die den Tausendfüßern und auch allen landlebenden Krebstieren fehlt.
Arthropoden besitzen einen in Segmente gegliederten Körper. Die Segmente weisen im Grundbauplan gleichen Bau mit gleichen Organen, Anhängen etc. auf (mit Ausnahme des ersten und des letzten). Bei allen Arthropoden ist dieser Grundbauplan durch zahlreiche Abwandlungen nur in Teilen tatsächlich verwirklicht, in der Regel sind die Segmente aber gut erkennbar gegeneinander abgegrenzt, und zahlreiche Organe (z. B. Gliedmaßen, Öffnungen des Herzens, Drüsen, Muskeln) sind segmental angelegt.
Traditionell wird angenommen, im Grundbauplan hätten die Gliederfüßer eine unbestimmte Anzahl in sich gleichartiger Segmente besessen, deren Anzahl erst später im Zuge der Spezialisierung fixiert worden sei. Diese Annahme beruht aber großenteils auf der Analogie mit den Ringelwürmern. Alle Gliederfüßer mit Ausnahme einiger Tausendfüßer, der Kiemenfußkrebse und der ausgestorbenen Trilobiten besitzen eine fixierte Segmentzahl; auch die aus dem Kambrium fossil überlieferten Lobopoden, die allgemein als Stammgruppenvertreter gelten, besaßen eine fixierte (und überschaubar große) Segmentzahl. Innerhalb der Forschung ist noch umstritten, ob die Segmentierung ein ursprüngliches Merkmal ist, welches die Gliederfüßer vom gemeinsamen Vorfahren der Bilateria geerbt haben, oder ob es sich um eine evolutive Neuheit handelt; in diesem Fall wäre bei den anderen im Grundbauplan segmentierten Tierstämmen (vor allem den Ringelwürmern und den Chordatieren) die Segmentierung konvergent entstanden.
An jedem Segment kann ein Paar ebenfalls in Abschnitte gegliederter Gliedmaßen (Beine, Mundwerkzeuge, Antennen) sitzen, welches aber an verschiedenen Segmenten auch fehlen kann. Die entwicklungsgeschichtliche Grundlage von Körpersegmenten und Beinabschnitten ist dabei vergleichbar, die Extremitäten können als sekundäre Körperachsen aufgefasst werden.
Auch das Nervensystem ist segmental angelegt und besteht aus einem auf der Bauchseite verlaufenden Doppelstrang mit einem Ganglion und einer Querverbindung (Kommissur) pro Segment („Strickleiternervensystem“). Typisch ist auch ein rückenseitiges (dorsales) Herz und ein offener Blutkreislauf.
Der ursprüngliche Aufbau eines Segments besteht aus dem Rumpfteil mit einem Ganglion und einem Paar zweiästiger (biramer) Gliedmaßen (Spaltbeine). Der (untere) Innenast wird Beinast oder Schreitbein genannt, der Außenast Kiemenast, womit auch die Funktionen angedeutet sind.
Bei der Diversifizierung der Gliederfüßer im Laufe der Evolution kam es zu Gruppenbildungen oder auch Verschmelzungen von Segmenten. Welche Segmente miteinander zu einem Tagma verschmolzen sind, ist bei unterschiedlichen Gruppen verschieden. Bei allen Gliederfüßern ist es zur Verschmelzung einiger der vorderen Segmente zum Kopf gekommen, der ein aus mehreren Ganglien verwachsenes Gehirn und zu Mundwerkzeugen und Antennen geformte ein- oder zweiästige Gliedmaßen besitzt. Bei den verschiedenen Gruppen der Gliederfüßer ist eine unterschiedliche Anzahl von Segmenten zum Kopf verschmolzen.
Bei urtümlichen Gliederfüßern wie den Remipedia besteht die Gliederung des Körpers hinter dem Kopf zum großen Teil noch aus nahezu identischen Segmenten. Die Trilobiten hatten einen ähnlich repetitiven Körperbau.
Bei den Tausendfüßern sind aus Vorfahren mit relativ wenigen Segmenten (je nach Stammgruppe 12 bis 15) sekundär wieder Tiere mit zahlreichen gleichartigen Segmenten hervorgegangen; bei den Doppelfüßern (Diplopoda) sind jeweils zwei aufeinanderfolgende Segmente miteinander zu einem Doppelsegment verschmolzen, so dass jedes „Segment“ des Rumpfes zwei Beinpaare hat; bei vielen Hundertfüßern folgen jeweils ein breites und ein schmales Segment aufeinander.
Bei den Insekten ist die Tagmatisierung weiter fortgeschritten: Jeweils mehrere Segmente sind zu Kopf, Mittel- und Hinterleib verschmolzen. Die drei so entstehenden Körperabschnitte haben eine sehr unterschiedliche Funktion. Die Gliedmaßen der ursprünglichen Segmente des Kopfes sind zu Antennen und Mundwerkzeugen umgebildet. Im Mittelleib (Thorax) sind drei Segmente zu einem Tagma verwachsen, dessen drei Paar Gliedmaßen den Außenast verloren haben und als Schreitbeine dienen. Solche Gliedmaßen heißen einästig (uniram). Beim Hinterleib können die Gliedmaßen vollständig zurückgebildet sein oder zu besonderen Organen (Kiemen, Begattungsorgane, Legebohrer) umgebildet sein, sie dienen aber nie als Laufbeine der Fortbewegung. Bei Gliederfüßern, die über keine sichtbaren Kiemen verfügen, erfolgt die Atmung entweder über in die Körperoberfläche eingestülpte Kiemen (Lungen) oder über ästig verzweigte Einstülpungen der Körperwand, die Tracheen. Der Hinterleib kann aus unterschiedlich vielen Segmenten ohne Gliedmaßen bestehen. Die Insektenflügel werden traditionell nicht als Gliedmaßen, sondern als umgebildete Seitenplatten der Brustsegmente gedeutet. Neuere genetische und entwicklungsbiologische Erkenntnisse deuten aber darauf hin, dass sie auf den Kiemenast des ursprünglichen Spaltbeins zurückgehen. (In diesem Fall sind die Kiemen wasserlebender Insektenlarven, z. B. der Eintagsfliegen, homologe Bildungen. Aus dem Mesozoikum existieren fossile Insektenlarven, die sich als Übergangsglieder deuten lassen.)
Bei den anderen Unterstämmen ist die Verschmelzung der Segmente in anderen Varianten erfolgt. Spinnen bestehen aus zwei Abschnitten. Der Vorderleib (Prosoma) besteht aus sechs verschmolzenen Segmenten. Eines der Beinpaare wurde zu Mundwerkzeugen, den Cheliceren, umgebildet, eines zu Tastern und vier zu Beinen. Die Gliedmaßen des Hinterleibs (Opisthosoma) haben den Beinast verloren, der Kiemenast liegt als Fächerlunge in Einstülpungen des Hinterleibs; bei manchen Arten wurden diese Fächerlungen in Tracheen umgewandelt. Aus den letzten Beinpaaren sind die Spinnwarzen entstanden. Bei den Weberknechten und Scheinskorpionen sind Vorder- und Hinterkörper zu einem kompakten Körper verschmolzen.
Bei den Pfeilschwanzkrebsen (Xiphosura) sind am Hinterleib noch Beine erhalten (Blattbeine), deren Kiemenäste Fächerkiemen bilden, die den Fächerlungen der Spinnen sehr ähnlich sind.
Bei den Krebstieren ist der Körper meist ebenfalls in Kopf, Thorax und Abdomen gegliedert, der Bauplan ist aber vielfältig abgewandelt. Bei zahlreichen Krebsordnungen sind ein oder zwei ursprüngliche Thoraxsegmente funktionell mit dem Kopf verschmolzen. Der Thorax weist eine sehr unterschiedliche Anzahl von beintragenden Segmenten auf. Ein gliedmaßenloses Abdomen ist je nach Verwandtschaftsgruppe vorhanden oder kann auch fehlen. Bei den Malacostraca ist anstelle des Abdomens ein gliedmaßentragendes Pleon ausgebildet. Am Körperende steht bei ihnen ein aus mehreren Anhängen verschmolzenes Schwanzruder. Viele andere Krebse tragen hier lange Anhänge (meist als Furca bezeichnet). Die Körpergliederung kann bei vielen Krebstieren durch eine Körperduplikatur, den Carapax, verdeckt sein. Diese kann einteilig oder zweischalig gegeneinander beweglich ausgebildet sein; bei einigen Gruppen (Ostracoda, Conchostraca) umhüllt sie den ganzen Körper, der dadurch muschelförmig aussieht. Bei den Malacostraca ist der ursprünglich nur am Kopf ansitzende Carapax mit dem Thorax fest verwachsen und bildet so einen einheitlichen, nicht mehr in sich beweglichen Cephalothorax. Bei einigen festsitzenden (Seepocken, Entenmuscheln), besonders aber bei parasitischen Gruppen kann der Körperbau vollkommen umgestaltet sein, so dass das Tier kaum noch als Gliederfüßer erkennbar ist.
Gliederfüßer besitzen am Kopf eine unterschiedliche Anzahl (bis zu neun) Augen mit sehr unterschiedlichem Aufbau und unterschiedlichen Funktionen.[3][4][5] Die Augen aller Gliederfüßer besitzen dabei einen Bauplan, der auf wenige Grundtypen zurückgeführt werden kann, deren teilweise stark abgewandelte Endpunkte durch Reihen miteinander verbunden werden können. Es ist daher anzunehmen, dass die Augen aller Gliederfüßer homologe Bildungen darstellen. Die Augen lassen sich in zwei Typen zusammenfassen, die jeweils durch Innervation, Rezeptor- und Transmittermoleküle gekennzeichnet sind, dies sind die Seitenaugen und die Mittelaugen.
Die nach außen gewölbten Seitenaugen sind Facettenaugen, die aus zahlreichen Einzelaugen bestehen (Komplexaugen). Die Einzelaugen werden Ommatidien genannt. Bei den Insekten und den Krebstieren ist der Bau der Ommatidien sehr einheitlich: Sie bestehen aus einer Netzhaut, die acht lichtempfindliche Sinneszellen enthält, darüber liegt ein durchsichtiger Kristallkegel aus vier sogenannten Semperzellen. Seitlich zu diesem liegen zwei Pigmentzellen, die seitlich einfallendes Licht abschirmen, diese scheiden auch die äußere Cornea ab. Die Cornea ist bei den meisten Gruppen linsenförmig verdickt. Neben diesen beiden sogenannten Hauptpigmentzellen existieren zahlreiche weitere Nebenpigmentzellen. Der Aufbau der Ommatidien kann in anderen Gruppen der Gliederfüßer sekundär abgewandelt oder vereinfacht sein.
Komplexaugen existieren bei den meisten Insekten (sie fehlen den anderen Hexapoda) und zahlreichen Krebstieren. Bei den Cheliceraten sind Komplexaugen nur in einer Gruppe vorhanden, bei den Pfeilschwanzkrebsen. Die einzigen Tausendfüßer, die Komplexaugen besitzen, sind die Spinnenläufer (Scutigeromorpha). Die Ommatidien dieser Gruppen weichen in ihrem Feinaufbau allerdings deutlich von denjenigen der Insekten und Krebstiere ab.[6][7] Beim Pfeilschwanzkrebs Limulus besteht jedes Ommatidium z. B. aus mehr als 300 Zellen.
Bei den Gruppen, bei denen sie vorhanden sind, sind die bildgebenden Komplexaugen in der Regel die wichtigsten Augen.[8][9] Die Sehzellen aller Retinazellen eines Ommatidiums wirken dabei zusammen und bilden einen Bildpunkt. Die Auflösung des Komplexauges richtet sich daher nach der Anzahl der Ommatidien, die von einigen wenigen über einige hundert (z. B. etwa 700 bei der Taufliege oder 1000 beim Pfeilschwanzkrebs) bis zu einem Maximum von 30.000 bei einigen Großlibellen-Arten reicht. Die räumliche Auflösung eines Komplexauges reicht dabei niemals an die hochentwickelten Linsenaugen der Wirbeltiere und der Kopffüßer heran, dafür ist ihre zeitliche Auflösung beim Bewegungssehen deutlich besser.
Neben dem räumlichen Sehen können zahlreiche Gliederfüßer aus allen Linien Farben unterscheiden. Wie beim Menschen geht das Farbsehen bei ihnen auf den Besitz unterschiedlicher Sehpigmente (wie generell im Tierreich Varianten von Opsin) zurück, die bei unterschiedlichen Wellenlängen ansprechen. Ebenfalls analog zum Menschen besitzen die meisten drei solcher Pigmente (trichromatisches Sehen). Im Unterschied zum Menschen ist allerdings die Empfindlichkeit vielfach zum kurzwelligen Teil des Spektrums verschoben. Viele Arten können deshalb kein Rot sehen, dafür einen Anteil im Ultraviolett. Eine weitere Sinnesqualität, die die Gliederfüßer dem Menschen voraus haben, ist das Sehen der Polarisationsebene von polarisiertem Licht. Dies ermöglicht ihnen zum Beispiel, bei bedecktem Himmel den Stand der Sonne zu erkennen, und erleichtert die Entdeckung von Wasserflächen.
Fast alle Gruppen, die Komplexaugen besitzen, haben daneben noch weitere Augen, die auf die Mittelaugen zurückgehen (Ausnahme: Mittelaugen fehlen bei allen Tausendfüßern). Bei den anderen Arthropoden, bei denen die Augen nicht komplett rückgebildet sind, kommen ebenfalls weitere Augentypen vor. Bei den Insekten sind auf der Stirn in der Regel drei Punktaugen (Ocellen) vorhanden, deren Funktion nicht in allen Fällen geklärt ist, zumindest bei einigen Gruppen dienen sie zur Lageinformation im Flug. Bei den Krebstieren bilden in vielen Gruppen drei Mittelaugen (nur bei den Phyllopoden vier) ein zentrales Sehorgan, das Naupliusauge. Dieses ist das wichtigste Sinnesorgan der Krebslarven, aber auch bei zahlreichen Adulti vorhanden. Bei den Malacostraca sind häufig ein Paar rückenseitige (dorsale) und ein Paar bauchseitige (ventrale) Mittelaugen ausgebildet, bei anderen Gruppen kann eine andere Anzahl vorhanden sein.[10] Bei den echten Spinnen, die niemals Komplexaugen besitzen, existieren nur ein Paar Mittelaugen und meist zwei oder drei Paar Seitenaugen. Die Seitenaugen dienen bei ihnen aber nie zum bildlichen Sehen, sie sind bei den Wolfsspinnen, den Springspinnen und den Krabbenspinnen Bewegungs-Detektoren, bei den übrigen Gruppen dienen sie nur zur räumlichen Navigation nach dem Sonnenstand, häufig durch Erkennung von polarisiertem Licht.[11]
In der artenreichen Gruppe der Gliederfüßer existieren in zahlreichen Gruppen Spezialanpassungen, die jeweils besondere Sinnesleistungen ermöglichen. So haben die Springspinnen vergrößerte Mittelaugen, die ihnen ein Bildsehen nach ähnlichen Prinzipien wie beim Linsenauge der Wirbeltiere ermöglicht. Einige Gruppen (so beim Copepoden Labidocera oder den Larven des Schwimmkäfers Thermonectes) haben Augen mit minimalem Gesichtsfeld und hoher Auflösung entwickelt, mit denen sie mit einer Scanner-Bewegung einem Lesekopf gleich ihre Umgebung abtasten. Besonders verbreitet sind Anpassungen an schwache Lichtverhältnisse. Viele dämmerungsaktive oder in größeren Meerestiefen lebende Arten haben die Komplexaugen so umgebildet, dass von einem größeren Ausschnitt der Umgebung Licht jede Rezeptoreinheit erreichen kann (Superpositions-Auge). Einige vor allem bodenlebende Gruppen haben allerdings Augen vollkommen aufgegeben. Sie sparen dadurch einen erheblichen Energieverbrauch ein, denn neben den Augen selbst dient ein erheblicher Anteil des Gehirns bei allen Arthropoden mit Fähigkeit zum Bildsehen der optischen Auflösung.
Neben den Augen besitzen zahlreiche Gliederfüßer zusätzlich einen Haut- oder Nervenlichtsinn, der vollkommen unabhängig vom Besitz von Augen ist.[12] Dieser kann z. B. zur Feinderkennung durch Schattenwurf dienen (z. B. beim Flusskrebs Procambarus clarkii) oder durch Erkennung der Tageslänge die Gonaden steuern, wie bei einigen Schmetterlingsarten.
Arthropoden nutzen hauptsächlich zwei unterschiedliche Wege, um mechanische Reize wahrzunehmen, durch Sinneshaare bzw. -borsten und durch Rezeptoren, die die Verformung der Cuticula (des Außenskeletts) registrieren. Diese Organe dienen zur Wahrnehmung der Lage von Körperteilen im Raum (Propriorezeptoren), zur Nahorientierung (Tastsinn) und zur Fernorientierung (Vibrations-, Strömungssinn, Hören). Obwohl zahlreiche Sensoren mehrere dieser Aufgaben parallel erledigen können, existieren teilweise hochspezialisierte Organe für besondere Zwecke.[13][14]
Der Tastsinn der Arthropoden beruht großenteils auf einfachen Sinneshaaren. Daneben haben vor allem die luftlebenden Insekten und Spinnentiere extrem dünne Sinneshaare entwickelt, die als Strömungs- und Vibrationsfühler dienen, diese werden bei Insekten Fadenhaare, bei Spinnentieren Trichobothrien genannt.[15] Im Unterschied zu normalen Haaren sind diese in der Regel in einer ringförmigen Einsenkung der Cuticula eingelenkt. Direkt in der Cuticula sitzen bei allen Gliederfüßern Sensillen, die sowohl Berührungsreize als auch Vibrationen wahrnehmen können. Die Rezeptoren (Scolopidien) können besondere Sinnesorgane ausbilden, wie die Spaltsinnesorgane der Spinnentiere, die bei den echten Spinnen gruppenweise (lyraförmiges Organ) gebündelt sind. Die nur bei den Insekten und Krebstieren nachgewiesenen Chordotonalorgane sind von außen überhaupt nicht sichtbar. Ihre Skolopidien bestehen aus ein bis vier Sinnesneuronen, einer Hüllzelle (Skolopalzelle), mehreren Anheftungszellen und umhüllenden Gliazellen. Ihre Funktionsweise ist im Detail noch nicht geklärt, mit Sicherheit dienen aber modifizierte Cilien der eigentlichen Reizaufnahme.[16] Bei den Insekten kommen zusätzlich sogenannte kuppelförmige (campaniforme) Sensillen vor. Wichtige Tast-, Vibrations- und Strömungssinnesorgane sind bei ihnen die Fühler, zur Wahrnehmung dient ein besonders Organ in der Fühlerbasis, das Johnstonsche Organ.
Spinnen und viele Insekten nehmen die Schwerkraft durch Zugkräfte des an den Beinen aufgehängten Körpers wahr, bei Spinnen dienen dazu lyraförmige Organe, manche Insekten nehmen mit den Johnstonschen Organen den Zug der Schwerkraft an den Antennen wahr. In Wasser lebende Krebstiere weisen vielfach dem umgebenden Medium gegenüber kaum Dichteunterschiede auf (sie können im Wasser schweben). Sie haben daher eigene Schwersinnesorgane entwickelt, die Statocysten. Diese bestehen aus einer Kapsel mit zahlreichen nach innen zeigenden Haarsinneszellen. In diese Kapsel werden bei der Häutung von außen einige Sandkörnchen eingefügt, die durch den Druck auf Haare einer Seite die Richtung der Schwerkraft anzeigen. Die Funktionsweise entspricht beinahe völlig dem Schweresinnesorgan der Wirbeltiere. Statocysten sitzen meist in den basalen Antennengliedern, in einigen Gruppen aber auch am Körperende in den Uropoden oder im Telson.
Wahrnehmung von Schallwellen ist eine Sonderform des Vibrationssinns. Zahlreiche Gliederfüßer können dafür dieselben Haarsensoren nutzen, mit denen sie Luftbewegungen (z. B. durch einen sich nähernden Räuber) oder Erschütterungen wahrnehmen. Bei einigen echten Spinnen ist so eine Schallwahrnehmung mit den Trichobothrien nachgewiesen worden. Bei Stechmücken erkennen die Männchen die Weibchen an dem Schall, der durch die schnelle Vibration der Flügel erzeugt wird (und den auch Menschen hören können).[17] Dazu sind bei ihnen die Antennen stark vergrößert, Hörorgan ist das Johnstonsche Organ. Akustische Kommunikation durch Vibrationen, die mit den Trichobothrien wahrgenommen werden, sind auch bei einem Geißelskorpion nachgewiesen worden und möglicherweise weit verbreitet.[18] Bei den Insekten sind neben diesen eher im Nahbereich arbeitenden Sensoren, die auf direkter Wahrnehmung der Partikelbewegung beruhen, besondere Organe entwickelt worden, die auch auf Druckunterschiede reagieren, d. h. Schallwellen wie die Wirbeltierohren wahrnehmen, diese werden Tympanalorgane genannt. Neben der Wahrnehmung von Feinden dient Schallwahrnehmung bei den auch zur Lauterzeugung fähigen Heuschrecken und Zikaden (und möglicherweise bei Wasserwanzen) der Geschlechterfindung. Parasitische Fliegen haben dies ausgenutzt, indem sie Tympanalorgane entwickelt haben, die die Gesänge von Grillen ebenfalls hören können, die sie so suchen können, um ihre Eier auf ihnen abzulegen.[19] Die meisten Insekten mit Tympanalorganen nutzen diese allerdings nur zu einem einzigen Zweck: um die Ultraschall-Ortungslaute von Fledermäusen zu hören. Diese Fähigkeit hat sich unabhängig voneinander bei sieben Schmetterlingsfamilien, bei Fangschrecken, Florfliegen und zwei Käfergruppen (Sandlaufkäfer und Blatthornkäfer) entwickelt. Die Tympanalorgane sind in jedem Fall abgewandelte Chordotonalorgane, bei denen die Sinneszellen auf Luftsäcken (erweiterten Tracheen) im Körperinnern liegen. Die Lage ist dabei extrem verschieden, im Hinterleib (Zikaden, die meisten Schmetterlinge, Kurzfühlerschrecken, Sandlaufkäfer), im Thorax (Eulenfalter, Fangschrecken), in den Beinen (Langfühlerschrecken), an der Flügelbasis (Schwärmer, Florfliegen), hinter dem Kopf (Fliegen, Blatthornkäfer), an den Mundwerkzeugen (Schwärmer).[20] Während viele Gruppen nur ein schmales Frequenzband im Ultraschall hören können, besitzen einige ein hoch entwickeltes Entfernungs- und Richtungshören.
Erst seit wenigen Jahren ist nachgewiesen, dass auch Krebse (Decapoda) einen Hörsinn besitzen.[21] Diese können niederfrequente Schallwellen von etwa 100 bis 3000 Hertz mit ihren Statocysten wahrnehmen.
Chemische Sinnesorgane zur Nahorientierung (Geschmackssinn) und zur Fernorientierung (Geruchssinn) liegen in Form einer Vielzahl von Sensillen-Typen bei allen Arthropoden vor.[22][23][24] Insbesondere bei allen Gruppen ohne hochentwickelte Augen handelt es sich in der Regel um die wichtigsten Organe zur Nahrungssuche. Außerdem beruht auch die soziale Kommunikation und die Geschlechterfindung bei zahlreichen Arthropoden auf Pheromonen, die über chemische Sinnesorgane wahrgenommen werden. Wichtigste Sinnesorgane zur chemischen Orientierung sind meistens die Antennen, häufig auch die Taster (Palpen) und die äußeren (distalen) Beinglieder. Fehlen Antennen und Taster, kann die Rezeptorausstattung der Beine ähnlich reichhaltig sein, wie z. B. bei den Geißelspinnen.[25] Sehr viele Gliederfüßer haben darüber hinaus chemische Sensoren verstreut auf der gesamten Körperoberfläche. Auf einer Gliederfüßer-Antenne werden regelmäßig ein bis mehrere hunderttausend Chemorezeptoren gefunden.
Die Sensillen vor allem der Antennen werden nach ihrer Morphologie in zwei verschiedene Gruppen eingeteilt, den Haarsensillen und den Grubensensillen, die durch Übergänge verbunden sind (stift- oder kegelförmige Fortsätze, die in einer Grube liegen). Des Weiteren werden ihrer Morphologie entsprechend verschiedene Arten von Haaren, nämlich kuppelförmige (campaniformia), flaschenförmige (ampullacea), kegelförmige (basiconia) und etliche andere Typen unterschieden. Zwar steht die Form der Rezeptoren in Beziehung zur Funktion, allerdings zeigt es sich, dass gleich geformte Sinnesorgane bei verschiedenen Tiergruppen unterschiedliche Funktion besitzen. Typisch für alle Typen von Chemorezeptoren ist es, dass ihre Oberfläche von Poren (im Mikro- bis Nanometerbereich) bedeckt ist. Durch diese Poren steht der hohle Innenraum (Lumen) der Sensille mit der Außenwelt in Kontakt. Hereindiffundierende Moleküle werden von spezialisierten Nervenzellen, die die eigentlichen Sinneszellen darstellen, registriert. Sehr häufig kann ein einzelnes Sinneshaar dabei verschiedene Sinneseindrücke verarbeiten, z. B. als kombinierter Chemo- und Mechanorezeptor, selten sogar drei (trimodal). Auf einer Antenne sind üblicherweise fünf bis zehn unterschiedliche Sensillentypen morphologisch unterscheidbar.
Dem Nahrungserwerb dienende Sensillen besitzen typischerweise ein relativ breites, aber zwischen verschiedenen Zellen recht differenziertes Sinnesspektrum für eine Vielzahl löslicher oder flüchtiger Substanzen wie Aminosäuren, Peptide, Zucker, Nukleotide oder Amine. Aus dem Erregungsmuster verschiedener Sensoren kann dann die entsprechende Substanz ermittelt werden (ähnlich wie der Geschmackssinn beim Menschen).[26] Daneben existieren vor allem für Botenstoffe (Pheromone) hochspezialisierte Sensoren, die nur diesen einzigen Stoff wahrnehmen.
Eine Reihe von Arthropodenarten, darunter Schmetterlinge, Mücken, Landasseln, Tausendfüßer und Zecken besitzen spezielle Sensoren, die spezifisch auf Kohlendioxid ansprechen.[27][28] Meist handelt es sich um Sensillen auf der Antenne (häufig flaschenförmige Sensillen), aber auch spezielle Sinnesorgane auf der Kopfoberfläche sind hierfür entwickelt worden, z. B. von Tausendfüßern. Der biologische Zweck dieser Sinnesleistung besteht z. B. in der Wirtsfindung bei blutsaugenden Arten oder in Vermeidungsreaktionen gegen Ersticken bei bodenlebenden Arten, die in engen Hohlräumen leben.
Obwohl es sich dabei streng genommen nicht um chemische Sinne handelt, werden sie von Gliederfüßern von ähnlich aufgebauten Sensillen wahrgenommen. Häufig dienen kegelförmige Sensillen zur Abdeckung beider Eigenschaften, oft sogar kombiniert miteinander. Berühmt geworden sind die spezialisierten Infrarot-Strahlungsdetektoren des „Feuerkäfers“ Melanophila. Diese Tiere können bis in 12 Kilometer Entfernung Waldbrände feststellen, die sie gezielt anfliegen, um ihre Eier in das verkohlte Holz abzulegen.
Wahrnehmung des Erdmagnetfelds ist im Tierreich weit verbreitet und auch bei vielen Gliederfüßern nachgewiesen. Zur Wahrnehmung dienen intrazelluläre Eisenoxid-Partikel, wie sie beispielsweise bei sozialen Insekten[29] und Krebsen[30] nachgewiesen worden sind. Der magnetische Sinn dient wohl in allen Fällen zur großräumigen Orientierung im Raum.
Während man früher dachte, ein Sinn für elektrische Felder würde bei Arthropoden fehlen, ist es inzwischen gelungen, einen solchen bei Krebsen doch nachzuweisen.[31] Dieser ist in seiner Sensitivität allerdings eine Größenordnung schlechter als die hochentwickelten elektrischen Sinnesorgane vieler Fischarten. Wofür die Krebse den Sinn nutzen, ist noch nicht geklärt, möglich wäre eine Beteiligung an der räumlichen Orientierung oder bei der Wahrnehmung von Beuteorganismen.
Während einerseits bei vielen Arten der Arthropoden Männchen und Weibchen nur anhand ihrer Geschlechtsorganen unterscheidbar sind, unterschieden sich andererseits die Geschlechter bei diversen Arten auch äußerlich erheblich.
Ein Beispiel für deutliche Farbunterschiede zwischen den Geschlechtern findet sich bei der Großen Goldschrecke (Chrysochraon dispar) aus der Ordnung der Kurzfühlerschrecken. Ein weiteres Beispiel für deutlich ausgeprägten Geschlechtsdimorphismus ist der Hirschkäfer (Lucanus cervus) aus der Ordnung der Käfer. Hier haben die Männchen deutlich vergrößerte, geweihähnliche Mandibeln, mit denen sie gegeneinander um die weniger auffälligen Weibchen kämpfen.
Bei manchen Arten gibt es Zwergmännchen, die erheblich kleiner sind als ihre Weibchen, so beispielsweise beim Schwan (Euproctis similis) aus der Ordnung der Schmetterlinge oder bei der Wespenspinne (Argiope bruennichi) aus der Ordnung der Webspinnen.
Es gibt bei den Insekten viele Fälle, in denen die Männchen im Gegensatz zu den Weibchen geflügelt sind, beispielsweise bei der Gemeinen Küchenschabe (Blatta orientalis) aus der Ordnung der Schaben oder dem Schlehen-Bürstenspinner (Orgyia antiqua), einem Schmetterling. Im Extremfall ist das Weibchen larvenähnlich und ziemlich groß, während das Männchen flugfähig ist, voll, entwickelt und erheblich besser ausgebildete Sinnesorgane hat. Ein Beispiel hierfür ist der Kleine Mottenspinner (Heterogynis penella), ebenfalls aus der Ordnung der Schmetterlinge.
Bei den Arthropoden haben sich verschiedene Wege entwickelt, den jeweiligen Geschlechtspartner anzulocken.
Bei vielen Schmetterlingen haben die Weibchen einfache fadenförmige Fühler, während die männlichen Tiere gekämmte Fühler haben, da sie die größere Oberfläche nutzen, um den Sexuallockstoff des Weibchens auf möglichst große Entfernung wahrnehmen zu können.
Andere Arten nutzen optische Signale: So winken die männlichen Winkerkrabben mit ihrer vergrößerten Schere, Leuchtkäfer locken ihre Männchen durch einen arttypischen Blinkrhythmus an.
Auch Geräusche werden genutzt: Wolfsspinnen balzen mit Klopfzeichen. Riesenkrabbenspinnen locken die Weibchen an, indem sie die Oberfläche, auf der sie sitzen, in Vibration versetzen. Langfühlerschrecken zirpen (Stridulation).
Bei den meisten Arthropoden findet eine innere Befruchtung statt. Ausnahmen wie bei Limulus sind selten. Die Spermien sind dabei oft in einer Spermatophore zusammengefasst, die vom Weibchen aufgenommen oder vom Männchen im oder am Weibchen befestigt wird. Bei einigen Arten wurden deshalb Beine zu Gonopoden umgebildet, die der Befruchtung dienen. Die Spermatophoren öffnen sich oft erst lange nach der Begattung, um die Eier zu befruchten.
Bei manchen Insekten passen die Geschlechtsorgane der Männchen nur zu denen der Weibchen derselben Art (Schlüssel-Schloss-Prinzip), so dass eine Befruchtung zwischen verschiedenen äußerlich nicht zu unterscheidenden Arten oft allein deshalb nicht möglich ist. Manchmal fressen die weiblichen Tiere nach der Begattung die Männchen, wie beispielsweise bei der Großen Winkelspinne.
Parthenogenese ist recht häufig bei Arthropoden. Sie tritt bei einigen Milben (Acari), Krebsen (Crustacea) und bei Insekten auf. Beispiele hierfür sind die Große Sägeschrecke und die Sommergenerationen der Blattläuse, im Herbst treten geflügelte Geschlechtstiere auf.
Zwittertum ist bei den Arthropoda eine seltene Ausnahme. Cyatura carinate (Isopoda) und viele Scherenasseln (Tanaidacea) sind zuerst Weibchen und dann Männchen (protogynes Zwittertum).
Im Bereich der Brutpflege gibt es bei den Gliederfüßern alle Varianten, von der Eiablage in freies Wasser ohne jegliche Fürsorge bis hin zu lebendgebärenden Tieren, wie bei Euscorpius mesotrichus.
Insekten legen ihre Eier oft an die Futterpflanzen. Endoparasiten wie Erzwespen und die Gemeine Eichengallwespe legen sie gleich in den Wirt. Weibliche Höhlenspinnen und Zitterspinnen der Gattung Pholcus umhüllen ihre Eier mit einem Gespinst und tragen den Eikokon mit sich herum. Der weibliche Edelkrebs trägt die Eier am Körper festgeklebt mit sich herum, bis sie schlüpfen. Einige Arten wie beispielsweise die Ohrwürmer bewachen ihre Eier in einer geschützten Höhle, bis sie schlüpfen.
Die Asseln haben ähnlich den Beuteltieren eine Brusttasche, in denen sie ihre Jungen herumtragen, wie bei Kellerassel und Wasserassel beschrieben. Die Fleckige Brutwanze bewacht und beschützt ihre Eier nicht nur, sie führt die Jungtiere später auch wie eine Ente ihre Jungen im Gänsemarsch zum Futter. Einige Arten wie der Gemeine Ohrwurm und die Kugelspinnen sowie die meisten staatenbildenden Insekten gehen noch weiter mit der Brutpflege und füttern ihre Jungen.
In Arthropodeneiern beginnt die Embryonalentwicklung mit einer totalen Furchung bei dotterarmen und oberflächlichen Furchung an verschiedenen Stellen der Oberfläche bei dotterreichen Eizellen. Bei oberflächlicher Furchung entsteht eine kugelförmige einzellige Schicht um den Dotter herum, häufig mit zahlreichen Zellkernen, aber ohne Zellgrenzen (Syncytium). Als Nächstes wandern die Zellen zu einer Seite des Dotters und bilden dort eine Keimscheibe, aus der der Embryo entsteht.
Nur bei primär wasserlebenden Gruppen schlüpft eine primäre Larve aus dem Ei, dies sind neben den meisten Krebstieren die zu den Chelicerata gehörenden Asselspinnen. Die Krebstiere schlüpfen mit drei Gliedmaßenpaare tragenden Segmenten (diejenigen der ersten und zweiten Antennen und der Mandibeln) als Naupliuslarve, Asselspinnen mit ebenfalls drei Segmenten (Anlagen der Cheliforen, Palpen und Brutbeine) als Protonymphon-Larve. Bei den folgenden Häutungen werden jeweils in einer hinteren Wachstumszone weitere Segmente angefügt. Die Gliedmaßen dieser Segmente werden häufig nur als ungegliederte Knospen angelegt und erhalten erst im Laufe weiterer Häutungen nach und nach funktionsfähige Gestalt. Auch die Gliedmaßen der bereits vorhandenen vorderen Extremitäten werden bei den späteren Häutungen umgestaltet, in manchen Fällen werden sie eingeschmolzen (d. h. sie sind bei den Larven, aber nicht bei adulten Tieren vorhanden). Bei den Krebstieren kann das Tier im Laufe einer langen Entwicklung mit zahlreichen Häutungen so nacheinander mehrere anatomisch und funktional völlig unterschiedliche Larvenformen mit unterschiedlicher Lebensweise durchlaufen. Diese für die Arthropoden ursprüngliche Entwicklung, bei der die Segmentzahl des adulten Tiers nach und nach erreicht wird, wird als Anamorphose (oder Anamerie) bezeichnet. Im ursprünglichsten Fall wird bei jeder Häutung genau ein Segment hinzugefügt. Bei vielen Gliederfüßern mit anamorpher Entwicklung können aber in einem Schritt gleich mehrere Segmente neu entstehen.
Bei zahlreichen Gruppen wird diese ursprüngliche Entwicklung dadurch abgewandelt, dass die frühen Entwicklungsstadien bereits im Ei durchlaufen werden, so dass das erste frei lebende Larvenstadium bereits in seiner Entwicklung fortgeschrittener ist (auch frei lebende Naupliuslarven vieler Krebstiere nehmen keine Nahrung auf). Dass es sich dabei um abgewandelte Formen handelt, kann sich zum Beispiel daran zeigen, dass der Embryo im entsprechenden Stadium eine Kutikula abscheidet. Schließlich ist es in zahlreichen Gruppen unabhängig voneinander dazu gekommen, dass alle Larvenstadien im Embryo durchlaufen werden, oder die Entwicklung wurde vollkommen abgewandelt, so dass keine Spur dieser Stadien übrigblieb. In diesen Fällen schlüpft aus dem Ei ein Jungtier, das in seiner Gestalt bereits dem Adultus weitgehend entspricht. Dieser Fall ist bei landlebenden Arthropoden die Regel, da dem Nauplius oder Protonymphon entsprechende Stadien hier nicht lebensfähig wären, dies gilt z. B. auch für die zu den Krebstieren gehörenden Landasseln. Besitzt die erste frei lebende Larve bereits alle Segmente des adulten Tiers, spricht man von Telomorphose (oder Telomerie). Die Entwicklung ohne (primäre) Larvenstadium wird direkte Entwicklung genannt. Larven, die dem adulten Tier in seiner Organisation weitgehend entsprechen, zeigen zum Beispiel auch die primär marinen Pfeilschwanzkrebse, die zu den Chelicerata gehören. Direkte Entwicklung ist typisch z. B. für alle echten Spinnen und die ursprünglicheren hemimetabolen Insekten. Bei den Tausendfüßern und bei den zu den Sechsfüßern gehörenden Beintastlern (Protura) entspricht das geschlüpfte Tier in Gestalt und Lebensweise weitgehend dem Adultus, es werden aber bei späteren Häutungen noch weitere Segmente gebildet.
Eine vollkommen abgewandelte Entwicklung zeichnet die holometabolen Insekten aus. Bei ihnen ist es sekundär wieder zur Ausbildung einer Larve gekommen, die eine vollkommen andere Gestalt und Lebensweise als die Imago aufweist. Die Herkunft dieser Larvenstadien ist nicht vollkommen geklärt, möglicherweise geht sie auf ein als Prolarve bezeichnetes Stadium zurück, das bei vielen hemimetabolen Insekten zwischen den Embryo und das eigentliche erste Larvenstadium eingeschoben ist. Das Gewebe der holometabolen Larve wird in einem Ruhestadium, der Puppe, weitgehend aufgelöst und die Imago aus knospenartigen Anlagen (den Imaginalscheiben) völlig neu gebildet.
Mit nur wenigen Segmenten (Anamorphose) schlüpfen
aus und erlangen ihre endgültige Segmentzahl erst nach dem Schlüpfen.
Mit der vollen Segmentzahl (Telomorphose) verlassen das Ei
Bei den Insekten kommen alle Übergänge vor zwischen Formen, die bereits im Embryo alle oder fast alle Segmente gleichzeitig anlegen („Langkeim“), und solchen, bei denen die Anamorphose bei der Embryonalentwicklung im Ei beibehalten worden ist („Kurzkeim“).
Die meisten Gliederfüßer sind kleine Tiere. Obwohl sie sich im Größenspektrum mit den Wirbeltieren überschneiden, sind sie weit überwiegend deutlich kleiner als diese. Wodurch die Körpergröße der sonst so vielgestaltigen und erfolgreichen Arthropoden nach oben begrenzt wird, ist eine alte Streitfrage. Eine Theorie geht davon aus, dass sich die wesentliche Begrenzung aus dem Außenskelett ergibt. Zwar ist ein auf Außenskelett beruhender Körperbau prinzipiell wohl zu beinahe jeder Größe imstande, zumal tiefe Einsenkungen (Apodeme genannt) als Muskelansatzstellen ins Innere verlagert sind. Achillesferse des Außenskeletts ist aber die Häutungsphase. Vor allem landlebende Riesen-Arthropoden würden während der Häutung unter ihrem eigenen Körpergewicht buchstäblich zerfließen. Eine weitere Theorie sieht im offenen Blutkreislauf und dem gegenüber den Wirbeltieren weniger leistungsfähigen Atmungssystem die wesentliche Begrenzung. Ein Argument für diese Ansicht ist, dass Rieseninsekten mit gegenüber rezenten Formen deutlich vergrößertem Körper in Perioden des Paläozoikums auftraten, in denen der Sauerstoff-Partialdruck der Atmosphäre gegenüber heute deutlich höher war.[32] Landlebende Gliederfüßer atmen über luftgefüllte Röhren (Tracheen), während das Blut für den Sauerstofftransport zwar nicht bedeutungslos, aber viel weniger wichtig als bei den Wirbeltieren ist. Entgegen mancher Ansicht ist der Lufttransport in den Tracheen auch nicht rein passiv, sondern die Tiere können sie gezielt zusammenpressen und damit die Atmung deutlich beschleunigen.[33][34] Dennoch ist vermutlich bei steigender Körpergröße ein überproportionales Ansteigen des Tracheen-Durchmessers erforderlich, der Effekt wird besonders in den Beinen deutlich.[35] Demzufolge ist irgendwann nicht mehr genügend Platz für andere lebenswichtige Organe. Gleichzeitig lässt der offene Blutkreislauf keinen wesentlichen Blutdruckanstieg zu, so dass der Sauerstofftransport über das Blut nur schwer optimiert werden kann. Eine dritte Theorie nimmt an, dass wegen der nicht von Schwannschen Zellen umhüllten, nackten Nervenfasern der Gliederfüßer bei höherer Körpergröße die Reaktionsgeschwindigkeit zu gering ist. Allerdings sind Myelinhüllen auch bei Krebstieren nachgewiesen worden.[36] Außerdem besitzen Gliederfüßer Riesenfasern, die für lebenswichtige Bewegungen (z. B. Fluchtreflexe) genauso hohe Übertragungsgeschwindigkeiten wie bei Wirbeltiernerven erreichen können. Möglicherweise ist es einfach so, dass durch das Vorhandensein der Wirbeltiere eine Vergrößerung keine ökologischen Vorteile mehr bietet, weil der zur Verfügung stehende Nischenraum bereits gefüllt ist. Eine Vergrößerung des Körpers bietet dabei durchaus nicht nur Vorteile. Erhebliche Nachteile sind z. B. Verlängerung der Entwicklungszeit (und damit längere Generationsdauer), höherer Nahrungsbedarf durch erhöhten Metabolismus und damit erhöhten Ruhestoffwechsel, erhöhtes Risiko, Prädatoren zum Opfer zu fallen. Ob es so etwas wie eine für den jeweiligen Bauplan optimale Körpergröße gibt und wo diese ggf. für die Gliederfüßer liegen würde, ist eine offene Streitfrage innerhalb der Wissenschaft.[37] Es fällt aber auf, dass es mehr mittelgroße Gliederfüßer mit Körpergrößen im Zentimeterbereich gibt als sehr große und sehr kleine.
Der größte bekannte lebende Arthropode ist eine im Meer lebende Krabbe, die Japanische Riesenkrabbe. Der Palmendieb (Birgus latro) ist mit einer Körperlänge von bis zu 40 cm und einer Masse von bis zu 4 kg der größte landlebende Vertreter der Arthropoden. Besonders kleine Gliederfüßer (um 0,1 Millimeter) gibt es u. a. innerhalb der Milben, der Springschwänze und der Ruderfußkrebse. Die kleinsten Insekten (parasitische Hautflügler) sind mit 0,15 mm kaum größer. Bei diesen Zwergformen sind viele Organsysteme reduziert. Auch die Segmentierung des Körpers und der Beine wird meist undeutlich.
Gliederfüßer bewohnen nahezu alle bekannten Lebensräume und besetzen dort die unterschiedlichsten ökologischen Nischen. Arthropoden können deshalb als die erfolgreichste Tierordnung der Erde gelten.
Die ursprünglichen Arthropoden lebten wie die Trilobiten im Meer. Von den heute lebenden (rezenten) Gruppen besitzen die Krebstiere die meisten marinen Vertreter. Bei den Cheliceraten sind die Asselspinnen und die Pfeilschwanzkrebse alle marin, aus den anderen Gruppen nahezu keine Art (Ausnahme: wenige marine Milben[38]). Weder Tausendfüßer noch Sechsfüßer haben echte marine Vertreter hervorgebracht, einige wenige Insektenarten wie die Zuckmücke Clunio sind nur bis ins Gezeitenwatt vorgedrungen.
Im Süßwasser leben ebenfalls zahlreiche Krebstiere, wobei es Gruppen gibt, die im Süßwasser weitaus artenreicher als im Meer sind, wie z. B. die Kiemenfußkrebse. Unter den Kieferklauenträgern sind die Milben mit den Süßwassermilben sekundär ins Süßwasser vorgedrungen, d. h. ihre Vorfahren waren landlebend. Auch die zahlreichen im Süßwasser lebenden Insektenarten gehen auf landlebende Vorfahren zurück. Dies zeigt sich z. B. daran, dass sie zur Atmung ein Tracheensystem besitzen, während die primär wasserlebenden Krebse mit Kiemen atmen. Viele wasserlebende Insektenlarven wie die Larven der Eintagsfliegen oder der Kleinlibellen haben sekundär wieder Kiemen entwickelt, die aber von (luftgefüllten) Tracheen durchzogen sind. Viele Insektenlarven nehmen Sauerstoff nur durch die Haut auf, einige besitzen zur Vergrößerung der Oberfläche mit Hämolymphe gefüllte einfache Schlauchkiemen.
Viele im Süßwasser lebende Gliederfüßer sind tatsächlich luftatmend. Sie besitzen dann ein Tracheensystem mit offenen Stigmen. Manche können mit schnorchelartigen Atemrohren an der Wasseroberfläche „Luft tanken“ wie die Larve der Mistbiene. Andere leben in einer Luftblase wie die Wasserspinne oder halten eine Lufthülle an der Körperoberfläche fest wie der Gelbrandkäfer.
Landlebend sind die allermeisten Insekten, beinahe alle Spinnentiere und alle Tausendfüßer. Von den Krebsen sind einige Entwicklungslinien unabhängig voneinander sekundär zum Landleben übergegangen. Am besten an das Landleben angepasst sind dabei die Landasseln, während landlebende Zehnfußkrebse (wie die Landeinsiedlerkrebse und Landkrabben) und Flohkrebse zumindest zur Fortpflanzung ins Wasser zurückkehren müssen.
Die Arthropoden sind neben den Wirbeltieren die einzige Gruppe des Tierreichs, die Organismen hervorgebracht hat, die auf Beinen laufen (d. h. sich nicht nur vorwärts stemmen). Die Funktionsweise des Arthropodenbeins ist dabei bei wasser- und landlebenden Formen genau entsprechend, es ist z. B. nicht möglich, anhand der Laufbeine zu entscheiden, ob eine fossil erhaltene Art wasser- oder landlebend war. Nur wenige Gruppen haben sekundär die Beine wieder aufgegeben, weit überwiegend parasitische Formen. Bei vielen Krebstieren besitzen die Extremitäten gleichzeitig zwei Funktionen: Sie dienen sowohl der Fortbewegung wie dem Nahrungserwerb. Bei anatomisch stärker abgeleiteten, moderneren Formen sind diese Funktionen meist getrennt, d. h. verschiedene Extremitäten sind auf die eine oder die andere Funktion spezialisiert. Die meisten wasserlebenden Entwicklungslinien haben schwimmende Formen hervorgebracht. Schwimmorgane sind in der Regel umgebildete und spezialisierte Extremitäten. Einige Entwicklungslinien der Krebstiere haben konvergent Blattbeine entwickelt, die nicht mehr zum Laufen geeignet sind. Zum Zooplankton gehörende Formen (neben Krebstieren einige Milben und Insektenlarven) sind Dauerschwimmer. Von extrem hoher Bedeutung für die Radiation der Insekten war die Entwicklung von Flügeln. Der Erwerb der Flugfähigkeit war vermutlich das Schlüsselmerkmal, das die Fluginsekten zur weitaus artenreichsten Organismengruppe des gesamten Planeten machte. Im Gegensatz zu Dauerschwimmern gibt es dabei keine Dauerflieger. Allerdings können z. B. viele Libellenarten mit ihren Beinen nicht laufen, sie dienen bei ihnen ausschließlich zum Festhalten und zum Nahrungserwerb. Häufig nicht beachtet wird, dass auch die Webspinnen in gewisser Weise eine Flugfähigkeit entwickelt haben. Der Flug vor allem von Jungspinnen an einem selbst abgegebenen Faden Spinnenseide („ballooning“) ermöglicht ihnen eine rasche Ausbreitung in alle geeigneten Lebensräume. Einige Entwicklungslinien haben besondere Arten der Fortbewegung entwickelt. Viele Arten wie Springspinnen, Flöhe und Springschwänze können sehr gut springen. Selten sind Formen, die wie Schnecken kriechen, wie etwa die Larven der Schneckenspinner.
Viele Krebse, die meisten Spinnen und diverse Insekten leben räuberisch.
Viele Arten saugen Blut, wie die Hirschlausfliege bei Hirschen, die Rote Vogelmilbe bei Vögeln, Fischläuse bei Fischen oder die Varroamilbe bei der Westlichen Honigbiene. Die Larve der Roten Samtmilbe saugt sich ähnlich wie der Holzbock (Ixodes ricinus) beim Menschen an Insekten fest.
Milbenbefall des Ohrs führt zu Ohrräude. Haarbalgmilben leben im Haarbalg von Säugetieren und Federmilben auf den Federn der Vögel. Viele dieser Arten sind für den Wirt unschädlich, andere rufen die Räude hervor.
Dasselfliegen leben im Magen, Nasen- und Rachenraum und/oder der Lunge von Säugetieren.
Schlupfwespen sind innere Parasiten, die zum Tod des Opfers führen, deshalb nennt man sie Parasitoide.
Diverse Arten wie Spinnmilben und Blattläuse saugen an Pflanzen.
Viele Schmetterlinge, die Honigbienen und diverse andere Tiere leben vom Nektar der Blütenpflanzen.
Larven von Schmetterlingen und Käfern fressen die Blätter von Pflanzen. Gallwespen legen ihre Eier in das Innere der Pflanzen.
Staatenbildende Insekten haben eine komplexe geruchliche Kommunikation, was einige Arten ausnutzen, um ungefährdet in die Staaten anderer Insekten eindringen zu können.
Der Totenkopfschwärmer ernährt sich von Honig direkt aus den Bienenwaben – von den Bienen wird er vermutlich wegen einer geruchlichen Tarnkappe nicht angegriffen.
Die Schlupfwespenart Ichneumon eumerus ist ein Hyperparasit: Sie parasitiert den Kreuzenzian-Ameisenbläuling (Maculinea rebeli), der selber bei Ameisen parasitiert.
Manche Ameisen züchten Pilze wie die Blattschneiderameisen oder halten Blattläuse als Haustiere wie die Rote Waldameise (Formica rufa) oder die Schwarze Wegameise (Lasius niger). Zu den Bläulingen haben Ameisen sehr unterschiedliche und interessante Beziehungen, die von Symbiose bis zu Parasitismus reichen können.
Die Systematik der Arthropoden beruht traditionell vor allem auf dem äußeren Körperbau mit der Abfolge der Segmente und Gliedmaßen. Seit Ende der 1990er Jahre werden verstärkt Merkmale des Nervensystems, molekulare Stammbäume und entwicklungsgenetische Merkmale in die Betrachtung einbezogen, daneben weitere Merkmale wie z. B. der Feinbau der Spermien. Zahlreiche sehr gut abgesichert geglaubte Gruppierungen des traditionellen Systems mussten daraufhin modifiziert oder aufgegeben werden. Wichtigstes Resultat der „neuen“ Systematik ist die Zweiteilung der Protostomia in zwei Großgruppen Ecdysozoa und Lophotrochozoa. Wenn dieses inzwischen auf verschiedenen Wegen gut abgesicherte Modell zutrifft, können die Ringelwürmer (Annelida) nicht die Schwestergruppe der (Pan-)Arthropoden sein. Diese Gruppierung (Gliedertiere oder Articulata) galt vorher als bestens abgesichert. Neuere Resultate deuten darauf hin, dass sowohl die Bildungsweise der Kopfsegmente[39] wie auch die Entwicklung der Körperanhänge (Beine bzw. Parapodien) in beiden Gruppen sehr ähnlich gesteuert wird. Da eine konvergente Bildung dieser komplizierten Abfolge nicht sehr wahrscheinlich erscheint, wäre es möglich, dass bereits der gemeinsame Vorfahre der Ecdysozoa und Lophotrochozoa („Ur-Protostomier“) oder sogar dessen gemeinsamer Vorfahre mit den Deuterostomia („Ur-Bilaterier“) segmentiert war. Problematisch an dieser Annahme ist es allerdings, dass dann in den zahlreichen nicht-segmentierten Tierstämmen die Segmentierung viele Male unabhängig voneinander sekundär aufgegeben worden sein müsste. Die heutigen Erkenntnisse der Forschung sind für eine Beantwortung dieser Frage nicht ausreichend.
Innerhalb der Ecdysozoa ist die Schwestergruppe der Panarthropoda ein Taxon aus wurmartigen Tieren mit fester, periodisch gehäuteter Körperhülle und einem „Gehirn“, das ringförmig den Verdauungstrakt umgibt, nach diesem Merkmal Cycloneuralia benannt. Eine weitere Gemeinsamkeit ist ein in fast allen Gruppen ausstülpbarer Mundabschnitt. Ungeklärt ist das Verhältnis zwischen den drei Gruppen, die gemeinsam die Panarthropoda ausmachen: Bärtierchen, Stummelfüßer, Gliederfüßer. Obwohl inzwischen die meisten Taxonomen ein Schwestergruppenverhältnis der Gliederfüßer und der Stummelfüßer für wahrscheinlicher halten,[40] hat auch die Gruppierung der Gliederfüßer mit den Bärtierchen weiter zahlreiche Anhänger. Die (traditionelle) Zusammenfassung der Bärtierchen und Stummelfüßer als gemeinsame Schwestergruppe wird weniger oft genannt, ist aber ebenfalls nicht auszuschließen.
Den neueren Untersuchungen zufolge gliedern sich die Gliedertiere in folgende monophyletische Gruppen:
Die Gruppierung der Sechsfüßer mit den Krebstieren ist inzwischen durch zahlreiche voneinander unabhängige molekulare Stammbäume recht gut abgesichert und wird auch durch die Anatomie des Nervensystems und des Auges gestützt. Obwohl auch etliche Studien die Monophylie der Krebstiere unterstützen, erscheint es nun am wahrscheinlichsten, dass diese in Bezug auf die Sechsfüßer paraphyletisch sind, das bedeutet, die Sechsfüßer sind mit einigen Krebsordnungen näher verwandt als diese untereinander. Die Sechsfüßer wären danach so etwas wie eine zum Landleben übergegangene Linie der Krebstiere. Welche Krebstiere Schwestergruppe der Sechsfüßer sein könnten (d. h. unter den Krebstieren am nächsten mit ihnen verwandt), ist noch nicht geklärt. Mögliche Kandidaten sind die „höheren“ Krebse (Malacostraca), die Kiemenfußkrebse (Branchiopoda) bzw. die urtümlichen Remipedia[41] und Cephalocarida. Die früher favorisierte Zusammenfassung der Sechsfüßer mit den Tausendfüßern („Atelocerata“ oder „Tracheata“) erscheint inzwischen kaum noch plausibel. Die wichtigsten Gemeinsamkeiten beider Gruppen sind vermutlich durch die beiden gemeinsame landlebende Lebensweise entstandene Konvergenzen. Das wichtige anatomische Argument, der in beiden Gruppen ähnliche Aufbau des Kopfes mit Beißmandibeln und Rückbildung der zweiten Antennen, ist allein nur eine schwache Begründung. Zwischenzeitlich aufgekommene Vermutungen, die Collembola (Springschwänze) könnten unabhängig von den anderen Hexapoda von Krebstieren abstammen, wodurch auch die Hexapoda gegenüber den Crustacea paraphyletisch wären, haben sich nicht bestätigt.
Noch umstritten ist die Stellung der Tausendfüßer. Aufgrund von morphologischen Befunden wurde eine Zusammenfassung mit den Tetraconata zu einer Mandibulata genannten Gruppe für gut abgesichert gehalten. Völlig überraschenderweise ergaben einige molekulare Stammbäume einen engeren Zusammenhang mit den Chelicerata;[42] die resultierende Gruppe, für die es zunächst überhaupt kein morphologisches Argument gab, wurde deshalb „Paradoxopoda“ genannt (auch: Myriochelata). Neuere molekulare Stammbäume auf verbesserter Datengrundlage stützen wieder überwiegend die Mandibulata-Hypothese, die Befunde sind aber keinesfalls eindeutig. Zusätzlich wurde (aufgrund der embryonalen Entwicklung des Nervensystems) auch ein morphologisches Argument für die Paradoxopoda vorgebracht,[43] während andere neuroanatomische Studien die Mandibulata stützen.[44] Obwohl die Sachlage damit keinesfalls als geklärt gelten kann, stützen neuere Daten überwiegend die Mandibulata. Inzwischen wurde vermutet, dass die auf Basis der ribosomalen RNA aufgestellten Stammbäume – die die wesentliche Grundlage für die Paradoxopoda-Hypothese bildeten – nicht geeignet sind, das Problem zu lösen.[45]
Weitere Hypothesen sind nur noch von historischem Interesse, z. B. die Zusammenfassung aller Gruppen mit einästigen Beinen (d. h. ohne Spaltbein) als „Uniramia“ oder die (vor allem bei Paläontologen beliebte) Vereinigung der Crustacea mit den Chelicerata („Schizoramia“).
Damit ergibt sich folgende Hypothese über die Verwandtschaftsverhältnisse (ohne fossile Gruppen):[46][47][48][49][50]
Gliederfüßer (Arthropoda) Mandibeltiere (Mandibulata)Tausendfüßer (Myriapoda)
Krebstiere (Crustacea)
Sechsfüßer (Hexapoda, unter anderem Insekten)
Kieferklauenträger (Chelicerata) (Pfeilschwänze und Spinnentiere)
Fossilien von Stammgruppenvertretern der Gliederfüßer sind seit dem frühesten Kambrium nachgewiesen. Ihre tatsächliche Entstehung fällt daher mit hoher Wahrscheinlichkeit ins ausgehende Präkambrium (Ediacarium), wenn auch aus dieser Zeit noch keine überzeugenden Fossilien vorliegen. Da auch die frühesten Fossilien von Spuren ins älteste Kambrium fallen, ist eine wesentlich frühere Entstehung des Stamms unwahrscheinlich. Im frühkambrischen Maotianshan-Schiefer aus China oder im mittelkambrischen Burgess-Schiefer aus Kanada sind Arthropoden bereits formenreich und differenziert mit zahlreichen Arten und Individuen vertreten.
Stammgruppe der Panarthropoden sind mit hoher Wahrscheinlichkeit die Lobopoden (oder auch Tardypolypoden), eine im Ordovizium ausgestorbene Gruppe fossiler „Würmer mit Beinen“, die in ihrem Organisationsniveau (nicht unbedingt in ihrer tatsächlichen Abstammung) an die rezenten Onychophora erinnern.[51][52] Primitive Formen wie Aysheaia zeigen einen gegliederten Körper mit repetitiven (gleichförmig wiederholten) Strukturen wie den geringelten Beinen, aber ohne erkennbare echte Segmente. An einem ungegliederten Kopf sitzen seitlich zwei längere Anhänge mit steifen Borsten, die vermutlich im Dienst der Nahrungsbeschaffung standen; diese sind die einzigen differenzierten Körperanhänge. Sklerotisierte Platten traten weder am Rumpf noch an den Beinen auf. Inzwischen ist ein ganzer „Zoo“ von teilweise bizarr abgewandelten Lobopoden bekannt, die etwa zeitgleich lebten. Einige von ihnen zeigen sklerotisierte Platten oder Dornen und einen primitiv segmentierten Kopf mit ein oder zwei Paar Anhängen (wie Onychodictyion oder die berühmte Hallucigenia). Eine Art, das „Kaktustier“ Diania cactiformis, wies Extremitäten mit gelenkig verbundenen Skleriten ähnlich dem Bein der Arthropoden auf.[53]
Fossilien, die als Stammgruppenvertreter der echten Arthropoden interpretiert werden wie z. B. Fuxianhuia, Canadaspis oder Shankouia, weisen gegenüber den Lobopoden eine Reihe neuer Merkmale auf.[55][56] Die Lage des Mundes verschiebt sich vom vorderen Körperende etwas bauchwärts (ventral). Am Vorderende sitzt ein von einem dorsalen Sklerit (Tergit) geschützter Kopf aus zwei Segmenten, deren erstes ein Paar gestielter Augen trägt, am zweiten sitzen sklerotisierte, gegliederte Anhänge, die antennen- oder beinähnlich ausgebildet sein können. Der übrige Körper ist in sich recht gleichförmig in Segmente gegliedert, die jeweils durch eine dorsale, stark sklerotisierte Platte (Tergit) geschützt sind. An der Unterseite sitzen stummel- oder blattförmig ausgebildete Beine, die meist nicht oder nur schwach sklerotisiert waren und die das Tier in einer Art schwimmend-laufender Bewegung über den Grund transportieren konnten. Außen an den Beinen sitzende blatt- oder lappenförmige Anhänge dienten vermutlich als Kiemen. Weiter fortgeschrittene Formen weisen einen Kopfschild auf, der neben dem Augen- und dem ersten Extremitätensegment drei weitere Segmente bedeckt, ohne dass deren Extremitäten sonderlich von den übrigen Rumpfgliedmaßen abweichen würden. Dieser fünfsegmentige Kopf wird von vielen Forschern als Grundbauplan der (Eu-)Arthropoden angesehen. Ob entsprechend gebaute Fossilien als Stammgruppenvertreter der Arthropoden oder als bereits fortgeschrittenere Vertreter einer bestimmten Arthropodenlinie wie der Crustacea oder der Chelicerata anzusehen sind, hängt in kritischer Weise von der Interpretation der vorderen Extremitäten und ihrer Segmente ab. Die meisten Forscher sehen in den „großen Anhängen“ zahlreicher Fossilien wie Leanchoilia oder Yohoia (beinartigen, stark beborsteten Mundanhängen, die vermutlich zum Ergreifen von Beutetieren dienten) homologe Bildungen der Cheliceren oder der (ersten) Antennen der lebenden Gliederfüßer. Folgt man dieser Deutung, können die meisten Formen als Stammgruppenvertreter der Chelicerata aufgefasst werden. Einige Forscher homologisieren diese Anhänge allerdings mit den Antennen der Onychophora, die am vordersten Kopfsegment (dem Segment, das den vordersten Gehirnabschnitt, das Protocerebrum, hervorbringt) sitzen, den Antennen oder Cheliceren der Arthropoden also nicht direkt entsprechen; dementsprechend gehören dieselben Fossilien ihrer Ansicht nach in die Stammgruppe aller Arthropoden.[57] Kylinxia war ein fünfäugiges ~5 cm großes garnelenartiges Tier, welches vor 518 Mio. Jahren lebte, mit einigen Unterscheidungsmerkmalen – wie den vorderen Anhängen – den "Missing Link" zwischen Anomalocaris (Radiodonta) und Deuteropoda (echten Gliederfüßern) schließt und damit laut einigen Wissenschaftlern der erste Gliederfüßer sein könnte bzw. deren evolutionären Wurzel zuzuordnen ist.[54][58]
Die Stammgruppe der Krebstiere ist bereits vom unteren Kambrium an (also parallel zu den Lobopoden) fossil überliefert. Besonders aussagekräftig sind körperlich erhaltene, phosphatisierte Fossilien (sogenannte „Orsten“), die allerdings nur mikrometer- bis millimetergroß werden, aber feinste Einzelheiten des Körperbaus erkennen lassen. Stammgruppenvertreter und Vertreter der modernen Krebse (der „Kronengruppe“) sind sich morphologisch sehr ähnlich, unterscheiden sich aber vor allem im Aufbau des Kopfes, der aus unterschiedlich vielen Segmenten bestehen kann. Die Antennenpaare von Arten wie Oelandocaris[59][60] dienten ihrem Aufbau nach wohl ebenso wie die „großen Anhänge“ primär der Nahrungsaufnahme.
Neben den Formen, die zu den modernen Gliederfüßern hinführen, existieren vom Kambrium an zahlreiche weitere Arthropodenlinien, die ohne moderne Nachkommen ausgestorben sind. Die berühmteste dieser Linien sind die Trilobiten. Bevor die reichen kambrischen Fossillagerstätten mit erhaltenen Weichteilen entdeckt worden waren, waren die Trilobiten die einzige fossile Quelle für die frühe Arthropodenentwicklung, weshalb sie die früheren Vorstellungen über die Stammgruppe der Gliederfüßer stark beeinflusst haben. Heute gelten die Trilobiten als bereits stark abgeleitete und morphologisch in vieler Hinsicht fortgeschrittene Sonderformen. Ihre traditionelle Zusammenfassung mit den Cheliceraten in ein Taxon Arachnomorpha wird neuerdings mit guten Argumenten bezweifelt.[61] Eine weitere berühmt gewordene Seitenlinie sind die Anomalocarididen. Anomalocaris zählt zu den berühmtesten Fossilien des Burgess-Schiefers, seine Verwandten waren mit Körpergrößen bis zwei Meter die größten Räuber im kambrischen Meer. Eine weitere Seitenlinie, die Thylacocephala, starben am Ende der Kreidezeit aus, sie sind z. B. in den Plattenkalken von Solnhofen gefunden worden. Kambrische Vorläufer wie Isoxys zählten damals zu den häufigsten frei schwimmenden Gliederfüßern im Meer.[62] Eine weitere Gruppe, die Euthycarcinoidea, sind trotz gut erhaltener Fossilien vollkommen rätselhaft geblieben und wurden mit so ziemlich jeder Verwandtschaftsgruppe der Arthropoden in Verbindung gebracht, ohne dass ihre tatsächliche Verwandtschaft geklärt werden konnte.[63] Wenn die Interpretation in Amerika gefundener kambrischer Spurenfossilien richtig ist, wären sie die ersten landlebenden Arthropoden gewesen.
Fossilien der primär landlebenden Hexapoda und Myriapoda liegen zwar in großer Zahl und teilweise exzellenter Erhaltung vor, sie werfen aber nur wenig Licht auf die Entstehung dieser Klassen. Die ältesten Fossilien beider Gruppen stammen aus der gleichen Lagerstätte, den devonischen Ablagerungen von Rhynie, Schottland. Die hier gefundenen Tiere waren bereits landlebend und besaßen einen im Großen und Ganzen recht modernen Körperbau. Fossilien von Stammgruppenvertretern (z. B. marinen Formen) sind unbekannt, alle entsprechend gedeuteten Fossilien sind entweder fragwürdig oder in ihrer Einordnung sogar widerlegt worden.
Die für Menschen vorrangige Rolle von Gliederfüßern liegt wohl in der Landwirtschaft, da sie eine herausragende Rolle bei der Bestäubung von Pflanzen und damit einem Gutteil unserer Nahrungsmittel spielen. Einige Arten werden selbst gegessen, wie beispielsweise der Hummer und die Nordseegarnele, in vielen Kulturen aber auch Insekten (Entomophagie). Die Honigbiene versorgt uns mit Honig. Diverse Gliederfüßer dienen als Nahrung für Speisefische. Andererseits gibt es unter den Gliederfüßern Agrarschädlinge wie die Blattlaus und Vorratsschädlinge wie den Mehlwurm, die sich von unseren Nahrungsmitteln ernähren.
Die Raupen des Seidenspinners produzieren beim Verpuppen Seide, die zur Herstellung von Stoffen verwendet wird. Im Gegensatz dazu fressen die Raupen der Kleidermotte Stoffe aus Wolle.
In der Forstwirtschaft treten Gliederfüßer als wichtige Holzschädlinge in Erscheinung, aber auch als deren Feinde und deshalb als Nützlinge. Holzschädlinge betreffen auch den Haus- und Möbelbau.
Während Gliederfüßer gewöhnlich nicht groß genug sind, um dem Menschen Verletzungen zuzufügen, die allein durch ihre Größe gefährlich sind, sind doch einige ihrer Stiche und Bisse sehr unangenehm, da die Tiere gleichzeitig Gift in der Wunde hinterlassen, wie das beispielsweise bei Wespen oder Wasserspinnen der Fall ist. Diese sind aber meist nicht ernsthaft gefährlich. Seltener sind Arten, bei denen schon ein einzelner Stich oder Biss tödlich sein kann, wie bei der Schwarzen Witwe (Latrodectrus tredecimguttatus, Sterblichkeit ohne Behandlung 5 %) oder dem Skorpion Leiurus quinquestriatus. Einige Insekten und Spinnen haben Brennhaare zur Abwehr von Fressfeinden, die auch für Menschen unangenehm werden können.
Auch als Parasiten des Menschen und Krankheitsüberträger spielen Gliederfüßer eine große Rolle.
Es gibt diverse Gliederfüßer, die beim Menschen Blut saugen, wie die Kopflaus, die Filzlaus, die Bettwanze und der Gemeine Holzbock, auch Zecke genannt (Ixodes ricinus). Einige davon sind gefürchtete Krankheitsüberträger, so überträgt der Rattenfloh die Pest, die Tsetsefliege verbreitet die Schlafkrankheit und die Anophelesmücke die Malaria. Zecken übertragen Erkrankungen wie Borreliose und Frühsommer-Meningoenzephalitis (FSME). Die Krätzmilbe frisst Bohrgänge in die oberste Hautschicht von Säugetier und Mensch und ruft dadurch die Krätze hervor.
Die Gliederfüßer (Arthropoda, von altgriechisch ἄρθρον árthron, deutsch ‚Glied, Gelenk‘ und πούς poús, Gen. ποδός podós, deutsch ‚Fuß‘, gelegentlich eingedeutscht als Arthropoden) sind ein Stamm des Tierreichs. Zu ihnen gehören Insekten, Tausendfüßer, Krebstiere (z. B. Krebse, Entenmuscheln), Spinnentiere (z. B. Spinnen, Skorpione, Milben) und die ausgestorbenen Trilobiten.
Gliederfüßer sind ein sehr erfolgreicher Stamm. Rund 80 Prozent aller bekannten rezenten (nicht fossilen) Tierarten sind Gliederfüßer. Sie verteilen sich auf beinahe eine Million Insektenarten, knapp 10.000 sonstige Sechsfüßer, etwa 16.000 Tausendfüßer, gut 100.000 Spinnentiere und knapp 50.000 Krebstiere (nur Größenordnungen, genaue Zahlen schwanken je nach Autor und Jahr der Publikation). Sie entstanden vermutlich während der kambrischen Explosion, als zu Beginn des Kambriums vor etwa 540 Millionen Jahren die mehrzelligen Tierstämme plötzlich in der Fossilüberlieferung auftraten.
Die nächsten Verwandten der Gliederfüßer sind die Bärtierchen (Tardigrada) und Stummelfüßer (Onychophora), mit denen sie zu den Panarthropoda zusammengefasst werden. Einige Taxonomen nennen diese übergeordnete Gruppe Arthropoda, die „eigentlichen“ Arthropoden werden dann als „Euarthropoda“ bezeichnet.
In der traditionellen Systematik wird diese Gruppe aus morphologischen Gründen mit den Ringelwürmern zu den Gliedertieren (Articulata) zusammengefasst. Neuere molekularbiologische Daten sprechen aber für eine enge Verwandtschaft der Gliederfüßer mit den Fadenwürmern (Nematoda) und deren engster Verwandtschaft (Cycloneuralia). Diese gemeinsame Gruppierung wird Häutungstiere (Ecdysozoa) genannt.
Arthropodi ni kundi kubwa la wanyama wenye mwili wa pingili na miguu ya kuunga wasio na ugwe wa mgongo. Mifano ni wadudu, nge, buibui au kaa. Katika uainishaji wa kisayansi wamejumlishwa katika faila ya Arthropoda.
Wote huwa na kiunzi cha nje kinachoundwa na khitini.
Spishi nyingi ni ndogo, hata chini ya mm 1 lakini spishi kadhaa ni kubwa sana, hadi zaidi ya m 1. K.m. kaa wa Japani anaweza kufikia uzito wa kilogramu 15-20. Faila hii ina spishi nyingi sana zinazopatikana kila sehemu ya dunia. Vifundo vya arithropodi ni kama vipashio vilivyounganishwa kuleta mtiririko wa mwili wao.
Mwainisho wa kawaida wa arithropodi unaonyeshwa katika sanduku la uainishaji. Lakini tangu mwanzo wa karne hii miainisho mibadala imependekezwa. Pendekezo la hivi karibuni ni hili (vifundo havikupewa tabaka)[1]:
Jongoo (Myriapoda)
Arthropodi ni kundi kubwa la wanyama wenye mwili wa pingili na miguu ya kuunga wasio na ugwe wa mgongo. Mifano ni wadudu, nge, buibui au kaa. Katika uainishaji wa kisayansi wamejumlishwa katika faila ya Arthropoda.
Wote huwa na kiunzi cha nje kinachoundwa na khitini.
Spishi nyingi ni ndogo, hata chini ya mm 1 lakini spishi kadhaa ni kubwa sana, hadi zaidi ya m 1. K.m. kaa wa Japani anaweza kufikia uzito wa kilogramu 15-20. Faila hii ina spishi nyingi sana zinazopatikana kila sehemu ya dunia. Vifundo vya arithropodi ni kama vipashio vilivyounganishwa kuleta mtiririko wa mwili wao.
Arthropoda is the phylum for aw the Arthropod speshie. Arthropoda is in the domain eukaryots, the kinrick Animalia, the subkinrick Eumetazoa, an the superphylum Ecdysozoa. Arthropods haes exoskelets, lithed bodies, an lithed limms. Some o the maist weel-kent arthropods is insects, speeders, an trilobytes.
Arthropoda is the phylum for aw the Arthropod speshie. Arthropoda is in the domain eukaryots, the kinrick Animalia, the subkinrick Eumetazoa, an the superphylum Ecdysozoa. Arthropods haes exoskelets, lithed bodies, an lithed limms. Some o the maist weel-kent arthropods is insects, speeders, an trilobytes.
Subphylum Trilobitomorpha Trilobita– trilobytes (extinct) Subphylum Chelicerata Arachnida– speeders, scorpions an that Merostomata– horseshae keavies an that Pycnogonida– sea speeders Eurypterid– sea scorpions (extinct) Subphylum Myriapoda Chilopoda– Jennie-hunder-legss Diplopoda– millipedes Pauropoda Symphyla Subphylum Hexapoda Insecta– insects Entognatha Subphylum Crustacea Branchiopoda– screws an that Remipedia Cephalocarida– horseshae shrimp Maxillopoda– barnacles, fish lice an that Ostracoda– seed shrimp Malacostraca– lobsters, keavies, shrimp an thatLe arthropodos es animales invertebrate cuje appendices es articulate.
Del greco Arthro: articulation e pod: pede.
Chelicerata (include Limulidae e Arachnida)
Arthropoda utawa kéwan mawa ros-rosan iku filum kang gedhé dhéwé ing donya sato kéwan lan nyakup gegremet, angga-angga, urang, kelabang lan kéwan liyané kang mèmper.
Papat saka limang pérangan (kang urip tekan saiki) saka spésies kéwan ya iku arthropoda, kanthi gunggung punjul sakyuta spésies modhèren kang tinemu lan rékor fosil kang nganti awal Cambrian. Arthropoda biyasa tinemu ing laut, banyu tawar, dharat, lan lingkungan udara, sarta kalebu manéka wujud simbiotis lan parasit.
Mèh 90% saka kabèh jinis kéwan kang diweruhi wong iku Arthropoda. Arthropoda dianggep duwé kerabat cerak karo Annelida, contoné ya iku Peripetus ing Afrika Kidul.
Suttafila e classi:
NOTA: Quarchi mudellu di classaficazzioni si mettinu
Myriapoda and Hexapoda ntrôn suttafilu
chiamatu Uniramia.
Lu filu Arthropoda rapprisenta lu cchiù granni filu d'armali, cumprinnennu li classi Insecta (nsetti) e Arachnida (p'asempiu: taràntuli, scurpiuni, ecetra), e lu suttafilu Crustacea (p'asempiu: granci e alaustri), e àutri armali sìmili. Circa quattru d'ogni cincu speci d'armali sunnu artropoda. Sunnu cumuni tra li nvironi marini e tirrituriali, e puru s'attròvanu li formi parassìtichi. In tèrmini di grannizza, s'attròvanu di chiddi microscòpici comu plankton (~0.25 millìmitri) finu a chiddi di quarchi metru longu.
Lu nomu arthropoda signìfica "pedi agghiunciutu". L'artropoda hannu li corpi sigmintati chî appendiggi p'ogni parti. Tutti l'artropoda sunnu nvirtibrati accuperti dûn esschèlitru durissimu. Ogni tantu l'artropoda perdi la sò peddi. Sta peddi priventi l'artropoda d'assicchiari, e puru di crìsciri troppu granni.
Lu successu di l'artropoda si tratta dû esschèlitru duru, sigmintazzioni, e appendiggi agghiunciuti. St'appendiggi vennu usati pi manciari, sintiri (nnû senzu ginirali), addifènnirisi e smuvìrisi.
Os Arthropoda u artropodos (d'o griego arthron «articulación» y podos «piet») son o Phyllum d'o Reino Animal que presenta o numero d'especies més gran, son aquaticos u terrestres, holotrofos, necrofagos u parasitos, pero a més gran part herbivoros.
Presentan simetría bilateral, cuerpo segmentato, pero a diferiencia d'os anelidos os segmentos se presentan en grupos (ditos tagmas), y o celoma ye redueito en l'adulto. Os diferents grupos de segmentos u tagmas son especializaus en diferents funcions y presentan apendices diferenciaus pa una clara división d'o treballo (por eixemplo pueden estar patas, antenas, mandibulas, etcetra...).
Bi ha un exoesqueleto (cuticla) segregau por a epidermis y formau por tres napas laminadas: epicuticla, exocuticla (de proteínas y lipidos) y endocuticla, (més dobla y formada por quitina y proteínas). A quitina ye un polisacarido nitrochenau duro, liuchero, resistent y insoluble a l'augua, alcalis u acidos febles. En beluns o exoesqueleto tien entrants (apodemas), pa que s'inserten os musclos.
A cada segmento li corresponden 2 apendices pars, encara que a vegadas han desapareixiu u s'han modificau. Con un exoesqueleto asinas de resistent un apéndiz sería immóbil, pero os artropodos pueden mover-los gracias a l'articulación: os apendices son palancas vuidas con musclos voluntarios y pelos sensitivos, y encara que en a evolución s'ha produeito una diversidat a saber-lo de formas, siempre bi ha dos seccions separadas por una articulación de caracter primario: -Sección coxal. -Sección d'o cabo.
O exoesqueleto da protección, millora y locomoción, y a més proteche d'a desecación, y ixo ha permeso a muitos artropodos de colonizar o medio terreste, pero no sin inconvenients, como por eixemplo o creiximiento y a mida.
O exoesqueleto ye un obstaclo pa o creximiento, y ixo se resuelve con o creiximiento a intervalos, producindo-se l'abandono d'una cuticla viella y o endurimiento d'una cuticla nueva. Por un regular bi ha 4-6 mudas. O exoesqueleto tamién imposa limitacions a la mida, y por ixo no pueden estar muito grans.
En o exoesqueleto puet haber-ie carbonato calcico, pigmentos (melanina, carotenoides, cromoproteidos, ezt...) y pueden tener e pelos (a vegadas sensitivos), punchas y sedas.
Bi ha un sistema muscular complexo inserto en o exoesqueleto, con musclos estriaus, pa accions rapedas y lisos pa movimientos viscerals.
A sección coxal s'implanta en a membrana pleural, podendo estar simple u subdividir-se en protopodio, subcoxa y coxa, y en atros tres artillos.
As estructuras d'a base de l'apéndiz pueden adhibir-se a las pleuras como escleritos pleurals. A sobén o protopodio leva exitos (epidopoditos), por un regular respiratorios (branquias) y enditos (gnatobases si tienen función masticadora).
A sección d'o cabo (telopodito) ye més variable que l'anterior, car s'ha adaptau a muitas funcions seguntes os grupos. En beluns s'ha dividiu en 2 partz: femoral y tibial, cadaguna formata por qualques podomeros, en atros s'ha dividiu en tres partz: femoral, patelar y tibial.
A respiración puet estar cutania, branquial, traquial y dica pulmonar.
O sistema circulatorio ye derivau de l'anelidiano pero simplificau. A circulación ye ubierta, y bi ha un conchunto de lacunas plenando entresillos y senos que forman o hemocele. No bi ha vasos verdaders, de fueras d'un corazón dorsal, tubular y un sistema arterial muito redueito que no ye un poquet desembolicau que en os de respiración branquial. O corazón ye en realidat un vaso pulsatil con dos foratos d'admisión (ostiolos) con valvulas. A hemolinfa cercula dende dezaga ta debant, sale d'o corazón por una aorta anterior, y as arterias la levan a los senos d'o hemocele. A hemolinfa cercula por o hemocele entre as visceras, tornando a la cambra pericardica y entrando en o corazón por os ostiolos, que tienen valvulas que no deixan que se'n torne ta dezaga.
A sistole se consigue por contracción cardiaca, a diastole por contracción de musclos de l'aparato locomotor.
L'aparato dichestivo ye completo, a boca tien apendices, como mandibulas y maxilas, pa ensopinar y masticar, y palpos pa tastar. Os apendices d'a boca son adaptaus a diferents formas d'alimentación.
O sistema niervoso ye pareixiu a lo d'os anelidos, encara que con ganglios més grans y organos d'os sentius més desembolicaus. Ye formau por un cerebro ganglionar dorsal que conecta a traviés d'un collar periesofachico con una doble catena niervosa ventral con 1 par de ganglios por segmento, con estructura en forma d'escalera. En os crustacios, insectos y miriapodos no bi ha que una cadena ventral de ganglios, producto d'a fusión d'os dos cordons niervosos, que encara pueden diferenciar-sen.
Presentan patrons d'intelichencia complexos en os que l'aprendizache chuega un papel important. Bi ha gran desembolique d'os organos sensorials, dende sensilas con función tactil, olfativa, auditiva, quimica y d'equilibrio dica o uello multifacetau.
Por un regular dioicos, con dimorfismo sexual y organos y conductos reproductos pars, Por un regular con fecundación interna. Pueden estar oviparos u ovoviviparos. A sobén bi ha metamorfosi. Bi ha casos de partenochenesi.
A clasificación en clases ye clara pero agrupar ixas clases en superclases y subphyllum ye controvertito, sobretot por o que fa a las clases fósils. Tamién bi ha unas clases que encara que sigan a man d'os artropodos verdaders no tienen totas as caracteristicas d'os artropodos, (onicoforos por eixemplo).
Dentro d'os artropodos bi ha exemplars d'intrés alimenticio, (marisco) d'intrés economico, (por os productos que producen u por os que destruyen), y d'intrés sanitario (foco d'infeccions u por estar animals de fizada verenosa).
Muitos crustacios se minchan, especialment os decapodos, (cangrillos), tenendo importancia economica os cangrillos marins. Tamién se minchan os percebes. Minchar insectos u aracnidos no ye bien visto en a cultura occidental, pero bi ha culturas en atros países on se minchan dreitament os insectos (por eixemplo os aborichens australians se minchan as formigas-bota y en atros países se minchan os escarbachos u os grillas de mida aceptable, y dica as tarantulas.
Encara que os insectos no se minchen en Occident, si que se consume a miel que producen as abellas, estando l'apicultura una actividat antiga y muito extendita. Os crustacios planctonicos, que no se minchan tradicionalment, pueden estar indicadors de bancos de pesca.
Bi ha insectos que fizan con o suyo fizón (abellas, viespas), as aranyas y ciengarras no tienen fizón y fizan con os quelicers y forciclas respectivament. Os arraclaus encara que sigan aracnidos fizan con un fizón y no pas con os quelicers.
A fizadura d'os mosquitos ye con l'aparato bucal y no con garra fizón y puet trasmitir microbios u nematodos que causan malautías (malaria, malautía d'o suenio, elefantiasis, ecetra...). Bi ha crustacios que pueden agüespar nematodos y platelmintos parasitos de l'hombre.
En agricultura os insectos se tienen en cuenta por o poder de esdevenir plagas. Ya en a Biblia mencionan as plagas de langostos. Atros insectos que pueden estar plaga son a mosca d'a fruita, o trips, as chinches, os polecons, a procesionaria, o piollo de Sant Chusé, o escarbacho d'a pataca y as polillas. Pero l'hombre puet fer servir un insecto contra atro, (depredadors u parasitoides), como ye o caso d'as viespas.
Bi ha artropodos con importancia en a industria u artesanía producindo sustancias d'intrés comercial. As abellas a més de producir miel producen cera. Os lepidoptero Bombus produce seda (cuco d'a seda). Un producto industrial produeito por un insecto ye o carmín y, d'atras especies, se'n obtién laca. D'os meloidos se puet obtener un afrodisiaco (spanish fly).
Os insectos son eslabons intermediarios de catenas alimentarias que tienen en o suyo cabalto especies útils pa l'hombre, tamién ye important en ixo a función polinizadora d'os insectos.
Los artropòdes (arthropoda, de grèc ἄρθρον, "articulat", e ποδός, "pè") constituisson l'embrancament (filum) mai grand d'animals e incluson, entre autres, insèctes, aracnids e crustacèus. La seuna diversitat es extraordinària; mai del 80% de las espècias animalas conegudas son d'artropòdes[1], es a dire, un milion d'espècias existentas descritas e un registre fossil que remonta fins l'èra proterozoïca.
Los artropòdes se son adaptat tant al mitan aqüatic (marin e aiga doça), coma al mitans terrèstre e aerian. Incluson totes tipes de formas de vida, tanben de formas simbiòticas e parasitas.
Malgrat aquesta granda diversitat los artropòdes presentan una estructura generala fòrça omogenèa. Se caracterizan per:[2]
Ang Arthropoda ay isang phylum sa kahariang Animalia.
This list is generated from data in Wikidata and is periodically updated by Listeriabot.
Edits made within the list area will be removed on the next update!
![]() Ang lathalaing ito na tungkol sa Hayop ay isang usbong. Makatutulong ka sa Wikipedia sa nito.
Ang lathalaing ito na tungkol sa Hayop ay isang usbong. Makatutulong ka sa Wikipedia sa nito.
Arthropoda nyaéta salah sahiji tina filum sato invertebrata nu dicirian sabab boga awak nu dijieun tina ségmén nu ngahiji, jeung miboga panutup teuas kawas kerang.[3] Lancah nyaéta salah sahiji jenis arthropoda.[3] Filum arthropoda miboga anggota panglobana di alam dunya.[4] Jumlah jenis arthropoda leuwih loba ti batan total sakabéh jenis filum sato nu séjén.[4] Maranéhna hirup di darat, cai tawar, jeung laut ogé di kabéh tempat di bumi.[4]
Arthropoda miboga ciri husus mangrupa suku nu bubukuan.[4] Suku arthropoda miboga persendian nu ngamungkinkeun bisa gerak jeung nyekel dahareun.[4] salian ti éta, sato ieu ogé boga eksoskeleton (rangka luar) teuas ni dijieun tina zat kitin, eksoskeleton ieu ngajaga manéhna tina gangguan lingkungan jeung sato séjén.[4]
Le arthropodos es animales invertebrate cuje appendices es articulate.
Arthropoda nyaéta salah sahiji tina filum sato invertebrata nu dicirian sabab boga awak nu dijieun tina ségmén nu ngahiji, jeung miboga panutup teuas kawas kerang. Lancah nyaéta salah sahiji jenis arthropoda. Filum arthropoda miboga anggota panglobana di alam dunya. Jumlah jenis arthropoda leuwih loba ti batan total sakabéh jenis filum sato nu séjén. Maranéhna hirup di darat, cai tawar, jeung laut ogé di kabéh tempat di bumi.
Os Arthropoda u artropodos (d'o griego arthron «articulación» y podos «piet») son o Phyllum d'o Reino Animal que presenta o numero d'especies més gran, son aquaticos u terrestres, holotrofos, necrofagos u parasitos, pero a més gran part herbivoros.
Lu filu Arthropoda rapprisenta lu cchiù granni filu d'armali, cumprinnennu li classi Insecta (nsetti) e Arachnida (p'asempiu: taràntuli, scurpiuni, ecetra), e lu suttafilu Crustacea (p'asempiu: granci e alaustri), e àutri armali sìmili. Circa quattru d'ogni cincu speci d'armali sunnu artropoda. Sunnu cumuni tra li nvironi marini e tirrituriali, e puru s'attròvanu li formi parassìtichi. In tèrmini di grannizza, s'attròvanu di chiddi microscòpici comu plankton (~0.25 millìmitri) finu a chiddi di quarchi metru longu.
Lu nomu arthropoda signìfica "pedi agghiunciutu". L'artropoda hannu li corpi sigmintati chî appendiggi p'ogni parti. Tutti l'artropoda sunnu nvirtibrati accuperti dûn esschèlitru durissimu. Ogni tantu l'artropoda perdi la sò peddi. Sta peddi priventi l'artropoda d'assicchiari, e puru di crìsciri troppu granni.
Arthropoda utawa kéwan mawa ros-rosan iku filum kang gedhé dhéwé ing donya sato kéwan lan nyakup gegremet, angga-angga, urang, kelabang lan kéwan liyané kang mèmper.
Papat saka limang pérangan (kang urip tekan saiki) saka spésies kéwan ya iku arthropoda, kanthi gunggung punjul sakyuta spésies modhèren kang tinemu lan rékor fosil kang nganti awal Cambrian. Arthropoda biyasa tinemu ing laut, banyu tawar, dharat, lan lingkungan udara, sarta kalebu manéka wujud simbiotis lan parasit.
Mèh 90% saka kabèh jinis kéwan kang diweruhi wong iku Arthropoda. Arthropoda dianggep duwé kerabat cerak karo Annelida, contoné ya iku Peripetus ing Afrika Kidul.
Los artropòdes (arthropoda, de grèc ἄρθρον, "articulat", e ποδός, "pè") constituisson l'embrancament (filum) mai grand d'animals e incluson, entre autres, insèctes, aracnids e crustacèus. La seuna diversitat es extraordinària; mai del 80% de las espècias animalas conegudas son d'artropòdes, es a dire, un milion d'espècias existentas descritas e un registre fossil que remonta fins l'èra proterozoïca.
Los artropòdes se son adaptat tant al mitan aqüatic (marin e aiga doça), coma al mitans terrèstre e aerian. Incluson totes tipes de formas de vida, tanben de formas simbiòticas e parasitas.
D'Arthropoden (Arthropoda) oder Glidderféisser sinn e Stamm aus dem Déiereräich. Si si mat 75% vun alle bekannten Aarten de gréisste Stamm aus dem Déiereräich.
Zu den Arthropode gehéieren ausser den Insekten, Dausendféisser, Spannendéieren, Milben a Kriibsdéieren och nach manner bekannt Gruppe wéi d'Séispannen an d'Feilschwanzkriibsen.
D'Gréisst geet vu mikroskopesch klengen Déieren wéi bis hin zu Homaren déi 75 cm laang kënne ginn.
All d'Glidderféisser hunn e baussenzegt Sklett dat de ganze Kierpe ëmschléisst an dat duerch Haitungen erneiert gëtt.
D'Arthropoden (Arthropoda) oder Glidderféisser sinn e Stamm aus dem Déiereräich. Si si mat 75% vun alle bekannten Aarten de gréisste Stamm aus dem Déiereräich.
Zu den Arthropode gehéieren ausser den Insekten, Dausendféisser, Spannendéieren, Milben a Kriibsdéieren och nach manner bekannt Gruppe wéi d'Séispannen an d'Feilschwanzkriibsen.
D'Gréisst geet vu mikroskopesch klengen Déieren wéi bis hin zu Homaren déi 75 cm laang kënne ginn.
All d'Glidderféisser hunn e baussenzegt Sklett dat de ganze Kierpe ëmschléisst an dat duerch Haitungen erneiert gëtt.
La artropodos es un filo de animales ce inclui esta sufilos e clases:
Atwopòd se yon bèt kò divize an segman.[1]
BOʻGʻIMOYOQLILAR (Arthropoda) - umurtqasiz hayvonlar tipi. B.ning ajdodi tuban tuzilgan dengiz halqali chuvalchanglari hisoblansada, ularning bitta yoki bir necha umumiy ajdodlardan kelib chiqqanligi toʻgʻrisidagi muammo hozirgacha uzilkesil hal etilmagan. Eng qad. boʻgʻimoyoklilardan trilobitasimonlar quyi kembriy qatlamlaridan boshlab uchraydi. Ordovik va silur davrida B. quruqlikka chiqishgan. B.ning tanasi va oyoqlari boʻgʻimlarga boʻlingan (B. nomi ham ana shundan olingan), ikki tomonlama simmetriyali. Tanasi qattiq xitin kutikula bilan qoplangan. Xitin qoplogʻich tashqi skelet funksiyasini ham bajaradi, unga ichki tomondan muskullar birikadi. Tana boʻgʻimlari geteronom, yaʼni ularning shakli va kattaligi har xil boʻladi. B. tanasi, odatda, bosh, koʻkrak va qorin boʻlimlaridan iborat. Lekin baʼzan tananing turli boʻlimlari oʻzaro har xil tarzda birikib ketishi mumkin. B.ning bosh boʻlimi bosh boʻlagi — akrondan va 4 tana boʻgʻimlarining qoʻshilishidan hosil boʻladi. Koʻkrak va qorin boʻlimlaridagi boʻgʻimlar har xil sistematik guruhlarda bir xil boʻlmaydi. B. tanasidagi har bir boʻgʻim dastlab bir juftdan nayga oʻxshash oʻsimtaga ega boʻlgan, keyinchalik bu oʻsimtalar jagʻlar, yurish oyoqlari, suzgich organlar, soʻrgʻichlar, oʻrgimchak bezlari va boshqa organlarga aylangan yoki yoʻqolib ketgan. B. jabra, traxeya yoki oʻpka orqali nafas oladi. Birlamchi ovqat hazm qilish sistemasi ektodermadan hosil boʻlgan oldingi va keyingi ichakdan, endodermadan hosil boʻlgan oʻrta ichakdan hamda ular bilan bogʻlangan soʻlak va hazm bezlaridan iborat. Ayirish organlari kelib chiqishi va funksiyasiga koʻra, halqali chuvalchanglarning metanefridiylariga oʻxshash boʻlgan antennal, koksal va maksillar bezlar — selomoduklardan yoki ichak oʻsimtalari — malpigi naychalaridan iborat. Qon aylanish sistemasi ochiq, qon aylanish organlari har xil darajada rivojlangan. Nerv sistemasi gangliylarning qoʻshilishidan hosil boʻladigan bosh miya va qorin nerv zanjiridan iborat. Qorin nerv zanjiridagi nerv gangliylari tana boʻgʻimlarining oʻzgarishiga moye holda turli darajada konsentratsiyalashgan boʻladi. Koʻpchilik B.ning yaxshi rivojlangan sezgi organlari, jumladan murakkab fasetkali koʻzlari, har xil mexano va xemoreseptorlari, eshitish, muvozanat saqlash, ovoz chiqarish organlari boʻladi. Deyarli barcha B. ayrim jinsli. Jinsiy dimorfizm yaxshi rivojlangan. Jinsiy yoʻl bilan (baʼzan partenogenez) koʻpayadi. Koʻpchilik turlari metamorfoz orqali rivojlanadi. Qattiq tana qoplogʻichi B.ning oʻsishiga toʻsqinlik qiladi. Shuning uchun B. faqat eski xitin qoplogʻichni tashlash (tullash), yangi xitin pust hosil qilish orqali oʻsadi. Tana boʻshligʻi aralash tipda (miksotsel). Muskullari toʻptoʻp boʻlib joylashgan. Koʻpchilik B.ning yuragi naysimon, koʻp kamerali, qoni tana suyuqligiga oʻxshash boʻlib, gemolimfa deyiladi. B. tipi 4 ta kenja tip: trilobitasimonlar (qazilma holda uchraydigan yagona trilobitlar sinfi), jabra bilan nafas oluvchilar (yagona qisqichbaqasimonlar sinfi), xelitseralilar (qilichdumlilar, oʻrgimchaksimonlar sinflari), traxeyalilar (koʻpoyoqlilar, hasharotlar sinflari) ga boʻlinadi. 2 mln.ga yaqin turi bor. Yerda mavjud boʻlgan hamma yashash muhitlarini egallagan. B.ning quruqlikda yashashga va havoda uchishga moslanishi hayvonot dunyosida sodir boʻlgan eng muhim oʻzgarishlardan biri hisoblanadi. B. biosferada beqiyos katta ahamiyatga ega. Koʻpchilik ekosistemalardagi tirik organizmlarning asosiy qismini tashkil etishi, xilmaxil oziqlanishi tufayli B. tabiatda moddalar aylanishi jarayonini amalga oshiradi. B. oziq-ovqat va kiyim-kechak uchun xom ashyo manbai sifatida ham katta ahamiyatga ega. B. orasida organik qrldiqlarini oʻzlashtirib, tuproq hosil boʻlishi jarayonida faol ishtirok etadigan; odam va hayvonlarda parazitlik qiladigan va kasallik tarqatadigan; gullarni changlatib, oʻsimliklarning hosildorligini oshirishga imkon beradigan; qishloq xoʻjaligi ekinlarini yeb, zararkunandalik qiladigan turlari ham koʻp.
BOʻGʻIMOYOQLILAR (Arthropoda) - umurtqasiz hayvonlar tipi. B.ning ajdodi tuban tuzilgan dengiz halqali chuvalchanglari hisoblansada, ularning bitta yoki bir necha umumiy ajdodlardan kelib chiqqanligi toʻgʻrisidagi muammo hozirgacha uzilkesil hal etilmagan. Eng qad. boʻgʻimoyoklilardan trilobitasimonlar quyi kembriy qatlamlaridan boshlab uchraydi. Ordovik va silur davrida B. quruqlikka chiqishgan. B.ning tanasi va oyoqlari boʻgʻimlarga boʻlingan (B. nomi ham ana shundan olingan), ikki tomonlama simmetriyali. Tanasi qattiq xitin kutikula bilan qoplangan. Xitin qoplogʻich tashqi skelet funksiyasini ham bajaradi, unga ichki tomondan muskullar birikadi. Tana boʻgʻimlari geteronom, yaʼni ularning shakli va kattaligi har xil boʻladi. B. tanasi, odatda, bosh, koʻkrak va qorin boʻlimlaridan iborat. Lekin baʼzan tananing turli boʻlimlari oʻzaro har xil tarzda birikib ketishi mumkin. B.ning bosh boʻlimi bosh boʻlagi — akrondan va 4 tana boʻgʻimlarining qoʻshilishidan hosil boʻladi. Koʻkrak va qorin boʻlimlaridagi boʻgʻimlar har xil sistematik guruhlarda bir xil boʻlmaydi. B. tanasidagi har bir boʻgʻim dastlab bir juftdan nayga oʻxshash oʻsimtaga ega boʻlgan, keyinchalik bu oʻsimtalar jagʻlar, yurish oyoqlari, suzgich organlar, soʻrgʻichlar, oʻrgimchak bezlari va boshqa organlarga aylangan yoki yoʻqolib ketgan. B. jabra, traxeya yoki oʻpka orqali nafas oladi. Birlamchi ovqat hazm qilish sistemasi ektodermadan hosil boʻlgan oldingi va keyingi ichakdan, endodermadan hosil boʻlgan oʻrta ichakdan hamda ular bilan bogʻlangan soʻlak va hazm bezlaridan iborat. Ayirish organlari kelib chiqishi va funksiyasiga koʻra, halqali chuvalchanglarning metanefridiylariga oʻxshash boʻlgan antennal, koksal va maksillar bezlar — selomoduklardan yoki ichak oʻsimtalari — malpigi naychalaridan iborat. Qon aylanish sistemasi ochiq, qon aylanish organlari har xil darajada rivojlangan. Nerv sistemasi gangliylarning qoʻshilishidan hosil boʻladigan bosh miya va qorin nerv zanjiridan iborat. Qorin nerv zanjiridagi nerv gangliylari tana boʻgʻimlarining oʻzgarishiga moye holda turli darajada konsentratsiyalashgan boʻladi. Koʻpchilik B.ning yaxshi rivojlangan sezgi organlari, jumladan murakkab fasetkali koʻzlari, har xil mexano va xemoreseptorlari, eshitish, muvozanat saqlash, ovoz chiqarish organlari boʻladi. Deyarli barcha B. ayrim jinsli. Jinsiy dimorfizm yaxshi rivojlangan. Jinsiy yoʻl bilan (baʼzan partenogenez) koʻpayadi. Koʻpchilik turlari metamorfoz orqali rivojlanadi. Qattiq tana qoplogʻichi B.ning oʻsishiga toʻsqinlik qiladi. Shuning uchun B. faqat eski xitin qoplogʻichni tashlash (tullash), yangi xitin pust hosil qilish orqali oʻsadi. Tana boʻshligʻi aralash tipda (miksotsel). Muskullari toʻptoʻp boʻlib joylashgan. Koʻpchilik B.ning yuragi naysimon, koʻp kamerali, qoni tana suyuqligiga oʻxshash boʻlib, gemolimfa deyiladi. B. tipi 4 ta kenja tip: trilobitasimonlar (qazilma holda uchraydigan yagona trilobitlar sinfi), jabra bilan nafas oluvchilar (yagona qisqichbaqasimonlar sinfi), xelitseralilar (qilichdumlilar, oʻrgimchaksimonlar sinflari), traxeyalilar (koʻpoyoqlilar, hasharotlar sinflari) ga boʻlinadi. 2 mln.ga yaqin turi bor. Yerda mavjud boʻlgan hamma yashash muhitlarini egallagan. B.ning quruqlikda yashashga va havoda uchishga moslanishi hayvonot dunyosida sodir boʻlgan eng muhim oʻzgarishlardan biri hisoblanadi. B. biosferada beqiyos katta ahamiyatga ega. Koʻpchilik ekosistemalardagi tirik organizmlarning asosiy qismini tashkil etishi, xilmaxil oziqlanishi tufayli B. tabiatda moddalar aylanishi jarayonini amalga oshiradi. B. oziq-ovqat va kiyim-kechak uchun xom ashyo manbai sifatida ham katta ahamiyatga ega. B. orasida organik qrldiqlarini oʻzlashtirib, tuproq hosil boʻlishi jarayonida faol ishtirok etadigan; odam va hayvonlarda parazitlik qiladigan va kasallik tarqatadigan; gullarni changlatib, oʻsimliklarning hosildorligini oshirishga imkon beradigan; qishloq xoʻjaligi ekinlarini yeb, zararkunandalik qiladigan turlari ham koʻp.
Chat-kha tōng-bu̍t (ha̍k-miâ: Arthropoda)
Geleedpôotign (Arthropoda) zyn e stamme van koudbloedige, oengewervelde bêestn met een uutwendig skelet (exoskelet), e gesegmenteerd lyf en geleede anhangsels. Z' omvattn d' insectn, de kobbn, de duusdpôotn en de kriftachtign. 't Woord Arthropoda komt van ’t Grieks ἄρθρον árthron, "gewricht" of "geleedienge", en πούς pous (gen. podos), "voet" of "pôot".
Geleedpôotign wordn in vuuf oenderstammn ingedêeld, woavan dat er êen uutgestorvn is:
Geleedpôotign (Arthropoda) zyn e stamme van koudbloedige, oengewervelde bêestn met een uutwendig skelet (exoskelet), e gesegmenteerd lyf en geleede anhangsels. Z' omvattn d' insectn, de kobbn, de duusdpôotn en de kriftachtign. 't Woord Arthropoda komt van ’t Grieks ἄρθρον árthron, "gewricht" of "geleedienge", en πούς pous (gen. podos), "voet" of "pôot".
De Gelidpoetege (Grieks-Latiense naom Arthropoda; aander meugelike vertaoling geleipoetege) zien 'n stam vaan 't Diereriek. 't Is vaan alle stamme wiedoet de groetste, mèt bekans ein miljoen soorte bekind. De meiste gelidpoetege zien diere in de orde vaan groetde vaan 10-2 m, allewel tot väöl soorte daobove of dao-oonder zitte. Ze höbbe, aanders es väöl primitiever diere, dudelek aonwiesbare herses en hert, wie primitief ouch. De pu bestoon oet mie stökskes. De indeiling in oonderstamme is zoe:
In de jaore '70 woorte de oet 't Cambrium aofkómstege fossiele vaan de Burgess Shale perceis bestudeerd. 't Mies-veurkoumend fossiel in de Burgess Shale is Marrella, dat ieder woort ingedeild bij de trilobiete. 't Bleek evels dat Marrella in gein vaan de veer bekènde gróppe kós weure oondergebrach: Marrella heet twie paar oetsteeksele aon de kop, bei veur de moond geplaots. In modern indeilinge vaan de Gelidpoetege weurt 't daoveur dèks mèt e paar aander verwaante soorte in 'nen eigen oonderstam geplaots, de Marrellomorpha.
Girêçikpê an Artropod (Arthropoda) ango, filûma Arthropoda, ji zimanê grekî ἄρθρον arthron, "girêçik" û ποδός podos, "pê") mezintirîn filûma heywan yan caneweran e û mêş, pîrik, pezpezok û gelek girêçikpiyên din dikevine vê filûmê, gor zanyaran ku der dorî %80 ji Artropodan e.
Pygnogonida
Xiphosura
Dûpişkê reş (Scorpiones)
Amblypygi
Amblypygi
Araneae
Araneae
Opiliones
Acari
Acari
Girêçikpê an Artropod (Arthropoda) ango, filûma Arthropoda, ji zimanê grekî ἄρθρον arthron, "girêçik" û ποδός podos, "pê") mezintirîn filûma heywan yan caneweran e û mêş, pîrik, pezpezok û gelek girêçikpiyên din dikevine vê filûmê, gor zanyaran ku der dorî %80 ji Artropodan e.
D Arthropode, vo griechisch arthron ‚Glänk‘ und griechisch pous, gen. podos ‚Fuess‘)[1] si e Stamm vom Dierriich. Zu iine ghööre so underschidligi Dier wie Insekte, Duusigfüessler, Chräbsdier (z. B. Chräbs, Äntemuschle), Spinnedier (z. B. Spinne, Skorpion, Milbe) und die usgstorbene Trilobite.
D Arthropode si e seer erfolgriiche Stamm. Öbbe 80 Brozänt vo alle bekannte rezänte (nit fossile) Dierarte si Arthropode. Si verdäile sich uf fast e Million Insektenarte, knapp 10'000 sünstigi Säggsfüessler, öbbe 16'000 Duusigfüessler, guet 100'000 Spinnedier und knapp 50'000 Chräbsdier[2] (nume Gröössenornige, gnauji Zaale schwanke je noch em Autor und Joor vo dr Publikazioon). Si si vermuetlig wääred dr kambrische Explosion entstande, wo am Aafang vom Kambrium vor öbbe 540 Millione Joor die meerzellige Dierstämm blötzlig in dr Fossilüberliiferig vorchoo si.
Die nöggste Verwandte vo de Arthropode si d Bärdierli (Tardigrada) und Stummelfüessler (Onychophora). Mit dene fasst mä sä zu de Panarthropoda zämme, au wenn d Onychophora käni gliiderete Bäi häi. Es git Taxonome die säge dere übergordnete Grubbe Arthropoda, de „äigentlige“ Arthropode säit mä denn „Euarthropoda“.
In dr dradizionelle Süstematik wird die Grubbe us morfologische Gründ mit de Ringelwürmer zu de Articulata oder Gliiderdier zämmegfasst. Noch nöijere molekularbiologische Date si si aber nid äng mit dene verwandt sondern mit de Faadewürmer (Nematoda) und andere Hütigsdier (Ecdysozoa).
D Arthropode, vo griechisch arthron ‚Glänk‘ und griechisch pous, gen. podos ‚Fuess‘) si e Stamm vom Dierriich. Zu iine ghööre so underschidligi Dier wie Insekte, Duusigfüessler, Chräbsdier (z. B. Chräbs, Äntemuschle), Spinnedier (z. B. Spinne, Skorpion, Milbe) und die usgstorbene Trilobite.
D Arthropode si e seer erfolgriiche Stamm. Öbbe 80 Brozänt vo alle bekannte rezänte (nit fossile) Dierarte si Arthropode. Si verdäile sich uf fast e Million Insektenarte, knapp 10'000 sünstigi Säggsfüessler, öbbe 16'000 Duusigfüessler, guet 100'000 Spinnedier und knapp 50'000 Chräbsdier (nume Gröössenornige, gnauji Zaale schwanke je noch em Autor und Joor vo dr Publikazioon). Si si vermuetlig wääred dr kambrische Explosion entstande, wo am Aafang vom Kambrium vor öbbe 540 Millione Joor die meerzellige Dierstämm blötzlig in dr Fossilüberliiferig vorchoo si.
Die nöggste Verwandte vo de Arthropode si d Bärdierli (Tardigrada) und Stummelfüessler (Onychophora). Mit dene fasst mä sä zu de Panarthropoda zämme, au wenn d Onychophora käni gliiderete Bäi häi. Es git Taxonome die säge dere übergordnete Grubbe Arthropoda, de „äigentlige“ Arthropode säit mä denn „Euarthropoda“.
In dr dradizionelle Süstematik wird die Grubbe us morfologische Gründ mit de Ringelwürmer zu de Articulata oder Gliiderdier zämmegfasst. Noch nöijere molekularbiologische Date si si aber nid äng mit dene verwandt sondern mit de Faadewürmer (Nematoda) und andere Hütigsdier (Ecdysozoa).
Këmbënyjorët përfaqësojnë grupin më të madh të llojeve shtazore. Përfshijnë mbi 1.000.000 lloje dhe përbëjnë afro 80% të tërë faunës së përhapur në rruzulin tokësor. Supozohet se numri i tyre është më i madh, meqenëse çdo ditë zbulohen lloje të reja. Janë të përhapur në mjedisin tokësor dhe ujor. Emri këmbënyjorë rrjedh nga greqishtja arthron=nyje dhe podhos=këmbë. Për nga ndërtimi i trupit, mënyra e të ushqyerit dhe tipi i aparatit gojor, këmbënyjëtuarit ndahen në disa grupe: ndër të cilat më të njohura janë gaforret, merimangat dhe insektet që jetojnë kryesisht në mjedisin tokësor e shumë më pak në atë ujor. Këtu duhet përmendur edhe këmbënyjëtuarit më të vjetër tribolitet, që kanë pasur ndërtim më primitiv. Këta kanë jetuar në kohëra të lashta dhe numri i tyre ka qenë afro 10.000 lloje. Mjedisi jetësor i tyre ka qenë uji. Sot hasen vetëm në formë të mbeturinave (fosileve).
Gaforret janë shtazë të cilat jetojnë në ujëra të ëmbël dhe të njelmët dhe përfaqësojnë këmbënyjëtuarit e sotëm më primitivë. Për njohjen e ndërtimit dhe mënyrën e jetesës si shembull merret gaforrja e lumit. Jeton në ujëra të ëmbël. Sipërfaqja e trupit është e mbuluar me një mbështjellës të fortë i përbërë nga lënda e quajtur kitina e forcuar me lëndë gëlqerore. Ky mbështjellës i fortë, i cili mbron organet e brendshme të gaforres, paraqet skeletin e jashtëm të saj. Trupi i gaforres së lumit përbëhet prej dy pjesëve: kokëkraharorit (cefalotoraksit) dhe barkut (abdomenit). Kokëkraharori formohet me bashkimin e kokës dhe të kraharorit. Në kokë gjendet goja, dy palë antena (dy të gjata dhe dy të shkurtëra) dhe dy sy tëpërbërë. Në krarharor ka pesë palë këmbë. Pala e parë e këmbëve ka formën e mashave dhe shëben për kapjen e ushqimit, ndërsa katër palët e tjera shërbejnë për ecje. Këmbët përbëhen prej gjashtë segmenteve (unazave) të vendosura njëra pas tjetrës. Në unazën e fundit e cila formon pendën e bishtit, gjendet vrima anale. Fundi i barkut mbaron me flatrën e bishtit që shërben për notimin e gaforres. Ushqimi, siç janë krimbat, peshqit e vegjël, bretkosat dhe bimët ujore, gaforrja e kap me anë të mashave dhe e fut në gojë, ku me ndihmën e nofullave e grimcon. Gaforrja merr frymë me anë të branshive, të cila janë të përshkuara me gypa gjaku. Velëzat janë të vendosura në dy anët e trupit. Sistemi i gjakut, i cili është në anën shpinore dhe gypave të gjakut. Sistemi nervor është i vendosur në anën barkore të trupit dhe përbëhet prej ganglioneve që gjenden në çdo segment të trupit. Në majë të kokës ka dy sy të përbërë prej numrit të madh të syve të thjeshtë, ndërsa antenat shërbejnë si organe të prekjes dhe të nuhatjes. P.sh.nëse i afrohemi gaforres, ajo ikën shumë shpejt duke u fshehur nën gurë. Shumimi i gaforres bëhet me anë të vezëve. Nga vezët zhvillohen gaforret e vogla, të cilat një kohë mjaft të gjatë i ushqen dhe i mbron e ëma duke i bartur me vete. Gaforrja e lumit mund të jetojë 15 deri 20 vjet. Karakteristikë e gaforreve është xhveshja në të cilin rast hedhet mbështjellësi i jashtëm, duke u formuar një i ri. Në dete dhe oqeane jeton numër mjaft i madh i gaforreve me formë dhe madhësi të ndryshme. Ndër ato të njohura janë aragosta dhe nefropsi, të cilat përdoren edhe si ushqim i njeriut. Këto lloje jetojnë në fund të ujit të fshehua nën gurë ose nën bimë të ndryshme. duke pritur gjahun e tyre. Me të parë gjahun me shpejtësi vërsulen mbi të duke kapur me mashat e fuqishme. Me formën dhe ngjyrën e trupit i përshtaten shumë mirë mjedisit jetësor, kështu që vështirë dallohen nga armiqtë nga armiqtë e tyre.
Pos gaforres së lumit, në ujëra amullë (të qeta) jetojnë edhe pleshti i ujit (dafina) dhe ciklopët. Pleshti i ujit është një gaforre e vogël me trup të tejdukshëm, i cili lëviz duke kërcyer. Është ushqim i mirë për peshqit. Ciklopët po ashtu janë gaforre të vogla, të cilat kanë vetëm një sy. Edhe këto gaforre shërbejnë si ushqim i mirë i peshqit
Janë këmbënyjëtuar te të cilat, pos disave, trupi është i ndarë në kokëkraharor dhe bark. Kokëkraharori është me mbulesë kitinore. Në kokë nuk ka antena. Në të janë të vendosur katër palë sy të thjeshtë. Unaza e parë e kokëkraharorit ka një alë shtojca të cilat përfundojnë me nga një grep, me të cilin mbyt gjahun pasi që në të lëshon helmin nga gjëndrrat që janë në lidhje me këto shtojca. Unaza e dytë, po ashtu, ka një palë shtojca për mbajtjen e gjahut. Në kokëkraharor ka edhe katër palë këmbë të nyjëtuara me të cilat ec dhe end rrjetin e saj. Barku është i butë dhe pa këmbë. Në anën e poshtme të barkut gjenden dy vrima për frymëmarrje që çojnë deri te një hapësirë që shërben si mushkëri e përshkruar me gypa të gjakut. Nga fundi i barkut gjenden gjëndrrat që prodhojnë materialin për endjen e rrjetit. Merimangat janë shtazë grabitqare, të cilat ushqehen me miza, gretha, flutura etj., të cilat kapen në rrjetën e tyre. Kur insekti bie në rrjetën të cilën e mbaron merimanga, ai del nga vendi ku ka qenë i fshehur, mbyt gjahun me helmin që del pas thumbimit, thithë pjesët e brendshme të insektit duke lënë vetëm skeletin hitinor të tij. Merimanga ha vetëm insektet të cilët kapen në rrjetën e saj. Numri më i madh i llojeve të merimangës nuk janë të rrezikshme për njeriun, megjithatë ka edhe prej tyre të cilat janë shumë helmuese. Ndër më të njohurat te ne është merimanga me kryq, ajo e shtëpisë. Në vendet e nxehta jeton merimanga shumë helmuese e quajtur vejusha e zezë, tarantulla etj.
Këta këmbë zgatur kanë bishtin e gjatë, i cili mbaron me një thumb që përmban helm. Me anë të thumbimit mbysin insekte dhe shtazë të tjera të imëta. Jetojnë nën gurë, drunj etj. dhe natën dalin kërkojnë ushqimin. Disa lloje të akrepave, të cilët jetojnë në viset e nxehta, janë helmues dhe shumë të rrezikshme edhe për njeriun, siç janë p.sh. skorpionet.
Janë shtazë të imëta, me madhësi mikroskopike e deri në 1 cm. Këpusha trupin e ka prej një pjese të formuar me bashkimin e kokës, të kraharorit dhe të barkut. Kanë katër palë këmbë mjaft të vogla. Dallohen për kah forma e trupit dhe mënyra e jetesës. Disa prej tyre ushqehen me shtazë të vogla ose me mbeturina, ndërsa të tjerat si parazitë. Prej llojeve parazite më të njohura është këpusha e zgjebës, e cila shkakton sëmundjen e zgjebës te njeriu. Ka madhësi deri në 0,5 mm dhe jeton në brendi të lëkurës së njeriut ku femra vendos vezët e saja. Sëmundja e zgjebës karakterizohet me kruarje të madhe të lëkurës dhe është ngjitëse. Sëmundja me anë të prekjes shumë lehtë bartet nga një njeri në tjetrin. Si masë paraprake për mbrojtje nga zgjeba është pastërtia personale.
Të njohur janë edhe riqrat, të cilët jetojnë në shtazët e egra dhe shtëpiake si dhe në lëkurën e njeriut duke thithur gjakun e amvisit, në ç'rast mund të shkaktojnë helmimin e tyre si dhe bartjen e sëmundjeve të ndryshme.
Këmbënyjorët përfaqësojnë grupin më të madh të llojeve shtazore. Përfshijnë mbi 1.000.000 lloje dhe përbëjnë afro 80% të tërë faunës së përhapur në rruzulin tokësor. Supozohet se numri i tyre është më i madh, meqenëse çdo ditë zbulohen lloje të reja. Janë të përhapur në mjedisin tokësor dhe ujor. Emri këmbënyjorë rrjedh nga greqishtja arthron=nyje dhe podhos=këmbë. Për nga ndërtimi i trupit, mënyra e të ushqyerit dhe tipi i aparatit gojor, këmbënyjëtuarit ndahen në disa grupe: ndër të cilat më të njohura janë gaforret, merimangat dhe insektet që jetojnë kryesisht në mjedisin tokësor e shumë më pak në atë ujor. Këtu duhet përmendur edhe këmbënyjëtuarit më të vjetër tribolitet, që kanë pasur ndërtim më primitiv. Këta kanë jetuar në kohëra të lashta dhe numri i tyre ka qenë afro 10.000 lloje. Mjedisi jetësor i tyre ka qenë uji. Sot hasen vetëm në formë të mbeturinave (fosileve).
A laspuateten (Arthropoda) san di gratst stam faan diarten.
A laspuateten (Arthropoda) san di gratst stam faan diarten.
De Liddfööt oder Liddpoten (Fachspraak: Arthropoden, greeksch: Arthropoda, Liddfööt) sünd en Stamm mank de Deerter. Dor höört so ünnerscheedliche Deerter to, as Insekten, Dusendfööt, Kreeften, Aantenmusseln, Spinnen, Skorpionen, Mieten un de Trilobiten, de utstorven sünd.
De Liddfööt sünd en Stamm, de bannig goot togange kümmt up de Eer. Bi 80 % vun al Aarden vun de Deerter, de dat hüdigendags överhaupt geven deit, sünd Liddfööt. De meisten vun jem höört to de Insekten to. Upkamen sünd se wohrschienlich bi de Kambrium-Exploschoon. In de Tiet an’n Anfang vun dat Kambrium vör üm un bi 540 Millionen Johren sünd sünd de mehrzelligen Tierstämm miteens uptreden. In den Burgess-Lei, de 505 Millionen Johr oolt is[1] hefft al de dree Ünnerstämm vun de Liddfööt, de Trilobiten un annere Gruppen, de dat hüdigendags gor nich mehr gifft, as Fossil de Tieden överduert. De Liddfööt ehre neegsten Verwandten sünd de Waterboren (Tardigrada) un de Stummelfööt. Towielen weert se mit de beiden annern to de Panarthropoda tohopensmeten, ofschoonst de doren gor keen Leden an jem ehre Beene hefft. In jungste Tiet hefft Forschers mit Hölp vun de Molekularbiologie rutfunnen, dat de Liddfööt veel ehr mit de Fadenwörm (Nematoda) un annere Huututtreckers (Ecdysozoa) verwandt sünd.
Al tohopen hefft se en Lief, dat ut verscheden Segmenten tohopenföögt is. An jedet Segment sitt hööchstens een Paar Leden, as Been, Mundwarktüüch un Föhlspriete. Al tohopen hefft se en Butenskelett, dat mit Hölp vun Sklerotin stiev warrt. Düt Skelett mutt bi dat Wassen mit Hölp vun dat Huuduttrecken verscheden Molen uttagen un nee anleggt weern. All hefft se ok en Hart an de Rüchsiet mit’n apen Kreisloop vun Hämolymphe un en Fallreepnervensystem an’e Buuksiet mit een Ganglion in jedet Segment. Sünnerlich bi de Aarden, de in’t Water leevt, kann in dat Bloot ok de Blootfarvstoff Hämocyanin insitten. Ähnlich as dat Hämoglobin in dat minschliche Bloot, is he dorför dor, dat Suerstoff wieterbröcht warrt.
De Liddfööt oder Liddpoten (Fachspraak: Arthropoden, greeksch: Arthropoda, Liddfööt) sünd en Stamm mank de Deerter. Dor höört so ünnerscheedliche Deerter to, as Insekten, Dusendfööt, Kreeften, Aantenmusseln, Spinnen, Skorpionen, Mieten un de Trilobiten, de utstorven sünd.
De Liddfööt sünd en Stamm, de bannig goot togange kümmt up de Eer. Bi 80 % vun al Aarden vun de Deerter, de dat hüdigendags överhaupt geven deit, sünd Liddfööt. De meisten vun jem höört to de Insekten to. Upkamen sünd se wohrschienlich bi de Kambrium-Exploschoon. In de Tiet an’n Anfang vun dat Kambrium vör üm un bi 540 Millionen Johren sünd sünd de mehrzelligen Tierstämm miteens uptreden. In den Burgess-Lei, de 505 Millionen Johr oolt is hefft al de dree Ünnerstämm vun de Liddfööt, de Trilobiten un annere Gruppen, de dat hüdigendags gor nich mehr gifft, as Fossil de Tieden överduert. De Liddfööt ehre neegsten Verwandten sünd de Waterboren (Tardigrada) un de Stummelfööt. Towielen weert se mit de beiden annern to de Panarthropoda tohopensmeten, ofschoonst de doren gor keen Leden an jem ehre Beene hefft. In jungste Tiet hefft Forschers mit Hölp vun de Molekularbiologie rutfunnen, dat de Liddfööt veel ehr mit de Fadenwörm (Nematoda) un annere Huututtreckers (Ecdysozoa) verwandt sünd.
Eitt liðadýr berður millum annað eyðkenni at tað hevur beinagrind uttaná kroppinum, sum í høvuðsdráttum er úr proteininum kitin. Liðadýr hava ikki eina beinagrind innaní.
Umi mymba ipy apytimby (Arthropoda) ha'e opaite oikovéva imichĩ, ipy apytykuéicha, ndoguerekóiva kangue rysýi ijatukupépe ha ikangue aty oĩ hete renondépe, ko'ã mymba'i ha'e pehẽ'a guasuvéva opa Yvy pegua, opaite tymbachu'i (taha'e eiru, tahýi, térã ha'éva panambi) ha'e niko mymba ipy apytimby, opaichagua ñandu, japeusaroto, guaiamu ha ambu'a ha'e avei mymba ipy apytimby.
Oĩve 1 200 000 mymba ipy apytimby juehegua ojekuaatee, umi tymbachu'i hetavéva (1 000 000),[1] upéva he'ise umíva ha'e 80% opaite pegua umi mymba juehegua ojeikuaáva.[2] Ojejuhu ko'ã mymba opa rendápe, heta umíva ikatu oveve avei, ha oĩ umíva ikatu oiko ýpe.
Umi mymba ipy apytimby (Arthropoda) ha'e opaite oikovéva imichĩ, ipy apytykuéicha, ndoguerekóiva kangue rysýi ijatukupépe ha ikangue aty oĩ hete renondépe, ko'ã mymba'i ha'e pehẽ'a guasuvéva opa Yvy pegua, opaite tymbachu'i (taha'e eiru, tahýi, térã ha'éva panambi) ha'e niko mymba ipy apytimby, opaichagua ñandu, japeusaroto, guaiamu ha ambu'a ha'e avei mymba ipy apytimby.
Oĩve 1 200 000 mymba ipy apytimby juehegua ojekuaatee, umi tymbachu'i hetavéva (1 000 000), upéva he'ise umíva ha'e 80% opaite pegua umi mymba juehegua ojeikuaáva. Ojejuhu ko'ã mymba opa rendápe, heta umíva ikatu oveve avei, ha oĩ umíva ikatu oiko ýpe.
Sillwichaki (Arthropoda) nisqakunaqa huk uywa rikch'aq putum. Sillwichakikunaqa khitina nisqamanta kapka qarayuqmi. Chakikunaqa raki-rakisqam, sillwikuna hina.
Kaymi rikch'aq sinrinkuna:
Sillwichaki (Arthropoda) nisqakunaqa huk uywa rikch'aq putum. Sillwichakikunaqa khitina nisqamanta kapka qarayuqmi. Chakikunaqa raki-rakisqam, sillwikuna hina.
Kaymi rikch'aq sinrinkuna:
Qaraqruyuwa (Crustacea) Cephalocarida Remipedia Samanachaki (Branchiopoda) Pura-pura qara (Malacostraca): Apanqura, Yukra, hukkunapas K'akichaki (Maxillopoda) Qallu kuru (Pentastomida) K'aki phapayuq (Chelicerata) Chakilla qhampu (Pycnogonida) Pakcha (Arachnida) (Awaq uru, Sira-sira, Chakisapa uru, hukkunapas) Chukichupa (Merostomata) Tunquchayuq (Tracheata) Waranqachaki (Myriapoda) Pachakchaki (Chilopoda) Ch'iñichaki (Symphyla) Iskaynintinchaki (Diplopoda) Asllachaki (Pauropoda) Palama (Insecta) Iskaychupa (Diplura) Llukchichaki (Protura) Phinkichupa (Collembola)Tiswermin (assaɣ usnan: Arthropod) d astirf ameqran yak deg tgelda n iɣersiwen dɣa tla (tesɛa) iburɣas, Tissisyin yakk d Tiruca., Taddadanin qqaren-t-d d akken 80% n iɣersiwen kecmen deg ustirf-a, Dɣa lant (sɛant) igmamen uddisen seg igezzumen iwermiyen d tiski tuffiɣt i tagaẓt, Akken i ttilin-t s waṭas deg yilel
Tiswermin d imuddaren armtegziten ilan (yesɛa) tafekka yettugezmer ar imuren
Asrgzem
Tiswermin (assaɣ usnan: Arthropod) d astirf ameqran yak deg tgelda n iɣersiwen dɣa tla (tesɛa) iburɣas, Tissisyin yakk d Tiruca., Taddadanin qqaren-t-d d akken 80% n iɣersiwen kecmen deg ustirf-a, Dɣa lant (sɛant) igmamen uddisen seg igezzumen iwermiyen d tiski tuffiɣt i tagaẓt, Akken i ttilin-t s waṭas deg yilel
Ko e veʻehokohoko (lea fakalatina (lea fakasaienisi): arthropoda) ko e vahe (phylum) ia ʻo e ngaahi monumanu ʻoku ʻi ai hanau vaʻe ʻoku hokohoko pe vahevaheʻi. Koeʻuhi ʻoku fefeka honau kili, kā ʻoku nau fiengaue. Tatau pehē, ʻoku vahevaheʻi honau sino, konga ʻe 2 he kulupu ʻe taha, 3 he taha, mo e hā fua.
Ko e vahe ia ʻo e ngaahi faʻahinga lahi taha, mahalo pe 80% he katoa.
ʻOku kau ko e ngaahi haʻa (classis) ki he vahé ni:
mo e ha ngaahi vahe siʻi, haʻa lahi mo e haʻa kehe.
ʻOku ʻi ai ha kau saienisi ʻoku ʻikai te nau ʻilo ʻa e "monumanu paipa mānava" pea ʻoku nau fakafaʻahinga ʻa e "veʻelahi" mo e "veʻeono" ʻi he vahe siʻi, pea toʻo ʻa e vahe lahi. Kaeʻumaʻā ko e fanga "veʻelahi" mahalo pē ʻoku kau kinautolu ki he "monumanu fingota ngeʻesi fefeka'.
Od svih beskičmenjaka zglavkari su postigli najviši evolucioni uspjeh. Poznato je oko milion vrsta. Žive na svim staništima: morska i slatka voda, kopno.
Tijelo im je građeno od glave, grudi i trbuha. Svaki od ovih dijelova sastoji se od više dijelova. Evolucioni uspjeh zglavkara zasniva se na razvoju vanjskog kostura. On im omogučava zaštitu, a postojanje većeg broja zglobova veću pokretljivost. Ovaj kostur je krut pa predstavlja prepreku rastu, koja je prevaziđena odbacivanjem starog kostura i njegova zamjena novim. U ovom periodu životinje su ranjive i nesposobne da se zaštite.
Osnovna osobina zglavkara je heterotomna segmentacija tijela. Za razliku od segmentacije anelida, kod kojih su svi segmenti međusobno jednaki, kod zglavkara se zglobovi jasno morfološki razlikuju.
Segmenti se grupišu u dva ili tri tjelesna regiona. Segmentisane su im i noge.
Osnovni tjelesni regioni su :
Kod pojedinih grupa ovi regioni se drugačije grupišu. Na površini tijela imaju čvrstu hitinsku kutikulu. Čvrst omotač je dobra zaštita tijelu. Naziva se spoljašnji kostur ili egzoskelet. Građen je od većeg broja ploča, koje pokrivaju tijelo i ekstremitete. U najvećem broju slučajeva skeletne ploče su međusobno pokretno spojene. Omogućava pokretljivost ovih organa. Služi kao zaštita unutrašnjih organa. Da bi rasli moraju se presvlačiti. Odbacuju staru kutikulu. Mišići i skeletne ploče funkcionišu kao sistem poluga.
Krvni sistem je otvorenog tipa. Srce se sastoji od jedne ili više komora. Linearno su raspoređene i imaju bočne linearno raspoređene otvore - ostije. Kroz njih krv (hemolimfa) iz perikardijalnog prostora ulazi u srce. Iz srca krvnim sudovima krv odlazi do tkiva i organa. Tu se ona izliva u šupljine i lakune oko tih organa.
Respiratorni organi vodenih zglavkara su škrge, a kod suhozemnih su traheje. Kod nekih oblika postoje pluća (pauci i škorpije) Traheje su razgranate cjevčice. One počinju sitnim otvorima na kutikuli (stigme). Zatim se granaju po cijelom tijelu.
Ekstetorni organi su različiti:
Nervni sistem je dostigao visok stepen razvoja. To je u vezi sa njihovim složenim ponašanjem. Imaju razvijen mozak. On se sastoji od 3 dijela:
Čula su dobro razvojena posebno čulo vida. Ono je glavno orijentaciono čulo. Mogu biti pokretljive na drškama ili nepokretljive na predenjem dijelu glave. Postoje proste (ocele) i facetovane oči. Facetovane se sastoje od nekoliko hiljada do nekoliko stotina hiljada ocela.
Polni organi su odvojeni čest polni diformizam. Polne žlijezde i njihovi odvodi su parni. Oplodnja se vrši u unutrašnjosti. Kod nekih predstavnika razviče je amebotalno tj. iz jajeta izlazi jedinka. Liči na odraslu jedinku.
Kod složenijih zglavkara razviće je sa metamorfozom. Može biti
Prelazni oblik između dvije grupe metazoa i zglavkara predstavljaju:
Po osobinama oni su slični prstenastim crvima i zglavkarima
Baršunasti crvi (Onychophora) otkriveni su 1826. Žive u tropskim i suptropskim predjelima, na vlažnim mjestima kao što je otpalo lišće, ispod kore mrtvog drveća. Ovo su noćne životinje. Hrane se ulovljenim insektima. Dužine su 1,4 do 15 cm, najčešće 2 cm. Liće na puževe golače, kišne gliste...
Njihovo tijelo je dugačko cilindričnog oblika podijeljeno na segmente od kojih svaki, sem glave, nosi po par nečlankovitih nožica sa stopalima na kojima je po par kandžica. Glava nije odvojena od tijela suženjem. Sastoji se od tri segmenta čeoni dio na kome su sva pipka i još dva na kojima je po jedno jednostavno oko i usta sa jakom čeljusti. Ekstretorni aparat sličan je onom kod prstenastih crva. Svaki nosi po par ekstretornih cjevčica. Sistem organa za krvotok i disanje sličan je onom kod zglavkara. Nožice podsječaju na parapodije prstenastih crva, ali imaju i snažne kandže slične onima kod zglavkara.
Živi ispod lišča tropskih šuma, krijući se danju , a noću izlazi u potragu za hranom.
Tartigrade (Tartigradae) su veoma sitne životinje. Dužina im rijetko prelazi 1 mm. Tijelo im je presvučeno debelom kutikulom ukrašenom šarama koje su često živo obojene. Ime su dobile po nezgrapnom karakterističnom načinom kretanja. Tijelo im podsječa na medvjeđi, pa ih nazivaju vodeni medvjedi.
Uvijek imaju 4 para zdepastih nožica koje su kao i kod baršunastih crva jednostavni valjkasti izdanci. Završavaju grupom sa po 4 do 8 oštrih šiljastih kandžica. Njima se hvataju za podlogu dok se kreću. Žive u pijesku morskog dna, slatkoj vodi, na lišajevima, mahovini. U periodu suše može da uzgubi tečnost, tako usporava metabolizam i u stanju su obamrlosti. Ovako mogu preživjeti nekoliko mjeseci.
Pentastomide (Pentastomidae) obuhvataju grupu sa oko 100 vrsta koje su paraziti. Dužina im je od nekoliko milimetara do 14 cm. Crvolikog su oblika. Na prednjem dijelu imaju pet izbočenja. Na grčkom ime im zunači petousti, a odnosi se na tih pet izdanaka. Samo jedan od njih predstavlja usta. Ostali zavržavaju kukicama ili kandžicama. Pomoču njih životinja se hvata za domaćina. Životni ciklus im je često složen i mogu, u toku razvoja , da se izredaju na više raznih domaćina. Odrasli oblici žive u disajnim putevima gmizavaca, ptica i sisara.
Lingituala je parazit nosne šupljine psa, nekad i drugih sisara pa i čovjeka. Kao larva živi na raznim organima zečeva i kunića.
Od svih beskičmenjaka zglavkari su postigli najviši evolucioni uspjeh. Poznato je oko milion vrsta. Žive na svim staništima: morska i slatka voda, kopno.
Tijelo im je građeno od glave, grudi i trbuha. Svaki od ovih dijelova sastoji se od više dijelova. Evolucioni uspjeh zglavkara zasniva se na razvoju vanjskog kostura. On im omogučava zaštitu, a postojanje većeg broja zglobova veću pokretljivost. Ovaj kostur je krut pa predstavlja prepreku rastu, koja je prevaziđena odbacivanjem starog kostura i njegova zamjena novim. U ovom periodu životinje su ranjive i nesposobne da se zaštite.
Τα Αρθρόποδα (Arthropoda) (ελλην. άρθρο + πόδι) είναι συνομοταξία από το βασίλειο των ζώων. Περίπου το 80% όλων των ειδών ζώων, που ζουν σήμερα, είναι αρθρόποδα.Έχουν καταγραφεί περίπου 1.100.000 είδη και πιθανότατα υπάρχουν πολύ περισσότερα.[1] Παρουσιάζουν εξαιρετική ποικιλομορφία. Απαντώνται σχεδόν σε όλο τον κόσμο, σε γλυκά και αλμυρά νερά καθώς και στην ξηρά. Πολλά είναι ιπτάμενα. Στα αρθρόποδα ανήκουν οι αράχνες, οι σκορπιοί, τα τσιμπούρια, τα καβούρια, τα έντομα κα., καθώς και πολλά απολιθωμένα είδη. Το μέγεθός τους κυμαίνεται από το μικροσκοπικό καρκινοειδές Stygotantulus έως το γιγάντιο γιαπωνέζικο καβούρι με μήκος σχεδόν 4 μέτρα.
Εμφανίζονται από την Κάμβρια περίοδο (ενδεχομένως και πιο νωρίς)[1] και περιλαμβάνουν μια μεγάλη ομάδα που εξαφανίστηκε τελείως, τους Τριλοβίτες.
Το επιδερμίδιό τους σχηματίζει έναν εξωσκελετό από χιτίνη και το σώμα τους διαμορφώνεται σε τμήματα που ονομάζονται μεταμερή. Ο εξωσκελετός αυτός προσφέρει στήριξη και προστασία από την αφυδάτωση. Ως εκ τούτου θεωρείται ο βασικός παράγοντας της επιτυχίας των αρθροπόδων στην χέρσο.[2] Φέρουν αρθρωτά άκρα ή εξαρτήματα, που μπορεί να διαφοροποιύνται σε πόδια (όπως συμβαίνει στα έντομα, τα μαλακόστρακα, και τα μυριάποδα) ή στοματικά μόρια ή κεραίες ή ενδιάμεσα άκρα (πχ. στοματικά πόδια). Ο αριθμός των ποδιών έχει χρησιμοποιηθεί παλαιότερα ως κριτήριο για την ταξινομική διαίρεση, όπως γίνεται αντιληπτό από τα ονόματα Δεκάποδα και Εξάποδα.
Είναι ζώα τριπλοβλαστικά. Το κοίλωμα στα ενήλικα άτομα είναι περιορισμένο και σχηματίζει το αιματόκοιλο. Επιπλέον πολλά χερσαία αρθρόποδα διαθέτουν ανεπτυγμένο τραχειακό σύστημα που εξασφαλίζει την αναπνοή τους. Τα περισσότερα υδρόβια αρθρόποδα διαθέτουν βράγχια. Εχουν κοιλιακή νευρική άλυσο με αρχικά ένα ζευγάρι γάγγλια κατά μέλος και ανοικτό κυκλοφορικό σύστημα με την καρδιά στη ράχη. Το μυϊκό τους σύστημα περιλαμβάνει γραμμωτούς και λείους μύες. Επιδεικνύουν μεγάλη ποικιλία αισθητήριων μηχανισμών και παρουσιάζουν ιδιαίτερα πρότυπα συμπεριφοράς, όπως οι σύνθετες κοινωνικές συμπεριφορές ορισμένων εντόμων (μυρμήγκια, μέλισσες , κα.).[1]
Σημαντικό χαρακτηριστικό τους είναι η έκδυση, η οποία έχει ιδιαίτερη σημασία για τα αρθρόποδα, καθώς το συμπαγές επιδερμίδιο εμποδίζει την αύξηση. Ως εκ τούτου ανα διαστήματα απορρίπτεται ολόκληρος ο εξωσκελετός και αντικαθίσταται από νέο. Σε πολλές περιπτώσεις μάλιστα, οι εκδύσεις οδηγούν σε μεγάλες μορφολογικές αλλαγές από ένα προνυμφικό στάδιο σε ένα ενήλικο, μια διαδικασία που ονομάζεται μεταμόρφωση.[2]
Κατά κύριο λόγο είναι γονοχωριστικά, και η γονιμοποίηση γίνεται εσωτερικά. Μπορεί να είναι ωοτόκα, ωοζωοτόκα ή ζωοτόκα.
Έχουν τα περισσότερα ενδιαιτήματα από κάθε άλλη ομάδα ζώων. Βρίσκονται από τα βάθη των ωκεανών ως τις πολικές περιοχές, ζώντας ακόμα και στις σχετικά πιο αφιλόξενες περιοχές της γης. Είναι κυρίως φυτοφάγα αλλά υπάρχουν τόσο σαρκοφάγα όσο και παμφάγα αρθρόποδα. Ελεύθερα ή και παρασιτικά. Μεταξύ των παρασιτικών είναι και τα Armillifer armillatus, τα οποία παρασιτούν κυρίως στην αναπνευστική οδός χερσαίων σπονδυλόζωων, κυρίως ερπετά, αλλά μπορούν να παρασιτούν και σε μεγάλα θηλαστικά όπως οι λεοπαρδάλεις[3].
Τα αρθρόποδα εμφανίζονται πρώτη φορά σε απολιθώματα του Καμβρίου, περιλαμβάνοντας πρωτόγονα καρκινοειδή, χηλοκερωτά καθώς και την εξαφανισμένη ομάδα των τριλοβιτών. Θεωρείται γενικά ότι τα αρθρόποδα προήλθαν εξελικτικά από έναν κοινό πρόγονο με τους δακτυλιοσκώληκες, καθώς και οι δύο ομάδες παρουσιάζουν μεταμέρεια και πολλές ομοιότητες στην φυσιολογία τους. Η διαφοροποίησή τους, που συχνά ονομάζεται «αρθροποδοποίηση» και περιλαμβάνει την δημιουργία του χιτινώδους εξωσκελετού και την ανάπτυξη των εξαρτημάτων, είναι ήδη εμφανής στο αρχείο των απολιθωμάτων. Είναι συνεπώς πολύ πιθανό ότι τα πρώτα αρθρόποδα εμφανίστηκαν ήδη από το Προκάμβριο. Ωστόσο η εξέλιξη αλλά και οι φυλογενετικές σχέσεις των επιμέρους ομάδων μεταξύ τους και με συγγενή φύλα παραμένουν αντικείμενο διαφωνίας.
Το σώμα των αρθροπόδων αποτελείται από πολλά διαδοχικά μεταμερή (σωμίτες), με ποικιλία προσαρτημένων αρθρωτών εξαρτημάτων, τα οποία λόγω της διαφοροποίησής τους συχνά σχηματίζουν διακριτές λειτουργικές ενότητες, τα τάγματα και συγκεκριμένα την κεφαλή, τον θώρακα και την κοιλία, χωρίς αυτή η διάκριση να είναι πάντα αρκετά εμφανής. Η κεφαλή φέρει αισθητήρια όργανα και άλλα στοματικά εξαρτήματα. Διακρίνεται στο πρωτοκέφαλο, που φέρει οφθαλμούς και κεραίες και το γναθοκέφαλο που φέρει στοματικά εξαρτήματα όπως γνάθους και γναθοπόδια. Το εμπρόσθιο άκρο του σώματος καλείται άκρο ενώ το οπίσθιο, τέλσον. [4] Κάθε σωμίτης περιβάλλεται από τέσσερις εξωσκελετικές πλάκες (σκληρίτες). Μια ραχιαία που ονομάζεται τεργίτης, μια κοιλιακή που ονομάζεται στερνίτης και δύο πλευρικές που ονομάζονται πλευρίτες. [5]
Τα διάφορα εξαρτήματα είναι σημαντικό χαρακτηριστικό της ομάδας αυτής (μιας και από αυτά πήρε το όνομά της) και ουσιαστικά απαρτίζουν τον μηχανισμό κίνησης και στήριξης. Εμφανίζονται ανά ζεύγη και γενικά θεωρείται ότι κατά την πρώιμη εμφάνισή τους, κάθε μεταμερές διέθετε από ένα ζεύγος εξαρτημάτων. [6] Τα εξαρτήματα πολλών αρθροπόδων όπως των καρκινοειδών, είναι δίκλαδα, και αποτελούνται από το ενδοπόδιο και το εξωπόδιο. Άλλων, όπως των εντόμων είναι μονόκλαδα.
Παρατηρείται σημαντική διαφοροποίηση των εξαρτημάτων τόσο σε επίπεδο ατόμου, εξασφαλίζοντας καταμερισμό εργασιών όσο και σε επίπεδο διαφορετικών ειδών, όπου φανερώνουν τις διάφορες οικολογικές πιέσεις που οδήγησαν την εξέλιξή τους. Η κίνηση των αρθρωτών ποδιών από γραμμωτούς μυς, εξασφαλίζει ταχύτητα στην κίνηση. Ο ρόλος τους μπορεί να είναι βαδιστικός, κολυμβητικός, πτητικός, τροφοληπτικός ή ακόμα και αισθητήριος, καθώς πολλά εξαρτήματα διαθέτουν αισθητήριες τρίχες και άλλες προσαρμογές.
Ο εξωσκελετός των αρθροπόδων αποτελεί το κυρίαρχο χαρακτηριστικό τους, καθώς προσφέρει προστασία από την αφυδάτωση και άλλους φυσικούς παράγοντες και επιπλέον μηχανική στήριξη, διασφαλίζοντας επαρκή κινητικότητα και ευελιξία. Είναι εξωδερμικής προέλευσης και σχηματίζεται από το επιδερμίδιο, ένα στρώμα χιτίνης και πρωτεϊνών που παράγεται από την υποκείμενη επιδερμίδα. Το πάχος του και η σκληρότητά του ποικίλει μεταξύ των επιμέρους ομάδων καθώς και μεταξύ των διαφόρων σταδίων ανάπτυξης (πχ. μια κάμπια με μια πεταλούδα).
Διακρίνονται δύο βασικές στοιβάδες, το λεπτό επιεπιδερμίδιο και το παχύ και σκληρό προεπιδερμίδιο. Και τα δύο σχηματίζονται από αλλεπάλληλα στρώματα (ελάσματα). Το πρώτο αποτελείται από μία λεπτή στρώση λιποπρωτεϊνών και (κυρίως σε χερσαία αρθρόποδα) κηρών και λιπαρών οξέων που αποτρέπουν την διαφυγή ή την ανεπιθύμητη είσοδο νερού στο σώμα. Αντίθετα το προεπιδερμίδιο συνίσταται κυρίως από πρωτεΐνη και χιτίνη σε αναλογία περίπου 1:1 (μεγαλύτερη συγκέντρωση χιτίνης στα καρκινοειδή). Η χιτίνη είναι ένα ιδιαίτερα ανθεκτικός αζωτούχος πολυσακχαρίτης, αδιάλυτος στο νερό και σε ασθενή οξέα και βάσεις. Η σύνδεση του με τις πρωτεΐνες, προς σχηματισμό πρωτεογλυκανών προσδίδει σκληρότητα και αντοχή στο επιδερμίδιο. Σε ένα ώριμο επιδερμίδιο μπορούν να διακριθούν δύο στρώσεις, το εξω- και το ενδο-επιδερμίδιο. Η διαφορά τους έγκειται στην παρουσία φαινολών στο εξωεπιδερμίδιο που σταθεροποιούν το δίκτυο χιτίνης-πρωτεϊνών.
Η χιτίνη από μόνη της παρά την ανθεκτικότητά και την ελαστικότητά της δεν είναι ιδιαίτερα σκληρή. Έτσι η σκληρότητα του επιδερμιδίου εξασφαλίζεται από δύο άλλους μηχανισμούς, την βιοορυκτοποίηση και την σκληροτινοποίηση. Η ορυκτοποίηση, δηλαδή η απόθεση ανόργανων ανθρακικών, φωσφορικών κλπ., αλάτων στο εξωεπιδερμίδιο, παρατηρείται σε μεγάλο ποσοστό στα καρκινοειδή και τα διπλόποδα. Η σκληροτινοποίηση από την άλλη, είναι μια ειδική πρωτεϊνική διεργασία και είναι εμφανίζεται κυρίως στα έντομα και άλλα χερσαία αρθρόποδα.[5]
Η επιδερμίδα που είναι υπεύθυνη για την παραγωγή του επιδερμιδίου είναι μονόστοιβo κυβικό επιθήλιο και φέρει αρκετούς αδένες ενώ αποτελεί την βάση για πολλούς εξωτερικούς σχηματισμούς όπως τρίχες, σμήριγγες, άκανθες κα. Διαχωρίζεται από το αιμόκοιλο μέσω της βασικής μεμβράνης.
Σε πολλά αρθρόποδα το επιδερμίδιο σχηματίζει εσωτερικές προεκβολές που ονομάζονται αποδέματα και στα οποία προσφύονται οι μύες.
Chelicerata
Pygnogonida
Xiphosura
Scorpiones
Amblypygi
Amblypygi
Araneae
Araneae
Opiliones
Acari
Acari
Trilobita




Crustaceae
Branchiopoda
Branchiopoda
Malacostraca, Dekapoda
Malacostraca, Isopoda
Malacostraca, Amphipoda
Cirripedia
Pemtastomida:
Tracheata
Myriapoda
Myriapoda
Hexapoda, Insecta
Hexapoda, Diplura
|first1= missing |last1= in Authors list (βοήθεια) Τα Αρθρόποδα (Arthropoda) (ελλην. άρθρο + πόδι) είναι συνομοταξία από το βασίλειο των ζώων. Περίπου το 80% όλων των ειδών ζώων, που ζουν σήμερα, είναι αρθρόποδα.Έχουν καταγραφεί περίπου 1.100.000 είδη και πιθανότατα υπάρχουν πολύ περισσότερα. Παρουσιάζουν εξαιρετική ποικιλομορφία. Απαντώνται σχεδόν σε όλο τον κόσμο, σε γλυκά και αλμυρά νερά καθώς και στην ξηρά. Πολλά είναι ιπτάμενα. Στα αρθρόποδα ανήκουν οι αράχνες, οι σκορπιοί, τα τσιμπούρια, τα καβούρια, τα έντομα κα., καθώς και πολλά απολιθωμένα είδη. Το μέγεθός τους κυμαίνεται από το μικροσκοπικό καρκινοειδές Stygotantulus έως το γιγάντιο γιαπωνέζικο καβούρι με μήκος σχεδόν 4 μέτρα.
Εμφανίζονται από την Κάμβρια περίοδο (ενδεχομένως και πιο νωρίς) και περιλαμβάνουν μια μεγάλη ομάδα που εξαφανίστηκε τελείως, τους Τριλοβίτες.
Το επιδερμίδιό τους σχηματίζει έναν εξωσκελετό από χιτίνη και το σώμα τους διαμορφώνεται σε τμήματα που ονομάζονται μεταμερή. Ο εξωσκελετός αυτός προσφέρει στήριξη και προστασία από την αφυδάτωση. Ως εκ τούτου θεωρείται ο βασικός παράγοντας της επιτυχίας των αρθροπόδων στην χέρσο. Φέρουν αρθρωτά άκρα ή εξαρτήματα, που μπορεί να διαφοροποιύνται σε πόδια (όπως συμβαίνει στα έντομα, τα μαλακόστρακα, και τα μυριάποδα) ή στοματικά μόρια ή κεραίες ή ενδιάμεσα άκρα (πχ. στοματικά πόδια). Ο αριθμός των ποδιών έχει χρησιμοποιηθεί παλαιότερα ως κριτήριο για την ταξινομική διαίρεση, όπως γίνεται αντιληπτό από τα ονόματα Δεκάποδα και Εξάποδα.
Είναι ζώα τριπλοβλαστικά. Το κοίλωμα στα ενήλικα άτομα είναι περιορισμένο και σχηματίζει το αιματόκοιλο. Επιπλέον πολλά χερσαία αρθρόποδα διαθέτουν ανεπτυγμένο τραχειακό σύστημα που εξασφαλίζει την αναπνοή τους. Τα περισσότερα υδρόβια αρθρόποδα διαθέτουν βράγχια. Εχουν κοιλιακή νευρική άλυσο με αρχικά ένα ζευγάρι γάγγλια κατά μέλος και ανοικτό κυκλοφορικό σύστημα με την καρδιά στη ράχη. Το μυϊκό τους σύστημα περιλαμβάνει γραμμωτούς και λείους μύες. Επιδεικνύουν μεγάλη ποικιλία αισθητήριων μηχανισμών και παρουσιάζουν ιδιαίτερα πρότυπα συμπεριφοράς, όπως οι σύνθετες κοινωνικές συμπεριφορές ορισμένων εντόμων (μυρμήγκια, μέλισσες , κα.).
Σημαντικό χαρακτηριστικό τους είναι η έκδυση, η οποία έχει ιδιαίτερη σημασία για τα αρθρόποδα, καθώς το συμπαγές επιδερμίδιο εμποδίζει την αύξηση. Ως εκ τούτου ανα διαστήματα απορρίπτεται ολόκληρος ο εξωσκελετός και αντικαθίσταται από νέο. Σε πολλές περιπτώσεις μάλιστα, οι εκδύσεις οδηγούν σε μεγάλες μορφολογικές αλλαγές από ένα προνυμφικό στάδιο σε ένα ενήλικο, μια διαδικασία που ονομάζεται μεταμόρφωση.
Κατά κύριο λόγο είναι γονοχωριστικά, και η γονιμοποίηση γίνεται εσωτερικά. Μπορεί να είναι ωοτόκα, ωοζωοτόκα ή ζωοτόκα.
Έχουν τα περισσότερα ενδιαιτήματα από κάθε άλλη ομάδα ζώων. Βρίσκονται από τα βάθη των ωκεανών ως τις πολικές περιοχές, ζώντας ακόμα και στις σχετικά πιο αφιλόξενες περιοχές της γης. Είναι κυρίως φυτοφάγα αλλά υπάρχουν τόσο σαρκοφάγα όσο και παμφάγα αρθρόποδα. Ελεύθερα ή και παρασιτικά. Μεταξύ των παρασιτικών είναι και τα Armillifer armillatus, τα οποία παρασιτούν κυρίως στην αναπνευστική οδός χερσαίων σπονδυλόζωων, κυρίως ερπετά, αλλά μπορούν να παρασιτούν και σε μεγάλα θηλαστικά όπως οι λεοπαρδάλεις.
Бандпойҳó (лот. Arthropoda, аз юн.-қад. ἄρθρον) — типи ҷонварони бемуҳра. Қариб 2 млн намуд доранд.
Дар Тоҷикистон тақрибан 10 ҳазор намуди бандпойҳо маълум аст. Бандпойҳо дар ҳаво, замин ва об зиндагӣ мекунанд. Баъзеашон алафхӯр, даранда ва баъзеи дигарашон паразитанд. Авлоди қадимаи бандпойҳо ҳалқакирмҳои баҳрӣ ба шумор мераванд. Харчангу тортанакшаклҳо, ҳашарот ва бисёрпойҳо синфҳои асосии бандпойҳо мебошанд. Тани бандпойҳо аз се қисм (сар, сина ва шикам) иборат аст. Бисёр вақт қисми сина бо сар васл шуда, сарсинаро ҳосил мекунад (масалан, дар тортанакшаклҳо, харчангҳои даҳпой ва ғайра). Пойҳои бандпойҳо эҳтимол дар аввал душоха будаанд. Ин аломат, ки масалан, хосси трилобитҳои давраи палеозой буд, дар харчангшаклҳои имрӯза низ боқӣ мондааст. Бадани бандпойҳо бо кутикула, ки сипари муҳофиз ҳосил мекунад (ба он мушакҳо часпидаанд), пӯшида шудааст. Бинобар ин, инкишофи бандпойҳо ба таври даврӣ дар фосилаи ба кутикулаи нав иваз шудани кутикулаи кӯҳна ба амал меояд. Силсилаи асаби бандпойҳо аз се ҷуфт ганглия (мағзи сар) ва асаби шикам иборат мебошад. Бандпойҳо бо узвҳои махсус нафас мегиранд, ки рӯйи онҳо кутикулаи нафис дорад. Дар бандпойҳои обӣ ғалсама вазифаи узви нафасгириро иҷро мекунад. Узвҳои ҳисси бандпойҳо хуб инкишоф ёфтаанд. Бандпойҳо бо таври ҷинсӣ, одатан, бо роҳи бордоршавӣ ва партеногенезӣ (як шакли афзоиши ҷинсии организмҳо, ки ҳангоми он ҳуҷайраҳои ҷинсии модина бидуни бордоршавӣ инкишоф меёбанд) афзоиш мекунанд.
Таснифи бандпойҳо гуногун аст. Дар асоси яке аз онҳо бандпойҳо ба чор зертип тақсим мешаванд:
Бисёр бандпойҳо ҷонварони муфид мебошанд. Онҳоро дар ғизо истифода мебаранд (масалан, харчангҳо); аз баъзе бандпойҳо (ору, кирмак) маҳсулоти пурбаҳо, масалан, асал, мум, абрешим ва ғайра мегиранд. Бандпойҳои зараррасон низ мавҷуданд (масалан, гамбуски колорадоӣ, малах, канаи хоришак, кайк, шабуш ва ғайра). Баъзе бандпойҳо ангезандаи бемориҳоро интиқол медиҳанд (кӯрпашшаҳо, канаҳо ва ғайра).
Буынтыкаяклылар (лат. Arthropoda), умырткасызлар тибы. 5 классы, Җир шарында таралган 2 млн артык төре билгеле. Ярым типлары: трилобитсыманнар (юкка чыккан трилобит классы белән бергә); хелицерлар (меростомлар һәм үрмәкүчсыманнар класслары белән бергә); саңак аша тын алучылар (кысласыманнар классы белән бергә); трахея аша тын алучылар, яки парлы мыеклылар (кыргаяклар һәм бөҗәкләр класслары белән бергә).
Гәүдәсе баш, күкрәк һәм корсактан тора; ике яклы симметрияле, саклагыч панцирен барлыкка китерүче хитин кутикула (аңа эчке мускуллар ябешә) белән капланган. Тән куышлыгы катнаш типлы (миксоцель). Гетероном сегментацияле. Эволюция процессында куыш, буынтыклы аягы авыз аппаратына, йөрү аягына, йөзү әгъзасына һ.б. әверелгән яки редукциягә бирелгән. Эчәклеге хитин белән капланган эктодермаль алгы һәм арткы эчәкләрдән һәм энтодермаль урта эчәктән [аның белән кайсы бер буынтыкаяклылар аш сеңдерү (бавыр-ашказан асты) бизләре, яки бавыр көпшәләре тоташкан] тора. Саңаклар (кысласыманнар), үпкәләр һәм (яки) трахеяләр (үрмәкүчсыманнар һәм бөҗәкләр) аша тын ала. Нерв системасы 3 пар тоташкан йоткылык өсте ганглиеннан (баш мие) һәм корсак нервы чылбырыннан тора. Күп буынтыкаяклыларның катлаулы фасеткалы күзе, механик һәм хеморецепторлары, ишетү әгъзалары һ.б. бар. Кан тамырлары системасы йомык түгел (аорта һәм артериялар сакланган), йөрәге метамер, артериаль. Бүлеп чыгару әгъзалары — метанефридия, яки мальпиги тамыры (җир өсте формаларының). Аягы буынтыклы, күчеп йөрү, тын алу, эләктерү һ.б. функцияләр башкара.
Күпчелек буынтыкаяклылар аерым җенесле. Җенси юл белән үрчи, кайсы вакыт партеногенез хас. Үсеше тулы булмаган (курчак стадиясе юк) яки тулы метаморфозлы. Буынтыкаяклылар көбәсен салган вакытта үсә. Фитофаг, сапрофаг, ш. ис. детритофаг, некрофаг, ерткыч, паразит һ.б.
Буынтыкаяклылар — экосистемалардагы туклану чылбыры компоненты, күп төрләрен (креветкалар, лангустлар, елга кыслалары, омарлар һ.б.) кеше азык итеп куллана. Үсемлекләрне серкәләндерә, яшәү эшчәнлеген кыйммәтле продуктларын (бал корты балавызы, бал, ефәк һ.б.) эшли, үрчетү объектлары (бал корты, тут ефәк күбәләге), кеше һәм хайван паразитлары (борча, бет, корчаңгы бетчәсе), инфекцион авыруларны таратучылар (малярия черкие, энцефалит талпаннары һ.б.), авыл хуҗалыгы һәм урман культуралары корткычлары булып тора.
Буынтыкаяклылар (лат. Arthropoda), умырткасызлар тибы. 5 классы, Җир шарында таралган 2 млн артык төре билгеле. Ярым типлары: трилобитсыманнар (юкка чыккан трилобит классы белән бергә); хелицерлар (меростомлар һәм үрмәкүчсыманнар класслары белән бергә); саңак аша тын алучылар (кысласыманнар классы белән бергә); трахея аша тын алучылар, яки парлы мыеклылар (кыргаяклар һәм бөҗәкләр класслары белән бергә).
Быуынтығаяҡлылар (рус. Членистоногие, лат. Arthropoda, бор. грек. ἄρθρον — ,sesy һәм πούς, ποδός) — кәүҙәләре бүлкәттәргә (сегменттар) бүленгән, ә аяҡтары (ослоҡтары) айырым-айырым быуынтыҡтарҙан торған умыртҡаһыҙ хайуандар тибы.
Төп үҙенсәлектәре:
Был тип 1,5 млн тирәһе төрҙө берләштерә. Бер нисә классты берләштерә. Мәҫәлән: ҡыҫала һымаҡтар, үрмәксе һымаҡтар, бөжәктәр һ. б.
Беренсе быуынтыҡаяҡлылар боронғо иң ябай төҙөлөшлө күп төклө селәүсендәрҙән килеп сыҡҡандар.
.
Быуынтығаяҡлылар (рус. Членистоногие, лат. Arthropoda, бор. грек. ἄρθρον — ,sesy һәм πούς, ποδός) — кәүҙәләре бүлкәттәргә (сегменттар) бүленгән, ә аяҡтары (ослоҡтары) айырым-айырым быуынтыҡтарҙан торған умыртҡаһыҙ хайуандар тибы.
Төп үҙенсәлектәре:
хитинлы кутикуланан торған тышҡы һөлдә, тәндәре төҙөлөштәре бер төрлө булмаған (гетероном) бүлкәттәрҙән — сегменттарҙан тора, үҙ-ара һәм тән менән хәрәкәтсән тоташҡан быуынтыҡтарҙан торған ослоҡтары булыуы.Был тип 1,5 млн тирәһе төрҙө берләштерә. Бер нисә классты берләштерә. Мәҫәлән: ҡыҫала һымаҡтар, үрмәксе һымаҡтар, бөжәктәр һ. б.
Суставаногія[1] (па-лацінску: Arthropoda; ад грэцкага Ἀρθρον — «сустаў» і Πούς / ποδός — «нага»), таксама — чэлясаногія[2] — тып першаснаротых жывёлаў, які ўключае вусякоў, ракападобных, павукападобных і шматножак. Паводле колькасьці відаў і сваёй распаўсюджанасьці лічыцца адной з самых разнастайных групаў жывых арганізмаў. Колькасьць відаў суставаногіх перавышае колькасьць відаў ўсіх астатніх жывёлаў разам узятых.
Асноўная асаблівасьць прадстаўнікоў дадзенай групы жывёлаў — наяўнасьць хітынавага вонкавага шкілета.
Суставаногія распаўсюджаныя практычна паўсюль і адыгрываюць даволі значную ролю. Вусякі, разам з сысунамі й птушкамі, — дамінуючая група наземных жывёлаў. Налічваецца ня менш за 1,5 мільёны відаў вусякоў, але магчыма іх зашмат больш. Павукапабодныя й шматножкі, хоць і не дасягнулі такой разнастайнасьці, як вусякі, але таксама шырока распаўсюджаныя. Нарэшце, ракападобныя насяляюць амаль усе шыроты й глыбіні Сусьветнага акіяну. У некаторых рэгіёнах ракападобныя — пануючая група зоаплянктона.
Паводле падлікаў суставаногія маюць каля 1 170 000 апісаных відаў, і на іх прыпадае больш за 80% усіх вядомых відаў існых жывёлаў[3]. Іншае дасьледаваньне налічвае ад 5 да 10 мільёнаў існых відаў суставаногіх, якія ўлучаюць як ужо апісаныя віды, гэтак яшчэ не апісаныя[4]. Зрабіць дакладную ацэнку агульнай колькасьці жывых віды надзвычай цяжка, таму што гэтая справа часьцяком залежыць ад шэрагу дапушчэньняў пры разьліке колькасьці ў пэўных месцах і потым вылічваньня гэтага паказчыку для ўсяго сьвету. Дасьледаваньне, праведзенае ў 1992 годзе, паказала, што ў Коста-Рыке існуе каля 500 тысячаў відаў жывёлаў і расьлін, зь якіх 365 тысячаў відаў належалі да тыпу суставаногіх. Суставаногія зьяўляюцца адным з самых распаўсюджаных тыпаў шматвузавых арганізмаў, выяўленых у біясфэры[5].
Яны зьяўляюцца важнымі чальцамі марскіх, прэснаводных, наземных і паветраных экасыстэмаў, а таксама зьяўляюцца адной з двух асноўных групаў жывёлаў, якія прыстасаваліся да жыцьця ў сухім навакольлі.
Суставаногія маюць шмат агульнага з колцавымі чарвякамі, асабліва поліхетамі. Аднак праз шэраг арамарфозаў яны дасягнулі вышэйшай ступені дыфэрэнцыяцыі тканінаў і органаў, перш за ўсё нэрвовай сыстэмы й органаў пачуцьцяў, некаторыя групы маюць складаныя паводзіны, якое праяўляецца ў клопаце пра нашчадзтва, здольнасьці да складанай будаўнічай дзейнасьці, грамадзкага жыцьця і нават мове, як то ў вышэйшых перапонкавакрылых.
Як і колцавыя чарвякі, суставаногія ёсьць мэтамэрнымі жывёламі, іхнае цела складаецца з шэрагу сэгмэнтаў, аднак мэтамэрыя суставаногіх рэзка гетэраномная. Групы падобных сэгмэнтаў аб’ядноўваюцца ў аддзелы цела — тагмы. Часьцей за ўсё вылучаюцца тры тагмэ: галава, грудзі й брушка. У павукападобных галава зьліваецца з грудзьмі ў адзіную тагме — галавагрудзі, у трылабітаў і шматножак ёсьць толькі дзьве тагмэ — галава й тулава. Асаблівае значэньне мае працэс цэфалізацыі — утварэньне галавы, дзе сканцэнтраваны органы пачуцьцяў, як то вусікі й вочы, і першаснай перапрацоўкі ежы, то бок ротавы апарат.
Галава складаецца з галоўнай акроны, і некалькіх пярэдніх сэгмэнтаў. На думку большасьці навукоўцаў, акрон гамалягічны прастаміюму колцавым чарвяком. Галава суставаногіх мае шэсьць сэгмэнтаў. Колькасьць сэгмэнтаў, якія ўваходзяць у склад грудзей і брушка, якое сканчаецца анальнай шчылінай, рэзка вагаецца, але ў межах тыпу назіраецца тэндэнцыя да стабілізацыі й памяншэньня іхнай колькасьці. У выніку тагматызацыі аб’ядноўваецца й унутранае ўтрыманьне сэгмэнтаў: адны органы разрастаюцца за кошт іншых раўнацэнных органаў. У прыватнасьці, назіраецца канцэнтрацыя гангліяў нэрвовай сыстэмы ў галаўны мозаг.
Палажэньне групы суставаногіх у таксанамічнай герархіі неадназначнае. З аднаго боку, некаторымі аўтарамі была прапанаваная тэорыя, паводле якой некаторыя падтыпы суставаногіх ня маюць агульнага продка з колцавымі чарвякамі (лац. Annelida). У прыватнасьці, меркавалася, што падтып трахейныя (лац. Uniramia) бліжэй да аніхафораў (лац. Onychophora), чым да астатніх суставаногіх. Іншымі словамі, меркавалася поліфілія суставаногіх. Аднак большасьць навукоўцаў не прыняло гэты пункт гледжаньня, і гэта таксама супярэчыць генэтычным дасьледаваньням.
Зь іншага пункту гледжаньня, больш традыцыйным варыянтам зьяўлялася сьцьвярджэньне, што колцавыя чарвякі - бліжэйшыя сваякі як суставаногіх, гэтак і аніхафораў, грунтуючыся на аднолькавай колькасьці сегмэнтаў. Пазьней некаторыя аўтары сталі казаць пра канвэргентную эвалюцыі, лічачы, што суставаногія бліжэй да нэматода (лац. Nematoda), маючы агульныя зь імі здольнасьці да лінькі. Два верагодныя радаводы вызначаюць два вышэйстаячыя таксоны, да якіх можна было б прылічыць групу суставаногіх жывёлаў: сустаўныя (лац. Articulata) і ліняючыя (лац. Ecdysozoa)[6].
Найстаражытны зь вядомых суставаногіх — спрыгіна (позьні пратэразой) — мае ў будове цела нямала агульнага з колцавымі чарвякамі. Зь іншага боку, не выключаецца, што спрыгіна ставілася да зьніклага тыпу, хоць і надзвычай блізкаму да суставаногіх.
Клясыфікацыя падтаксонаў унутры групы суставаногіх таксама неадназначная. Пяць асноўных падгрупаў адносяць то да падтыпаў, то да клясах. Акрамя гэтых падгрупаў маецца таксама некаторы лік выкапнёвых жывёлаў, у большасьці сваім зь ніжнекембрыйскага пэрыяду, якія цяжка аднесьці да якой-небудзь падгрупы альбо па прычыне непадабенства зь вядомымі групамі, альбо з-за няяснасьці іхных роднасных узаемаадносінаў. Шматножкі й казуркі часьцяком аб’ядноўваюцца ў адну групу непоўнавусых. Некаторыя апошнія дасьледаваньні, аднак, сьведчаць, што шматножкі не бліжэй да казурак, чым да ракападобных.
![]() — сховішча мультымэдыйных матэрыялаў
— сховішча мультымэдыйных матэрыялаў
Суставаногія (па-лацінску: Arthropoda; ад грэцкага Ἀρθρον — «сустаў» і Πούς / ποδός — «нага»), таксама — чэлясаногія — тып першаснаротых жывёлаў, які ўключае вусякоў, ракападобных, павукападобных і шматножак. Паводле колькасьці відаў і сваёй распаўсюджанасьці лічыцца адной з самых разнастайных групаў жывых арганізмаў. Колькасьць відаў суставаногіх перавышае колькасьць відаў ўсіх астатніх жывёлаў разам узятых.
Асноўная асаблівасьць прадстаўнікоў дадзенай групы жывёлаў — наяўнасьць хітынавага вонкавага шкілета.
Суставаногія распаўсюджаныя практычна паўсюль і адыгрываюць даволі значную ролю. Вусякі, разам з сысунамі й птушкамі, — дамінуючая група наземных жывёлаў. Налічваецца ня менш за 1,5 мільёны відаў вусякоў, але магчыма іх зашмат больш. Павукапабодныя й шматножкі, хоць і не дасягнулі такой разнастайнасьці, як вусякі, але таксама шырока распаўсюджаныя. Нарэшце, ракападобныя насяляюць амаль усе шыроты й глыбіні Сусьветнага акіяну. У некаторых рэгіёнах ракападобныя — пануючая група зоаплянктона.
Членконогите (лат. Arthropoda од грчки: ἀρθρον — членче и грчки: πούς/ποδός — нога) се најголемиот и најразновидниот животински тип. Во членконоги спаѓаат раковите, инсектите, многуногите, клештарите и изумрените трилобити. Членконогите се карактеризираат со сегментирано тело прекриено со сегментиран надворешен скелет (егзоскелет), со парни сегментирани додатоци на секој сегмент. Присутен е комплексен нервен систем со дорзален мозок (ганглија), со конективни нерви кои минуваат низ антериорниот крај на дигестивниот систем и вентрален нерв со ганглија во секој телесен сегмент. Имаат отворен тип на циркулаторен систем со дорзално срце во кое крвта тече низ парни отвори (остии) и многу редуцирана телесна празнина (целом). Поради тоа што поврзаниот егзоскелет го блокира растот на организмот, тој мора периоднично да се менува по пат на преслекување. Овој феномен наречен екдиза е карактеристична особина за овој тип животни; овозможува брзо растење и значајна промена во формата на телото сè додека новиот егзоскелет, секретиран од животното, не се стврдне. Членконогите се главно копнени животни, но водните претставници се исто така доста добро познати. Постојат неколку поттипови со шеснаесет класи. Меѓутоа, се среќаваат и разни други систематики.
Кај членконогите телото е покриено со еднослојна кожа што излачува цврст заштитен слој наречен хитин. Хитинот не е подеднакво дебел по целото тело. Помеѓу членчињата тој е потенок и еластичен што овозможува движење на одделни делови на телото. Хитинот го заштитува телото од механички повреди, од губење вода, а воедно тој преставува надворешен скелет за кој се прицврстени мускулите.
Успешноста на членконогите во природата се должи на нивниот цврст надворешен скелет, сегментацијата и поврзаните телесни додатоци. Додатоците се користат за хранење, сетилна рецепција, одбрана и движење. Мускулниот систем е повеќе или помалку потпомогнат од хидрауликата која доаѓа од крвниот притисок создаден од срцето. Хидрауличниот систем е особено добро развиен кај пајаците.
Акватичните членконоги користат жабри за гасна размена. Овие жабри имаат екстензивна површина која е во контакт со околната вода. Копнените членконоги имаат внатрешни биолошки површини кои се специјализирани за размена на гасови. Инсектите и повеќето други копнени видови имаат трахеални системи: воздушни вреќички на кои се надоврзуваат каналчиња чиј друг крај се отвора во пори (наречени спиракули) надвор од телото во епидермалната кутикула. Другите имаат листовидни бели дробови, односно модифицирани жабри за дишење, присутни кај видовите како кокосната краба. Некои делови од нозете на војничките краби се покриени со кислородно апсорбирачка мембрана. Жабрените комори кај копнените краби понекогаш имаат две различни структури: една која е жабреста и служи за дишење во вода и друга специјално адаптирана за преземање на кислородот.
Членконогите имаат отворен циркулаторен систем. Хемолимфата со хемоцијанинот, респираторен пигмент базиран на бакар, се пропелира од страна на серии на срца во телесната празнина каде доаѓа во директен контакт со ткивата. Членконогите се протостоми. Кај нив постои целом, но е редуциран во тенка празнина околу репродуктивните и екскреторните органи, при што доминантна телесна празнина е хемоцелот исполнет со хемолимфа во која се капат органите. Членконожното тело е поделено на серии од различни сегменти, како и од предсегментален акрон кој обично ги потпира сложените и простите очи и постсегментален телсон. Овие сегменти се групирани во најразлични, специјализирани телесни региони наречени тагмата. Секој сегмент поддржува пар на телесни додатоци.
Кутикулата кај членконогите образува ригиден егзоскелет, составен главно од хитин, кој периодично се менува како што животното расте. Овие животни содржат и внатрешна зона (прокутикула) која е изградена од протеин и хитин и е одговорна за силината на надворешниот скелет. Надворешната зона (епикутикула) лежи на површината на прокутикулата. Таа е нехитинозна и претставува комплекс од протеини и липиди. Овозможува влажност и заштита на прокутикулата. Егзоскелетот има форма на чинии наречени склерити на сегментите, како и прстени на додатоците кои ги делат нив на сегменти одделени со зглобови. Овие особини ги одделуваат членконогите од нивните роднини онихофорите и тардиградите, наречени и лобопода.
Трилобитите сочинуваат целосно изумрена група на примитивни морски членконоги. Тие биле многу распространети во камбриумот и ордовикот, а станале истребени цо перм. Нивното зарамнето, овално тело било составено од глава покриена со дорзален штит, торакс, и терминален сегмент (пигидиум). Повеќе од 3.900 видви биле долги од 2,5 до 10 см, а некои планктонски форми биле помали, додека некои биле долги околу 76 см. Riarthrus eatoni е фосилен трилобит, чест во морињата од периодот ордовик.
Мандибулатите ја сочинуваат најголемата и најваријабилната членконожна група и се карактеризираат со присуството на модифицирани телесни додатоци (мандибули) на страната од устата кои ги користат како вилици. Постојат шест класи, сите карактеризирани со различните аспекти на телесната форма.
Членовите на оваа класа се карактеризираат со два пара на антени и два пара на модифицирани додатоци (магзили) кои се користат при хранењето. Постојат околу 40,000 видови на ракови (крустацеи), меѓу кои се јастозите, крабите, копеподите и голем број на минијатурни планктонски форми. Раковите се единствените членконоги кои се главно акватични (повеќето се морски видови). Некои, пак, се распространети во влажни станишта близу до вода. Имаат жабри за респирација. Градниот регион типично има нозе за одење (переиоподи) и опашен дел составен од пар на додатоци (уроподи), како и телсон. Нивните екскреторни органи се модифицирани нефридии што произведуваат разредена урина која содржи голем дел амонијак.
Класата Chilopoda вклучува 5,000 видови на центипеди, од кои сите се копнени. Центипедите се месојади и предатори, кои го имобилизираат својот плен (обично помали артроподи) со помош на нивните отровни канџи. Телото е составено од глава со пар на антени, пар на мандибули и два пара на магзили, како и труп со еден пар на нозе на секој сегмент. Предниот пар на додатоци на трупот (прехензори) е опремен со отровни жлезди. Јувенилните единки можат да имаат помалку додатоци отколку кај адултните. Нови сегменти се додаваат за време на растењето. Хилоподоте се наоѓаат во тропските и умерените климатски појаси.
Постојат околу 8.000 видови во оваа класа, која ги сочинува многуногите. Тие се среќаваат насекаде. Главата има пар на антени , пар на мандибули и два пара на магзили кои обично се споени во единствена уста, хилогнатариум. Многуногите имаат трахеален систем за респирација. Тие се растенојади или сапроби на мртов растителен материјал. Многу претставници се заштитени со жлезди кои произведуваат отров или непријатни соединенија.
Има околу 500 познати видови кои припаѓаат на оваа група. Пауроподите се со меки тела, мали (0,5 до 2 мм во должина), почвени членконоги и се космополити. Тие се елонгирани и имаат 9-11 пара на нозе, но немаат трахеи и срце.
Членовите на класата Symphyla се брзи трчачи кои се долги од 2,5 до 10 см. Класата вклучува околу 160 видови. Тие се главно сапроби на изумрена вегетација, но еден вид, Scutigerella immaculata, е сериозен штетник на одредени посеви. Симфилите имаат дванаесет пара на нозе и се претставени од центипедите.
Класата Insecta е најголемата артроподна класа која се состои од илјадници видови. Освен неколку примитивни или високо модифицирани форми, инсектите се карактеризираат со еден или два пара на крила прикрепени за тораксот. Главата има пар антени, пар мандибули и два пара на модифицирани магзили кои ги формираат усните делови. Абдоменот е добро издиференциран од тораксот и нема додатоци освен редуцирани кои се модифицирани во репродуктивни органи. Типичната глава на инсектите има сложени очи и едно или повеќе прости очи и е прекриена со егзоскелетен штит. Тораксот се состои од три сегменти, од кои секој носи пар на нозе. Последните два сегмента обично имаат пар на крила. Инсектите се главно копнени суштества и поседуваат трахеи за гасна размена. Се карактеризираат со единствени екскреторни органи, познати како Малпигиеви цевчиња, кои се корисни при зачувувањето (конзервирањето) на водата.
Членконогите (лат. Arthropoda од грчки: ἀρθρον — членче и грчки: πούς/ποδός — нога) се најголемиот и најразновидниот животински тип. Во членконоги спаѓаат раковите, инсектите, многуногите, клештарите и изумрените трилобити. Членконогите се карактеризираат со сегментирано тело прекриено со сегментиран надворешен скелет (егзоскелет), со парни сегментирани додатоци на секој сегмент. Присутен е комплексен нервен систем со дорзален мозок (ганглија), со конективни нерви кои минуваат низ антериорниот крај на дигестивниот систем и вентрален нерв со ганглија во секој телесен сегмент. Имаат отворен тип на циркулаторен систем со дорзално срце во кое крвта тече низ парни отвори (остии) и многу редуцирана телесна празнина (целом). Поради тоа што поврзаниот егзоскелет го блокира растот на организмот, тој мора периоднично да се менува по пат на преслекување. Овој феномен наречен екдиза е карактеристична особина за овој тип животни; овозможува брзо растење и значајна промена во формата на телото сè додека новиот егзоскелет, секретиран од животното, не се стврдне. Членконогите се главно копнени животни, но водните претставници се исто така доста добро познати. Постојат неколку поттипови со шеснаесет класи. Меѓутоа, се среќаваат и разни други систематики.
Кај членконогите телото е покриено со еднослојна кожа што излачува цврст заштитен слој наречен хитин. Хитинот не е подеднакво дебел по целото тело. Помеѓу членчињата тој е потенок и еластичен што овозможува движење на одделни делови на телото. Хитинот го заштитува телото од механички повреди, од губење вода, а воедно тој преставува надворешен скелет за кој се прицврстени мускулите.
Үет хөлтний хүрээ (Arthropoda хүрээ, Грек: ἄρθρον буюу arthron - "нийлмэл", ποδός буюу podos - "хөл" гэсэн утгатай) нь шавьж, хавч хэлбэртэн зэргийг багтаасан амьтны аймгийн хамгийн том хүрээ юм. Энэ хүрээнд 1,500,000 орчим зүйл амьтад багтана. Үет хөлтний гол онцлог нь тэдгээрийн бие нь дор хаяж нэг сигмент бүхий үелэсэн бүтэцтэй байна. Хоол боловсруулах систем нь өмнөд, дунд, хойд гэсэн гурван хэсгээс тогтно. Хитин бүрхүүлээр бүрхэгдэж гадаад араг яс үүсгэнэ.
Одоо оршин буй амьтдын>80% нь үет хөлтөн байдаг байна[1]. Үет хөлтөн нь тэнгис, цэнгэг ус, эх газарт түгээмэл тааралдана. Биеийн хэмжээ нь планктоноос эхлээд хэдэн метр хүрнэ. Одоо буй хамгийн том үет хөлтөн бол "Японы аалзан хавч" бөгөөд хөлнийх нь хэмжээг оруулан тооцвол нийт урт нь 3,5 метр хүрдэг байна.Үет хөлтөн нь тав болон түүнээс олон хөлтэй байна.
Үет хөлтний хүрээг дотор нь хэд хэдэн дэд хүрээ болгодог.
Үет хөлтний хүрээ (Arthropoda хүрээ, Грек: ἄρθρον буюу arthron - "нийлмэл", ποδός буюу podos - "хөл" гэсэн утгатай) нь шавьж, хавч хэлбэртэн зэргийг багтаасан амьтны аймгийн хамгийн том хүрээ юм. Энэ хүрээнд 1,500,000 орчим зүйл амьтад багтана. Үет хөлтний гол онцлог нь тэдгээрийн бие нь дор хаяж нэг сигмент бүхий үелэсэн бүтэцтэй байна. Хоол боловсруулах систем нь өмнөд, дунд, хойд гэсэн гурван хэсгээс тогтно. Хитин бүрхүүлээр бүрхэгдэж гадаад араг яс үүсгэнэ.
Одоо оршин буй амьтдын>80% нь үет хөлтөн байдаг байна. Үет хөлтөн нь тэнгис, цэнгэг ус, эх газарт түгээмэл тааралдана. Биеийн хэмжээ нь планктоноос эхлээд хэдэн метр хүрнэ. Одоо буй хамгийн том үет хөлтөн бол "Японы аалзан хавч" бөгөөд хөлнийх нь хэмжээг оруулан тооцвол нийт урт нь 3,5 метр хүрдэг байна.Үет хөлтөн нь тав болон түүнээс олон хөлтэй байна.
Үет хөлтний хүрээг дотор нь хэд хэдэн дэд хүрээ болгодог.
Заламгай амьсгалууртан (Branchiata) Трахейтан (Tracheata) Трилобит хэлбэртэн (Trilobitomorpha) Хелицертэн (Chelicerata) зэрэг.अर्थोपोडा( अंंग्रेजी भाषा: Arthropoda|) एक मेरुदण्ड नभएको जीवहरूको फाइलम हो । यसमा पर्ने जीवहरूको जोर्नीदार खुुुुट्टहरू हुन्छन। यिनिहरू मध्ये केही जमिनमा, केेही पानीमा र केेही हावामा रहन्छन्। यो जन्तुु विज्ञानको सबैभन्दा बढी विविधता भएको फाइलम हो। यसमा पाँँच वटा सव-फाइलमहरू पर्छन् ती हुन्
१• क्रुुस्टासिया(Crustacea)
२• मेरियापोडा(Meriapoda)
३• इन्सेक्ट्रा(Insectra)
४• एराक्निडा(Arachnida)
सन्धिपाद (अर्थोपोडा) प्राणी जगत का सबसे बड़ा संघ है। पृथ्वी पर सन्धिपाद की लगभग दो तिहाई जातियाँ हैं, इसमें कीट भी सम्मिलित हैं। इनका शरीर सिर, वक्ष और उदर में बँटा रहता है। शरीर के चारों ओर एक खोल जैसी रचना मिलती है। प्रायः सभी खंडों के पार्श्व की ओर एक संधियुक्त शाखांग होते हैं। सिर पर दो संयुक्त नेत्र होते हैं। ये जन्तु एकलिंगी होते हैं और जल तथा स्थल दोनों स्थानों पर मिलते हैं। तिलचट्टा, मच्छर, मक्खी, शतपाद, झिंगा, केकड़ा आदि इस संघ के प्रमुख जन्तु हैं।
सन्धिपाद (अर्थोपोडा) प्राणी जगत का सबसे बड़ा संघ है। पृथ्वी पर सन्धिपाद की लगभग दो तिहाई जातियाँ हैं, इसमें कीट भी सम्मिलित हैं। इनका शरीर सिर, वक्ष और उदर में बँटा रहता है। शरीर के चारों ओर एक खोल जैसी रचना मिलती है। प्रायः सभी खंडों के पार्श्व की ओर एक संधियुक्त शाखांग होते हैं। सिर पर दो संयुक्त नेत्र होते हैं। ये जन्तु एकलिंगी होते हैं और जल तथा स्थल दोनों स्थानों पर मिलते हैं। तिलचट्टा, मच्छर, मक्खी, शतपाद, झिंगा, केकड़ा आदि इस संघ के प्रमुख जन्तु हैं।
সন্ধিপদী প্ৰাণী হৈছে বহিঃকংকাল যুক্ত অমেৰুদণ্ডী প্ৰাণী, খণ্ডিত দেহ আৰু উপাংগবোৰ সন্ধিযুক্ত বা জোৰালগা। পতংগ আদিকে ধৰি সন্ধিপদীবোৰ, আৰ্থপডা (Arthropoda) পৰ্বৰ অন্তৰ্গত। সন্ধিপদীৰ প্ৰধান লক্ষণ হৈছে, সিঁহতৰ পদবোৰ (ভৰি) জোৰালগা আৰু α-কাইটিন আৰু/নাইবা কেলচিয়াম কাৰ্বনেটৰ দ্বাৰা গঠিত কিউটিকল্ থাকে। সন্ধিপদীৰ দেহটো কেইবাটাও খণ্ডেৰে গঠিত আৰু প্ৰতিটো খণ্ডতে এজোৰা উপাংগ থাকে। সন্ধিপদীবোৰে পৰ্যায়ক্ৰমে মোট সলাই সিঁহতৰ কঠিন খোলাটো পৰিবৰ্তন কৰি থাকে। পৰিৱেশৰ লগত খাপ খাব পৰা বিশেষ যোগ্যতাৰ ফলত ইহঁত আটাইতকৈ বেছি প্ৰজাতিযুক্ত জীৱ বিচাপে পৰিগণিত হৈছে। বৰ্তমানলৈকে প্ৰায় এক নিযুটতকৈও অধিক সন্ধিপদী প্ৰাণীৰ বিষয়ে জনা যায়, যিটো জ্ঞাত জীৱিত প্ৰাণীৰ প্ৰায় ৮০% ত কৈও অধিক, ইয়াৰে কিছুমান, অন্য প্ৰাণী জীয়াই থাকিব নোৱাৰা শুকাম পৰিৱেশতো বসবাস কৰে। ইহঁতৰ আকাৰ আনুবীক্ষণিক শৈৱালৰ পৰা কেইবা মিটাৰ পৰ্যন্ত দীঘল হব পাৰে। সন্ধিপদীৰ প্ৰাথমিক আভ্যন্তৰিক গহ্বৰটোকহিম'চেল বোলে, তাতে ইহঁতৰ আভ্যন্তৰিণ অংগবোৰ থাকে, আৰু ইয়াৰ মাজেৰেই হিম'লিম্ফ - তেজৰ সমধৰ্মী - চলাচল কৰে; ইহঁত মুক্ত পৰিবহন তন্ত্ৰী। বাহ্যক অংগবোৰৰ দৰই, সন্ধিপদীৰ আভ্যন্তৰিণ অংগবোৰো খণ্ডযুক্ত। সিঁহতৰ স্নায়ু তন্ত্ৰ "চামৰা-সদৃশ", যুৰীয়া পৃষ্ঠীয় স্নায়ু ৰজ্জু প্ৰতিটো খণ্ডৰ মাজেৰে গৈ প্ৰতিটো খণ্ডতে যুৰীয়া গেংলিয়া গঠন কৰে। সিঁহতৰ মুণ্ড বহুতো খণ্ডেৰে গঠিত, আৰু মগজুটো এই খণ্ডবোৰত থকা গেংলিয়াবোৰ লগ লাগি গঠন হয় আৰু খাদ্যনলীৰ চাৰিওফালে আগুৰি থাকে। সন্ধিপদীবোৰ বসবাস কৰা পৰিৱেশ আৰু উপপৰ্বৰ ওপৰত ভিত্তি কৰি সিহঁতৰ শ্বাসতন্ত্ৰ আৰু ৰেচন তন্ত্ৰৰ ভিন্নতা পৰিলক্ষিত হয়।
দেহৰ প্ৰতিটো খণ্ডত এজোৰকৈ উপাঙ্গ থাকে। ইহঁতৰ দেহ ত্ৰিস্তৰীয় আৰু দেহত প্ৰকৃত চিলম বা গহ্বৰ আছে। ইহঁতৰ পাচন তন্ত্ৰ সম্পূৰ্ণ আৰু ই মুখছিদ্ৰৰ পৰা পায়ুছিদ্ৰ পৰ্যন্ত বিস্তৃত। দেহৰ পৃষ্ঠদেশত হৃৎপিণ্ড আৰু ধমনীৰ দ্বাৰা মুক্ত সংবহনতন্ত্ৰ গঠিত। ইহঁতৰ দেহত বুক গিল (book gill), বুক লাঙ (book lung) বা শ্বাসনলী (trachea) ইত্যাদিয়ে শ্বসন অঙ্গৰ কাম কৰে। মালপিজিয়ান নলিকা (malpighian tubule), সেউজীয়া গ্ৰন্থি (green gland), কক্সাল গ্ৰন্থি (coxal gland) ইত্যাদিয়ে ৰেচন অঙ্গৰ কাম কৰে। স্নায়ুতন্ত্ৰ উন্নত ধৰণৰ, অৰ্থাৎ মস্তিষ্ক আৰু এযোৰ নিৰেট স্নায়ুৰজ্জু, বক্ষ আৰু উদৰ গেংগ্লিয়াৰ দ্বাৰা গঠিত হয়। ইহঁত সাধাৰণত্ একলিঙ্গ প্ৰাণী। যৌন দ্বিৰূপতা দেখা যায়। পূঞ্জাক্ষি বা সৰলাক্ষী থাকে।
কেঁকোৰা, মিছামাছ, ক্ৰেফিছ আদি ক্ৰাষ্টাচিয়ান বহু যুগৰে পৰা মানুহৰ খাদ্য হিচাপে ব্যৱহাৰ হৈ আহিছে আৰু বৰ্তমান ব্যৱসায়িক ভাৱে ইহঁতৰ যথেষ্ট চাহিদা আছে।[3] পতংগ আৰু ইহঁতৰ পলুবোৰ বিশ্বজুৰি কেঁচাকৈ বা সিজাই মাংস হিচাপে গ্ৰহণ কৰা হয়। কিন্তু কিছুমান ইউৰোপীয়ান, হিন্দু আৰু ইছলাম ধৰ্মী সমাজত কমকৈ ব্যৱহাৰ কৰা হয়।[4][5] কম্বডিয়াত সিদ্ধ টেৰেণ্টুলাক অতি সুস্বাদু আহাৰ হিচাপে গণ্য কৰা হয়।[6][7][8] আৰু দক্ষিণ ভেনেজুৱেলাৰ পায়াৰোৱা (Piaroa) জনগোষ্ঠীয়ে ইহঁতৰ মুখ্য প্ৰতিৰক্ষা তন্ত্ৰ অৰ্থাৎ শুং বোৰ গুচাই ভক্ষণ কৰে।[9]
কৃষি ক্ষেত্ৰতো সন্ধিপদী প্ৰাণীৰ অৱদান উল্লেখযোগ্য। পৰাগ সংযোগৰ মূল বাহক হিচাপে ইহঁতে খাদ্য উৎপাদন তথা উদ্ভিদৰ প্ৰজননত অতি গুৰুত্বপূৰ্ণ ভূমিকা পালন কৰে।[10] পৰাগ সংযোগৰ উপৰিও, মৌ মাখিয়ে মৌ উৎপাদন কৰে, এই মৌ ৰস হৈছে বহুতো উদ্যোগ আৰু আন্তঃৰাষ্ট্ৰীয় ব্যৱসায়ৰ মূল উপাদান।[11]
মধ্য আমেৰিকাত মায়া সভ্যতাত এটা পতংগ প্ৰজাতিৰ পৰা ৰঙা ডাই উৎপাদন কৰা হৈছিল।[12]
সন্ধিপদী প্ৰাণীয়ে মানুহৰ বহু উপকাৰ কৰে যদিও যথেষ্ট অপকাৰো সাধন কৰে। পতংগৰ পৰাই বহুতো সংক্ৰামক ৰোগ বিয়পে।
বহুতো সন্ধিপদী বিষাক্ত, ইহঁতৰ দংশন আৰু কামোৰত মানুহৰ ওপৰত বিভিন্ন প্ৰভাৱ পৰে। বহুতো পতংগই দংশন কৰাৰ ফলৰ বিভিন্ন ৰোগৰ সৃষ্টি হয়। কিছুমান পতংগই মানুহৰ তেজ শোষণ কৰে।[13] টিক (Tick)ৰ পৰা মানুহৰ টিক পেৰালাইছিছ আৰু বহুতো পৰজীৱি−জনিত ৰোগ হয়।[14] কিছুমান পতংগ তথা চাঁহীয়ে কামুহিলে খজুৱতী হয়,[15] আৰু অন্য কিছুমানে এলাৰ্জি ৰোগ, যেনে হেই ফিভাৰ, এজমা, আৰু এস্থমা আদি সৃষ্টি কৰে।[16]
আনহাতে বহুতো সন্ধিপদী বিশেষকৈ পতংগ আৰু চাঁহী (মাইট) কৃষি তথা অৰণ্যত অপকাৰী কীট হিচাপে যথেষ্ট ক্ষতিসাধনো কৰে।[17][18] ভাৰোৱা ডেষ্ট্ৰাক্টৰ (Varroa destructor) নামৰ চাঁহী বিধ বিশ্বজুৰি মৌ পালক সকলৰ বাবে এটা আটাইতকৈ ডাঙৰ সমস্যাত পৰিণত হৈছে।[19] এই কীট সমূহ নিয়ন্ত্ৰণৰ বাবে ব্যৱহাৰ কৰা কীতনাশক দৰৱে মানুহৰ স্বাস্থ্যৰ যথেষ্ট ক্ষতি সাধন কৰাৰ লগতে জীৱ বৈচিত্ৰৰ ওপৰতো প্ৰভাৱ পেলাই।[20] বৰ্তমান বিজ্ঞানীসকলে কিছুমান চিকাৰী চাঁহী ব্যৱহাৰ কৰি অপকাৰী চাঁহী নিয়ন্ত্ৰণৰ কৌশল আবিস্কাৰ কৰিছে।[21][22]
সন্ধিপদী প্ৰাণী হৈছে বহিঃকংকাল যুক্ত অমেৰুদণ্ডী প্ৰাণী, খণ্ডিত দেহ আৰু উপাংগবোৰ সন্ধিযুক্ত বা জোৰালগা। পতংগ আদিকে ধৰি সন্ধিপদীবোৰ, আৰ্থপডা (Arthropoda) পৰ্বৰ অন্তৰ্গত। সন্ধিপদীৰ প্ৰধান লক্ষণ হৈছে, সিঁহতৰ পদবোৰ (ভৰি) জোৰালগা আৰু α-কাইটিন আৰু/নাইবা কেলচিয়াম কাৰ্বনেটৰ দ্বাৰা গঠিত কিউটিকল্ থাকে। সন্ধিপদীৰ দেহটো কেইবাটাও খণ্ডেৰে গঠিত আৰু প্ৰতিটো খণ্ডতে এজোৰা উপাংগ থাকে। সন্ধিপদীবোৰে পৰ্যায়ক্ৰমে মোট সলাই সিঁহতৰ কঠিন খোলাটো পৰিবৰ্তন কৰি থাকে। পৰিৱেশৰ লগত খাপ খাব পৰা বিশেষ যোগ্যতাৰ ফলত ইহঁত আটাইতকৈ বেছি প্ৰজাতিযুক্ত জীৱ বিচাপে পৰিগণিত হৈছে। বৰ্তমানলৈকে প্ৰায় এক নিযুটতকৈও অধিক সন্ধিপদী প্ৰাণীৰ বিষয়ে জনা যায়, যিটো জ্ঞাত জীৱিত প্ৰাণীৰ প্ৰায় ৮০% ত কৈও অধিক, ইয়াৰে কিছুমান, অন্য প্ৰাণী জীয়াই থাকিব নোৱাৰা শুকাম পৰিৱেশতো বসবাস কৰে। ইহঁতৰ আকাৰ আনুবীক্ষণিক শৈৱালৰ পৰা কেইবা মিটাৰ পৰ্যন্ত দীঘল হব পাৰে। সন্ধিপদীৰ প্ৰাথমিক আভ্যন্তৰিক গহ্বৰটোকহিম'চেল বোলে, তাতে ইহঁতৰ আভ্যন্তৰিণ অংগবোৰ থাকে, আৰু ইয়াৰ মাজেৰেই হিম'লিম্ফ - তেজৰ সমধৰ্মী - চলাচল কৰে; ইহঁত মুক্ত পৰিবহন তন্ত্ৰী। বাহ্যক অংগবোৰৰ দৰই, সন্ধিপদীৰ আভ্যন্তৰিণ অংগবোৰো খণ্ডযুক্ত। সিঁহতৰ স্নায়ু তন্ত্ৰ "চামৰা-সদৃশ", যুৰীয়া পৃষ্ঠীয় স্নায়ু ৰজ্জু প্ৰতিটো খণ্ডৰ মাজেৰে গৈ প্ৰতিটো খণ্ডতে যুৰীয়া গেংলিয়া গঠন কৰে। সিঁহতৰ মুণ্ড বহুতো খণ্ডেৰে গঠিত, আৰু মগজুটো এই খণ্ডবোৰত থকা গেংলিয়াবোৰ লগ লাগি গঠন হয় আৰু খাদ্যনলীৰ চাৰিওফালে আগুৰি থাকে। সন্ধিপদীবোৰ বসবাস কৰা পৰিৱেশ আৰু উপপৰ্বৰ ওপৰত ভিত্তি কৰি সিহঁতৰ শ্বাসতন্ত্ৰ আৰু ৰেচন তন্ত্ৰৰ ভিন্নতা পৰিলক্ষিত হয়।
ਆਰਥਰੋਪੋਡਇੱਕ ਰੀੜ੍ਹਰਹਿਤ ਜੰਤੂ ਹੈ ਜਿਸਦੇ ਸਰੀਰ ਦੇ ਚਾਰੇ ਪਾਸੇ ਇੱਕ ਖੋਲ ਵਰਗੀ ਰਚਨਾ ਮਿਲਦੀ ਹੈ। ਇਨ੍ਹਾਂ ਦਾ ਸਰੀਰ ਸਿਰ, ਛਾਤੀ ਅਤੇ ਉਦਰ ਵਿੱਚ ਵੰਡਿਆ ਹੁੰਦਾ ਹੈ। ਆਰਥਰੋਪੋਡ ਆਰਥਰੋਪੋਡਾ ਸੰਘ ਦੇ ਰੁਕਨ ਹੁੰਦੇ ਹਨ (ਯੂਨਾਨੀ ਭਾਸ਼ਾ ἄρθρον árthron, "ਜੋੜ", ਅਤੇ πούς pous (gen. ਪੋਡੋਜ ਤੋਂ), ਅਰਥਾਤ "ਪੈਰ" ਜਾਂ "ਲੱਤ", ਇਸ ਦਾ ਮਿਲਵਾਂ ਅਰਥ "ਜੁੜੀਆਂ ਲੱਤਾਂ" ਹੈ।[2]) ਇਹ ਪ੍ਰਾਣੀ ਜਗਤ ਦਾ ਸਭ ਤੋਂ ਵੱਡਾ ਸੰਘ ਹੈ। ਧਰਤੀ ਤੇ ਲਗਪਗ ਦੋ ਤਿਹਾਈ ਜਾਤੀਆਂ ਆਰਥਰੋਪੋਡ ਹਨ।
સંધિપાદ (અંગ્રેજી: Arthropod) એ પ્રાણીસૃષ્ટિનો સૌથી મોટો સમુદાય છે. આ પ્રકારના પ્રાણીઓ સાંધાવાળા ઉપાંગો ધરાવે છે તેથી તે સંધિપાદ કહેવામાં આવે છે. જીવનની શક્યતા હોય તેવી કોઈ પણ જગ્યાએ આ સમુદાયનાં પ્રાણીઓ જોવા મળે છે. તેઓ જમીનમાં, જમીન ઉપર, મીઠા પાણીમાં, ખારા પાણીમાં-સમુદ્રમાં કોઈ પણ ઊંડાઈએ, ઉષ્ણકટિબંધ તેમજ શીત કટિબંધ પ્રદેશોમાં અને હવામાં વસવાટ કરે છે. તેમાંના કેટલાક પ્રાણીઓ સમુહમાં જીવન ગાળે છે જેવા કે, મધમાખી, કીડી, ઊધઈ વગેરે કીટકો. આ સમુદાયનાં સામાન્ય પ્રાણીઓ આદિસંધિપાદ (ઉદા. પેરિપેટસ); સ્તરકવછી (ઉદા. કરચલાં, જિંગા); કીટક (ઉદા. પતંગિયાં, માખી, વંદો વગેરે) અને અષ્ટપાદ (ઉદા. કરોળિયા, વીંછી વગેરે) છે. [૧]
આ સમુદાયનાં પ્રાણીઓમાં કાઈટીનયુક્ત બાહ્યકંકાલ અને સાંધાવાળા પગ તે મુખ્ય વિશેષતા છે. શરીર દ્રિપાર્શ્વ સમરચના ધરાવે છે. તેઓ તેમનું જીવન સ્વોપજીવી કે પરોપજીવી તરીકે ગુજારે છે. આ પ્રાણીઓ ત્રિગર્ભસ્તરીય અને ખંડયુક્ત શરીરવાળાં હોય છે. ખંડો બહારથી જોડાયેલા હોય છે અને શરીર મુખ્ય ત્રણ ભાગોમાં વહેંચાયેલું દેખાય છે: અગ્રથી પશ્વબાજુ તરફ જતાં આ વિભાગોને અનુક્રમે શિર્ષ, ઉરસ અને ઉદર કહેવામાં આવે છે. કેટલાક પ્રાણીઓમાં શિર્ષ અને ઉરસ જોડાઈ સંયુક્ત ભાગરૂપે શિરોરસ બને છે તો કેટલાકમાં ઉરસ તેમજ ઉદર જોડાઇ જઈ ઉરોદર બને છે. મહદંશે આ પ્રાણીઓનાં સાંધાવાળા ઉપાંગો પ્રત્યેક ખંડમાં એક જોડરૂપે આવેલા હોય છે. શરીરમાં રેખિત સ્નાયુઓ વિપુલ પ્રમાણમાં હોય છે. શરિર પરનું બાહ્યકંકાલ કાઈટીનનું બનેલું હોય મજબૂત હોય છે. આ બાહ્યકંકાલ અવારનવાર શરીર પરથી નિર્મોચન ક્રિયા દ્વારા ઉતરી જાય છે અને નવું બાહ્યકંકાલ અધિચર્મમાંથી કાઈટીનનાં સ્ત્રાવથી તૈયાર થાય છે. અગ્રાંત્ર, મધ્યાંત્ર અને પશ્ચાંત્ર એમ ત્રણ ભાગમાં વગેંચાયેલું વિકસિત પાચનતંત્ર તેઓ ધરાવે છે. આ પ્રાણીઓમાં શ્વસન માટે ઝાલરો, શ્વસનનલિકાઓ, ફેફસાપોથી વગેરે વિશિષ્ટ અવયવો આવેલાં છે. રુધિરાભિસરણ તંત્ર ખુલ્લા પ્રકારમુ હોય છે. શરીરગુહાનું સ્થાન રુધિરગુહાએ લીધેલુ હોય છે, તેથી ઉત્સર્ગિકાઓનો અભાવ હોય છે. હ્રદય પૃષ્ઠબાહુએ આવેલું હોય છે. ઉત્સર્જન હરિતપિંડ, માલ્પિધિની નલિકાઓ વગેરે અવયવો દ્વારા થાય છે. નિર્મોચન ક્રિયા પણ ઉત્સર્જનમાં ભાગ ભજવે છે. આ સમુદાયના બધા પ્રાણીઓ એકલિંગી હોય છે. તેમનામાં ફલન આંતરફલનથી થાય છે. મહંદશે તેઓ ઈંડામાંથિ રૂપાંતરણ પામી પુખ્તાવસ્થા પ્રાપ્ત કરે છે..[૨]
સંધિપાદ (અંગ્રેજી: Arthropod) એ પ્રાણીસૃષ્ટિનો સૌથી મોટો સમુદાય છે. આ પ્રકારના પ્રાણીઓ સાંધાવાળા ઉપાંગો ધરાવે છે તેથી તે સંધિપાદ કહેવામાં આવે છે. જીવનની શક્યતા હોય તેવી કોઈ પણ જગ્યાએ આ સમુદાયનાં પ્રાણીઓ જોવા મળે છે. તેઓ જમીનમાં, જમીન ઉપર, મીઠા પાણીમાં, ખારા પાણીમાં-સમુદ્રમાં કોઈ પણ ઊંડાઈએ, ઉષ્ણકટિબંધ તેમજ શીત કટિબંધ પ્રદેશોમાં અને હવામાં વસવાટ કરે છે. તેમાંના કેટલાક પ્રાણીઓ સમુહમાં જીવન ગાળે છે જેવા કે, મધમાખી, કીડી, ઊધઈ વગેરે કીટકો. આ સમુદાયનાં સામાન્ય પ્રાણીઓ આદિસંધિપાદ (ઉદા. પેરિપેટસ); સ્તરકવછી (ઉદા. કરચલાં, જિંગા); કીટક (ઉદા. પતંગિયાં, માખી, વંદો વગેરે) અને અષ્ટપાદ (ઉદા. કરોળિયા, વીંછી વગેરે) છે.
கணுக்காலிகள் ((![]() ஒலிப்பு ) (Arthropod) என்பவை விலங்குகளின் பிரிவில் ஒரு மிகப் பெரிய தொகுதி. ஆறு கால்கள் கொண்ட பூச்சிகளும், வண்டுகளும், தேளினங்களும், எட்டுக்கால் பூச்சி, சிலந்திகளும், நண்டுகளும் அடங்கிய மிகப் பெரிய உயிரினப்பகுப்பு. இன்று மனிதர்களால் அறிந்து விளக்கப்பட்ட விலங்குகளில் 80% க்கும் மேல் கணுக்காலிகளைப் பற்றியவைதாம். கணுக்காலிகளை அறிவியலில் ஆர்த்ரோபோடா (Arthropoda) என்பர். இது கிரேக்க மொழியில் உள்ள ἄρθρον ஆர்த்ரோ (= இணைக்கப்பட்ட, கணு) என்னும் சொல்லும் ποδός (போடொஸ் = கால்) என்னும் சொல்லும் சேர்ந்து ஆக்கப்பட்டது. கணுக்காலிகளுக்குக் கடினமான புறவன்கூடு உண்டு. இவற்றின் உடல், பகுதி பகுதியாக, அதாவது கண்டங்களாகப் பிரிக்கப்பட்டுள்ள அமைப்பு கொண்டது. இவற்றின் கால்கள் இணை இணையாக ஒவ்வொரு உடற் கண்டத்திலும் உள்ளன. பிப்ரவரி 29, 2016 அன்று, 2013ல் தெற்குச் சீனாவில் கண்டெடுக்கப்பட்ட சுமார் 52 கோடி (520 மில்லியன்) ஆண்டுகள் பழமையான கணுக்காலியின் தொல்படிம மேட்டின் கூடுதல் படிமம் மற்றும் கள ஆய்வில் அதன் வயிற்றின் கீழ்ப்பகுதியில் செல்லும் கயிறு போன்ற ஒன்று அதன் கீழ்நரம்பு வடம் எனப் புரசீடிங்சு ஓப் நேச்சரல் அகாடமி ஆப் சயன்சசு இதழில் அறிக்கை வெளியிட்டுள்ளனர். [1]
ஒலிப்பு ) (Arthropod) என்பவை விலங்குகளின் பிரிவில் ஒரு மிகப் பெரிய தொகுதி. ஆறு கால்கள் கொண்ட பூச்சிகளும், வண்டுகளும், தேளினங்களும், எட்டுக்கால் பூச்சி, சிலந்திகளும், நண்டுகளும் அடங்கிய மிகப் பெரிய உயிரினப்பகுப்பு. இன்று மனிதர்களால் அறிந்து விளக்கப்பட்ட விலங்குகளில் 80% க்கும் மேல் கணுக்காலிகளைப் பற்றியவைதாம். கணுக்காலிகளை அறிவியலில் ஆர்த்ரோபோடா (Arthropoda) என்பர். இது கிரேக்க மொழியில் உள்ள ἄρθρον ஆர்த்ரோ (= இணைக்கப்பட்ட, கணு) என்னும் சொல்லும் ποδός (போடொஸ் = கால்) என்னும் சொல்லும் சேர்ந்து ஆக்கப்பட்டது. கணுக்காலிகளுக்குக் கடினமான புறவன்கூடு உண்டு. இவற்றின் உடல், பகுதி பகுதியாக, அதாவது கண்டங்களாகப் பிரிக்கப்பட்டுள்ள அமைப்பு கொண்டது. இவற்றின் கால்கள் இணை இணையாக ஒவ்வொரு உடற் கண்டத்திலும் உள்ளன. பிப்ரவரி 29, 2016 அன்று, 2013ல் தெற்குச் சீனாவில் கண்டெடுக்கப்பட்ட சுமார் 52 கோடி (520 மில்லியன்) ஆண்டுகள் பழமையான கணுக்காலியின் தொல்படிம மேட்டின் கூடுதல் படிமம் மற்றும் கள ஆய்வில் அதன் வயிற்றின் கீழ்ப்பகுதியில் செல்லும் கயிறு போன்ற ஒன்று அதன் கீழ்நரம்பு வடம் எனப் புரசீடிங்சு ஓப் நேச்சரல் அகாடமி ஆப் சயன்சசு இதழில் அறிக்கை வெளியிட்டுள்ளனர். [1]
கணுக்காலிகளில் பிளாங்க்டன் (plankton) வகையைச் சேர்ந்தவையின் 0.25 மி.மீ. அளவில் இருந்து, 20 கிலோ கிராம் எடையுடன் 4 மீட்டர் (12-13 அடிகள்) கால் விரிப்பு அகலம் கொண்ட மிகப்பெரிய நிப்பானிய எட்டுக்கால் நண்டு வரை மிகப்பல வகைகள் உள்ளன.
கணுக்காலிகளை வேறுபடுத்திக் காட்டும் முக்கிய மூன்று இயல்புகள்:
இவை பார்வைக்காகக் கூட்டுக்கண்களையும் ஒசிலி எனும் பார்வைப் புலனங்கத்தையும் பயன்படுத்துகின்றன. பூச்சிகளில் கூட்டுக்கண்ணே பிரதானமான பார்வைப் புலனங்கமென்றாலும், சிலந்திகளில் ஒசிலிகளே பிரதான பார்வைப் புலனங்கங்களாகும். பூச்சிகளின் ஒசிலிக்களால் ஒளி வரும் திசையை மாத்திரமே கணிக்க முடியும். எனினும் சிலந்திகளின் ஒசிலிக்களால் முழு உருவத்தையும் கணித்துக்கொள்ள முடியும். கேம்பிரியன் காலப்பகுதியிலிருந்து கணுக்காலிகள் பூமியில் உள்ளன. அவற்றின் உடலகக் கட்டமைப்பின் சிறப்புத் தன்மையால், விலங்குகளில் அதிகளவான இனங்களும், அதிகளவான தனியன்களும் கணுக்காலிகளாக உள்ளன. எனவே இவையே விலங்குக் கூட்டங்களுள் அதிக வெற்றியுடைய கூட்டமாக உள்ளன. எனினும் உலகிலுள்ள உயிரினங்களுள் பாக்டீரியாக்களே அதிக இனங்களையும், அதிக தனியன்களையும் கொண்ட கூட்டமாகும்.
கணுக்காலிகள் மனித வாழ்வில் பல்வேறு தாக்கங்களைக் காட்டுகின்றன. நாம் உண்ணும் உணவில் முக்கியப் பங்கை வகிக்கும் இறால், நண்டு என்பன கணுக்காலிகளே. ஒவ்வொரு வருடமும் பயிர்களின் அறுவடைக்கு முன்னமும் பின்னரும் பயிரையும் விளைச்சலையும் அழித்துப் பல கோடி ரூபாய் நஷ்டத்தை ஏற்படுத்தும் பீடைகளில் அதிகமானவை கணுக்காலிகளே. தாவரங்களின் தேனை உறிஞ்சி அவற்றின் மகரந்தச் சேர்க்கைக்கு உதவும் வண்ணத்துப் பூச்சிகளும், தேனீக்களும் கணுக்காலிகளாகும். எறும்புகளும் இக்கூட்டத்தைச் சேர்ந்தவையே ஆகும். எனவே உயிரினப் பல்வகைமையில் கணுக்காலிகள் முக்கியப் பங்கை வகிக்கின்றன.
ஒரு கணிப்பின் படி இதுவரை அறியப்பட்ட கணுக்காலி இனங்களின் எண்ணிக்கை கிட்டத்தட்ட 1,170,000 ஆகும். எனினும் புவியில் வாழும் கணுக்காலி இனங்களின் உண்மையான எண்ணிக்கை 5-10 மில்லியன்கள் எனக் கணிப்பிடப்பட்டுள்ளது. இவற்றின் எண்ணிக்கை ஒரு பிரதேசத்தின் சராசரி இனப்பல்வகைமையைக் கொண்டே கணிக்கப்படுகின்றது. எனினும் துருவப் பிரதேசங்களை விட அயனமண்டலப் பிரதேசங்களின் கணுக்காலி இனங்களின் பல்வகைமை மிக அதிகமாகும். 1992ஆம் ஆண்டில் மேற்கொள்ளப்பட்ட ஆய்வின் படி கோஸ்டா ரிகா எனும் சிறிய அயன மண்டல நாட்டில் மாத்திரம் 365,000 கணுக்காலி இனங்கள் உள்ளன. எனினும் துருவங்களுக்கருகிலுள்ள மிகப்பெரிய நாடுகளிலும் இவ்வெண்ணிக்கை இதை விடக் குறைவாகவும் காணப்படலாம்.
கணுக்காலிகளின் மிக முக்கியமான தனித்துவமான இயல்பு மூட்டுள்ள தூக்கங்களைக் கொண்டிருத்தலும், கைட்டினாலான புறவன்கூட்டைக் கொண்டிருத்தலுமாகும். இம்மூட்டுள்ள தூக்கங்கள் கணுக்காலிகளின் இடப்பெயர்ச்சி, உணவுண்ணல், புலனுகர்ச்சி போன்றவற்றிற்கு உதவும். இவை இருபக்கச் சமச்சீருள்ள, முப்படை கொண்ட, அனுப்பாத்து துண்டுபட்ட உடலுடைய விலங்குகளாகும். இவற்றிற்குப் பூரணமான உணவுக்கால்வாய் உண்டு. கணுக்காலிகள் திறந்த குருதிச்சுற்றோட்டத் தொகுதியை உடையவையாகும். இவை ஒடுக்கப்பட்ட உடற்குழியையும் நன்கு விருத்தியடைந்த குருதிக் குழியையும் கொண்டுள்ளன. நன்கு விருத்தியடைந்த வரித்தசை காணப்படும். இக்கூட்டத்தில் தலையாகு செயலும், புறத்தே குறித்த எண்ணிக்கையான உடற்துண்டங்கள் இணைவதன் மூலம் தக்குமா ஆகும் செயலும் சிறப்படைந்து காணப்படுபவை. பூச்சிகளில் தலை, நெஞ்சறை, வயிறு என மூன்று தக்குமாக்களும் (Tagma) நண்டு, இறால் பொன்ற கிரஸ்டீசியன்களில் தலைநெஞ்சு, வயிறு என இரண்டு தக்குமாக்களும் உள்ளன. முளையவிருத்தி இயல்புகளின் அடிப்படையில் கணுக்காலிகள் புரொட்டோஸ்டோம் விலங்குகளாகும். அதாவது இவற்றின் முளைய விருத்தியில் சமிபாட்டுத் தொகுதியில் முதலில் விருத்தியடைவது அவற்றின் வாயாகும். எனினும் முள்ளந்தண்டுளிகள் போன்ற Deutorosome விலங்குகளில் குதமே முதலில் விருத்தியடைகின்றது. வாழும் சூழலுக்கேற்றபடி இனங்களின் சுவாசத்தொகுதியும் இனப்பெருக்க முறையும் வேறுபடுகின்றன. நீரில் வாழும் கணுக்காலிகள் பொதுவாகப் புறக்கருக்கட்டல் முறையையும், நில வாழ் கணுக்காலிகள் அகக்கருக்கட்டல் முறையையும் பின்பற்றுகின்றன.
கணுக்காலிகளின் புறவன்கூடு ஓரளவுக்கு மிகவும் பலமானதாகும். இது அவற்றை அதிக வெப்பநிலை, நீரிழப்பு, பாதகமான கதிர்வீச்சிலிருந்து பாதுகாக்கின்றது. எனினும் இவ்வாறு உறுதியான புறவன்கூட்டுடன் கணுக்காலியொன்றால் வளர்ச்சியடைய முடியாது. எனவே இவற்றின் வளர்ச்சிக்காலத்தில் புறவன்கூட்டை அகற்றிய பின்னரே வளர்கின்றன. வளர்ந்த பின்னர் புதிய உறுதியான புறவன்கூட்டை ஆக்குகின்றன. அடுத்த வளர்ச்சிக் கட்டத்தில் மீண்டும் இவ்வாறு தோல் கழற்றப்படும். எனவே இவற்றின் வளர்ச்சியொழுங்கு நேரியதாகக் காணப்படாது. புறத்தோலை அகற்ற முன்னர் இவை உணவுண்பதை நிறுத்தி விடும். பின்னர் நொதியங்களைச் சுரந்து புறவன்கூடு உடலுடன் பொருந்தியுள்ள பகுதியை நீக்கி விடுகின்றன. அதிகளவான வளியையும், நீரையும் உடலினுள் எடுத்து உடலை ஊதச் செய்து பழைய புறவன்கூட்டை உடைத்து நீக்கி விடுகின்றன. அனேகமான கணுக்காலிகள் தங்களது நீக்கப்பட்ட பழைய புறவன்கூட்டை உண்டு விடுகின்றன. பழைய புறவன்கூட்டை நீக்கும் போதே மெல்லிய உறுதியற்ற புதிய புறவன்கூடு உருவாகியிருக்கும். எனினும் இது உறுதி பெறும் வரை கணுக்காலிக்குச் சிறந்த பாதுகாப்பை அளிக்காது. எனவே வளர்ச்சியடையும் காலத்திலேயே அனேகமான கணுக்காலிகள் ஊனுண்ணிகளால் தாக்கப்பட்டு இறக்கின்றன.
அனேகமான நீர்வாழ் கிரஸ்டேசியன்களில் புறக்கருக்கட்டல் நிகழும். அனைத்து நிலவாழ் கணுக்காலிகளிலும் அகக்கருக்கட்டலே நிகழ்கின்றது. அதாவது சூழ் பெண்ணங்கியின் உடலினுள்ளேயே இருக்கும் படியாக அசையும் விந்துக்கள் மூலம் கருக்கட்டப்படுவது. சில அங்கிகளில் மாத்திரம் கன்னிப்பிறப்பு காணப்படுகின்றது. அதாவது ஆணங்கியின் துணையின்றி சந்ததியை உருவாக்கல். கருக்கட்டலின் பின்னர் அனேகமான கணுக்காலிகள் முட்டையீனுகின்றன. சில தேள் இனங்கள் மட்டும் சூற்பிள்ளையீனல் (முட்டை தாயினுள்ளேயே பொரித்து குட்டி ஈனப்படல்) காணப்படுகின்றது. பல கணுக்காலிகள் உருமாற்ற விருத்தியைக் காண்பிக்கின்றன. சில குடம்பிப் பருவமுள்ள நிறையுருமாற்றத்தையும், சில அணங்குப் பருவமுள்ள குறையுருமாற்றத்தையும் காண்பிக்கின்றன. கிறஸ்டேசியன்களில் பொதுவாக முட்டையிலிருந்து நோப்பிளியஸ் எனும் குடம்பிப் பருவம் வெளிவரும். சில கணுக்காலிகளில் நேர்விருத்தி (முட்டையிலிருந்து சிறிய நிறையுடலி வெளிவரல்) முறையும் உள்ளது.
கணுக்காலிகள் ஐந்து பிரதான உபகணங்களுக்குள் வகைப்படுத்தப்படுகின்றன. இவற்றில் ஒரு கணம் இனமழிந்த உயிரினங்களை உள்ளடக்கியதாகும்.[2]
Panarthropoda
Onchyophora
Tardigrada
செலிசரேட்டா
மைரியபோடா
Ostracoda, Branchiura, Pentastomida, Mystacocarida
Branchiopoda, Copepoda, Malacostraca, Thecostraca
ஆறுகாலிகள் (Hexapoda)
கணுக்காலிகள் பெரும் பிரிவைச்சேர்ந்த ஓர் உயிரினம்-மெக்சிகோ டராண்ட்டூலா என்னும் சிலந்தி
கரும்பு வேர்வண்டு (Diaprepes abbreviatus) ஒரு வகை கணுக்காலி
எறும்புகளும் கணுக்காலிகள் தொகுப்பைச் சேர்ந்த உயிரினங்கள்தாம். படத்தில் உள்ளது ஆஸ்திரேலியாவைச் சேர்ந்த ஊன் உண்ணும் ஒருவகை எறும்பு (Iridomyrmex purpureus)
எட்டுக்கால் பூச்சி, தமிழ்நாடு
கணுக்காலிகள் ((![]() ஒலிப்பு ) (Arthropod) என்பவை விலங்குகளின் பிரிவில் ஒரு மிகப் பெரிய தொகுதி. ஆறு கால்கள் கொண்ட பூச்சிகளும், வண்டுகளும், தேளினங்களும், எட்டுக்கால் பூச்சி, சிலந்திகளும், நண்டுகளும் அடங்கிய மிகப் பெரிய உயிரினப்பகுப்பு. இன்று மனிதர்களால் அறிந்து விளக்கப்பட்ட விலங்குகளில் 80% க்கும் மேல் கணுக்காலிகளைப் பற்றியவைதாம். கணுக்காலிகளை அறிவியலில் ஆர்த்ரோபோடா (Arthropoda) என்பர். இது கிரேக்க மொழியில் உள்ள ἄρθρον ஆர்த்ரோ (= இணைக்கப்பட்ட, கணு) என்னும் சொல்லும் ποδός (போடொஸ் = கால்) என்னும் சொல்லும் சேர்ந்து ஆக்கப்பட்டது. கணுக்காலிகளுக்குக் கடினமான புறவன்கூடு உண்டு. இவற்றின் உடல், பகுதி பகுதியாக, அதாவது கண்டங்களாகப் பிரிக்கப்பட்டுள்ள அமைப்பு கொண்டது. இவற்றின் கால்கள் இணை இணையாக ஒவ்வொரு உடற் கண்டத்திலும் உள்ளன. பிப்ரவரி 29, 2016 அன்று, 2013ல் தெற்குச் சீனாவில் கண்டெடுக்கப்பட்ட சுமார் 52 கோடி (520 மில்லியன்) ஆண்டுகள் பழமையான கணுக்காலியின் தொல்படிம மேட்டின் கூடுதல் படிமம் மற்றும் கள ஆய்வில் அதன் வயிற்றின் கீழ்ப்பகுதியில் செல்லும் கயிறு போன்ற ஒன்று அதன் கீழ்நரம்பு வடம் எனப் புரசீடிங்சு ஓப் நேச்சரல் அகாடமி ஆப் சயன்சசு இதழில் அறிக்கை வெளியிட்டுள்ளனர்.
ஒலிப்பு ) (Arthropod) என்பவை விலங்குகளின் பிரிவில் ஒரு மிகப் பெரிய தொகுதி. ஆறு கால்கள் கொண்ட பூச்சிகளும், வண்டுகளும், தேளினங்களும், எட்டுக்கால் பூச்சி, சிலந்திகளும், நண்டுகளும் அடங்கிய மிகப் பெரிய உயிரினப்பகுப்பு. இன்று மனிதர்களால் அறிந்து விளக்கப்பட்ட விலங்குகளில் 80% க்கும் மேல் கணுக்காலிகளைப் பற்றியவைதாம். கணுக்காலிகளை அறிவியலில் ஆர்த்ரோபோடா (Arthropoda) என்பர். இது கிரேக்க மொழியில் உள்ள ἄρθρον ஆர்த்ரோ (= இணைக்கப்பட்ட, கணு) என்னும் சொல்லும் ποδός (போடொஸ் = கால்) என்னும் சொல்லும் சேர்ந்து ஆக்கப்பட்டது. கணுக்காலிகளுக்குக் கடினமான புறவன்கூடு உண்டு. இவற்றின் உடல், பகுதி பகுதியாக, அதாவது கண்டங்களாகப் பிரிக்கப்பட்டுள்ள அமைப்பு கொண்டது. இவற்றின் கால்கள் இணை இணையாக ஒவ்வொரு உடற் கண்டத்திலும் உள்ளன. பிப்ரவரி 29, 2016 அன்று, 2013ல் தெற்குச் சீனாவில் கண்டெடுக்கப்பட்ட சுமார் 52 கோடி (520 மில்லியன்) ஆண்டுகள் பழமையான கணுக்காலியின் தொல்படிம மேட்டின் கூடுதல் படிமம் மற்றும் கள ஆய்வில் அதன் வயிற்றின் கீழ்ப்பகுதியில் செல்லும் கயிறு போன்ற ஒன்று அதன் கீழ்நரம்பு வடம் எனப் புரசீடிங்சு ஓப் நேச்சரல் அகாடமி ஆப் சயன்சசு இதழில் அறிக்கை வெளியிட்டுள்ளனர்.
கணுக்காலிகளில் பிளாங்க்டன் (plankton) வகையைச் சேர்ந்தவையின் 0.25 மி.மீ. அளவில் இருந்து, 20 கிலோ கிராம் எடையுடன் 4 மீட்டர் (12-13 அடிகள்) கால் விரிப்பு அகலம் கொண்ட மிகப்பெரிய நிப்பானிய எட்டுக்கால் நண்டு வரை மிகப்பல வகைகள் உள்ளன.
கணுக்காலிகளை வேறுபடுத்திக் காட்டும் முக்கிய மூன்று இயல்புகள்:
கைட்டினாலான புறவன்கூடு துண்டுகளாலான உடல் மூட்டுள்ள தூக்கங்கள் (மூட்டுள்ள கால்கள்)இவை பார்வைக்காகக் கூட்டுக்கண்களையும் ஒசிலி எனும் பார்வைப் புலனங்கத்தையும் பயன்படுத்துகின்றன. பூச்சிகளில் கூட்டுக்கண்ணே பிரதானமான பார்வைப் புலனங்கமென்றாலும், சிலந்திகளில் ஒசிலிகளே பிரதான பார்வைப் புலனங்கங்களாகும். பூச்சிகளின் ஒசிலிக்களால் ஒளி வரும் திசையை மாத்திரமே கணிக்க முடியும். எனினும் சிலந்திகளின் ஒசிலிக்களால் முழு உருவத்தையும் கணித்துக்கொள்ள முடியும். கேம்பிரியன் காலப்பகுதியிலிருந்து கணுக்காலிகள் பூமியில் உள்ளன. அவற்றின் உடலகக் கட்டமைப்பின் சிறப்புத் தன்மையால், விலங்குகளில் அதிகளவான இனங்களும், அதிகளவான தனியன்களும் கணுக்காலிகளாக உள்ளன. எனவே இவையே விலங்குக் கூட்டங்களுள் அதிக வெற்றியுடைய கூட்டமாக உள்ளன. எனினும் உலகிலுள்ள உயிரினங்களுள் பாக்டீரியாக்களே அதிக இனங்களையும், அதிக தனியன்களையும் கொண்ட கூட்டமாகும்.
கணுக்காலிகள் மனித வாழ்வில் பல்வேறு தாக்கங்களைக் காட்டுகின்றன. நாம் உண்ணும் உணவில் முக்கியப் பங்கை வகிக்கும் இறால், நண்டு என்பன கணுக்காலிகளே. ஒவ்வொரு வருடமும் பயிர்களின் அறுவடைக்கு முன்னமும் பின்னரும் பயிரையும் விளைச்சலையும் அழித்துப் பல கோடி ரூபாய் நஷ்டத்தை ஏற்படுத்தும் பீடைகளில் அதிகமானவை கணுக்காலிகளே. தாவரங்களின் தேனை உறிஞ்சி அவற்றின் மகரந்தச் சேர்க்கைக்கு உதவும் வண்ணத்துப் பூச்சிகளும், தேனீக்களும் கணுக்காலிகளாகும். எறும்புகளும் இக்கூட்டத்தைச் சேர்ந்தவையே ஆகும். எனவே உயிரினப் பல்வகைமையில் கணுக்காலிகள் முக்கியப் பங்கை வகிக்கின்றன.
Melldrosogyon yw divellkeynoges y'n fylum Arthropoda. Yma a-dro dhe 1,230,000 ehen dheskrifys y'n bagas, ow komprehendya hwesker, ethgarrogyon ha krestenogyon.[1] Yma garrow mellek ha korf eskern a-ves dhedha ha rynnys yw aga horfow yn trehow.[2]
Melldrosogyon yw divellkeynoges y'n fylum Arthropoda. Yma a-dro dhe 1,230,000 ehen dheskrifys y'n bagas, ow komprehendya hwesker, ethgarrogyon ha krestenogyon. Yma garrow mellek ha korf eskern a-ves dhedha ha rynnys yw aga horfow yn trehow.
Үетэ хүлтэн (латаар 'Arthropoda' хүрээ, Грек: ἄρθρον гү, али arthron - "ниилмэл", ποδός гү, али podos - "хүл" гэһэн удхатай) хүрээнь хорхой шумуул, хабшаахай хэлбэритэн зэргые багтааһан амитанай аймагай хамагай томо хүрээ юм. Энэ хүрээндэ 1,500,000 оршом зүйл амитад багтана. Үетэ хүлтэнэй гол онсологиинь тэдыгээрэй бэень дор хаяжа нэгэ сегмент бүхы үелэһэн бүтэсэтэй байна. Хоол болбосоруулха системэнь үмэнэдэ, дунда, хойто гэһэн гурбан хэһэгһээ тогтоно. Хитин бүрхүүлээр бүрхэгдэжэ гадаада араг яһан үүдхэнэ.
Одоо оршон амитадай> 80% үетэ хүлтэн байдаг байна[1]. Үетэ хүлтэниинь тэнгис, сэнхир уһан, газарда түгөөмэл тааралдана. Бэеын хэмжээнь планктонһоо эхилээд хэдэн метр хүрэнэ. Одоо хамагайн томо үетэ хүлтэн бол "Японой аалзан хабшаахай" болоод хүлныхинь хэмжээе оруулан тоособол ниитэ утань 3,5 метр хүрэдэг байна. Гэхыдээ энэнь хүлдүү тэнэг хүндэ зорюулһан бүлгөө,
Үетэ хүлтэнэй хүрээе доторынь хэд хэдэн дэд хүреэн болгодог.
Үетэ хүлтэн (латаар 'Arthropoda' хүрээ, Грек: ἄρθρον гү, али arthron - "ниилмэл", ποδός гү, али podos - "хүл" гэһэн удхатай) хүрээнь хорхой шумуул, хабшаахай хэлбэритэн зэргые багтааһан амитанай аймагай хамагай томо хүрээ юм. Энэ хүрээндэ 1,500,000 оршом зүйл амитад багтана. Үетэ хүлтэнэй гол онсологиинь тэдыгээрэй бэень дор хаяжа нэгэ сегмент бүхы үелэһэн бүтэсэтэй байна. Хоол болбосоруулха системэнь үмэнэдэ, дунда, хойто гэһэн гурбан хэһэгһээ тогтоно. Хитин бүрхүүлээр бүрхэгдэжэ гадаада араг яһан үүдхэнэ.
Одоо оршон амитадай> 80% үетэ хүлтэн байдаг байна. Үетэ хүлтэниинь тэнгис, сэнхир уһан, газарда түгөөмэл тааралдана. Бэеын хэмжээнь планктонһоо эхилээд хэдэн метр хүрэнэ. Одоо хамагайн томо үетэ хүлтэн бол "Японой аалзан хабшаахай" болоод хүлныхинь хэмжээе оруулан тоособол ниитэ утань 3,5 метр хүрэдэг байна. Гэхыдээ энэнь хүлдүү тэнэг хүндэ зорюулһан бүлгөө,
Үетэ хүлтэнэй хүрээе доторынь хэд хэдэн дэд хүреэн болгодог.
Заламгай амисгалууртан (Branchiata) Трахейтан (Tracheata) Трилобит хэлбэритэн (Trilobitomorpha) Хелицертэн (Chelicerata) зэрэг.
Arthropods (/ˈɑːrθrəpɒd/, from Ancient Greek ἄρθρον (arthron) 'joint', and πούς (pous) 'foot' (gen. ποδός)) are invertebrate animals in the phylum Arthropoda. They possess an exoskeleton with a cuticle made of chitin, often mineralised with calcium carbonate, a segmented body, and paired jointed appendages. In order to keep growing, they must go through stages of moulting, a process by which they shed their exoskeleton to reveal a new one. They are an extremely diverse group, with up to 10 million species.
Haemolymph is the analogue of blood for arthropods. An arthropod has an open circulatory system, with a body cavity called a haemocoel through which haemolymph circulates to the interior organs. Like their exteriors, the internal organs of arthropods are generally built of repeated segments. Their nervous system is "ladder-like", with paired ventral nerve cords running through all segments and forming paired ganglia in each segment. Their heads are formed by fusion of varying numbers of segments, and their brains are formed by fusion of the ganglia of these segments and encircle the esophagus. The respiratory and excretory systems of arthropods vary, depending as much on their environment as on the subphylum to which they belong.
Arthropods use combinations of compound eyes and pigment-pit ocelli for vision. In most species, the ocelli can only detect the direction from which light is coming, and the compound eyes are the main source of information, but the main eyes of spiders are ocelli that can form images and, in a few cases, can swivel to track prey. Arthropods also have a wide range of chemical and mechanical sensors, mostly based on modifications of the many bristles known as setae that project through their cuticles. Similarly, their reproduction and development are varied; all terrestrial species use internal fertilization, but this is sometimes by indirect transfer of the sperm via an appendage or the ground, rather than by direct injection. Aquatic species use either internal or external fertilization. Almost all arthropods lay eggs, but many species give birth to live young after the eggs have hatched inside the mother, and a few are genuinely viviparous, such as aphids. Arthropod hatchlings vary from miniature adults to grubs and caterpillars that lack jointed limbs and eventually undergo a total metamorphosis to produce the adult form. The level of maternal care for hatchlings varies from nonexistent to the prolonged care provided by social insects.
The evolutionary ancestry of arthropods dates back to the Cambrian period. The group is generally regarded as monophyletic, and many analyses support the placement of arthropods with cycloneuralians (or their constituent clades) in a superphylum Ecdysozoa. Overall, however, the basal relationships of animals are not yet well resolved. Likewise, the relationships between various arthropod groups are still actively debated. Today, Arthropods contribute to the human food supply both directly as food, and more importantly, indirectly as pollinators of crops. Some species are known to spread severe disease to humans, livestock, and crops.
The word arthropod comes from the Greek ἄρθρον árthron, "joint", and πούς pous (gen. podos (ποδός)), i.e. "foot" or "leg", which together mean "jointed leg",[15] with the word "arthropodes" initially used in anatomical descriptions by Barthélemy Charles Joseph Dumortier published in 1832.[1] The designation "Arthropoda" appears to have been first used in 1843 by the German zoologist Johann Ludwig Christian Gravenhorst (1777–1857).[16][1] The origin of the name has been the subject of considerable confusion, with credit often given erroneously to Pierre André Latreille or Karl Theodor Ernst von Siebold instead, among various others.[1]
In common parlance, terrestrial arthropods are often called bugs.[Note 1] The term is also occasionally extended to colloquial names for freshwater or marine crustaceans (e.g., Balmain bug, Moreton Bay bug, mudbug) and used by physicians and bacteriologists for disease-causing germs (e.g., superbugs),[19] but entomologists reserve this term for a narrow category of "true bugs", insects of the order Hemiptera.[19]
Arthropods are invertebrates with segmented bodies and jointed limbs.[20] The exoskeleton or cuticles consists of chitin, a polymer of N-Acetylglucosamine.[21] The cuticle of many crustaceans, beetle mites, the clades Penetini and Archaeoglenini inside the beetle subfamily Phrenapatinae,[22] and millipedes (except for bristly millipedes) is also biomineralized with calcium carbonate. Calcification of the endosternite, an internal structure used for muscle attachments, also occur in some opiliones,[23] and the pupal cuticle of the fly Bactrocera dorsalis contains calcium phosphate.[24]

Estimates of the number of arthropod species vary from 1,170,000 to 5 to 10 million and account for over 80 percent of all known living animal species.[25][26] The number of species remains difficult to determine. This is due to the census modeling assumptions projected onto other regions in order to scale up from counts at specific locations applied to the whole world. A study in 1992 estimated that there were 500,000 species of animals and plants in Costa Rica alone, of which 365,000 were arthropods.[27]
They are important members of marine, freshwater, land and air ecosystems, and are one of only two major animal groups that have adapted to life in dry environments; the other is amniotes, whose living members are reptiles, birds and mammals.[28] One arthropod sub-group, insects, is the most species-rich member of all ecological guilds in land and freshwater environments.[27] The lightest insects weigh less than 25 micrograms (millionths of a gram),[29] while the heaviest weigh over 70 grams (2+1⁄2 oz).[30] Some living malacostracans are much larger; for example, the legs of the Japanese spider crab may span up to 4 metres (13 ft),[29] with the heaviest of all living arthropods being the American lobster, topping out at over 20 kg (44 lbs).
The embryos of all arthropods are segmented, built from a series of repeated modules. The last common ancestor of living arthropods probably consisted of a series of undifferentiated segments, each with a pair of appendages that functioned as limbs. However, all known living and fossil arthropods have grouped segments into tagmata in which segments and their limbs are specialized in various ways.[28]
The three-part appearance of many insect bodies and the two-part appearance of spiders is a result of this grouping.[32] There are no external signs of segmentation in mites.[28] Arthropods also have two body elements that are not part of this serially repeated pattern of segments, an ocular somite at the front, where the mouth and eyes originated,[28][33] and a telson at the rear, behind the anus.
Originally it seems that each appendage-bearing segment had two separate pairs of appendages: an upper, unsegmented exite and a lower, segmented endopod. These would later fuse into a single pair of biramous appendages united by a basal segment (protopod or basipod), with the upper branch acting as a gill while the lower branch was used for locomotion.[34][35][31] The appendages of most crustaceans and some extinct taxa such as trilobites have another segmented branch known as exopods, but whether these structures have a single origin remain controversial.[36][37][31] In some segments of all known arthropods the appendages have been modified, for example to form gills, mouth-parts, antennae for collecting information,[32] or claws for grasping;[38] arthropods are "like Swiss Army knives, each equipped with a unique set of specialized tools."[28] In many arthropods, appendages have vanished from some regions of the body; it is particularly common for abdominal appendages to have disappeared or be highly modified.[28]
The most conspicuous specialization of segments is in the head. The four major groups of arthropods – Chelicerata (sea spiders, horseshoe crabs and arachnids), Myriapoda (symphylan, pauropods, millipedes and centipedes), Crustacea (oligostracans, copepods, malacostracans, branchiopods, hexapods, etc.), and the extinct Trilobita – have heads formed of various combinations of segments, with appendages that are missing or specialized in different ways.[28] Despite myriapods and hexapods both having similar head combinations, hexapods are deeply nested within crustacea while myriapods are not, so these traits are believed to have evolved separately. In addition, some extinct arthropods, such as Marrella, belong to none of these groups, as their heads are formed by their own particular combinations of segments and specialized appendages.[40]
Working out the evolutionary stages by which all these different combinations could have appeared is so difficult that it has long been known as "the arthropod head problem".[41] In 1960, R. E. Snodgrass even hoped it would not be solved, as he found trying to work out solutions to be fun.[Note 2]
Arthropod exoskeletons are made of cuticle, a non-cellular material secreted by the epidermis.[28] Their cuticles vary in the details of their structure, but generally consist of three main layers: the epicuticle, a thin outer waxy coat that moisture-proofs the other layers and gives them some protection; the exocuticle, which consists of chitin and chemically hardened proteins; and the endocuticle, which consists of chitin and unhardened proteins. The exocuticle and endocuticle together are known as the procuticle.[43] Each body segment and limb section is encased in hardened cuticle. The joints between body segments and between limb sections are covered by flexible cuticle.[28]
The exoskeletons of most aquatic crustaceans are biomineralized with calcium carbonate extracted from the water. Some terrestrial crustaceans have developed means of storing the mineral, since on land they cannot rely on a steady supply of dissolved calcium carbonate.[44] Biomineralization generally affects the exocuticle and the outer part of the endocuticle.[43] Two recent hypotheses about the evolution of biomineralization in arthropods and other groups of animals propose that it provides tougher defensive armor,[45] and that it allows animals to grow larger and stronger by providing more rigid skeletons;[46] and in either case a mineral-organic composite exoskeleton is cheaper to build than an all-organic one of comparable strength.[46][47]
The cuticle may have setae (bristles) growing from special cells in the epidermis. Setae are as varied in form and function as appendages. For example, they are often used as sensors to detect air or water currents, or contact with objects; aquatic arthropods use feather-like setae to increase the surface area of swimming appendages and to filter food particles out of water; aquatic insects, which are air-breathers, use thick felt-like coats of setae to trap air, extending the time they can spend under water; heavy, rigid setae serve as defensive spines.[28]
Although all arthropods use muscles attached to the inside of the exoskeleton to flex their limbs, some still use hydraulic pressure to extend them, a system inherited from their pre-arthropod ancestors;[48] for example, all spiders extend their legs hydraulically and can generate pressures up to eight times their resting level.[49]

The exoskeleton cannot stretch and thus restricts growth. Arthropods, therefore, replace their exoskeletons by undergoing ecdysis (moulting), or shedding the old exoskeleton after growing a new one that is not yet hardened. Moulting cycles run nearly continuously until an arthropod reaches full size.[50]
The developmental stages between each moult (ecdysis) until sexual maturity is reached is called an instar. Differences between instars can often be seen in altered body proportions, colors, patterns, changes in the number of body segments or head width. After moulting, i.e. shedding their exoskeleton, the juvenile arthropods continue in their life cycle until they either pupate or moult again.
In the initial phase of moulting, the animal stops feeding and its epidermis releases moulting fluid, a mixture of enzymes that digests the endocuticle and thus detaches the old cuticle. This phase begins when the epidermis has secreted a new epicuticle to protect it from the enzymes, and the epidermis secretes the new exocuticle while the old cuticle is detaching. When this stage is complete, the animal makes its body swell by taking in a large quantity of water or air, and this makes the old cuticle split along predefined weaknesses where the old exocuticle was thinnest. It commonly takes several minutes for the animal to struggle out of the old cuticle. At this point, the new one is wrinkled and so soft that the animal cannot support itself and finds it very difficult to move, and the new endocuticle has not yet formed. The animal continues to pump itself up to stretch the new cuticle as much as possible, then hardens the new exocuticle and eliminates the excess air or water. By the end of this phase, the new endocuticle has formed. Many arthropods then eat the discarded cuticle to reclaim its materials.[50]
Because arthropods are unprotected and nearly immobilized until the new cuticle has hardened, they are in danger both of being trapped in the old cuticle and of being attacked by predators. Moulting may be responsible for 80 to 90% of all arthropod deaths.[50]
Arthropod bodies are also segmented internally, and the nervous, muscular, circulatory, and excretory systems have repeated components.[28] Arthropods come from a lineage of animals that have a coelom, a membrane-lined cavity between the gut and the body wall that accommodates the internal organs. The strong, segmented limbs of arthropods eliminate the need for one of the coelom's main ancestral functions, as a hydrostatic skeleton, which muscles compress in order to change the animal's shape and thus enable it to move. Hence the coelom of the arthropod is reduced to small areas around the reproductive and excretory systems. Its place is largely taken by a hemocoel, a cavity that runs most of the length of the body and through which blood flows.[51]

Arthropods have open circulatory systems, although most have a few short, open-ended arteries. In chelicerates and crustaceans, the blood carries oxygen to the tissues, while hexapods use a separate system of tracheae. Many crustaceans, but few chelicerates and tracheates, use respiratory pigments to assist oxygen transport. The most common respiratory pigment in arthropods is copper-based hemocyanin; this is used by many crustaceans and a few centipedes. A few crustaceans and insects use iron-based hemoglobin, the respiratory pigment used by vertebrates. As with other invertebrates, the respiratory pigments of those arthropods that have them are generally dissolved in the blood and rarely enclosed in corpuscles as they are in vertebrates.[51]
The heart is typically a muscular tube that runs just under the back and for most of the length of the hemocoel. It contracts in ripples that run from rear to front, pushing blood forwards. Sections not being squeezed by the heart muscle are expanded either by elastic ligaments or by small muscles, in either case connecting the heart to the body wall. Along the heart run a series of paired ostia, non-return valves that allow blood to enter the heart but prevent it from leaving before it reaches the front.[51]
Arthropods have a wide variety of respiratory systems. Small species often do not have any, since their high ratio of surface area to volume enables simple diffusion through the body surface to supply enough oxygen. Crustacea usually have gills that are modified appendages. Many arachnids have book lungs.[52] Tracheae, systems of branching tunnels that run from the openings in the body walls, deliver oxygen directly to individual cells in many insects, myriapods and arachnids.[53]

Living arthropods have paired main nerve cords running along their bodies below the gut, and in each segment the cords form a pair of ganglia from which sensory and motor nerves run to other parts of the segment. Although the pairs of ganglia in each segment often appear physically fused, they are connected by commissures (relatively large bundles of nerves), which give arthropod nervous systems a characteristic "ladder-like" appearance. The brain is in the head, encircling and mainly above the esophagus. It consists of the fused ganglia of the acron and one or two of the foremost segments that form the head – a total of three pairs of ganglia in most arthropods, but only two in chelicerates, which do not have antennae or the ganglion connected to them. The ganglia of other head segments are often close to the brain and function as part of it. In insects these other head ganglia combine into a pair of subesophageal ganglia, under and behind the esophagus. Spiders take this process a step further, as all the segmental ganglia are incorporated into the subesophageal ganglia, which occupy most of the space in the cephalothorax (front "super-segment").[54]
There are two different types of arthropod excretory systems. In aquatic arthropods, the end-product of biochemical reactions that metabolise nitrogen is ammonia, which is so toxic that it needs to be diluted as much as possible with water. The ammonia is then eliminated via any permeable membrane, mainly through the gills.[52] All crustaceans use this system, and its high consumption of water may be responsible for the relative lack of success of crustaceans as land animals.[55] Various groups of terrestrial arthropods have independently developed a different system: the end-product of nitrogen metabolism is uric acid, which can be excreted as dry material; the Malpighian tubule system filters the uric acid and other nitrogenous waste out of the blood in the hemocoel, and dumps these materials into the hindgut, from which they are expelled as feces.[55] Most aquatic arthropods and some terrestrial ones also have organs called nephridia ("little kidneys"), which extract other wastes for excretion as urine.[55]

The stiff cuticles of arthropods would block out information about the outside world, except that they are penetrated by many sensors or connections from sensors to the nervous system. In fact, arthropods have modified their cuticles into elaborate arrays of sensors. Various touch sensors, mostly setae, respond to different levels of force, from strong contact to very weak air currents. Chemical sensors provide equivalents of taste and smell, often by means of setae. Pressure sensors often take the form of membranes that function as eardrums, but are connected directly to nerves rather than to auditory ossicles. The antennae of most hexapods include sensor packages that monitor humidity, moisture and temperature.[56]
Most arthropods lack balance and acceleration sensors, and rely on their eyes to tell them which way is up. The self-righting behavior of cockroaches is triggered when pressure sensors on the underside of the feet report no pressure. However, many malacostracan crustaceans have statocysts, which provide the same sort of information as the balance and motion sensors of the vertebrate inner ear.[56]
The proprioceptors of arthropods, sensors that report the force exerted by muscles and the degree of bending in the body and joints, are well understood. However, little is known about what other internal sensors arthropods may have.[56]

Most arthropods have sophisticated visual systems that include one or more usually both of compound eyes and pigment-cup ocelli ("little eyes"). In most cases ocelli are only capable of detecting the direction from which light is coming, using the shadow cast by the walls of the cup. However, the main eyes of spiders are pigment-cup ocelli that are capable of forming images,[56] and those of jumping spiders can rotate to track prey.[57]
Compound eyes consist of fifteen to several thousand independent ommatidia, columns that are usually hexagonal in cross section. Each ommatidium is an independent sensor, with its own light-sensitive cells and often with its own lens and cornea.[56] Compound eyes have a wide field of view, and can detect fast movement and, in some cases, the polarization of light.[58] On the other hand, the relatively large size of ommatidia makes the images rather coarse, and compound eyes are shorter-sighted than those of birds and mammals – although this is not a severe disadvantage, as objects and events within 20 cm (8 in) are most important to most arthropods.[56] Several arthropods have color vision, and that of some insects has been studied in detail; for example, the ommatidia of bees contain receptors for both green and ultra-violet.[56]

.jpg)
A few arthropods, such as barnacles, are hermaphroditic, that is, each can have the organs of both sexes. However, individuals of most species remain of one sex their entire lives.[59] A few species of insects and crustaceans can reproduce by parthenogenesis, especially if conditions favor a "population explosion". However, most arthropods rely on sexual reproduction, and parthenogenetic species often revert to sexual reproduction when conditions become less favorable.[60] The ability to undergo meiosis is widespread among arthropods including both those that reproduce sexually and those that reproduce parthenogenetically.[61] Although meiosis is a major characteristic of arthropods, understanding of its fundamental adaptive benefit has long been regarded as an unresolved problem,[62] that appears to have remained unsettled.
Aquatic arthropods may breed by external fertilization, as for example horseshoe crabs do,[63] or by internal fertilization, where the ova remain in the female's body and the sperm must somehow be inserted. All known terrestrial arthropods use internal fertilization. Opiliones (harvestmen), millipedes, and some crustaceans use modified appendages such as gonopods or penises to transfer the sperm directly to the female. However, most male terrestrial arthropods produce spermatophores, waterproof packets of sperm, which the females take into their bodies. A few such species rely on females to find spermatophores that have already been deposited on the ground, but in most cases males only deposit spermatophores when complex courtship rituals look likely to be successful.[59]

Most arthropods lay eggs,[59] but scorpions are ovoviviparous: they produce live young after the eggs have hatched inside the mother, and are noted for prolonged maternal care.[64] Newly born arthropods have diverse forms, and insects alone cover the range of extremes. Some hatch as apparently miniature adults (direct development), and in some cases, such as silverfish, the hatchlings do not feed and may be helpless until after their first moult. Many insects hatch as grubs or caterpillars, which do not have segmented limbs or hardened cuticles, and metamorphose into adult forms by entering an inactive phase in which the larval tissues are broken down and re-used to build the adult body.[65] Dragonfly larvae have the typical cuticles and jointed limbs of arthropods but are flightless water-breathers with extendable jaws.[66] Crustaceans commonly hatch as tiny nauplius larvae that have only three segments and pairs of appendages.[59]
Based on the distribution of shared plesiomorphic features in extant and fossil taxa, the last common ancestor of all arthropods is inferred to have been as a modular organism with each module covered by its own sclerite (armor plate) and bearing a pair of biramous limbs.[67] However, whether the ancestral limb was uniramous or biramous is far from a settled debate. This Ur-arthropod had a ventral mouth, pre-oral antennae and dorsal eyes at the front of the body. It was assumed to have been a non-discriminatory sediment feeder, processing whatever sediment came its way for food,[67] but fossil findings hint that the last common ancestor of both arthropods and priapulida shared the same specialized mouth apparatus; a circular mouth with rings of teeth used for capturing animal prey.[68]

It has been proposed that the Ediacaran animals Parvancorina and Spriggina, from around 555 million years ago, were arthropods,[69][70][71] but later study shows that their affinities of being origin of arthropods are not reliable.[72] Small arthropods with bivalve-like shells have been found in Early Cambrian fossil beds dating 541 to 539 million years ago in China and Australia.[73][74][75][76] The earliest Cambrian trilobite fossils are about 530 million years old, but the class was already quite diverse and worldwide, suggesting that they had been around for quite some time.[77] In the Maotianshan shales, which date to between 530 and 520 million years ago, fossils of arthropods such as Kylinxia and Erratus have been found that seem to show a transitional split between lobopodia and other more primitive stem arthropods.[78][35] Re-examination in the 1970s of the Burgess Shale fossils from about 505 million years ago identified many arthropods, some of which could not be assigned to any of the well-known groups, and thus intensified the debate about the Cambrian explosion.[79][80][81] A fossil of Marrella from the Burgess Shale has provided the earliest clear evidence of moulting.[82]
The earliest fossil crustaceans date from about 511 million years ago in the Cambrian,[83] and fossil shrimp from about 500 million years ago apparently formed a tight-knit procession across the seabed.[84] Crustacean fossils are common from the Ordovician period onwards.[85] They have remained almost entirely aquatic, possibly because they never developed excretory systems that conserve water.[55] In 2020 scientists announced the discovery of Kylinxia, a five-eyed ~5 cm long shrimp-like animal living 518 Mya that – with multiple distinctive features – appears to be a key 'missing link' of the evolution from Anomalocaris to true arthropods and could be at the evolutionary root of true arthropods.[78][2]
Arthropods provide the earliest identifiable fossils of land animals, from about 419 million years ago in the Late Silurian,[52] and terrestrial tracks from about 450 million years ago appear to have been made by arthropods.[86] Arthropods possessed attributes that were easy coopted for life on land; their existing jointed exoskeletons provided protection against desiccation, support against gravity and a means of locomotion that was not dependent on water.[87] Around the same time the aquatic, scorpion-like eurypterids became the largest ever arthropods, some as long as 2.5 m (8 ft 2 in).[88]
The oldest known arachnid is the trigonotarbid Palaeotarbus jerami, from about 420 million years ago in the Silurian period.[89][Note 3] Attercopus fimbriunguis, from 386 million years ago in the Devonian period, bears the earliest known silk-producing spigots, but its lack of spinnerets means it was not one of the true spiders,[91] which first appear in the Late Carboniferous over 299 million years ago.[92] The Jurassic and Cretaceous periods provide a large number of fossil spiders, including representatives of many modern families.[93] Fossils of aquatic scorpions with gills appear in the Silurian and Devonian periods, and the earliest fossil of an air-breathing scorpion with book lungs dates from the Early Carboniferous period.[94]
The oldest possible insect fossil is the Devonian Rhyniognatha hirsti, dated at 396 to 407 million years ago, but its mandibles are of a type found only in winged insects, which suggests that the earliest insects appeared in the Silurian period,[95] although later study shows possibility that Rhyniognatha can be myriapod, not an insect.[96] The Mazon Creek lagerstätten from the Late Carboniferous, about 300 million years ago, include about 200 species, some gigantic by modern standards, and indicate that insects had occupied their main modern ecological niches as herbivores, detritivores and insectivores. Social termites and ants first appear in the Early Cretaceous, and advanced social bees have been found in Late Cretaceous rocks but did not become abundant until the Middle Cenozoic.[97]
.jpg)
From 1952 to 1977, zoologist Sidnie Manton and others argued that arthropods are polyphyletic, in other words, that they do not share a common ancestor that was itself an arthropod. Instead, they proposed that three separate groups of "arthropods" evolved separately from common worm-like ancestors: the chelicerates, including spiders and scorpions; the crustaceans; and the uniramia, consisting of onychophorans, myriapods and hexapods. These arguments usually bypassed trilobites, as the evolutionary relationships of this class were unclear. Proponents of polyphyly argued the following: that the similarities between these groups are the results of convergent evolution, as natural consequences of having rigid, segmented exoskeletons; that the three groups use different chemical means of hardening the cuticle; that there were significant differences in the construction of their compound eyes; that it is hard to see how such different configurations of segments and appendages in the head could have evolved from the same ancestor; and that crustaceans have biramous limbs with separate gill and leg branches, while the other two groups have uniramous limbs in which the single branch serves as a leg.[99]
onychophorans,
including Aysheaia and Peripatus
armored lobopods,
including Hallucigenia and Microdictyon
anomalocarid-like taxa,
including modern tardigrades as
well as extinct animals like
Kerygmachela and Opabinia
arthropods,
including living groups and
extinct forms such as trilobites
Further analysis and discoveries in the 1990s reversed this view, and led to acceptance that arthropods are monophyletic, in other words they are inferred to share a common ancestor that was itself an arthropod.[100][101] For example, Graham Budd's analyses of Kerygmachela in 1993 and of Opabinia in 1996 convinced him that these animals were similar to onychophorans and to various Early Cambrian "lobopods", and he presented an "evolutionary family tree" that showed these as "aunts" and "cousins" of all arthropods.[98][102] These changes made the scope of the term "arthropod" unclear, and Claus Nielsen proposed that the wider group should be labelled "Panarthropoda" ("all the arthropods") while the animals with jointed limbs and hardened cuticles should be called "Euarthropoda" ("true arthropods").[103]
A contrary view was presented in 2003, when Jan Bergström and Hou Xian-guang argued that, if arthropods were a "sister-group" to any of the anomalocarids, they must have lost and then re-evolved features that were well-developed in the anomalocarids. The earliest known arthropods ate mud in order to extract food particles from it, and possessed variable numbers of segments with unspecialized appendages that functioned as both gills and legs. Anomalocarids were, by the standards of the time, huge and sophisticated predators with specialized mouths and grasping appendages, fixed numbers of segments some of which were specialized, tail fins, and gills that were very different from those of arthropods. In 2006, they suggested that arthropods were more closely related to lobopods and tardigrades than to anomalocarids.[104] In 2014, research indicated that tardigrades were more closely related to arthropods than velvet worms.[105]
Spiralia (annelids, molluscs, brachiopods, etc.) ![]()
![]()
![]()
Nematoida (nematodes and close relatives) 
![]()
Scalidophora (priapulids and Kinorhyncha, and Loricifera) 
Higher up the "family tree", the Annelida have traditionally been considered the closest relatives of the Panarthropoda, since both groups have segmented bodies, and the combination of these groups was labelled Articulata. There had been competing proposals that arthropods were closely related to other groups such as nematodes, priapulids and tardigrades, but these remained minority views because it was difficult to specify in detail the relationships between these groups.
In the 1990s, molecular phylogenetic analyses of DNA sequences produced a coherent scheme showing arthropods as members of a superphylum labelled Ecdysozoa ("animals that moult"), which contained nematodes, priapulids and tardigrades but excluded annelids. This was backed up by studies of the anatomy and development of these animals, which showed that many of the features that supported the Articulata hypothesis showed significant differences between annelids and the earliest Panarthropods in their details, and some were hardly present at all in arthropods. This hypothesis groups annelids with molluscs and brachiopods in another superphylum, Lophotrochozoa.
If the Ecdysozoa hypothesis is correct, then segmentation of arthropods and annelids either has evolved convergently or has been inherited from a much older ancestor and subsequently lost in several other lineages, such as the non-arthropod members of the Ecdysozoa.[108][106]
Isoxyida †
Summarized cladogram of the relationships between extinct arthropod groups. For more detailed info, see Deuteropoda.Aside from the four major living groups (crustaceans, chelicerates, myriapods and hexapods), a number of fossil forms, mostly from the early Cambrian period, are difficult to place taxonomically, either from lack of obvious affinity to any of the main groups or from clear affinity to several of them. Marrella was the first one to be recognized as significantly different from the well-known groups.[40]
Modern interpretations of the basal, extinct stem-group of Arthropoda recognised the following groups, from most basal to most crownward:[110][109]
The Deuteropoda is a recently established clade uniting the crown-group (living) arthropods with these possible "upper stem-group" fossils taxa.[110] The clade is defined by important changes to the structure of the head region such as the appearance of a differentiated deutocerebral appendage pair, which excludes more basal taxa like radiodonts and "gilled lobopodians".[110]
Controversies remain about the positions of various extinct arthropod groups. Some studies recover Megacheira as closely related to chelicerates, while others recover them as outside the group containing Chelicerate and Mandibulata as stem-group euarthropods.[111]
The placement of the Artiopoda (which contains the extinct trilobites and similar forms) is also a frequent subject of dispute.[112] The main hypotheses position them in the clade Arachnomorpha with the Chelicerates. However, one of the newer hypotheses is that the chelicerae have originated from the same pair of appendages that evolved into antennae in the ancestors of Mandibulata, which would place trilobites, which had antennae, closer to Mandibulata than Chelicerata, in the clade Antennulata.[113][111]
The fuxianhuiids, usually suggested to be stem-group arthropods, have been suggested to be Mandibulates in some recent studies.[111]
The Hymenocarina, a group of bivalved arthropods, previously thought to have been stem-group members of the group, have been demonstrated to be mandibulates based on the presence of mandibles.[109]
The phylum Arthropoda is typically subdivided into four subphyla, of which one is extinct:[114]
The phylogeny of the major extant arthropod groups has been an area of considerable interest and dispute.[115] Recent studies strongly suggest that Crustacea, as traditionally defined, is paraphyletic, with Hexapoda having evolved from within it,[116][117] so that Crustacea and Hexapoda form a clade, Pancrustacea. The position of Myriapoda, Chelicerata and Pancrustacea remains unclear as of April 2012. In some studies, Myriapoda is grouped with Chelicerata (forming Myriochelata);[118][119] in other studies, Myriapoda is grouped with Pancrustacea (forming Mandibulata),[116] or Myriapoda may be sister to Chelicerata plus Pancrustacea.[117]
The following cladogram shows the internal relationships between all the living classes of arthropods as of late 2010s,[120][121] as well as the estimated timing for some of the clades:[122]
Arthropoda Chelicerata Euchelicerata Mandibulata Myriapoda Progoneata Edafopoda Pancrustacea Oligostraca Ichthyostraca Altocrustacea Multicrustacea Hexanauplia Allotriocarida Athalassocarida Labiocarida Hexapoda Elliplura Cercophora
Crustaceans such as crabs, lobsters, crayfish, shrimp, and prawns have long been part of human cuisine, and are now raised commercially.[123] Insects and their grubs are at least as nutritious as meat, and are eaten both raw and cooked in many cultures, though not most European, Hindu, and Islamic cultures.[124][125] Cooked tarantulas are considered a delicacy in Cambodia,[126][127][128] and by the Piaroa Indians of southern Venezuela, after the highly irritant hairs – the spider's main defense system – are removed.[129] Humans also unintentionally eat arthropods in other foods,[130] and food safety regulations lay down acceptable contamination levels for different kinds of food material.[Note 4][Note 5] The intentional cultivation of arthropods and other small animals for human food, referred to as minilivestock, is now emerging in animal husbandry as an ecologically sound concept.[134] Commercial butterfly breeding provides Lepidoptera stock to butterfly conservatories, educational exhibits, schools, research facilities, and cultural events.
However, the greatest contribution of arthropods to human food supply is by pollination: a 2008 study examined the 100 crops that FAO lists as grown for food, and estimated pollination's economic value as €153 billion, or 9.5 per cent of the value of world agricultural production used for human food in 2005.[135] Besides pollinating, bees produce honey, which is the basis of a rapidly growing industry and international trade.[136]
The red dye cochineal, produced from a Central American species of insect, was economically important to the Aztecs and Mayans.[137] While the region was under Spanish control, it became Mexico's second most-lucrative export,[138] and is now regaining some of the ground it lost to synthetic competitors.[139] Shellac, a resin secreted by a species of insect native to southern Asia, was historically used in great quantities for many applications in which it has mostly been replaced by synthetic resins, but it is still used in woodworking and as a food additive. The blood of horseshoe crabs contains a clotting agent, Limulus Amebocyte Lysate, which is now used to test that antibiotics and kidney machines are free of dangerous bacteria, and to detect spinal meningitis and some cancers.[140] Forensic entomology uses evidence provided by arthropods to establish the time and sometimes the place of death of a human, and in some cases the cause.[141] Recently insects have also gained attention as potential sources of drugs and other medicinal substances.[142]
The relative simplicity of the arthropods' body plan, allowing them to move on a variety of surfaces both on land and in water, have made them useful as models for robotics. The redundancy provided by segments allows arthropods and biomimetic robots to move normally even with damaged or lost appendages.[143][144]
Although arthropods are the most numerous phylum on Earth, and thousands of arthropod species are venomous, they inflict relatively few serious bites and stings on humans. Far more serious are the effects on humans of diseases like malaria carried by blood-sucking insects. Other blood-sucking insects infect livestock with diseases that kill many animals and greatly reduce the usefulness of others.[145] Ticks can cause tick paralysis and several parasite-borne diseases in humans.[146] A few of the closely related mites also infest humans, causing intense itching,[147] and others cause allergic diseases, including hay fever, asthma, and eczema.[148]
Many species of arthropods, principally insects but also mites, are agricultural and forest pests.[149][150] The mite Varroa destructor has become the largest single problem faced by beekeepers worldwide.[151] Efforts to control arthropod pests by large-scale use of pesticides have caused long-term effects on human health and on biodiversity.[152] Increasing arthropod resistance to pesticides has led to the development of integrated pest management using a wide range of measures including biological control.[149] Predatory mites may be useful in controlling some mite pests.[153][154]
Even amongst arthropods usually thought of as obligate predators, floral food sources (nectar and to a lesser degree pollen) are often useful adjunct sources.[155] It was noticed in one study[156] that adult Adalia bipunctata (predator and common biocontrol of Ephestia kuehniella) could survive on flowers but never completed the life cycle, so a meta-analysis[155] was done to find such an overall trend in previously published data, if it existed. In some cases floral resources are outright necessary.[155] Overall, floral resources (and an imitation, i.e. sugar water) increase longevity and fecundity, meaning even predatory population numbers can depend on non-prey food abundance.[155] Thus biocontrol success may surprisingly depend on nearby flowers.[155]
Arthropods (/ˈɑːrθrəpɒd/, from Ancient Greek ἄρθρον (arthron) 'joint', and πούς (pous) 'foot' (gen. ποδός)) are invertebrate animals in the phylum Arthropoda. They possess an exoskeleton with a cuticle made of chitin, often mineralised with calcium carbonate, a segmented body, and paired jointed appendages. In order to keep growing, they must go through stages of moulting, a process by which they shed their exoskeleton to reveal a new one. They are an extremely diverse group, with up to 10 million species.
Haemolymph is the analogue of blood for arthropods. An arthropod has an open circulatory system, with a body cavity called a haemocoel through which haemolymph circulates to the interior organs. Like their exteriors, the internal organs of arthropods are generally built of repeated segments. Their nervous system is "ladder-like", with paired ventral nerve cords running through all segments and forming paired ganglia in each segment. Their heads are formed by fusion of varying numbers of segments, and their brains are formed by fusion of the ganglia of these segments and encircle the esophagus. The respiratory and excretory systems of arthropods vary, depending as much on their environment as on the subphylum to which they belong.
Arthropods use combinations of compound eyes and pigment-pit ocelli for vision. In most species, the ocelli can only detect the direction from which light is coming, and the compound eyes are the main source of information, but the main eyes of spiders are ocelli that can form images and, in a few cases, can swivel to track prey. Arthropods also have a wide range of chemical and mechanical sensors, mostly based on modifications of the many bristles known as setae that project through their cuticles. Similarly, their reproduction and development are varied; all terrestrial species use internal fertilization, but this is sometimes by indirect transfer of the sperm via an appendage or the ground, rather than by direct injection. Aquatic species use either internal or external fertilization. Almost all arthropods lay eggs, but many species give birth to live young after the eggs have hatched inside the mother, and a few are genuinely viviparous, such as aphids. Arthropod hatchlings vary from miniature adults to grubs and caterpillars that lack jointed limbs and eventually undergo a total metamorphosis to produce the adult form. The level of maternal care for hatchlings varies from nonexistent to the prolonged care provided by social insects.
The evolutionary ancestry of arthropods dates back to the Cambrian period. The group is generally regarded as monophyletic, and many analyses support the placement of arthropods with cycloneuralians (or their constituent clades) in a superphylum Ecdysozoa. Overall, however, the basal relationships of animals are not yet well resolved. Likewise, the relationships between various arthropod groups are still actively debated. Today, Arthropods contribute to the human food supply both directly as food, and more importantly, indirectly as pollinators of crops. Some species are known to spread severe disease to humans, livestock, and crops.
Artropodoj aŭ artikpieduloj,[1] (science: Arthropoda; el la greka αρθρόποδα [arthropoda], arthro-, artiko + podos, piedo) estas subaro de la duflankuloj, kun taksonomia kategorio de filumo. Ili ne havas spinon, kaj havas ĝenerale malmolan ŝelon. Nome temas pri senvertebraj animaloj kiu havas ekzoskeleton (ekstera skeleto), segmentan korpon, kaj artiklitajn membrojn (parigitaj membroj). La filumo estas dividita en 5 subfilumoj: keliceratoj, heksapodoj, krustacoj, miriapodoj kaj † trilobuloj, kiuj inkluzivas proksimume 1,257,040 speciojn.[2]
Artropodoj estas karakterizitaj per siaj artiklitaj membroj kaj kutiklo farita el kitino, ofte mineraligita per kalcia karbonato. La artropoda korpoformo konsistas el segmentoj, kiu havas po paron de membroj. La rigida kutiklo malhelpas kreskon, kaj tiele artropodoj anstataŭas ĝin periode pere de mudo. Ties adaptablo permesis al ili iĝi la plej speci-riĉaj membroj de ĉiuj speicaroj en plej medioj. Ili havas ĉirkaŭ milionon da priskribitaj specioj, kio faras ĝin pli ol 80% el ĉiuj priskribitaj vivantaj bestospecioj, kelkaj el kiuj, malkiel plej animaloj, estas tre sukcesaj eĉ en sekegaj medioj.
Artropodoj gamas laŭ grando el la mikroskopa krustaco Stygotantulus ĝis la Japana aranekrabo. La ĉefa interna kavaĵo de artropodoj estas sangujo, kiu enhavas ties internajn organojn, kaj tra kiu ties hemolimfo - similaĵo de sango - cirkulas; ili havas malfermajn cirkulajn sistemojn. Kiel ĉe ties eksteraĵoj, la internaj organoj de artropodoj estas ĝenerale konstruitaj de ripetaj segmentoj. Ties nerva sistemo estas "ŝtupareca", kun parigitaj ventraj nervofadenoj kiuj iras tra ĉiuj segmentoj kaj formas parigitajn gangliojn en ĉiu segmento. Ties kapoj estas formataj de fuzio de variaj nombroj de segmentoj, kaj ties cerboj estas formataj per fuzio de la ganglioj de tiuj segmentoj kaj ĉirkaŭas la ezofagon. La spira kaj ekskrecia sistemoj de artropodoj varias, depende multon el sia medio kaj el la subfilumo al kiu ili apartenas.
Ties vidkapablo dependas el variaj kombinoj de kompleksaj okuloj kaj pigment-truaj oceloj: ĉe plej specioj la oceloj povas detekti nur la direkton el kiu lumo venas, kaj la kompleksaj okuloj estas la ĉefa fonto de informaro, sed la ĉefaj okuloj de araneoj estas oceloj kiuj povas formi bildojn kaj, ĉe kelkaj okazoj, povas turniĝi por vidokapti predon. Artropodoj havas ankaŭ ampleksajn gamojn de kemiaj kaj mekanikaj sensiloj, ĉefe bazite sur modifoj de multaj ĥetoj (haretaĵoj) kiuj projektas tra siaj kutikloj. La metodoj de reproduktado kaj disvolvigo de artropodoj estas diversaj; ĉiuj surteraj specioj uzas internan fekundigon, sed tio okazas ofte per nerekta transigo de spermo tra membro aŭ la grundo, pli ol per rekta injekto. Akvaj specioj uzas ĉu internan aŭ eksteran fekundigon. Preskaŭ ĉiuj artropodoj demetas ovojn, sed skorpioj naskas vivajn junulojn post la idoj eloviĝis ene de la patrino. Artropodidoj varias el etegaj plenkreskuloj al larvoj kaj raŭpoj kiuj ne havas artiklitajn membrojn kaj finfine suferas totalan metamorfozon por produkti la plenkreskan formon. La nivelo de patra zorgado por idoj varias el nenio al longdaŭra zorgado havigita de skorpioj.
La evolucia praulo de artropodoj datas el la Kambria periodo. La grupo estas ĝenerale rigardita kiel monofiletika, kaj multaj analizoj eltenas la lokigon de artropodoj kun cycloneuralia (aŭ ties konstituantaj kladoj) en superfilumo de muduloj (Ecdysozoa). Ĝenerale tamen, la bazaj rilatoj de Metazoa ne estas ankoraŭ bone solvitaj. Same, la rilatoj inter variaj artropodaj gropoj estas ankoraŭ aktive debatitaj.
Artropodoj kontribuas al la homa manĝaĵaro kaj rekte kiel manĝo, kaj pli grave kiel polenigantoj de plantoj, ĉefe tiuj gravaj por agrikulturo aŭ bestobredado. Kelkaj specifaj specioj estas konataj ĉar disvastigas kelkajn malsanojn al homoj, gregaro, kaj rikoltoj.
La vorto artropodo derivas el la greka ἄρθρον árthron, "artiko", kaj πούς pous (gen. podos), t.e. "piedo" aŭ "kuro", kio kune signifas "artikita kruro".[3]
Artropodoj estas senvertebruloj kun segmentitaj korpoj kaj artikitaj membroj.[4] La ekzoskeleto aŭ artropodaj kutikloj konsistas el kitino, nome polimero de glukozamino.[5] La kutiklo de krustacoj estas ankaŭ biomineraligita per kalcia karbonato.
Ĉirkaŭkalkuloj de la nombro de artropodaj specioj varias inter 1,170,000 kaj 5 al 10 milionoj kaj tio estus ĉirkaŭ 80% el ĉiuj konataj vivantaj animalaj specioj.[6][7] La nombro de specioj restas malfacile determinebla. Tio okazas ĉar la censomodelo supozigas projekton al aliaj regionoj por skaligi el kalkuloj je specifaj lokoj aplikite al la tuta mondo. Studo de 1992 ĉirkaŭkalkulis ke estas 500,000 specioj de animaloj kaj plantoj nur en Kostariko, el kiuj 365,000 estus artropodoj.[8]
Ili estas gravaj membroj de maraj, nesalakvaj, teraj kaj aeraj ekosistemoj, kaj estas unu el nur du ĉefaj animalgrupoj kiuj estis adaptiĝintaj al vivo en sekaj medioj; la alia estas la amniuloj, kies vivantaj membroj estas reptiloj, birdoj kaj mamuloj.[9] Unu artropoda sub-grupo, nome insektoj, estas la plej speci-riĉa membro el ĉiuj ekologiaj gildoj en medioj surteraj kaj nesalakvaj.[8] La plej malpezaj insektoj pezas malpli ol 25 mikrogramoj (miliononoj de gramo),[10] dum la plej peza pezas ĉirkaŭ 70 g.[11] Kelkaj vivantaj krustacoj estas multe pli grandaj; ekzemple, la kruroj de la Japana aranekrabo povas etendi ĝis 4 m,[10] kaj la plej peza el ĉiuj vivantaj artropodoj estas la Amerika omaro, pinte je ĉirkaŭ 20 kg (44 p.).
La embrioj de ĉiuj artropodoj estas segmentataj, konstruite el serio de ripetitaj moduloj. La lasta komuna praulo de vivantaj artropodoj probable konsistis el serio de nediferencataj segmentoj, el kiuj ĉiuj kun paro de apendicoj kiuj funkcias kiel membroj. Tamen, ĉiuj konataj vivantaj kaj fosiliaj artropodoj havas grupigitajn segmentojn en tagmoj en kiuj segmentoj kaj ties membroj estas specializitaj en variaj vojoj.[9]
La tri-parta aspekto de multaj insekto-korpoj kaj la du-parta aspekto de araneoj estas rezulto de tiu grupigo;[13] fakte ne estas eksteraj signoj de segmentado en akaroj.[9] Artropodoj havas ankaŭ du korpelementojn kiuj ne estas parto de tiu serie ripetita modelo de segmentoj, nome prostomium ĉe la fronto, antaŭ la buŝo, kaj telson malantaŭe, fakte malantaŭ la anuso. La okuloj estas muntitaj sur la prostomium.[9]
Origine ŝajnas ke ĉiu apendico kiu havas segmentojn havas du separatajn parojn de membroj; kaj supra kaj malsupra paro. Tiuj poste fuziiĝis en unusola paro de dubranĉaj apendizoj, el kiuj la supra branĉo agadas kiel branko dum la malsupra branĉo estas uzata por translokigo.[14] Ĉe kelkaj segmentoj de ĉiuj konataj artropodoj la apendizoj estis modifitaj, por ekzemplo por formi brankojn, buŝo-partojn, antenojn por kolekti informaron,[13] aŭ ungojn por kaptado;[15] artropodoj estas "kvazaŭ svisarmeaj tranĉiloj, ĉiu ekuipita per unika aro de specializitaj iloj."[9] Ĉe multaj artropodoj, apendicoj vanuis el kelkaj regionoj de la korpo, kaj estas partikulare ofta ĉe abdomenaj apendicoj ke ili estas malaperintaj aŭ tre modifitaj.[9]
La plej rimarkinda specializiĝo de segmentoj estas en la kapo. La kvar ĉefaj grupoj de artropodoj – nome kelikeruloj (inkludas araneojn kaj skorpiojn), krustuloj (salikokoj, omaroj, kraboj, ktp.), traĥeuloj (artropodoj kiuj spiras tra kanaloj en siaj korpoj; inklude insektojn kaj miriapodojn), kaj la formortintaj trilobitoj – havas kapojn formitaj de variaj kombinoj de segmentoj, kun apendicoj kiuj estas perditaj aŭ specializitaj laŭ diversaj vojoj.[9] Aldone kelkaj formortintaj artropodoj, kiaj Marrella, apartenas al neniu el tiuj grupoj, ĉar ties kapoj estas formataj de siaj propraj partikularaj kombinoj de segmentoj kaj specializitaj apendicoj.[16]
Prilabori la evoluciajn stadiojn laŭ kiuj ĉiuj tiuj diversaj kombinoj povus esti aperintaj estas tiom malfacile ke oni konis delonge kiel "la artropodkapa problemo".[17] En 1960 R. E. Snodgrass eĉ montris deziron ke ĝi ne estu solvita, ĉar klopodi prilabori solvojn estis multe pli amuza.[18]
Artropodaj ekzoskeletoj estas formataj de kutiklo, ne-ĉela materialo sekreciata de la epidermo.[19] Ties kutikloj varis en la detaloj de ties strukturo, sed ĝenerale konsistas el tri ĉefaj tavoloj: nome epikutiklo, fajna ekstera vakseca mantelo kiu defendas kontraŭ malsekeco la aliajn tavolojn kaj havigas krome iom da protekto; la eksokutiklo, kiu konsistas el ĥitino kaj kemie plifortigitaj proteinoj; kaj la endokutiklo, kiu konsistas el ĥitino kaj nefortigitaj proteinoj. La eksokutiklo kaj endokutiklo kune estas konataj kiel prokutiklo.[20] Ĉiu korposegmento kaj membroparto estas enskatoligita en fortigita kutiklo. la kuniĝoj inter korposegmentoj kaj inter membropartoj estas kovritaj de fleksebla kutiklo.[21]
La ekzoskeleto de plej akvaj krustuloj estas biomineraligitaj per kalcia karbonato elprenita el akvo. Kelkaj surteraj krustacoj estis disvolvigintaj rimedojn por stoki la mineralon, ĉar surtere ili ne povas dependi el preta havigo de solvita kalcia karbonato.[22] Biomineraligo ĝenerale tuŝas la eksokutiklon kaj la eksteran parton de la endokutiklo.[23] Du ĵusaj hipotezoj pri la evoluo de biomineraligo en artropodoj kaj aliaj grupoj de animaloj proponas ke ĝi havigas pli fortan defendan armaĵon,[24] kaj ke ĝi ebligas al animaloj kreskiĝi pli grande kaj pli forte havigante al ili pli rigidajn skeletojn;[25] kaj ajnaokaze mineral-organike komponita ekzoskeleto estas pli malmultekosta konstruebla ol tut-organika ekzoskeleto de komparebla forto.[26][27]
La kutiklo povas havi ĥetojn (haregoj) kiuj kreskiĝas el specialaj ĉeloj en la epidermo. Ĥetoj estas variaj laŭ formo kaj funkcio kiel apendicoj. Por ekzemplo, ili estas ofte uzataj kiel sentiloj por detekti aeron aŭ akvofluojn, aŭ kontakton kun objektoj; akvaj artropodoj uzas plumecajn ĥetojn por pliigi la surfacan areon de naĝ-apendicoj kaj por filtri manĝerojn el la akvo; akvaj insektoj, kiuj estas aer-spirantoj, uzas dikajn mantelojn el ĥetoj por kapti aeron, etendante la tempon kiun ili povas esti subakve; fortaj, rigidaj ĥetoj utilas kiel defendaj spinoj.[28]
Kvankam ĉiuj artropodoj uzas muskolojn ligitajn al la interno de ekzoskeleto por fleksi siajn membrojn, kelkaj ankoraŭ uzas hidraŭlikan premon por etendi ilin, nome sistemo heredita el ties pra-artropodaj prauloj;[29] por ekzemplo, ĉiuj araneoj etendas siajn krurojn hidraŭlike kaj povas generi premojn de ĝis ok fojoj ties ripoza nivelo.[30]
La ekzoskeleto ne povas etendiĝi kaj tiele limigas kreskon. Artropodoj tiele anstataŭas sian ekzoskeleton pere de mudo, aŭ forigo de la malnova ekzoskeleto post kresko de nova skeleto kiu ne estas ankoraŭ plifortigita. Mudocikloj povas okazi flue preskaŭ kontinue ĝis la artropodo atingas plenan grandon.[31]
En la komenca fazo de mudo, la animalo ĉesas manĝi kaj ties epidermo liverigas mudofluaĵon, nome mikso de enzimoj kiuj digestas la endokutiklon kaj tiele forigas la malnovan kutiklon. Tiu fazo ekas kiam la epidermo estis sekreciinta novan epikutiklon por protekti ĝin el la enzimoj, kaj la epidermo sekrecias la novan eksokutiklon dum la malnova kutiklo estas forigata. Kiam tiu stadio estas kompleta, la animalo faras ke sia korpo ŝvelas prenante grandan kvanton da akvo aŭ aero, kaj tio faras ke la malnova kutiklo disiĝas laŭlonge de antaŭdifinita malfortejo kie la malnova eksokutiklo estas pli fajna. Plej ofte tio bezonas kelkajn minutojn por ke la animalo elŝoviĝas el la malnova kutiklo. Je tiu punkto la nova estas elĉerpita kaj tiom milda ke la animalo ne povas subteni sin kaj malfacile moveblas, kaj la nova endokutiklo ankoraŭ ne estis formata. La animalo plue pligrandigas sin por etendi la novan kutiklon tiom multe kiom eblas, poste plifortigas la novan eksokutiklon kaj forigas la troan aeron aŭ akvon. Fine de tiu fazo la nova endokutiklo estis formita. Multaj artropodoj tiam manĝas la forigitan kutiklon por rekuperi ties materialojn.[31]
Ĉar artropodoj estas neprotektitaj kaj preskaŭ senmovigitaj ĝis la nova kutiklo estas plifortigita, ili estas en danĝero ĉar povas esti kaptitaj en la malnova kutiklo kaj povas esti atakita de predantoj. Mudo povas esti tialo de 80 al 90% de ĉiuj mortoj de artropodoj.[31]
Artropodaj korpoj estas segmentataj ankaŭ interne, kaj la nerva, muskola, cirkula, kaj ekskreta sistemoj havas ripetajn komponantojn.[9] Artropodoj venas el stirpo de animaloj kiuj havas celomon, nome membran-linia kavaĵo inter la ventro kaj la korpomuro kiu protektas la internajn organojn. La fortaj, segmentataj membroj de artropodoj nuligas la neceson de unu el la ĉefaj praaj funkcioj de la celomo, kiel hidrostata skeleto, kiun la muskoloj premas por ŝanĝi la formon de la animalo kaj tiele ebligi ĝin moviĝi. El tio la celomo de la artropodo estas limigita al malgrandaj areoj ĉirkaŭ la reprodukta kaj ekskreta sistemoj. Ties loko estas tre okupita de hemocelo, nome kavaĵo kiu etendas plej el la longo de la korpo kaj tra kiu la sango fluas.[32]
Artropodoj havas malfermajn cirkulajn sistemojn, kvankam plej havas kelkajn mallongajn, malferm-finajn arteriojn. Ĉe keliceruloj kaj krustacoj, la sango portas oksigenon al la histoj, dum heksapodoj uzas separatan sistemon de traĥeoj. Multaj krustacoj, sed malmultaj keliceruloj kaj traĥeuloj, uzas spiran pigmenton por helpi la transporton de oksigeno. La plej ofta spira pigmento ĉe artropodoj estas kupro-bazita hemocianino; tiu estas uzata de multaj krustacoj kaj kelkaj centpieduloj. Kelkaj krustacoj kaj insektoj uzas fer-bazitan hemoglobinon, nome la spira pigmento uzata de vertebruloj. Kiel ĉe aliaj senvertebruloj, la spiraj pigmentoj de tiuj artropodoj kiuj havas ilin estas ĝenerale dissolvitaj en la sango kaj rare enfermitaj en korpuskloj kiel ili estas en vertebruloj.[32]
La koro estas tipe muskola tubo kiu etendas ĝuste sub la dorso kaj laŭ plej el la longo de hemocelo. Ĝi kuntiriĝas laŭ ondoj kiuj iras el malantaŭe antaŭen, pelanta sangon antaŭen. Sekcioj kiuj ne estas premitaj de la kormuskolo estas etenditaj ĉu de elastaj ligamentoj aŭ de malgrandaj muskoloj, ĉiam konektante la koron al la korpomuro. Laŭlonge de la koro estas serio de paraj ostia, nome ne-revenaj valvoj kiuj ebligas ke la sango eniru al la koro sed evitas ke ĝi eliru antaŭ ĝi atingas la antaŭon.[32]
Artropodoj havas ampleksan varion de spiraj sistemoj. Malgrandaj specioj ofte ne havas tion, ĉar ties alta proporcio de surfaca areo al volum ebligas simplan disvastigon tra la korpa surfaco por havigi sufiĉan oksigenon. Krustacoj kutime havas brankojn kiuj estas modifitaj apendicoj. Multaj araknidoj havas folipulmojn.[33] Traĥeoj, sistemoj de brankaj tuneloj kiuj estas el la malfermaĵoj de la korpomuroj, liveras oksigenon rekte al la unuopaj ĉeloj ĉe multaj insektoj, miriapodoj kaj araknidoj.[34]
Vivantaj artropodoj havas parajn ĉefe nervajn ĥordojn laŭlonge de siaj korpoj sub la ventro, kaj en ĉiu segmento la ĥordoj formas paron de ganglioj el kiuj la sensaj kaj movaj nervoj estas cxe aliaj partoj de la segmento. Kvankam la paroj de ganglioj en ĉiu segmento ofte aperas fizike fuziitaj, ili estas konektitaj pere de komisuroj (nme relative grandaj faskoj da nervoj), kiuj donas al la artropoda nervosistemoj karakterajn "ŝtuparecan" aspekton. La cerbo estas en la kapo, encirklante kaj ĉefe super la ezofago. Ĝi konsistas el fuziitaj ganglioj de la prostomium kaj unu aŭ du el la plej antaŭaj segmentoj kiuj formas la kapon – totalo de tri paroj de ganglioj ĉe plej artropodoj, sed nur du ĉe keliceruloj, kiuj ne havas antenojn aŭ la ganglion konektita el ili. La ganglio de aliaj kaposegmentoj estas ofte proksima al la cerbo kaj funkcias kiel ties parto. Ĉe insektoj tiuj aliaj kapoganglioj kombinas en unu paro de subezofagaj ganglioj, sub kaj malantaŭ la ezofago. Araneoj faras tiun procezon iom plue, ĉar ĉiuj la segmentaj ganglioj estas aligitaj en la subezofagaj ganglioj, kiuj okupas plej el la spaco en la cefalotorako (antaŭa "super-segmento").[35]
Estas du diferencaj tipoj de artropodaj ekskretaj sistemoj. En akvaj artropodoj, la fina produkto de biokemiaj reagoj kiuj metabolas nitrogenon estas amoniako, kiu estas tiom venena ke ĝi bezonas esti solvita en akvo tiom multe kiom eblas. La amoniako estas tiam forigita tra iu permeabla membrano, ĉefe tra la brankoj.[33] Ĉiuj krustacoj uzas tiun sistemon, kaj ties alta konsumo de akvo povas esti responsa pri la relativa manko de sukceso de krustacoj kiel surteraj animaloj.[36] Variaj grupoj de surteraj artropodoj sendepende disvolvigis diferencan sistemon: la fina produkto de nitrogena metabolo estas ureata acido, kiu povas esti ekskreciata kiel seka materialo; la Malpiga tubo filtras la ureatan acidon kaj alian nitrogenan forĵetaĵon ekster la sango en la hemocelon, kaj oni forigas tiujn materialojn en la malantaŭan inteston, el kiu ili estas elpelitaj kiel fekaĵo.[36] Plej akvaj artropodoj kaj kelkaj surteremaj havas ankaŭ organojn nome nefridioj ("malgrandaj renoj"), kiuj elprenas aliajn forĵetaĵojn por ekskretado kiel urino.[36]
La rigidaj kutikloj de artropodoj forblokigus informaron pri la ekstera mondo, escepte ke ili estas penetrataj de multaj sensiloj aŭ konektiloj el sensiloj al la nervosistemo. Fakte, artropodoj havas modifitajn kutiklojn en prilaboritaj radioj de sensiloj. Variaj tuŝosensiloj, ĉefe ĥetoj, rilatas al diferencaj niveloj de forto, el forta kontakto al tre malfortaj aerfluoj. Kemiaj sensiloj havigas ekvivalentojn de gusto kaj flaro, ofte pere de ĥetoj. Premosensiloj ofte havas la formon de membranoj kiuj funkcias kiel miringoj, sed estas konektitaj rekte al nervoj pli ol al la orelostetoj. La antenoj de plej heksapodoj inkludas sensilarojn kiuj monitoras humidecon, humidecon kaj temperaturon.[37]
Plej artropodoj havas kompleksajn vidsistemojn kiuj inkludas unu aŭ pli ofte ambaŭ el komponitaj okuloj kaj simplajn pigmentotasajn okulojn ("okuletoj"). Plej ofte ocelli estas kapablaj nur detekti la direkton el kiu venas la lumo, uzante la ombron ĉe la tasomuroj. Tamen, la okuloj de araneoj estas pigmentotasaj ocelli kiuj estas kapablaj formi bildojn,[37] kaj tiuj de saltaraneoj povas turniĝi por kapti predon.[38]
Komponitaj okuloj konsistas el dekkvin al kelkaj miloj da sendependaj ommatidia, kolonoj kiuj estas kutime seslateraj en krucosekco. Ĉiu ommatidium estas sendependa sensilo, kun siaj propraj lum-sensivaj ĉeloj kaj ofte kun siaj propraj okullensoj kaj korneoj.[37] Komponitaj okuloj havas ampleksan vidkampon, kaj povas detekti rapidajn movojn kaj foje la lumpolarizadon.[39] Aliflanke, la relativa grando de ommatidia faras la bildojn iom fuŝaj, kaj komponitaj okuloj estas pli mallongaj - kompare kun tiuj de birdoj kaj mamuloj – kvankam tiu ne estas akra malavantaĝo, ĉar objektoj kaj okazoj ene de 20 centimetroj estas plej gravaj por plej artropodoj.[37] Kelkaj artropodoj havas koloran vidkapablon, kaj tiu de kelkaj insektoj estis studita detale; por ekzemplo, la ommatidia de abeloj enhavas ricevilojn por kaj verda kaj ultraviola.[37]
Plej artropodoj ne havas sensilojn por ekvilibro kaj akcelo, kaj dependas el siaj okuloj por diri al ili kien iri. La memfida konduto de blatoj estas ŝaltita kiam premosensiloj en la suba flanko de la piedoj ne informas premon. Tamen, multaj malakostrakaj krustacoj havas statocistojn, kiuj havigas la saman specon de informaro kiel la ekvilibraj kaj movajn sensilojn de la vertebrula interna orelo.[37]
La proprioceptoj de artropodoj, nome sensiloj kiuj informas pri la forto plenumita de muskoloj kaj pri la grado kliniĝi en la korpo kaj artikloj, estas bone komprenitaj. Tamen oni scias malmulte pri kiujn aliajn internajn sensilojn artropodoj povas havi.[37]
Insekta flarkapablo referencas al la funkcio de kemiaj riceviloj kiuj ebligas insektojn por detekti kaj identigi volatilajn komponaĵojn por manĝado, predevito, trovado de partneroj por pariĝado (pere de feromonoj) kaj lokigo de habitatoj por ovodemetado.[40] Tiele, ĝi estas la plej grava senso por insektoj.[40] Plej gravaj insektokondutoj devas esti tempigitaj perfekte kio estas dependa el tio kion ili flaras kaj el kiam ili flaras ĝin.[41] Por ekzemplo flarado estas esenca por ĉasado ĉe multaj specioj de vespoj, inklude Polybia sericea.
La du organoj kiujn insektoj ĉefe uzas por detekti odoroj estas la antenoj kaj specializitaj buŝopartoj nome makzelaj palpiloj.[42] Ene de tiuj flaraj organoj estas neŭronoj nome flar-ricevaj neŭronoj kiuj, kiel la nomo sugestas, estas hejmoriceviloj por odoraj molekuloj en ties ĉelmembranoj. La majoritato de flar-ricevaj neŭronoj tipe kuŝas en la antenoj. Tiuj neŭronoj povas esti tre abundaj, por ekzemplo muŝoj de la genro Drosophila havas ĉirkaŭ 2,600 flar-ricevaj neŭronoj.[42]
Insektoj estas kapablaj flari kaj diferenci inter miloj da volatilaj komponaĵoj kaj sensive kaj selektive.[40][43]
______ Heksapodoj (insektoj, dipluroj, proturoj, kolemboloj) _________/ / ______ Krustacoj (= krustuloj; kraboj, langustoj, saliketo, ktp) / / ___ Paŭropodoj / / / /____ Diplopodoj (milpieduloj) /____Miriapodoj_/ / _____ Kilopodoj (centpieduloj) / __/ ___ Simfiloj _____ Araneoidoj (araneoj, akaroj, skorpioj, ktp) / ___Ĥeliceruloj_/______ Merostomoj _____ Piknogonidoj



Artropodoj aŭ artikpieduloj, (science: Arthropoda; el la greka αρθρόποδα [arthropoda], arthro-, artiko + podos, piedo) estas subaro de la duflankuloj, kun taksonomia kategorio de filumo. Ili ne havas spinon, kaj havas ĝenerale malmolan ŝelon. Nome temas pri senvertebraj animaloj kiu havas ekzoskeleton (ekstera skeleto), segmentan korpon, kaj artiklitajn membrojn (parigitaj membroj). La filumo estas dividita en 5 subfilumoj: keliceratoj, heksapodoj, krustacoj, miriapodoj kaj † trilobuloj, kiuj inkluzivas proksimume 1,257,040 speciojn.
Artropodoj estas karakterizitaj per siaj artiklitaj membroj kaj kutiklo farita el kitino, ofte mineraligita per kalcia karbonato. La artropoda korpoformo konsistas el segmentoj, kiu havas po paron de membroj. La rigida kutiklo malhelpas kreskon, kaj tiele artropodoj anstataŭas ĝin periode pere de mudo. Ties adaptablo permesis al ili iĝi la plej speci-riĉaj membroj de ĉiuj speicaroj en plej medioj. Ili havas ĉirkaŭ milionon da priskribitaj specioj, kio faras ĝin pli ol 80% el ĉiuj priskribitaj vivantaj bestospecioj, kelkaj el kiuj, malkiel plej animaloj, estas tre sukcesaj eĉ en sekegaj medioj.
Artropodoj gamas laŭ grando el la mikroskopa krustaco Stygotantulus ĝis la Japana aranekrabo. La ĉefa interna kavaĵo de artropodoj estas sangujo, kiu enhavas ties internajn organojn, kaj tra kiu ties hemolimfo - similaĵo de sango - cirkulas; ili havas malfermajn cirkulajn sistemojn. Kiel ĉe ties eksteraĵoj, la internaj organoj de artropodoj estas ĝenerale konstruitaj de ripetaj segmentoj. Ties nerva sistemo estas "ŝtupareca", kun parigitaj ventraj nervofadenoj kiuj iras tra ĉiuj segmentoj kaj formas parigitajn gangliojn en ĉiu segmento. Ties kapoj estas formataj de fuzio de variaj nombroj de segmentoj, kaj ties cerboj estas formataj per fuzio de la ganglioj de tiuj segmentoj kaj ĉirkaŭas la ezofagon. La spira kaj ekskrecia sistemoj de artropodoj varias, depende multon el sia medio kaj el la subfilumo al kiu ili apartenas.
Ties vidkapablo dependas el variaj kombinoj de kompleksaj okuloj kaj pigment-truaj oceloj: ĉe plej specioj la oceloj povas detekti nur la direkton el kiu lumo venas, kaj la kompleksaj okuloj estas la ĉefa fonto de informaro, sed la ĉefaj okuloj de araneoj estas oceloj kiuj povas formi bildojn kaj, ĉe kelkaj okazoj, povas turniĝi por vidokapti predon. Artropodoj havas ankaŭ ampleksajn gamojn de kemiaj kaj mekanikaj sensiloj, ĉefe bazite sur modifoj de multaj ĥetoj (haretaĵoj) kiuj projektas tra siaj kutikloj. La metodoj de reproduktado kaj disvolvigo de artropodoj estas diversaj; ĉiuj surteraj specioj uzas internan fekundigon, sed tio okazas ofte per nerekta transigo de spermo tra membro aŭ la grundo, pli ol per rekta injekto. Akvaj specioj uzas ĉu internan aŭ eksteran fekundigon. Preskaŭ ĉiuj artropodoj demetas ovojn, sed skorpioj naskas vivajn junulojn post la idoj eloviĝis ene de la patrino. Artropodidoj varias el etegaj plenkreskuloj al larvoj kaj raŭpoj kiuj ne havas artiklitajn membrojn kaj finfine suferas totalan metamorfozon por produkti la plenkreskan formon. La nivelo de patra zorgado por idoj varias el nenio al longdaŭra zorgado havigita de skorpioj.
La evolucia praulo de artropodoj datas el la Kambria periodo. La grupo estas ĝenerale rigardita kiel monofiletika, kaj multaj analizoj eltenas la lokigon de artropodoj kun cycloneuralia (aŭ ties konstituantaj kladoj) en superfilumo de muduloj (Ecdysozoa). Ĝenerale tamen, la bazaj rilatoj de Metazoa ne estas ankoraŭ bone solvitaj. Same, la rilatoj inter variaj artropodaj gropoj estas ankoraŭ aktive debatitaj.
Artropodoj kontribuas al la homa manĝaĵaro kaj rekte kiel manĝo, kaj pli grave kiel polenigantoj de plantoj, ĉefe tiuj gravaj por agrikulturo aŭ bestobredado. Kelkaj specifaj specioj estas konataj ĉar disvastigas kelkajn malsanojn al homoj, gregaro, kaj rikoltoj.
Los artrópodos (Arthropoda, del griego ἄρθρον, árthron, «articulación» y πούς, poús, «pie»), constituyen el filo más numeroso y diverso del reino animal (Animalia). El grupo incluye animales invertebrados dotados de un esqueleto externo y apéndices articulados; los insectos, arácnidos, crustáceos y miriápodos, entre otros.
Hay más de 1 300 000 especies descritas,[1] en su mayoría insectos (estimados entre 941 000 a 1 000 000 de especies),[2][3][4] que representan al menos el 80 % de todas las especies animales conocidas.[5] Son importantes miembros de ecosistemas marinos, de agua dulce, terrestres y aéreos. Varios grupos de artrópodos están perfectamente adaptados a la vida en ambientes secos, al igual que los vertebrados amniotas y a diferencia de todos los demás filos de animales, que son acuáticos o requieren ambientes húmedos.
Si bien no hay una especialidad de la zoología que estudia específicamente a todos los artrópodos en su conjunto, hay ciencias que estudian específicamente a los insectos (entomología), arácnidos (aracnología) y crustáceos (carcinología).
Los artrópodos fueron clasificados tradicionalmente junto con los anélidos dentro del clado Articulata.[6] Por lo tanto, se creía que los primeros artrópodos podrían haber sido similares a los anélidos; aunque dado que los restos fósiles de anélidos eran poco comunes, la clasificación de los artrópodos en Articulata estaba basada principalmente en similitudes morfológicas.[7]
El amplio registro fósil de los artrópodos y los análisis moleculares descartan la agrupación en Articulata, incluyendo a los artrópodos en el clado Ecdysozoa; y a los anélidos, en Spiralia. Actualmente, se piensa que los primeros artrópodos fueron animales pequeños, segmentados, y con apéndices; denominados lobópodos. Se cree que estos animales, entre otros grupos de clasificación incierta, representan varios taxones del grupo troncal del filo Arthropoda.[8]
Por otra parte, se ha propuesto que ciertos organismos pertenecientes a la biota de Ediacara (de hace 555 millones de años) están relacionadas con los artrópodos.[9][10][11] Dichos animales del periodo Ediacárico son formas "seudo-bilaterales" que han sido incluidos dentro del grupo Proarticulata,[12] como los géneros Dickinsonia, Spriggina y Vendia. Aunque poseen una segmentación diferente a la de los actuales artrópodos.
Los artrópodos constituyen uno de los grandes filos del reino animal, subdividido en diversas clases, algunas de las cuales cuentan con gran número de géneros y especies. Se los denomina de esta manera por estar provistos de patas articuladas. Todo el cuerpo de los artrópodos está formado por varios segmentos unidos entre sí por medio de membranas articulares.
A pesar de su gran variedad y su disparidad, los artrópodos poseen en común características morfológicas y fisiológicas fundamentales:
El exoesqueleto de los artrópodos es una cubierta continua llamada cutícula, que se extiende incluso por los dos extremos del tubo digestivo y por las vías o cavidades respiratorias, y que está situada por encima de la epidermis (llamada en estos por ese motivo hipodermis), que es la que la secreta.
La composición del exoesqueleto es glucopeptídica (con una parte glucídica y una parte peptídica). El componente principal y más característico pertenece al primero de estos dos tipos, y es la quitina, un polisacárido derivado del aminoazúcar N-acetil-2-D-glucosamina que se encuentra también, por ejemplo, en la pared celular de los hongos. En muchos casos la consistencia del exoesqueleto gana por el añadido de sustancias minerales, como en el caso de los cangrejos y otros crustáceos decápodos cuya cutícula aparece calcificada, por depósito de carbonato cálcico.
El espesor y dureza de la cutícula no es igual en toda su extensión. Por el contrario, aparece formando zonas endurecidas llamadas escleritos, separadas o unidas entre sí por zonas más delgadas y flexibles. Los escleritos reciben denominaciones complejas que varían en cada grupo, pero de manera general se denominan terguitos a los de ubicación dorsal; esternitos, los de ubicación ventral; y pleuritos, los laterales. Pueden existir además crestas del exoesqueleto desarrolladas hacia adentro llamadas apodemas y otras llamadas apófisis, ambas invaginaciones de la pared del cuerpo forman procesos rígidos que sirven para la inserción de músculos y para dar fortaleza o rigidez al exoesqueleto.[17]
El exoesqueleto está estructurado en las siguientes capas:
La cutícula aparece muy frecuentemente cubierta de quetas (pelos) de diversa función, incluida la sensorial táctil.
La coloración de los artrópodos suele depender de la cutícula. En la procutícula se depositan pigmentos coloreados o cristales de guanina. La epicutícula puede presentar estriaciones finas que producen colores físicos (no químicos), como la apariencia metálica o irisada de muchos insectos.
El esqueleto externo tiene una desventaja y es que, para poder crecer, el animal debe desprenderse de él. Lo hace en un proceso, controlado hormonalmente, de ecdisis o muda. La hipodermis secreta enzimas que ablandan y digieren en parte la capa más inferior de la cutícula (la endocutícula), provocando que el resto se desprenda. Inmediatamente comienza la secreción de una cutícula nueva, primero la epicutícula y luego, debajo de ella, la procutícula. Hasta que se endurece esta nueva cubierta el animal está relativamente indefenso, con menos posibilidad de escapar o resistirse. Todo el proceso de la muda está controlado hormonalmente; la ecdisona u "hormona de la muda" es la sustancia responsable de que estos cambios se produzcan. Se llaman estadios o instares a las sucesivas fases de la existencia del animal entre muda y muda. Este rasgo lo comparten los artrópodos con algunos otros filos, como los nemátodos que también tienen una cutícula y mudan.
Los apéndices son expansiones articuladas del exoesqueleto, en cuyo interior se sitúan los músculos estriados que, adheridos a las articulaciones, les proporcionan versatilidad y rapidez de movimientos. Se llama artejos (voz que deriva del latín artículo, "articulado") a las piezas articuladas que forman los apéndices.
Existen dos tipos básicos de apéndices, los unirrámeos, formados por un solo eje, propios de los artrópodos terrestres (arácnidos, miriápodos e insectos); y los birrámeos formados por dos ejes y propios de los artrópodos acuáticos (trilobites y crustáceos). No hay acuerdo sobre cuál fue el apéndice ancestral.
En el curso de la evolución ha existido la tendencia a restringir los apéndices a determinadas regiones del cuerpo y a especializarlos para funciones distintas. Los apéndices de la cabeza están adaptados para la percepción sensorial, la defensa y para manipular los alimentos; los del tórax sirven para la locomoción y natación; los abdominales cumplen funciones respiratorias y reproductoras, como retener los huevos o aferrarse a la pareja durante la cópula. Otros se han modificado de tal modo que cuesta reconocerlos como tales (hileras de las arañas, peines de los escorpiones).
El aparato digestivo de los artrópodos se divide en tres regiones bien diferenciadas, el estomodeo, el mesodeo y el proctodeo. Estomodeo y proctodeo son las regiones situadas en el extremo anterior y posterior, respectivamente; están recubiertas de cutícula que se renueva cada vez que el animal muda. La parte media del tubo digestivo, el mesodeo, deriva del endodermo (segunda hoja blastodérmica) y es la que produce las secreciones digestivas y donde se realiza la mayor parte de la absorción de nutrientes; frecuentemente presenta derivaciones o ciegos laterales que amplían su superficie.
Muchos artrópodos son demasiado pequeños como para tener o necesitar órganos respiratorios. Los artrópodos acuáticos suelen presentar branquias, apéndices internamente más vascularizados que los otros órganos. Se encuentran en los crustáceos, como especializaciones de la rama dorsal de los apéndices torácicos, y de la misma manera en los xifosuros o en los euriptéridos o los primeros escorpiones fósiles. También se encuentran branquias secundarias (derivadas de las tráqueas) en las larvas acuáticas de algunos insectos, como las efímeras.
Como es general en los animales, los miembros del grupo de vida aérea respiran por órganos internalizados, que en los artrópodos pueden ser de dos tipos:
El aparato circulatorio de los artrópodos es abierto, es decir, no existe un circuito cerrado de vasos por el que circule un líquido diferenciado, lo que propiamente se podría llamar sangre. Lo que existe es un vaso especializado que recorre longitudinalmente gran parte del cuerpo llamado corazón dorsal. Al contraerse dicho órgano, mueve el líquido corporal interno: la hemolinfa, que recibe de vasos posteriores abiertos e impulsa hacia adelante por vasos igualmente abiertos (separados mediante válvulas). El tramo del vaso que se encuentra conectado al cerebro se denomina aorta, en el cual irriga directamente la cavidad cerebral y los órganos cercanos a este para luego retornar a la red de vasos y llegar nuevamente al corazón dorsal. La hemolinfa llega a las extremidades a través de bombas musculares que, con la ayuda de contracciones musculares, actúan como "corazones auxiliares" permitiendo al fluido circulatorio alcanzar aquella zona del cuerpo.[18][19]
La red de vasos está siempre escasamente desarrollada, salvo en las branquias de los artrópodos acuáticos. No hay células sanguíneas especializadas en el transporte de oxígeno, aunque, como en todos los animales existen amebocitos (células ameboideas) con funciones de inmunidad celular y hemostasis (coagulación y cicatrización). Sí puede haber pigmentos respiratorios, pero disueltos en la hemolinfa.[19]
Los crustáceos presentan para la excreción glándulas antenales y maxilares, en la base de esos apéndices. Los arácnidos suelen disponer de glándulas coxales, que desembocan en la base de las patas locomotoras. En insectos y en miriápodos aparecen órganos tubulares característicos, llamados tubos o conductos de Malpighi, que desembocan entre el intestino medio y el intestino posterior (proctodeo); sus productos se suman a la composición de las heces.
Los artrópodos terrestres suelen ser uricotélicos, es decir, que para la excreción nitrogenada no producen amoníaco o urea, sino ácido úrico o, a veces, guanina.
En los artrópodos es frecuente la excreción por acumulación, como alternativa o complemento de la excreción por secreción. En este caso se acumulan los productos de excreción en nefrocitos, células pericárdicas o directamente en la cutícula. La acumulación suele ser de uratos o guanina, bases nitrogenadas muy poco solubles que forman depósitos sólidos. En este último caso las mudas sirven para la función añadida de librarse de esas excretas.
Como corresponde a los protóstomos, el sistema nervioso se desarrolla en el lado ventral del cuerpo, y como corresponde a animales metaméricos, su organización es segmentaria. En cada segmento aparece un par de ganglios, de posición más o menos ventrolateral, con los dos ganglios de un par soldados o unidos por una comisura transversal y los de pares consecutivos unidos por nervios conectivos.
En los artrópodos es un órgano de tipo anelidiano, por tanto, tiene una estructura primariamente en forma de escalera de cuerda, o sea, dos cordones nerviosos longitudinales que recorren la parte ventral del cuerpo, con un par de ganglios por metámero unidos transversalmente por comisuras; no obstante, se producen procesos de concentración de ganglios debidos a la formación de tagmas.
Normalmente está formado por tres pares de ganglios que se asocian, correspondientes al procéfalon. Se pueden diferenciar tres regiones:
En el protocerebro y deutocerebro, no se diferencian comisuras ni conectivos. El tritocerebro está formado por un par de ganglios que se unen a los anteriores en las cabezas denominadas tritocefálicas, perdiéndose los conectivos, mientras que en las cabezas deutocefálicas, se mantiene independiente, conservando los conectivos con el deutocerebro. Esto ocurre en algunos crustáceos como branquiópodos o cefalocáridos. En todos los casos, se diferencia la comisura, que es subesofágica.
Dentro de la cápsula cefálica, el cerebro tiene posición vertical; el protocerebro y el deutocerebro se sitúan hacia arriba, y el tritocerebro es inferior y se dirige hacia atrás.
Está formada por un par de ganglios por metámero que en principio presentan conectivos y comisuras. En grupos primitivos, los ganglios de cada par de segmentos se presentan disociados, y la estructura recuerda a una escalera de cuerda. Los grados de concentración y de acortamiento se deben a la supresión de las comisuras y los conectivos respectivamente.
Destaca el ganglio subesofágico; en hexápodos es resultado de la fusión de tres pares de ganglios ventrales correspondientes a los metámeros IV, V y VI e inerva las piezas bucales (las mandíbulas y los dos pares de maxilas) y por ello se llama gnatocerebro; en los decápodos, son seis los ganglios que se asocian (pues se incluyen los tres ganglios de los maxilípedos.
Neuronas sensitivas y motoras que forman ganglios y que se sitúan sobre las paredes del estomodeo. Este sistema está relacionado con el sistema nervioso central y con el sistema endocrino. En el sistema nervioso simpático se diferencian dos partes.
La mayoría de los artrópodos están dotados de ojos, de los que existen varios modelos distintos.
La visión de muchos artrópodos presenta ventajas que suelen faltar en vertebrados, como la habilidad para ver en un espectro extendido que incluye el ultravioleta próximo, o para distinguir la dirección de polarización de la luz. La visión del color está casi siempre presente y puede ser muy rica; el crustáceo Squilla mantis presenta trece pigmentos distintos con diferente sensibilidad al color, lo que contrasta con el pobre sistema tricromático (de tres pigmentos) de la mayoría de los primates, incluida nuestra especie.
Distribuidos por todo el cuerpo, pueden encontrarse sensilias, que son receptores sensibles a los estímulos químicos, como los del gusto o el olfato, y receptores táctiles, asociado a antenas y palpos y también a setas táctiles, pelos que está asociados a una célula sensible. Algunos insectos disponen de un sentido del oído, lo que es revelado por la existencia de señales auditivas de comunicación intraespecífica, como por ejemplo en los grillos. Muchos son sensibles a las vibraciones del suelo, por las que detectan la presencia de presas o depredadores; otros, como las moscas, poseen tricobotrios capaces de percibir mínimos cambios de presión ambiental.
Los artrópodos suelen estar dotados de sensores de posición, sencillos, pero eficaces, que les ayudan a mantener la posición y el equilibrio, como los órganos cordotonales que un díptero tiene en los halterios.
En la reproducción sexual, las hembras, tras ser fecundadas por los machos, ponen huevos. El desarrollo, a partir del huevo, puede ser directo o indirecto.
Se dan frecuentes casos de partenogénesis (la hembra produce un cigoto sin haber sido fecundada), sobre todo en crustáceos e insectos. También hay casos de reproducción por embriogénesis que es uno de los tipos de fragmentación donde las larvas, estados jóvenes o embrionarios se dividen en nuevos individuos.
También se dan casos de hermafroditismo que aparecen sobre todo en especies parásitas o sésiles.
Durante muchas décadas, las relaciones filogenéticas de los celomados se basaron en la concepción de los articulados (Articulata) de Cuvier,[20] un clado formado por anélidos y artrópodos. Numerosos análisis morfológicos modernos basados en principios cladistas han corroborado la existencia del clado Articulata, por ejemplo, Brusca & Brusca,[3] Nielsen[15] o Nielsen et al.,[16] entre otros.
No obstante, diversos análisis cladísticos como el de datos combinados de Zrzavý et al (1998)[13] y otros han llegando a la conclusión de que anélidos y artrópodos no están directamente relacionados. La presencia de metamerización en anélidos y artrópodos debe considerarse un caso de convergencia evolutiva. Por el contrario, estos estudios propusieron el clado Ecdysozoa en el que los artrópodos muestran estrechas relaciones filogenéticas con grupos pseudocelomados, como nematodos, nematomorfos, priapúlidos y quinorrincos, por la presencia compartida de una cutícula quitinosa y un proceso de muda (ecdisis) de la misma. Ecdysozoa, en su definición actual, incluye a los miembros de tres subclados putativos: Nematoida (Nematoda y Nematomorpha), Scalidophora (Priapulida, Loricifera y Kinorhyncha) y Panarthropoda (Arthropoda, Onychophora y Tardigrada).[14]
La filogenia de los artrópodos ha sido muy controvertida, con una enfrentada polémica entre los partidarios del monofiletismo y los del polifiletismo. Snodgrass[21] y Cisne[22] han defendido el monofiletismo, aunque el primero contempla los artrópodos divididos en arácnidos + mandibulados, y el segundo los interpreta divididos en esquizorrámeos y atelocerados. Tiegs & Manton[23] defendieron el difiletismo, con los artrópodos divididos en esquizorrámeos + unirrámeos y los onicóforos como grupo hermano de miriápodos + hexápodos. Posteriormente, Manton[24] y Anderson[25] sostuvieron el polifiletismo del grupo (ver Uniramia).
Con la aparición de los primeros estudios basados en datos moleculares y análisis combinados de datos morfológicos y moleculares, parece que la antigua polémica sobre monofilia y polifilia ha quedado superada, ya que todos ellos corroboran que los artrópodos son un grupo monofilético en el que incluyen también los onicóforos y tardígrados (el clado se ha dado en llamar panartrópodos); la mayoría también proponen la existencia del clado Mandibulata que reúne a los subfilos Hexapoda, Crustacea y Myriapoda. Principalmente se había propuesto que los hexápodos podrían estar emparentados con los crustáceos o los miriápodos,[26] sin embargo, la hipótesis de que estarían relacionados con los miriápodos se ha descartado en gran medida, debido a varios análisis moleculares y evidencias fósiles que sugieren que los crustáceos están estrechamente con los hexápodos y que de hecho son un grupo parafilético de estos últimos, por tanto las relaciones filogenéticas de los artrópodos quedarían de la siguiente manera:[27][28][29][30][31]
Arthropoda Chelicerata Prosomapoda Mandibulata Myriapoda Progoneata Edafopoda Pancrustacea Oligostraca Ichthyostraca Altocrustacea Multicrustacea Hexanauplia Thecostraca Allotriocarida Athalassocarida Labiocarida Hexapoda Ellipura EuentomataEl filo artrópodos (Arthropoda) se divide en cuatro subfilos.
Hexapoda (Insecta, Collembola, Diplura, Protura)
Crustacea (cangrejos, gambas, Isopoda, etc.)
MyriapodaDiplopoda (milpiés)
Chilopoda (ciempiés)
ChelicerataArachnida (arañas, escorpiones, etc.)
Eurypterida (gigantostráceos: extintos)
Xiphosura (xifosuros)
Pycnogonida (picnogónidos)
Trilobites (extinto)
Árbol filogenético de los artrópodos y grupos relacionados[34]
De acuerdo con otra propuesta clasificatoria,[4] el filo se divide en cinco subfilos:
Los artrópodos (Arthropoda, del griego ἄρθρον, árthron, «articulación» y πούς, poús, «pie»), constituyen el filo más numeroso y diverso del reino animal (Animalia). El grupo incluye animales invertebrados dotados de un esqueleto externo y apéndices articulados; los insectos, arácnidos, crustáceos y miriápodos, entre otros.
Hay más de 1 300 000 especies descritas, en su mayoría insectos (estimados entre 941 000 a 1 000 000 de especies), que representan al menos el 80 % de todas las especies animales conocidas. Son importantes miembros de ecosistemas marinos, de agua dulce, terrestres y aéreos. Varios grupos de artrópodos están perfectamente adaptados a la vida en ambientes secos, al igual que los vertebrados amniotas y a diferencia de todos los demás filos de animales, que son acuáticos o requieren ambientes húmedos.
Si bien no hay una especialidad de la zoología que estudia específicamente a todos los artrópodos en su conjunto, hay ciencias que estudian específicamente a los insectos (entomología), arácnidos (aracnología) y crustáceos (carcinología).
Lülijalgsed ehk artropoodid (Arthropoda) on loomariigi hõimkond, kuhu kuulub üle 80% tänapäevastest loomaliikidest. Nad kuuluvad alamriiki pärishulkraksed.
Lülijalgsete kehaehituse iseloomulikumateks tunnuseks on kitiinist välisskelett ja lüliline kehaehitus. Lülijalgsed jagunevad klassideks, millest suurimad on putukad (Insecta), ämblikulaadsed (Arachnida) ja vähid (Crustacea).
Lülijalgsed on traditsiooniliselt jagatud 5 alamhõimkonda, millest 1 on väljasurnud.
Lülijalgsed ehk artropoodid (Arthropoda) on loomariigi hõimkond, kuhu kuulub üle 80% tänapäevastest loomaliikidest. Nad kuuluvad alamriiki pärishulkraksed.
Lülijalgsete kehaehituse iseloomulikumateks tunnuseks on kitiinist välisskelett ja lüliline kehaehitus. Lülijalgsed jagunevad klassideks, millest suurimad on putukad (Insecta), ämblikulaadsed (Arachnida) ja vähid (Crustacea).
Artropodoak (antzinako grezieraz: ἄρθρον, arthron, "artikulazio" eta πούς, podos, "hanka") exoeskeleto bat, gorputz segmentatua eta apendize bikoitiak dituzten animalia ornogabeak dira. Artropodoek Euarthropoda filuma osatzen dute[1][2], eta euren barruan intsektuak, araknidoak, miriapodoak eta krustazeoak daude, besteak beste. Hasiera batean Arthropoda terminoa Euartropodoak eta onikoforoak izendatzeko erabili zen. Artropodoen ezaugarririk nabarmenena artikulatutako adarrak eta kitinaz osatutako kutikulak dira, batzuetan kaltzio karbonatoan mineralizatzen direnak. Artropodoen gorputz-planak segmentuak ditu, horietako bakoitza apendize pare batekin. Kutikula zurrunak hazkuntza gelditzen du, beraz artropodoek mudak egiten dituzte erregularki. Simetria bilaterala dute, eta gorputzak kanpo eskeletoa du. Espezie batzuek hegalak dituzte.
Bere aldakortasunari esker, elkarketa-ekologiko guztietako espezierik ohikoenak bihurtu dira ingurune gehienetan. Gutxi gorabehera milioi bat espezie deskribatu dira, eta animalia espezie guztien %80 baino gehiago dira. Horietako batzuk, beste animalia gehienek ez bezala, giro lehorrean arrakasta handia dute. Artropodoak neurriz aldatzen dira Stygotantulus krustazeo mikroskopikotik Japoniako Macrocheira kaempferi karramarro erraldoiraino.
Artropodoen barne kabitate nagusia hemozeloa da, bere barne-organoak biltzen dituena; bertatik, odolaren analogoa den hemolinfak zirkulatzen du. Euren zirkulazio-sistema irekia da. Kanpoko egituran gertatzen den bezala, artropodoen barne organoak ere errepikapen bidez eratu ohi dira. Euren nerbio-sistema eskailera baten antzekoa da, nerbio-korda bentralen bikoteekin segmentu guztietan, ganglio parea sortuz segmentu bakoitzean. Burua segmentu kopuru ezberdinen fusio bidez osatzen dira, eta euren garuna ganglio horietako batzuen batuketatik osatzen da, esofagoaren inguruan. Arnasketa-sistema eta iraizketa-sistemak aldakorrak dira artropodo ezberdinetan, batez ere euren ingurunearen eta osatzen dute subfilumaren araberakoa. Ikusmena begi-konposatu eta begi-sinpleen konbinazio batekin eman ohi da: espezie gehienetan ocelliek argia nondik datorren zehaztu dezakete, eta begi-konposatuak dira informazio iturririk nagusiena, baina armiarmen begi nagusiak ocelli sinpleak dira. Artropodoek sentsore kimiko eta mekaniko kopuru handia dute, gehienak euren septuen modifikazio bidez lortuak. Artropodoen ugalketa anitza da; espezie lurtar guztiek barne ernalketa dute, baina gehienetan lurrera botatzen diren espermatozoideak apendize batekin hartuz lortzen da, sarketa bidez izan beharrean.
Artropodoen arbaso ebolutiboa Kanbriar garaikoa da. Taldea monofiletikoa dela uste da, eta ikerketa askok proposatzen dute artropodoak eta Cycloneuralia (edo bere barneko kladoak) Ecdysozoa superfilumean sailkatu beharko liratekeela. Orokorrean, hala ere, Metazoaren oinarrizko harremanak zeintzuk diren ez da ondo ebaztu, momentuz. Artropodoen barne taldeen arteko harremanak ere eztabaidagarriak dira oraindik ere. Espezie urtarrek barne zein kanpo ernalketa erabiltzen dute. Artropodo gehienek arrautzak uzten dituzte, baina eskorpioek amaren barruan hazi diren kumeak erditzen dituzte. Artropodo kumeen egitura oso ezberdina izan daiteke, batzuk helduen bertsio txikiak dira, biana beste batzuek metaforfosi osoa izaten dute kume itxuratik heldu itxurara. Eskorpioak kumeengatik kezkatzen diren bitartean, beste espezie askotan ez dago inongo ardurarik hurrengo belaunaldiarekiko.
Artropodoak oso garrantzitsuak dira gizakien elikadurarako, zuzenean elikagai gisa edo jaten diren fruituen polinizazioa eginez. Espezie batzuek gaixotasunak hedatzen dituzte, gizaki, etxabere edo nekazaritza-produktuetan.
Artropodoen gorputz segmentatua dute eta segmentu bakoitzak adarrak ditu[3]. Bihotza alde dortsalean (goikaldean) dute eta nerbio-sistema bentralean (behekaldean). Artropodo guztiek exoeskeleto bat dute euren gorputza. Exoeskeleto hau kitinaz osatua dago, glukosaminazko polimero bat[4]. Artropodoek, noizbehinka, euren exoeskeletoa apurtu, atera eta handiagoa den beste bat eraikitzen dute. Sistema honek artropodoei lehortzea eragozten die, baina baita gehiegi hazten ere.
Artropodo espezie kopuruaren estimazioak oso aldakorrak dira; txikienak dio 1.170.000 espezie inguru direla, baina badira 5 edo 10 milioi direla dioten ikerketak ere, espezie bizidun guztien %80 inguru[5][6]. Espezie kopuru zehatza zein den kalkulu zaila da. Izan re, mundu mailan erabiltzen diren zentsu-sistemak ezberdinak dira, eta batzuetan leku zehatz batean egiten diren kontaketak mundu osorako baliagarritzat hartzen dira. 1992an egindako ikerketa batek zioen bakarrik Costa Rican 500.000 animalia eta landare espezie zeudela, horietatik 365.000 artropodoak[7].
Itsasoan, ur gezan, lehorrean eta aireko ekosistematan kide garrantzitsuak dira, eta ingurune lehorrera adaptatu diren bi animalia talde handietako bat da; bestea Amniota da, narrastiak, hegaztiak eta ugaztunak batzen dituen taldea[8]. Artropodoen barneko azpi-talde bakar bat, intsektuak, lehorreko eta ur-gezako ingurune guztietan espezie gehien dituena da. Intsekturik arinenak 25 mikrogramo pisatzen ditu[9], pisutsuenak 70 gramo dituen bitartean[10]. Krustazeo batzuk handiagoak dira; adibidez, Japoniako karramarro erraldoiak 4 metroko hankak izan ditzake, eta Homarus americanusak 20 kilogramoko pisua izan dezake.
Artropodo guztien enbrioiak segmentatuak dira, modulu errepikatuen segida batez eratuak. Gaur egun bizi diren artropodo guztien azken arabaso komunak, ziurrenik, diferentziatu gabeko segmentuz osatua zegoen, bakoitza adar gisa funtzionatzen zuen apendize pare batekin. Hala ere, ezagutzen ditugun artropodo bizidun eta fosil guztiek tagma izeneko segmentuetan taldekatuta dituzte segmentu horiek, eta adarrek espezializazioren bat dute[8].
Intsektu gehienen hiru ataletako itxura eta armiarmen bi partetako itxura taldekatzeko honen ondorioa da[11]; berez, ez dago kanpo segmentazio itxurarik akaroetan[8]. Artropodoek badituzte bi gorputz-elementu ez direna errepikatutako segmentuen parte, prostomioa gorputzaren aurrealdean, ahoaren aurretik, eta telsona atzean, ipurtzuloaren atzetik. Begiak prostomioaren gainean daude[8].
Artropodoek sistema zirkulatorio ireki bat daukate. Euren odola kuprean oinarrituta dago eta bere bulkada egiteko gorputzan zehar dauden hainbat bihotzek eragiten dute. Odola modu zuzenean dago kontaktuan ehun guztiekin. Artropodoak protostomak dira, hau da, gorputzaren barnekaldean espazioa dute, baina hau oso txikia da eta iraitz eta ugalketa aparatuei loturik dago. Era berean begiak dituzte kanpokaldean eta telson izeneko segmentu bat gorputzaren atzekaldean. Segmentu bakoitzak apendize pare bat ditu, baina batzuetan apendizetako bat galdu egin da eboluzioan zehar (unirramioak).
Gaur egun honako sailkapen hau proposatzen da artropodoentzat:
Phylum Arthropoda
Autore batzuek Myriapoda eta Hexapoda talde berdinean sartzen dute, Uniramia izeneko azpiphylumean, hain zuzen ere.
Oraindik dauden erlazioak oso argi ez badaude ere (autore batzuek unirramioak Onychophorarekin erlazionatzen dute), hainbat gauza nahiko ziurtzat ematen dira.
Tradizionalki anelidoak oso gertu dauden senide gisa hartu izan dira, biek dutelako segmentazioa gorputzean. Hala ere, gaur egun, eboluzio konbergentea dela uste da. Artropodoak pseudozelomatuak dira eta beraz hauekin ere erlazionaturik daudela uste da. (Ikusi Ecdyosozoa eta Articulata).
Sailkapena egiten dutenen arabera dauden 5 azpitaldeak (Trilobita, Chelicerata, Myriapoda, Hexapoda, eta Crustacea) batzuetan azpiphyla moduan sailkatzen dira eta beste batzuetan klase moduan, autoreraren irizpidearen arabera.
Burguess Shaleko Kanbriarreko artropodo askok ez dute gaur egungo taldeekin harremanik eta euren sailkapenerako hainbat autorek azpiphylum berriak eraikitzea proposatu dute.
Artropodoak (antzinako grezieraz: ἄρθρον, arthron, "artikulazio" eta πούς, podos, "hanka") exoeskeleto bat, gorputz segmentatua eta apendize bikoitiak dituzten animalia ornogabeak dira. Artropodoek Euarthropoda filuma osatzen dute, eta euren barruan intsektuak, araknidoak, miriapodoak eta krustazeoak daude, besteak beste. Hasiera batean Arthropoda terminoa Euartropodoak eta onikoforoak izendatzeko erabili zen. Artropodoen ezaugarririk nabarmenena artikulatutako adarrak eta kitinaz osatutako kutikulak dira, batzuetan kaltzio karbonatoan mineralizatzen direnak. Artropodoen gorputz-planak segmentuak ditu, horietako bakoitza apendize pare batekin. Kutikula zurrunak hazkuntza gelditzen du, beraz artropodoek mudak egiten dituzte erregularki. Simetria bilaterala dute, eta gorputzak kanpo eskeletoa du. Espezie batzuek hegalak dituzte.
Bere aldakortasunari esker, elkarketa-ekologiko guztietako espezierik ohikoenak bihurtu dira ingurune gehienetan. Gutxi gorabehera milioi bat espezie deskribatu dira, eta animalia espezie guztien %80 baino gehiago dira. Horietako batzuk, beste animalia gehienek ez bezala, giro lehorrean arrakasta handia dute. Artropodoak neurriz aldatzen dira Stygotantulus krustazeo mikroskopikotik Japoniako Macrocheira kaempferi karramarro erraldoiraino.
Artropodoen barne kabitate nagusia hemozeloa da, bere barne-organoak biltzen dituena; bertatik, odolaren analogoa den hemolinfak zirkulatzen du. Euren zirkulazio-sistema irekia da. Kanpoko egituran gertatzen den bezala, artropodoen barne organoak ere errepikapen bidez eratu ohi dira. Euren nerbio-sistema eskailera baten antzekoa da, nerbio-korda bentralen bikoteekin segmentu guztietan, ganglio parea sortuz segmentu bakoitzean. Burua segmentu kopuru ezberdinen fusio bidez osatzen dira, eta euren garuna ganglio horietako batzuen batuketatik osatzen da, esofagoaren inguruan. Arnasketa-sistema eta iraizketa-sistemak aldakorrak dira artropodo ezberdinetan, batez ere euren ingurunearen eta osatzen dute subfilumaren araberakoa. Ikusmena begi-konposatu eta begi-sinpleen konbinazio batekin eman ohi da: espezie gehienetan ocelliek argia nondik datorren zehaztu dezakete, eta begi-konposatuak dira informazio iturririk nagusiena, baina armiarmen begi nagusiak ocelli sinpleak dira. Artropodoek sentsore kimiko eta mekaniko kopuru handia dute, gehienak euren septuen modifikazio bidez lortuak. Artropodoen ugalketa anitza da; espezie lurtar guztiek barne ernalketa dute, baina gehienetan lurrera botatzen diren espermatozoideak apendize batekin hartuz lortzen da, sarketa bidez izan beharrean.
Artropodoen arbaso ebolutiboa Kanbriar garaikoa da. Taldea monofiletikoa dela uste da, eta ikerketa askok proposatzen dute artropodoak eta Cycloneuralia (edo bere barneko kladoak) Ecdysozoa superfilumean sailkatu beharko liratekeela. Orokorrean, hala ere, Metazoaren oinarrizko harremanak zeintzuk diren ez da ondo ebaztu, momentuz. Artropodoen barne taldeen arteko harremanak ere eztabaidagarriak dira oraindik ere. Espezie urtarrek barne zein kanpo ernalketa erabiltzen dute. Artropodo gehienek arrautzak uzten dituzte, baina eskorpioek amaren barruan hazi diren kumeak erditzen dituzte. Artropodo kumeen egitura oso ezberdina izan daiteke, batzuk helduen bertsio txikiak dira, biana beste batzuek metaforfosi osoa izaten dute kume itxuratik heldu itxurara. Eskorpioak kumeengatik kezkatzen diren bitartean, beste espezie askotan ez dago inongo ardurarik hurrengo belaunaldiarekiko.
Artropodoak oso garrantzitsuak dira gizakien elikadurarako, zuzenean elikagai gisa edo jaten diren fruituen polinizazioa eginez. Espezie batzuek gaixotasunak hedatzen dituzte, gizaki, etxabere edo nekazaritza-produktuetan.
Niveljalkaiset (Arthropoda) on ylivoimaisesti suurin eläinkunnan pääjakso. Niveljalkaisiin kuuluvia lajeja tunnetaan jo yli miljoona, mutta niitä arvellaan olevan vielä paljon enemmän. Arvioidaan, että kaikista nykyään elävistä eläinlajeista jopa 85 % on niveljalkaisia. Niveljalkaisilla on kuoriosa ja parilliset raajat. Niveljalkaisiin kuuluvat muun muassa trilobiitit, hämähäkkieläimet, äyriäiset, hyönteiset ja tuhatjalkaiset.[1]
Termi niveljalkainen eli latinaksi arthropod on johdettu Kreikan kielen sanasta kreik. ἄρθρον árthron, ”nivel”, ja kreik. πούς pous (genetiivi podos), ”jalka”. Sen esitteli 1845 saksalainen eläintieteilijä Karl Theodor Ernst von Siebold (1804-1885).[2]
Niveljalkaisille on ominaista kitiinipitoinen suojaava tukirunko, jonka sisäpintaan lihakset ovat kiinnittyneet. Kitiinirunko ei pysty laajenemaan, joten niveljalkaisten on siis aika ajoin ”luotava nahkansa”, jotta ne voivat kasvaa.[3] Ruumis koostuu jaokkeista, joiden välissä on nivelet. Jaokkeissa on yleensä jalkoja tai muita ulokkeita, joissa on myös nivel, joista koko pääjakso on saanut nimensä.[1] Ruumiin nivelten parillisista hermosolmukkeista ja niitä yhdistävistä pituussuuntaisista hermosyistä koostuu vatsapuolella sijaitseva hermorunko eli vatsaydin . Niveljalkaisten verenkierto on avoin. Putkimainen sydän ja siihen liittyvä verisuonisto sijaitsee selkäpuolella. Hengityselimistö on maalla yleensä ilmaputkisto, vedessä kidukset.[3]
Vallitsevana on suvullinen lisääntyminen, mutta suvutonta lisääntymistä tavataan myös.[1]
Niveljalkaiset jaetaan yleensä neljään alajaksoon: Kuusijalkaiset (Hexapoda), leukakoukulliset (Chelicerata), tuhatjalkaiset (Myriapoda) ja äyriäiset (Crustacea).[1]
Useimmat niveljalkaiset ovat pienikokoisia. Ne voivat kasvaa kohtalaisen kokoisiksi vain meressä, missä vesi antaa niiden tukirungolle lisätukea. Merten niveljalkaisista suurin on jopa 20 kiloa painava jättiläistaskurapu. Maalla elävät hyönteiset tai hämähäkit eivät paina aikuisina yli sataa grammaa, mutta yhden kovakuoriaislajin toukka on yli 200-grammainen.[1]
Niveljalkaisia tavataan lähes kaikkialla maapallolla, jopa 4000 metrin syvyydessä merissä ja yli 6700 metrin korkeudessa vuorilla.[1]
Niveljalkaiset (Arthropoda) on ylivoimaisesti suurin eläinkunnan pääjakso. Niveljalkaisiin kuuluvia lajeja tunnetaan jo yli miljoona, mutta niitä arvellaan olevan vielä paljon enemmän. Arvioidaan, että kaikista nykyään elävistä eläinlajeista jopa 85 % on niveljalkaisia. Niveljalkaisilla on kuoriosa ja parilliset raajat. Niveljalkaisiin kuuluvat muun muassa trilobiitit, hämähäkkieläimet, äyriäiset, hyönteiset ja tuhatjalkaiset.
Termi niveljalkainen eli latinaksi arthropod on johdettu Kreikan kielen sanasta kreik. ἄρθρον árthron, ”nivel”, ja kreik. πούς pous (genetiivi podos), ”jalka”. Sen esitteli 1845 saksalainen eläintieteilijä Karl Theodor Ernst von Siebold (1804-1885).
Niveljalkaisille on ominaista kitiinipitoinen suojaava tukirunko, jonka sisäpintaan lihakset ovat kiinnittyneet. Kitiinirunko ei pysty laajenemaan, joten niveljalkaisten on siis aika ajoin ”luotava nahkansa”, jotta ne voivat kasvaa. Ruumis koostuu jaokkeista, joiden välissä on nivelet. Jaokkeissa on yleensä jalkoja tai muita ulokkeita, joissa on myös nivel, joista koko pääjakso on saanut nimensä. Ruumiin nivelten parillisista hermosolmukkeista ja niitä yhdistävistä pituussuuntaisista hermosyistä koostuu vatsapuolella sijaitseva hermorunko eli vatsaydin . Niveljalkaisten verenkierto on avoin. Putkimainen sydän ja siihen liittyvä verisuonisto sijaitsee selkäpuolella. Hengityselimistö on maalla yleensä ilmaputkisto, vedessä kidukset.
Vallitsevana on suvullinen lisääntyminen, mutta suvutonta lisääntymistä tavataan myös.
Niveljalkaiset jaetaan yleensä neljään alajaksoon: Kuusijalkaiset (Hexapoda), leukakoukulliset (Chelicerata), tuhatjalkaiset (Myriapoda) ja äyriäiset (Crustacea).
Useimmat niveljalkaiset ovat pienikokoisia. Ne voivat kasvaa kohtalaisen kokoisiksi vain meressä, missä vesi antaa niiden tukirungolle lisätukea. Merten niveljalkaisista suurin on jopa 20 kiloa painava jättiläistaskurapu. Maalla elävät hyönteiset tai hämähäkit eivät paina aikuisina yli sataa grammaa, mutta yhden kovakuoriaislajin toukka on yli 200-grammainen.
Niveljalkaisia tavataan lähes kaikkialla maapallolla, jopa 4000 metrin syvyydessä merissä ja yli 6700 metrin korkeudessa vuorilla.
Arthropoda
Les Arthropodes (Arthropoda[1]) — du grec arthron « articulation » et podos « pied », aussi appelés « articulés » — sont un embranchement d'animaux protostomiens dont le plan d'organisation est caractérisé par un corps segmenté. Ils sont formés de métamères hétéronomes munis chacun d'une paire d'appendices articulés et recouvert d'une cuticule ou d'une carapace rigide, qui constitue leur exosquelette, dans la plupart des cas constitué de chitine. Leur mue permet, en changeant périodiquement leur squelette externe, de grandir en taille (mue de croissance) ou d'acquérir de nouveaux organes, voire de changer de forme (mue de métamorphose). Ils seraient apparus il y a 543 Ma.
L'embranchement des arthropodes est de très loin celui qui possède le plus d'espèces et le plus d'individus de tout le règne animal et des eucaryotes (80 % des espèces connues), tels les myriapodes, crustacés, arachnides, insectes, etc. On compte plus d'un million et demi d'espèces actuelles d'arthropodes qui présentent des modes de vie (guildes écologiques) les plus variés possibles grâce notamment à leur tagmatisation. Les membres de ce taxon sont aussi extrêmement nombreux : ils sont principalement représentés par les insectes dont 1019 (10 milliards de milliards) d'individus seraient vivants en même temps à un instant donné selon des estimations[2]. Les arthropodes forment un groupe cosmopolite qui s'est adapté dans des environnements naturels (déserts, forêts, abysses, montagnes, etc.) ou d’origine anthropique (habitations, puits de pétrole, etc.) et sont parmi les premiers animaux à avoir colonisé la terre ferme[3].
Leur nombre diminue toutefois considérablement. À l'échelle de trois régions allemandes, la biomasse d’arthropodes a ainsi chuté de 67 % au cours de la décennie 2010[4].
Les micro-arthropodes (arthropodes de la microfaune) sont les plus petits d'entre eux, de taille inférieure à 0,2 mm. Bien que discrets, ceux d'entre eux qui sont des décomposeurs jouent un rôle essentiel dans les réseaux trophiques en assurant le recyclage de la nécromasse, notamment dans le sol où avec les champignons décomposeurs, ils contribuent à produire l'humus.
Les arthropodes ou articulés sont :
Les articulations sont rendues nécessaires par la présence de chitine, matière coriace, à la surface de la peau. Les appendices articulés s'adaptent aux plus diverses fonctions.
La cavité générale est réduite à un ensemble de lacunes où circule l'hémolymphe.
Le système nerveux est ganglionnaire.
Au cours du développement se produisent des mues, et très souvent des métamorphoses.
La chitine (du grec chiton χιτών signifiant « tunique ») est une substance azotée sécrétée par l'ectoderme. D'abord mince et flexible, elle s'épaissit en une carapace résistante, qui revêt non seulement l'extérieur du corps, mais aussi les parties antérieures et postérieures du tube digestif.
On considère la chitine comme le caractère des arthropodes dominant auquel sont subordonnés les caractères suivants :
Conséquence de la présence d'une cuticule rigide et inextensible (l'exosquelette), les arthropodes sont pourvus d'appendices segmentaires articulés : chaque métamère (appelé aussi segment) est relié à ses voisins par des membranes articulaires et porte fondamentalement un nombre pair d'appendices articulés aux fonctions variées (antennes, pièces buccales, pattes locomotrices, nageuses ou marcheuses…). La tagmatisation induit la présence, l'absence ou la réduction de ces appendices. Ils ne sont pas souples comme les parapodes des annélides, mais composés de plusieurs articles durs (articles appelés aussi segments, à ne pas confondre avec les métamères), liés entre eux par des membranes articulaires flexibles qui permettent les mouvements[6].
Les appendices se composent typiquement d'une base, ou protopodite, formée de deux segments qui portent chacun deux segments latéraux.
Ces diverses parties ne restent simples que chez les crustacés inférieurs. Chez tous les autres arthropodes, elles se complexifient et s'adaptent à diverses fonctions. Il en résulte des appendices sensoriels (antennules, antennes), masticateurs (mandibules, maxillule, maxille, certains maxillipèdes (ou pattes-mâchoires), certains pédipalpes), préhenseurs (certains maxillipèdes (ou pattes-mâchoires), certains pédipalpes, pinces), locomoteurs (pattes à partir des 5 premières paires d'appendices, nageoires à partir des paires 11 jusqu'aux paires 16), reproducteurs (organes d'accouplement) à partir de la 11e paire d'appendices chez le mâle, etc.
Par contre, les ailes ne sont pas des appendices d'un point de vue morphologique puisqu'elles ne disposent pas de ces deux segments.
L'appareil circulatoire est un ensemble de lacunes. Il n'y a ni capillaires, ni veines. Les artères sont elles-mêmes réduites. Les vésicules cœlomiques métamérisées ont fusionné avec le blastocœle embryonnaire (par disparition des dissépiments, les parois entre deux segments) pour former l’hémocœle dans lequel circule l'hémolymphe, liquide circulatoire dont le rôle est analogue au sang et au liquide interstitiel chez les vertébrés[7].
Le cœur, situé dorsalement, est formé d'une ou de plusieurs poches en série linéaire et percées chacune d'une paire d'orifices ou ostioles. À chaque dilatation, le sang est aspiré à travers les ostioles. À chaque contraction, il est chassé dans les artères.
Le sang est incolore ou bleuté (hémocyanine), et contient seulement des globules blancs.
L'appareil excréteur est très variable selon les groupes. Il est formé de quelques glandes qui dérivent peut-être de néphridies.
Le système nerveux est ganglionnaire et ventral.
Il y a toujours une paire de ganglions cérébroïdes et une paire de ganglions sous-œsophagiens reliés par un collier périphagien. Le reste du système nerveux est en « échelle de corde » ou « corde à nœuds ».
Les ganglions de chaque paire sont unis par une commissure. Les paires successives sont unies par des connectifs.
Chez les types supérieurs, les ganglions tendent à fusionner en une ou plusieurs grosses masses.
Les yeux sont simples (ocelles) ou composés (yeux à facettes). Un œil composé peut être formé de plusieurs centaines d'ommatidies. Chaque ommatidie comprend :
Les ommatidies sont séparées les unes des autres par des cellules noires.
Il est probable que les yeux composés donnent à leur possesseur une vue panoramique (large champ visuel), mais très imprécise.
Les arthropodes se dirigent surtout par le toucher et par l'odorat, qui ont leur siège dans des poils sensoriels. Ceux-ci sont creux et contiennent le prolongement d'un neurone sensitif.
Il peut aussi y avoir des organes auditifs et des organes d'équilibre.
Pour se protéger, quelques espèces d'arthropodes utilisent le mimétisme, voire le Myrmécomorphisme.
Les sexes sont généralement séparés. Les œufs, assez chargés de vitellus (œufs centrolécithe), ont une segmentation partielle ou inégale.
À l'occasion des mues se produisent souvent, au cours du développement, des changements brusques de forme, autrement dit des métamorphoses.
Beaucoup d'arthropodes parasites, piqueurs ou suceurs sont vecteurs de maladies[8]. En matière de responsabilité au regard de l'importance épidémiologique mondiale pour l'homme, les moustiques sont considérés comme le premier groupe de vecteurs, le second groupe étant celui des arthropodes hématophages (ex. : acariens : tiques, insectes : puces), qui ont leur pendant chez les végétaux (ex. : puceron, punaises).
Nombre d'arthropodes vecteurs de maladies semblent héberger durablement des bactéries intracellulaires, ayant des effets variés sur leurs hôtes et transmises verticalement. Il s'agit d'interactions durables et peut-être parfois, voire souvent de véritables symbioses, certaines bactéries étant bénéfiques voire nécessaires, par exemple pour fournir à leur hôte des acides aminés essentiels, des vitamines ou un rôle fonctionnel vital (on parle alors de symbiote primaire et pour l'exemple cité de symbiose nutritionnelle). Les bactéries utiles à l'hôte, mais facultatives sont dites symbiotes secondaires pour leur hôte ; elles leur apportent par exemple un complément métabolique indispensable chez les espèces vivant sur des milieux pauvres (certains arthropodes hématophages) par exemple. Il est fréquent qu'un arthropode véhicule un symbiote primaire et plusieurs symbiotes secondaires, chez le puceron, le charançon, la mouche tsé-tsé par exemple, ou chez de nombreux aleurodes.
La bactérie symbiote peut fortement modifier le comportement de son hôte (et même le sexe-ratio dans certains cas). Il semble parfois exister une triple symbiose ou au moins une relation triangulaire ; des insectes phytophages étant par exemple vecteurs de virus qui infectent plus facilement leur plante hôte en présence de la bactérie symbiotique. Du point de vue épidémiologique global et des interactions « hôte-parasite », les arthropodes piqueurs-suceurs pourraient jouer un rôle important pour l'immunité de leurs hôtes et/ou de leurs populations, permettant une coévolution moins brutale des espèces avec la plupart de leurs parasites et pathogènes.
Sur la formule générale des vers, dont ils conservent d'ailleurs souvent l'aspect à l'état larvaire, les arthropodes ont superposé plusieurs innovations :
Cette formule gagnante correspond à la forme générale des mille-pattes. Elle a été immédiatement à l'origine d'une nouvelle explosion radiative, qui a exploré différentes formules pour transformer tel ou tel groupe de pattes en mâchoires, antennes, pattes spécialisées, ou les laisser régresser dans la queue. La variable qui semble avoir structuré sa répartition est le nombre de pattes.
Les trilobites ont été les animaux emblématiques du monde marin à l'ère paléozoïque. C'est un groupe à présent éteint.
Les trilobites sont bien connus car ils sont l'un des groupes fossiles les plus répandus. Par ailleurs, ils ont donné les fossiles les plus diversifiés : on recense entre neuf et quinze mille espèces. La plupart d'entre elles étaient des animaux marins simples et petits, qui filtraient la vase pour s'alimenter.
Les trilobites variaient en taille, d'un millimètre à plus de soixante-dix centimètres, avec une moyenne entre deux et sept centimètres. Le plus grand trilobite connu, Isotelus rex, mesurait 72 cm de long, et fut retrouvé en 1998 par des scientifiques canadiens sur les rivages de la baie d'Hudson.
Les zoologistes ont répertorié environ 55 000 espèces de crustacés.
Les plus connus sont les crabes, crevettes, homards et autres langoustes, tous sont des crustacés décapodes (à dix pattes). Le nombre de pattes est cependant très variable parmi les crustacés.
Le plus grand crustacé terrestre est le crabe de cocotier.
Le cloporte est également un crustacé, un isopode.
Pendant l'ère paléozoïque, au Cambrien, il y a de cela environ 540 millions d'années[réf. nécessaire], alors que la mer grouillait de trilobites et que les frontières des forêts primitives étaient peuplées de géants rampants (des mille-pattes de 1,50 m), apparurent les premiers arthropodes à pinces du sous-phylum des chélicérés, dont la plupart s'adaptèrent ensuite à la vie terrestre.
C'est à cette époque que l'on rencontre les premiers arachnides possédant des chélicères. Plus tard, dès le Silurien supérieur, nous découvrons des espèces de scorpions et d'araignées fort semblables à ceux et celles que nous pouvons trouver à l'heure actuelle.
On a recensé à ce jour environ 80 000 espèces d'arachnides, dont plus de 1 500 espèces de scorpions et 50 000 espèces d'araignées vivant dans tous les biotopes existants, des régions tropicales aux régions polaires. La plupart des arachnides sont cependant terrestres.
Les autres chélicérés sont très marginaux, les plus connus sont les tiques et les limules.
Les ptérygotes sont les porteurs d'une évolution majeure : les ailes. Elles leur permettent de conquérir les airs, et alliées à la simplification qu'apporte l'animation de seulement six pattes pour se déplacer, ont été à l'origine d'une troisième explosion radiative.
C'est par la superposition de ces trois explosions radiatives (vermiformes, arthropodes, insectes) que les insectes dominent le monde par leur variété : ils représentent aujourd'hui près de 80 % des espèces animales décrites.
L'embranchement fossile des lobopodes, rattaché aux onychophores, contient des animaux qui ressemblent à un ver mince au corps mou et possédant des paires de robustes appendices articulés et épineux, formant ainsi un exosquelette, défense originale contre les prédateurs. Ainsi Diania cactiformis, surnommé le « cactus ambulant », est une espèce lobopodienne qui en possède 10 paires. Daté de 520 millions d'années, les chercheurs suggèrent que cet organisme serait le plus proche parent connu des arthropodes modernes, même s'il n'est pas leur ancêtre commun[9].
Les études phylogénétiques indiquent que les radiodontes (en) sont le groupe le plus proche de tous les euarthropodes qui ont des appendices frontaux sur le deuxième segment de la tête (deutéropodes). Les relations entre les principaux groupes d'euarthropodes cambriens restant controversées, il était difficile d'aller plus loin dans l'évaluation des étapes évolutives entre radiodontes et deutéropodes. En 2020, un euarthropode datant du Cambrien inférieur, découvert dans le site fossilifère de Chengjiang (Chine) et dénommé Kylinxia zhangi, possède à la fois des caractéristiques des deutéropodes (un bouclier de tête fusionné, un tronc entièrement arthrodisé et des endopodites articulés) et des radiodontes (cinq yeux comme Opabinia, des appendices prédateurs frontaux de type radiodonte). Kylinxia est un taxon transitionnel entre Radiodonta et Deuteropoda[10].
La classification traditionnelle définissait comme arthropodes tous les métazoaires à squelette externe, ou cuticule, à corps en segments articulés-porteurs d'appendices eux-mêmes articulés, et une croissance par mues).
Cette classification les distinguait principalement en opposant les Chélicérés (Xiphosures, Arachnides…) aux mandibulés (ou antennates) ; avec parmi ces derniers, 3 classes :
La classification moderne intègre les progrès apportés par les analyses génétiques et biomoléculaires, qui ont montré que les Insectes (Hexapoda) sont un groupe-frère des Malacostracés, ce qui les réunit à l'ensemble des Crustacés en un gigantesque taxon qu'on a appelé Pancrustacea[11].
D'après World Register of Marine Species (9 mai 2016.)[12] :
embranchement Arthropoda
Arachnide (Argiope bruennichi)
Limule (Limulus polyphemus)
Pycnogonide (Ascorhynchus compactus)
Branchiopode (Triops longicaudatus)
Ichthyostracé (Argulus coregoni)
Malacostracé (Cancer pagurus)
Maxillopode (Lepas anatifera)
Ostracode
Rémipède (Speleonectes tanumekes)
Entognathe (Acerentomon sp.)
Insecte (Morpho didius)
Chilopode (Lithobius forficatus)
Diplopode (Narceus americanus)
Pauropode
Symphyle
Trilobite
Avec les vers Nématodes et quelques autres groupes, ils constituent les Ecdysozoaires, clade qui rend obsolète l'ancienne notion d'articulés (en), classification de Georges Cuvier en 1817 qui regroupait les arthropodes et les annélides[13].
Les arthropodes sont un groupe qui réunit à la fois des taxons vivants et fossiles.
Pour des raisons de difficultés de classification de la plupart de ces espèces fossiles, que certains paléontologues placent parmi les arthropodes et d'autres dans des phylums différents, les Arthropodes actuels sont réunis dans le taxon monophylétique des Euarthropodes, inclus dans le taxon des Arthropodes au sens général, lui-même placé dans le clade plus vaste des Panarthropodes qui réunit aussi les Tardigrades et les Onychophores.
Cladogramme des Panarthropodes parmi les Ecdysozoaires[14] incluant les Euthycarcinoidea[15]
ProtostomesSpiralia (annélides, mollusques, brachiopodes, etc.)
Nematoida (nématodes et apparentés)
Scalidophora (Priapulida, Kinorhyncha et Loricifera)
Les arthropodes sont en grande partie des décomposeurs ou de petits prédateurs dépendant de ces décomposeurs ou d'autres décomposeurs (champignons notamment). Ils jouent un rôle très important dans les forêts[16]. Au moins 79 % des arthropodes terrestres prédateurs consomment des proies relativement grandes en transformant leur cuticule épaisse par digestion extra-orale (exodigestion grâce à l'injection d'enzymes hydrolytiques)[17].
Ils sont vulnérables à certains polluants (pesticides notamment). Ils sont sensibles à l'humidité du sol[18], vulnérables aux longues inondations et très vulnérables aux sécheresses ; des stress hydriques importants sur les arbres sont associés à une régression de la communauté entière d’arthropodes vivant sous les arbres (ainsi d'ailleurs que de la communauté mycorhizienne)[16].
Par exemple, sous des pins (Pinus edulis) étudiés dans une zone présentant un gradient dans la gravité d'une sécheresse intense, l'intensité des symptômes en ce qui concerne le retard de croissance et le dépérissement était associée à la gravité du recul des communautés d'arthropodes. Les modèles tirés de l'observation confirment pour différentes espèces fondatrices de la forêt (ici du sud-ouest des États-Unis) que la sévérité des sécheresses correspond à une sévérité des impacts sur les communautés des arbres avec un effondrement de la richesse en espèces et de l'abondance des arthropodes (huit à dix fois moins d'arthropodes dans les lieux qui ont le plus souffert). Remarque : dans ce cas, l'épaisseur des cernes et l'état du feuillage correspondait à la qualité et quantité des populations d'arthropodes[16].
Les espèces ont répondu différemment au stress de la plante, mais la plupart ont été affectés négativement. Ces résultats laissent craindre que des sécheresses récurrentes affectant les espèces fondatrices d'arbres sont susceptibles de fortement diminuer la diversité multi-trophique et la composition des communautés d'arthropodes, avec de possibles effets en cascade sur d'autres taxons, la qualité et la résilience écologique des sols[16].
Arthropoda
Les Arthropodes (Arthropoda) — du grec arthron « articulation » et podos « pied », aussi appelés « articulés » — sont un embranchement d'animaux protostomiens dont le plan d'organisation est caractérisé par un corps segmenté. Ils sont formés de métamères hétéronomes munis chacun d'une paire d'appendices articulés et recouvert d'une cuticule ou d'une carapace rigide, qui constitue leur exosquelette, dans la plupart des cas constitué de chitine. Leur mue permet, en changeant périodiquement leur squelette externe, de grandir en taille (mue de croissance) ou d'acquérir de nouveaux organes, voire de changer de forme (mue de métamorphose). Ils seraient apparus il y a 543 Ma.
L'embranchement des arthropodes est de très loin celui qui possède le plus d'espèces et le plus d'individus de tout le règne animal et des eucaryotes (80 % des espèces connues), tels les myriapodes, crustacés, arachnides, insectes, etc. On compte plus d'un million et demi d'espèces actuelles d'arthropodes qui présentent des modes de vie (guildes écologiques) les plus variés possibles grâce notamment à leur tagmatisation. Les membres de ce taxon sont aussi extrêmement nombreux : ils sont principalement représentés par les insectes dont 1019 (10 milliards de milliards) d'individus seraient vivants en même temps à un instant donné selon des estimations. Les arthropodes forment un groupe cosmopolite qui s'est adapté dans des environnements naturels (déserts, forêts, abysses, montagnes, etc.) ou d’origine anthropique (habitations, puits de pétrole, etc.) et sont parmi les premiers animaux à avoir colonisé la terre ferme.
Leur nombre diminue toutefois considérablement. À l'échelle de trois régions allemandes, la biomasse d’arthropodes a ainsi chuté de 67 % au cours de la décennie 2010.
Les micro-arthropodes (arthropodes de la microfaune) sont les plus petits d'entre eux, de taille inférieure à 0,2 mm. Bien que discrets, ceux d'entre eux qui sont des décomposeurs jouent un rôle essentiel dans les réseaux trophiques en assurant le recyclage de la nécromasse, notamment dans le sol où avec les champignons décomposeurs, ils contribuent à produire l'humus.
Is fíleam ainmhithe iad na hartrapóid (Phylum Arthropoda, ón nGréigis ἄρθρον arthron, "alt", agus ποδός podos, "troigh"). Tá eiseachnámharlach, deighilt, agus géagán gabhal acu. I measc na n-ainmhithe san fhíleam seo, tá feithidí, crústaigh, araicnidí, srl.
O dos artrópodos (Arthropoda, do grego ἄρθρον, árthron, «articulación» e πούς, poús, «pé») é un filo de animais caracterizados por teren un exoesqueleto (esqueleto externo), o corpo segmentado e provisto de apéndices articulados.
Constitúen o filo máis numeroso e diversificado do reino animal, e inclúe, entre outros, aos insectos, arácnidos, crustáceos e miriápodos. Coñécense case 1 200 000 especies descritas, a súa maioría insectos (un millón),[1][2] que representan polo menos o 80 % de todas as especies animais coñecidas.
Varios grupos de artrópodos están perfectamente adaptados á vida no aire, igual que os vertebrados amniotas, e a diferenza de todos os demais filos de animais, que son acuáticos ou requiren ambientes húmidos.
A súa anatomía, a súa fisioloxía e o seu comportamento revelan un deseño simple pero admirabelmente eficaz. Os artrópodos existen en todos os ambientes da terra: no mar, na auga doce, no medio terrestre e no aire. Existen tamén moitas formas parasitas e simbióticas.
Pode ser que os primeiros artrópodos foran similares aos seus presuntos antecesores, os anélidos. O seu corpo era longo e brando, e tiña moitos segmentos ou metámeros, todos eles moi similares entre si e equipados cun par de patas. Posteriormente a superficie do corpo endureceuse até formar un esqueleto externo ou cutícula constituído por quitina, proteínas, lípidos e sales de calcio.
Hai rexistos fósiles de artrópodos desde o período cámbrico.
Pénsase que as formas Parvacornia e Spriggina da fauna de Ediacara, de hai uns 555 millóns de anos, eran artrópodos.[3][4][5]
Pequenos artrópodos con cunchas semellantes ás dos bivalvos encontráronse en xacementos de fósiles do cámbrico temperán, de 541 a 539 millóns de anos de antigüidade, en China.[6][7]
Os primeiros fósiles de trilobites cámbricos datan duns 530 millóns de anos, pero a clase xa estaba bastante diversificada en todo o mundo, o que suxire que xa existira durante algún tempo antes.[8] A revisión nos anos 1970 dos fósiles dos xistos de Burgess de hai cerca de 505 millóns de anos permitiron identificar numeosos artópodos, algúns dos cales non se poderían asignar a ningún dos grupos coñecidos, o que intensificou o debate acerca da explosión cámbrica.[9]
Os fósiles máis antigos de crustáceos datan de hai aproximadamente 513 millóns de anos, no cámbrico.[10] O arácnido máis antigo coñecido é Palaeotarbus jerami, de hai sobre uns 420 millóns de anos, no silúrico.[11] E o máis antigo insecto fósil é Rhyniognatha hirsti, do Devoniano, datado entre 396 e 407 millóns de anos, pero as súas mandíbulas son dun tipo que se encontra só en insectos alados, o que suxire que os primeiros insectos apareceron no período silúrico.[12]
Os artrópodos constitúen unha das grandes divisións do reino animal, subdividida á súa vez en diversas clases, algunhas das cales contan con gran número de xéneros e especies. Denomínanse así por estaren provistos de patas articuladas. Pero en realidade non son sá as patas, senón todo o corpo o que está formado por varios segmentos unidos entre sí por medio de articulacións, así como os sertmentos dos apéndices.
A pesar da súa variedade e disparidade, os artrópodos posúen en común características morfolóxicas e fisiolóxicas comúns fundamentais:
Os artrópodos teñen o corpo segmentado (2), envolvido nun exosqueleto quitinoso, con varios apéndices articulados (1). Os apéndices están especializados para a alimentación, para a percepción sensorial, para defensa e para a locomoción. Son estas "patas articuladas" as que dan o nome ao filo e os que o separan dos filos máis próximos, os onicóforos e os tardígrados.
Os segmentos poden estaren agrupados en rexións do corpo, denominadas tagmas, tipicamente:
Porén, en varios grupos, como os cangrexos, arañas, escorpións e lagostas, a cabeza e o tórax están unidos, formando o chamado cefalotórax protexido por unha peza única da casca, o cacho. Nos quelicerados, o corpo está dividido en prosoma e opistosoma.
O primeiro segmento da cabeza é denominado acron, e normalmente leva os ollos, que poden ser simples ou compostos. O último segmento do abdome (3) remata no telson. Cada segmento contén, polo menos primitivamente, un par de apéndices.
Os artrópodos respiran por un sistema de traqueas, túbulos que abren ao exterior a través de poros ou pequenos orificios na cutícula denominados espiráculos, que se estenden por todo o corpo facilitando o intercambio de gases. Os artrópodos acuáticos teñen branquias ligadas ao sistema de traqueas.
O sistema circulatorio dos artrópodos consiste nun conxunto de corazóns que se dispoñen ao longo do corpo e que bombean a hemolinfa (o "sangue" destes animais, que moitas veces non contén hemoglobina, que contén ferro, senón hemocianina, que contén cobre), e que se acha bañando os tecidos.
Os artrópodos son protóstomos, e posúen un celoma reducido a un espazo ao redor dos órganos da reprodución e da excreción.
Uns poucos artrópodos, como os cirrípedes, son hermafroditas, isto é, cada individuo pode ter os órganos de ambos sexos. Porén, os individuos da maioría das especies permanecen nun único sexo toda a súa vida.[16] Unhas poucas especies de insectos e crustáceos poden reproducirse por partenoxénese, especialmente se as condicións favorecen unha "explosión demográfica". Así a todo, moitos artrópodos confían na reprodución sexual, e as especies partenoxenética reverten á reprodución sexual cando as condicións se tornan menos favorables.[17] Os artrópodos acuáticos poden reproducirse mediante fecundación externa como, por exemplo, fan as ras; ou por fecundación interna, na que o óvulo permanece no corpo da femia e o esperma debe ser introducido dalgunha maneira. Todos os artrópodos terrestres coñecidos usan a fecundación interna. Os opilións, os diplópodos, e algúns crustáceos usan apéndices modificados como gonópodos ou penes para transferir o esperma directamente á femia. Non obstante, moitos machos de artrópodos terrestres producen espermatóforos, bolsas de esperma impermeables, que as femias toman dentro dos seus corpos. Unhas poucas especies confían nas femias para atopar espermatóforos que previamente foron depositados no chan, pero en moitos casos os machos só depositan espermatóforos cando parecen ter éxito rituais complexos de cortexo.[16]
A maioría dos artrópodos poñen ovos,[16] mais os escorpións son ovovíparos: nacen as crías despois de que os ovos foron incubados dentro da nai, e son obxecto dun prolongado coidado materno[18] Os artrópodos que acaban de nacer teñen diversas formas, só os insectos abranguen un amplo rango de formas extremas. Algunha crías que acaban de saír da casca do ovo aparentan adultos en miniatura (desenvolvemento directo), e noutros casos, coma no peixiño de prata, as crías non se alimentan e poden estar desamparados ata despois da súa primeira muda. Moitas crías de insecto, coma as larvas dos coleópteros ou as eirugas, que non teñen membros segmentados ou cutículas endurecidas, metamorfosean en formas adultas entrando nunha fase inactiva na que rompen os tecidos larvarios e se reutilizan para construír o corpo adulto.[19] A larva da libélula ten as cutículas típicas e os membros articulados dos artrópodos pero teñen branquias que lles permiten a respiración acuática e mandíbulas extensibles.[20] As crías dos crustáceos nacen comunmente como as delgadas larvas dos peneidos con só tres segmentos e pares de apéndices.[16]
Durante moitas décadas, as relacións filoxenéticas dos celomados baseáronse na concepción dos articulados de Cuvier,[21] un clado formado por anélidos e artrópodos. Numerosas análises morfolóxicas modernas baseados en principios cladísticos corroboraron a existencia do clado Articulata, por exemplo, Brusca & Brusca,[2] Nielsen[14] ou Nielsen et al.,[15] entre outros.
Porén diversas análises cladísticas, como a de datos combinados de Zrzavý et al.[13] chegaron á conclusión de que anélidos e artrópodos non están directamente relacionados. A presenza de metamerización en anélidos e artrópodos debería considerarse, pois, como unha converxencia evolutiva. Polo contrario, estes estudos propoñen o clado Ecdysozoa no que os artrópodos mostran estreitas relacións filoxenéticas con grupos de pseudocelomados, como nematodos, nematomorfos, priapúlidos e quinorrincos, pola presenza compartida dunha cutícula quitinosa e un proceso de muda (écdise) da mesma.
A filoxenia dos artrópodos foi moi controvertida, cunha enfrontada polémica entre os partidarios do monofiletismo e os do polifiletismo. Snodgrass [22] e Cisne [23] defenderon o monofiletismo, aínda que o primeiro considera os artrópodos divididos en Arachnata + Mandibulata, e o segundo interpreta que se dividirían en esquizorrámeos e atelocerados. Tiegs e Manton [24] defenderon o difiletismo, cos artrópodos divididos en esquizorrámeos + unirrámeos e os onicóforos como grupo irmán de miriápodos + hexápodos.
Posteriormente, Manton [25] e Anderson [26] sostiveron o polifiletismo do grupo.
Coa aparición dos primeiros estudos baseados en datos moleculares e análises combinadas de datos morfolóxicos e moleculares, parece que a antiga polémica sobre monofilia e polifilia quedou superada, xa que todos eles corroboran que os artrópodos son un grupo monofilético en que se inclúen tamén os tardígrados (o clado deuse en chamar panartrópodos); a maioría tamén propoñen a existencia do clado mandibulados. Porén, xurdiron novas controversias, sobre todo ao redor de dúas hipóteses alternativas mutuamente excluíntes que están sendo debatidas en numerosos artigos sobre filoxenia e evolución de artrópodos: atelocerados (miriápodos + hexápodos) (Wheeler)[27]
Cladograma A comprado co cladograma B [pancrustáceos (crustáceos + hexápodos)]. (Segundo Giribert & Ribera)[28]O filo dos artrópodos foi descrito en 1829 polo zoólogo francés Pierre André Latreille, na súa obra Les crustacés, les arachnides et les insectes: distribués en familles naturelles.
O termo do latín científico Arthropoda está construído cos elementos arthro- e -poda, tirados do grego antigo ἄρθρον, árthron, "articulación" e πούς, ποδός poús, podós, "pé", "perna", "pata", coa desinencia -a, indicando substantivo neutro plural. Literalmente, "os que teñen as patas articuladas".
As clasificacións tradicionais definían como artrópodos a todos os metazoos con esqueleto externo, ou cutícula, co corpo dividido en segmentos articulados portadores de apéndices así mesmo articulados, e que medraban por mudas). Estas clasificacións distinguían principalmente dous grupos, os os quelicerados (xifosuros, arácnidos...) e os mandibulados (ou antenados), estros últimos divididos en tres clases:
A clasificación máis moderna, tendo en conta os avances da xenética e as análises da bioloxía molecular, demostrou que os hexápodos (que comprende os insectos) é un grupo irmán dos crustáceos malacostráceos, polo que inclúe a todos os crustáceos e aos hexápodos nun taxon xigantesco chamado Pancrustacea.[29]
Porén, na bibliografía usualmente manexada séguese esta clasificación, seguindo ao WoRMS:[30]
Filo Arthropoda
Trilobitomorfo †
(un trilobite)
Quelicerado
(unha araña)
O dos artrópodos (Arthropoda, do grego ἄρθρον, árthron, «articulación» e πούς, poús, «pé») é un filo de animais caracterizados por teren un exoesqueleto (esqueleto externo), o corpo segmentado e provisto de apéndices articulados.
Constitúen o filo máis numeroso e diversificado do reino animal, e inclúe, entre outros, aos insectos, arácnidos, crustáceos e miriápodos. Coñécense case 1 200 000 especies descritas, a súa maioría insectos (un millón), que representan polo menos o 80 % de todas as especies animais coñecidas.
Varios grupos de artrópodos están perfectamente adaptados á vida no aire, igual que os vertebrados amniotas, e a diferenza de todos os demais filos de animais, que son acuáticos ou requiren ambientes húmidos.
A súa anatomía, a súa fisioloxía e o seu comportamento revelan un deseño simple pero admirabelmente eficaz. Os artrópodos existen en todos os ambientes da terra: no mar, na auga doce, no medio terrestre e no aire. Existen tamén moitas formas parasitas e simbióticas.
Člankonošci; Arthropoda, predstavljaju najbrojniju i najrasprostranjeniju skupinu beskralježnjaka. Više od tri četvrtine današnjeg životinjskog svijeta pripada člankonošcima. Čine ih tri osnovne skupine: trorežnjaci (Trilobita), kliještari (Chelicerata) i čeljusnici (Mandibulata). [1] U njih se ubrajaju rakovi, pauci, kukci i stonoge. Ime su dobili po člankovitim nogama.
Tijelo člankonožaca je kolutićavo, bilateralno simetrično. Kolutići su međusobno spojeni u dvije ili tri funkcionalne cjeline. Pokriveno je hitinskom pokrovom. Pokrov sprječava kontinuirani rast člankonožaca pa se oni moraju presvlačiti. Jednostavniji člankonošci se presvlače mnogo puta npr. raci, a kod složenijih člankonožaca npr. kukaca, broj presvlačenja je ograničen i događa se samo u stanju ličinke. Tri su glavna dijela tijela: glava, prsa i zadak. Glava i prsa srasli su u glavopršnjak. Zadak je člankovite građe. člankovite noge su parni izdanci na svakom kolutiću njihova tijela. Pauci imaju četiri para nogu koje su prekrivene dlakama (s pomoću njih osjeti kada je u blizini plijen).
Sustav za disanje ovisi o tome žive li člankonošci u vodi ili u zraku. Vodeni člankonošci dišu škrgama a zračni člankonošci dišu većinom uzdušnicama, trahejama.
Optjecajni sustav je jednostavan, srce se nalazi s leđne strane iznad crijeva u prostoru odijeljenom od ostale tjelesne šupljine.
Osjetila za opip, miris, sluh, ravnotežu i vid ovise o načinu života. Najjednostavniji mehanoreceptori su pokretne osjetne dlačice koje reagiraju na podražaje dodira, kretanja zraka, izmjena tlaka i sl. Kemoreceptori (osjetila mirisa) su najrazvijeniji kod kopnenih kukaca. Fotoreceptori (oči) člankonožaca čine retinularne stanice. Sastavljene oči (kukaca i rakova) sastoje se od većeg broja produljenih jednostavnih očiju, npr. oči obada zauzimaju čitavu leđnu i bočnu stranu glave. Neki kukci mogu proizvoditi glasove posebnim zvučnim, stridulacijskim, organima, npr. skakavci trenjem hrapavih dijelova krila i nogu ili cvrčci titranjem bubnjića.
Podijeljeni su na 16 razreda i poseban red Symphyla u koji su uključeni rod Remysymphyla i porodice Scolopendrellidae i Scutigerellidae[2]
Člankonošci; Arthropoda, predstavljaju najbrojniju i najrasprostranjeniju skupinu beskralježnjaka. Više od tri četvrtine današnjeg životinjskog svijeta pripada člankonošcima. Čine ih tri osnovne skupine: trorežnjaci (Trilobita), kliještari (Chelicerata) i čeljusnici (Mandibulata). U njih se ubrajaju rakovi, pauci, kukci i stonoge. Ime su dobili po člankovitim nogama.
Artropoda adalah filum yang paling besar dalam dunia hewan dan mencakup serangga, laba-laba, udang, lipan, dan hewan sejenis lainnya. Artropoda biasa ditemukan di laut, air tawar, darat, dan lingkungan udara, termasuk berbagai bentuk simbiosis dan parasit. Kata artropoda berasal dari bahasa Yunani ἄρθρον árthron, "ruas, buku, atau segmen", dan πούς pous (podos), "kaki", yang jika disatukan berarti "kaki berbuku-buku"[2]. Artropoda juga dikenal dengan nama hewan berbuku-buku atau hewan beruas.
Karakteristik yang membedakan artropoda dengan filum yang lain yaitu: tubuh bersegmen, segmen biasanya bersatu menjadi dua atau tiga daerah yang jelas, anggota tubuh bersegmen berpasangan (asal penamaan Artropoda), simetribu bilateral, eksoskeleton berkitin. Secara berkala mengalir dan diperbaharui sebagai pertumbuhan hewan, kanal alimentari seperti pipa dengan mulut dan anus, sistem sirkulasi terbuka, hanya pembuluh darah yang biasanya berwujud sebuah struktur dorsal seperti pipa menuju kanal alimentar dengan bukaan lateral di daerah abdomen, rongga tubuh; sebuah rongga darah atau hemosol dan selom tereduksi.
Sistem saraf mereka seperti tangga tali, terdiri atas sebuah ganglion anterior atau otak yang berlokasi di atas kanal alimentari, sepasang penghubung yang menyalurkan dari otak ke sekitar kanal alimentari dan tali saraf ganglion yang berlokasi di bawah kanal alimentary, ekskresi biasanya oleh tubulus malphigi. Tabung kosong yang masuk kanal alimentari dan material hasil ekskresi melintas keluar lewat anus, respirasi dengan insang atau trakhea dan spirakel, tidak ada silia atau nefridia.
Penglihatan mereka bergantung pada berbagai variasi kombinasi mata majemuk dan ocelli (mata tunggal) yang berbentuk mangkuk pigmen: pada kebanyakan spesies ocelli hanya dapat mendeteksi arah dari sumber cahaya, dan mata majemuk adalah sumber informasi utama, tetapi mata utama pada laba-laba adalah ocelli yang dapat membentuk bayangan dan dalam beberapa kasus, dapat berputar untuk melacak mangsa. Arthopoda juga memiliki berbagai sensor kimia dan mekanik, sebagian besar didasarkan pada modifikasi dari banyak setae (bulu) yang keluar melalui kutikula mereka. Metode repoduksi artropoda beragam; semua spesies darat menggunakan pembuahan dalam, tetapi sering dilakukan dengan cara pemindahan sperma tidak langsung. Spesies air menggunakan pembuahan dalam atau pembuahan luar. Hampir semua artropoda bertelur, tetapi kalajengking melahirkan setelah telurnya menetas di dalam tubuh induknya. Atropoda muda bervariasi bentuknya, mulai dari menyerupai dewasa dengan ukuran lebih kecil hingga berupa larva dan ulat yang tidak memiliki kaki berbuku-buku yang akhirnya akan mengalami metamorfosis total untuk mencapai bentuk dewasa. Penjagaan induk terhadap artropoda muda bervariasi dari yang tidak ada sama sekali hingga penjagaan yang lama, seperti yang dilakukan oleh kalajengking.
Garis evolusi artropoda dapat ditelusuri dari periode Kambrium. Kelompok ini umumnya dianggap sebagai monofiletik dan banyak analisis mendukung penempatan artropoda bersama dengan cycloneuralia dalam superfilum Ecdysozoa. Tetapi, hubungan dasar Metazoa belum diketahui secara pasti. Begitu pula hubungan di antara sesama kelompok artropoda masih sering diperdebatkan.
Artropoda memiliki kontribusi terhadap suplai makanan manusia, baik langsung (sebagai makanan) maupun tidak langsung, seperti membantu penyerbukan. Beberapa spesies tertentu diketahui merupakan vektor penyakit berbahaya bagi manusia, hewan, dan tumbuhan.
Artropoda adalah avertebrata dengan tubuh bersegmen dan kaki berbuku-buku.[3] Anggota-anggota tubuh membentuk suatu eksooskeleton, yang utamanya terdiri atas kitin α, salah satu turunan dari glukosa.[4] Kutikula krustasea juga mengalami biomineralisasi dengan kalsium karbonat.
Suatu kelompok hewan lain, tetrapoda, memiliki anggota tubuh berbuku-buku, tetapi tetrapoda adalah vertebrata dan karenanya memiliki endoskeleton.[5]
Estimasi jumlah spesies artropoda bervariasi antara 1.170.000 dan 5 hingga 10 juta serta meliputi lebih dari 80% seluruh spesies hewan yang diketahui masih hidup saat ini.[6][7] Jumlah spesiesnya masih sulit ditentukan karena penghitungannya berdasarkan model asumsi yang diproyeksikan ke wilayah-wilayah lain dari dari penghitungan di lokasi-lokasi tertentu diterapkan dengan skala ke seluruh dunia. Sebuah studi tahun 1992 mengestimasikan bahwa ada 500.000 spesies hewan dan tumbuhan hanya di Kosta Rika, dan 365.000 di antaranya adalah artropoda.[8]
Mereka adalah anggota penting dari ekosistem laut, air tawar, daratan, dan udara, dan merupakan salah satu dari hanya dua kelompok hewan besar yang telah beradaptasi untuk hidup dalam lingkungan kering. Kelompok lainnya adalah amniota yang anggota-anggotanya adalah reptil, burung, dan mamalia.[9] Salah satu anggota kelompok artropoda, serangga, adalah kelompok yang paling banyak spesies di antara semua kelompok ekologis di lingkungan darat dan air tawar.[8] Spesies paling ringan memiliki berat kurang dari 25 mikrogram (satu per satu juta gram),[10] sementara yang paling berat memiliki berat lebih dari 70 gram (2,5 oz).[11] Beberapa krustasea yang hidup saat ini berukuran jauh lebih besar, sebagai contoh, kaki kepiting laba-laba jepang dapat mencapai 4 meter (13 ft),[10] dan artropoda yang paling berat saat ini adalah lobster amerika dengan berat lebih dari 20 kg (44 lbs).
Embrio semua artropoda bersegmen, terdiri atas serangkaian modul yang berulang. Nenek moyang terakhir dari artropoda mungkin memiliki tubuh yang terdiri atas serangkaian segmen yang belum terdiferensiasi, masing-masing dengan sepasang embelan yang berfungsi sebagai anggota tubuh. Tetapi, semua artropoda, baik yang masih hidup maupun yang telah punah, memiliki segmen-segmen yang terkelompok-kelompok menjadi tagmata tempat segmen dan anggota tubuhnya terspesialisasi dalam berbagai cara.[9] Hasil pengelompokan itu adalah banyak tubuh serangga yang terlihat terdiri atas tiga bagian dan tubuh laba-laba terdiri atas dua bagian.[12] Faktanya tidak ada tanda-tanda segmentasi pada tubuh tungau.[9] Artropoda juga memiliki dua elemen tubuh yang bukan merupakan bagian dari rangkaian segmen berulang ini, sebuah akron di depan mulut dan sebuah telson di belakang anus. Mata menempel pada akron.[9]
Artropoda umumnya diklasifikasikan dalam lima upafilum, yang salah satunya telah punah:[13]
Artropoda dalam dunia hewan merupakan filum yang terbesar di dunia. Empat dari lima bagian spesies hewan adalah arthropoda, dengan jumlah di atas satu juta spesies modern yang ditemukan dan rekor fosil yang mencapai awal Cambrian. Jumlah spesiesnya yaitu sekitar 900.000 spesies dengan beragam variasi. Jumlah ini kira-kira 80% dari spesies hewan yang diketahui sekarang. Artropoda dapat hidup di air tawar, laut, tanah, dan praktis semua permukaan bumi dipenuhi oleh spesies ini. Artropoda dianggap berkerabat dekat dengan Annelida, contohnya adalah Peripetus di Afrika Selatan.
Artropoda mungkin satu-satunya yang dapat hidup di Antartika dan liang-liang batu terjal di pegunungan yang tinggi. Semua anggota filum ini mempunyai tubuh beruas-ruas dan kerangka luar yang tersusun dari kitin. Rongga tubuh utama disebut hemocoel. Hemocoel terdiri dari sejumlah ruangan kecil yang dipompa oleh jantung. Jantung terletak pada sisi dorsal dari tubuhnya.
|date= (bantuan) |date= (bantuan) |accessdate= (bantuan)Pemeliharaan CS1: Bahasa yang tidak diketahui (link) |accessdate=, |date= (bantuan) |accessdate= (bantuan) Artropoda adalah filum yang paling besar dalam dunia hewan dan mencakup serangga, laba-laba, udang, lipan, dan hewan sejenis lainnya. Artropoda biasa ditemukan di laut, air tawar, darat, dan lingkungan udara, termasuk berbagai bentuk simbiosis dan parasit. Kata artropoda berasal dari bahasa Yunani ἄρθρον árthron, "ruas, buku, atau segmen", dan πούς pous (podos), "kaki", yang jika disatukan berarti "kaki berbuku-buku". Artropoda juga dikenal dengan nama hewan berbuku-buku atau hewan beruas.
Karakteristik yang membedakan artropoda dengan filum yang lain yaitu: tubuh bersegmen, segmen biasanya bersatu menjadi dua atau tiga daerah yang jelas, anggota tubuh bersegmen berpasangan (asal penamaan Artropoda), simetribu bilateral, eksoskeleton berkitin. Secara berkala mengalir dan diperbaharui sebagai pertumbuhan hewan, kanal alimentari seperti pipa dengan mulut dan anus, sistem sirkulasi terbuka, hanya pembuluh darah yang biasanya berwujud sebuah struktur dorsal seperti pipa menuju kanal alimentar dengan bukaan lateral di daerah abdomen, rongga tubuh; sebuah rongga darah atau hemosol dan selom tereduksi.
Sistem saraf mereka seperti tangga tali, terdiri atas sebuah ganglion anterior atau otak yang berlokasi di atas kanal alimentari, sepasang penghubung yang menyalurkan dari otak ke sekitar kanal alimentari dan tali saraf ganglion yang berlokasi di bawah kanal alimentary, ekskresi biasanya oleh tubulus malphigi. Tabung kosong yang masuk kanal alimentari dan material hasil ekskresi melintas keluar lewat anus, respirasi dengan insang atau trakhea dan spirakel, tidak ada silia atau nefridia.
Penglihatan mereka bergantung pada berbagai variasi kombinasi mata majemuk dan ocelli (mata tunggal) yang berbentuk mangkuk pigmen: pada kebanyakan spesies ocelli hanya dapat mendeteksi arah dari sumber cahaya, dan mata majemuk adalah sumber informasi utama, tetapi mata utama pada laba-laba adalah ocelli yang dapat membentuk bayangan dan dalam beberapa kasus, dapat berputar untuk melacak mangsa. Arthopoda juga memiliki berbagai sensor kimia dan mekanik, sebagian besar didasarkan pada modifikasi dari banyak setae (bulu) yang keluar melalui kutikula mereka. Metode repoduksi artropoda beragam; semua spesies darat menggunakan pembuahan dalam, tetapi sering dilakukan dengan cara pemindahan sperma tidak langsung. Spesies air menggunakan pembuahan dalam atau pembuahan luar. Hampir semua artropoda bertelur, tetapi kalajengking melahirkan setelah telurnya menetas di dalam tubuh induknya. Atropoda muda bervariasi bentuknya, mulai dari menyerupai dewasa dengan ukuran lebih kecil hingga berupa larva dan ulat yang tidak memiliki kaki berbuku-buku yang akhirnya akan mengalami metamorfosis total untuk mencapai bentuk dewasa. Penjagaan induk terhadap artropoda muda bervariasi dari yang tidak ada sama sekali hingga penjagaan yang lama, seperti yang dilakukan oleh kalajengking.
Garis evolusi artropoda dapat ditelusuri dari periode Kambrium. Kelompok ini umumnya dianggap sebagai monofiletik dan banyak analisis mendukung penempatan artropoda bersama dengan cycloneuralia dalam superfilum Ecdysozoa. Tetapi, hubungan dasar Metazoa belum diketahui secara pasti. Begitu pula hubungan di antara sesama kelompok artropoda masih sering diperdebatkan.
Artropoda memiliki kontribusi terhadap suplai makanan manusia, baik langsung (sebagai makanan) maupun tidak langsung, seperti membantu penyerbukan. Beberapa spesies tertentu diketahui merupakan vektor penyakit berbahaya bagi manusia, hewan, dan tumbuhan.
Liðdýr (fræðiheiti: Arthropoda) eru stærsta fylking dýra. Til liðdýra teljast meðal annars skordýr, krabbadýr, áttfætlur og svipuð dýr sem einkennast af því að vera með liðskiptan líkama og ytri stoðgrind úr kítíni.
Gli artropodi (Arthropoda Latreille, 1829) sono un phylum (o tipo) di animali invertebrati protostomi celomati, che comprende circa i 5/6 delle specie finora classificate.
Il fatto che siano state descritte oltre un milione di specie di artropodi (e si stima che ne esistano 5 o forse 10 milioni[1]) dimostra come la loro struttura di base sia versatile e adattabile a diversi modi di vita.
Il termine "artropodi" viene dai termini greci ἄρθρον (àrthron), giunto, articolazione, e ποδόι (podòi), piedi, nel senso di "piedi articolati".
Alcune caratteristiche che distinguono gli artropodi.
Tutti gli artropodi si riproducono per mezzo di uova, anche se alcuni, in particolare gli scorpioni, sono ovovivipari, cioè le uova restano all'interno del corpo materno fino alla schiusa.
Le larve in alcuni casi assomigliano all'adulto, ma nella maggior parte dei casi presentano differenze significative.
La crescita degli artropodi è ostacolata dalle caratteristiche dell'esoscheletro, che non è in grado di espandersi in funzione con l'aumento di grandezza e larghezza del corpo. Pertanto tutti gli artropodi subiscono periodicamente delle mute con le quali l'esoscheletro esistente viene sostituito da uno nuovo.
Alle mute si accompagnano, in moltissimi artropodi, trasformazioni del corpo che non si limitano alla semplice crescita dimensionale (metamorfosi).
Nei crostacei è comune la comparsa di segmenti e piedi aggiuntivi, che corrispondono tipicamente alle tre fasi dette nauplio, zoea e adulto (con varianti a seconda degli ordini e delle specie).
Negli insetti la metamorfosi comporta trasformazioni varie, tra cui quasi sempre la comparsa delle ali, sempre assenti nelle larve. Negli insetti olometaboli la metamorfosi assume il suo grado massimo, con la presenza di tre fasi: la larva propriamente detta (fase che comprende vari stadi separati da mute); la pupa, fase immobile (preceduta da un eventuale stadio di pre-pupa); infine la cosiddetta immagine, che emerge dalla pupa e ha le caratteristiche dell'adulto.
Gli artropodi si sono originati in ambienti marini e ancora oggi un grandissimo numero di artropodi abita i mari e gli oceani. Molti gruppi si sono adattati anche alle acque dolci.
Alcuni gruppi di artropodi (aracnidi, insetti, ecc.) hanno avuto peraltro un grande successo nel colonizzare gli ambienti terrestri, persino se aridi. Gli insetti hanno infine colonizzato anche gli ambienti aerei, mediante l'acquisizione della capacità di volare, propria in diverso grado di quasi tutte le specie.
Nel giacimento cinese di Chengjiang sono state ritrovate lunghe catene di artropodi antiche almeno 525 milioni di anni, legati l'uno all'altro; sembrerebbe la più remota forma di comportamento cooperativo pervenuto ai nostri giorni. Una delle spiegazioni plausibili per questo comportamento è quella di una migrazione di massa, resa in tal modo più sicura[3].
Quello degli Arthropoda è il phylum più ricco di taxa e che conta il maggior numero di organismi viventi nel regno animale. Tradizionalmente sono riconosciuti quattro subphyla[4][5]:
Fuori da questi quattro sottotipi, gli artropodi comprendono un certo numero di forme fossili, alcune risalenti al Cambriano, difficili da collocare nella filogenesi del gruppo per la mancanza di una chiara affinità verso un qualsiasi altro gruppo associata a somiglianze con più di uno.
I trilobiti rappresentano uno dei più noti esempi di artropodi ormai estinti. Sono un gruppo di organismi marini scomparso durante l'estinzione permo-triassica, nonostante avessero già subito una forte riduzione dopo l'estinzione del Tardo Devoniano[6].
Dopo diversi anni di dibattito la comunità scientifica è concorde nell'affermare che i sopramenzionati quattro subphyla formino un gruppo monofiletico chiamato Euarthropoda, ben individuato da caratteri morfologici e molecolari[7][8]. Tuttavia le relazioni fra questi rimangono parzialmente incerte, come del resto è oggetto di dibattito la supposta parentela degli euartropodi con i tardigradi e gli onicofori che racchiuderebbe questi tre gruppi in un clade monofiletico denominato Panarthropoda[9]. Recenti studi, fortemente supportati, suggeriscono che il subphylum Crustacea sia parafiletico, in quanto gli esapodi si sarebbero evoluti da un clade, di collocazione ancora incerta, all'interno di questo gruppo. Secondo questa ipotesi filogenetica, i crostacei e gli esapodi formerebbero un clade, detto Pancrustacea[10][11]. Questa interpretazione si contrappone alla più antica idea di un clade, gli Atelocerata (o Tracheata, o Uniramia stricto sensu) che raccoglieva Esapoda con Myriapoda, escludendo i Crustacea, sulla base di caratteri morfologici[12]. Un numero crescente di evidenze molecolari smentisce questa filogenesi.
Euarthropoda Mandibulata Atelocerata Euarthropoda Mandibulata PancrustaceaAll'interno degli euartropodi, il maggior punto di disaccordo è stato a lungo rappresentato dalla posizione dei miriapodi. In alcuni studi i miriapodi vengono raggruppati con i chelicerati a formare un clade detto Myriochelata, sister-group dei pancrustacei[13]; altre analisi, più recenti e maggiormente supportate da dataset molecolari, suggeriscono che i miriapodi siano strettamente imparentati con i Pancrustacea, a formare un clade, i Mandibulata, che esclude i chelicerati[14][15].
Euarthropoda Myriochelata Euarthropoda MandibulataNell'ambito del più ampio raggruppamento degli Ecdysozoa, gli euartropodi mostrano tratti condivisi, tra cui la segmentazione e la presenza di appendici, con i Tardigrada e gli Onychophora, tuttavia una stretta relazione fra questi tre phyla non è stata chiaramente supportata da dati molecolari. Una parentela fra onicofori ed euartropodi è ampiamente accettata, ma le affinità con i tardigradi sono meno chiare, tant'è che questi ultimi sono spesso stati raggruppati con i nematodi in diversi studi filogenetici[16][17][18]. Recentemente, studi che hanno limitato artefatti filogenetici come long-branch attraction e compositional attraction, dovuti a fast-evolving taxa e a bias nella composizione in basi delle sequenze nucleotidiche di certi taxa del dataset, rigettano l'ipotesi che Nematoda e Tardigrada siano sister-group e mostrano una comune origine dei tre phyla che costituiscono i Panarthropoda[15][19]. Analisi più specifiche, volte a risolvere le relazioni di parentela fra tardigradi, onicofori e artropodi, sono state condotte nel 2010 sui marcatori mitocondriali e sono risultate nell'unione di Tardigrada e Onychophora come sister-group, una topologia che non trova un supporto morfologico[20]. Una parte degli autori di questa filogenesi ha rimesso in discussione questa stessa ipotesi l'anno successivo, sostenendo il sister-group Arthropoda + Onychophora sulla base di un dataset costituito da EST e microRNA e utilizzando un modello di sostituzione più adeguato ai dati disponibili (CAT-GTR)[21].
Panarthropoda Euarthropoda Mandibulata PancrustaceaGli artropodi (Arthropoda Latreille, 1829) sono un phylum (o tipo) di animali invertebrati protostomi celomati, che comprende circa i 5/6 delle specie finora classificate.
Il fatto che siano state descritte oltre un milione di specie di artropodi (e si stima che ne esistano 5 o forse 10 milioni) dimostra come la loro struttura di base sia versatile e adattabile a diversi modi di vita.
Arthropoda (ex verbis Graecis ἄρθρον 'articulus' + πούς 'pes' = 'pedes articulati') sunt animalia invertebrata quae exosceletum (sceletum exteriorem), corpus segmentosum, appendicesque articulatas habent. Ea sunt membra phyli inter quae sunt insecta, arachnida, crustacea, et alia animalia familiaria. Eorum maiores proprietates sunt membra articularia et cuticula, quae plerumque ex α-chitin constat; crustaceorum praeterea cuticula carbonato calcii biomineralizatur. Rigiditas cuticulae auctum inhibet; ergo arthropoda eam cuti exuta? interdum substituunt. Conformatio corporis arthropodorum in segmentis iteratis consistit, quorum quodque par habet appendicum. Ea est tam versatilis quod cum cultro exercitus Helvetici comparata est, atque arthropoda frequentissima omnium collegiorum oecologicorum in plurimis circumiectibus membra effecit. Sunt plus quam 1 000 000 specierum descriptarum, plus quam 80 centesimae omnium animalium vivorum specierum descriptarum, et ea sunt unus ex duobus gregibus animalibus quae in circumiectibus persiccis vigent (alius grex amniota comprehendit). Arthropoda magnitudine inter plancton microscopicos et formas nonnulla metra longas variantur.
Primaria arthropodorum lacuna interna est haemocoelum,? quod eorum organa accommodat et per quod sanguis circumagitur; eis sunt aperta systemata circulatoria. Sicut partes exteriores, interna arthropodorum organa plerumque ex segmentis iteratis constant. Systema nervosum est scaliforme, ventralibus restibus nervosis coniunctis per omnia segmenta transientibus et ganglia coniuncta in quoque segmento figurantibus. Caput per coniunctionem variorum numerorum segmentorum, cerebrum per coniunctionem gangliorum horum segmentorum nascitur, oesophagum circumplectens. Systemata respiratorium et excretorium variantur, tantum secundum circumiectum quantum subphylum ad quod pertinent.
Arthropoda (ex verbis Graecis ἄρθρον 'articulus' + πούς 'pes' = 'pedes articulati') sunt animalia invertebrata quae exosceletum (sceletum exteriorem), corpus segmentosum, appendicesque articulatas habent. Ea sunt membra phyli inter quae sunt insecta, arachnida, crustacea, et alia animalia familiaria. Eorum maiores proprietates sunt membra articularia et cuticula, quae plerumque ex α-chitin constat; crustaceorum praeterea cuticula carbonato calcii biomineralizatur. Rigiditas cuticulae auctum inhibet; ergo arthropoda eam cuti exuta? interdum substituunt. Conformatio corporis arthropodorum in segmentis iteratis consistit, quorum quodque par habet appendicum. Ea est tam versatilis quod cum cultro exercitus Helvetici comparata est, atque arthropoda frequentissima omnium collegiorum oecologicorum in plurimis circumiectibus membra effecit. Sunt plus quam 1 000 000 specierum descriptarum, plus quam 80 centesimae omnium animalium vivorum specierum descriptarum, et ea sunt unus ex duobus gregibus animalibus quae in circumiectibus persiccis vigent (alius grex amniota comprehendit). Arthropoda magnitudine inter plancton microscopicos et formas nonnulla metra longas variantur.
Primaria arthropodorum lacuna interna est haemocoelum,? quod eorum organa accommodat et per quod sanguis circumagitur; eis sunt aperta systemata circulatoria. Sicut partes exteriores, interna arthropodorum organa plerumque ex segmentis iteratis constant. Systema nervosum est scaliforme, ventralibus restibus nervosis coniunctis per omnia segmenta transientibus et ganglia coniuncta in quoque segmento figurantibus. Caput per coniunctionem variorum numerorum segmentorum, cerebrum per coniunctionem gangliorum horum segmentorum nascitur, oesophagum circumplectens. Systemata respiratorium et excretorium variantur, tantum secundum circumiectum quantum subphylum ad quod pertinent.
Nariuotakojai (Arthropoda) – pagal rūšių skaičių gausiausias gyvūnų karalystės tipas. Nariuotakojai – kosmopolitai, paplitę visame pasaulyje, išskyrus Antarktidos ledynus. Tai trisluoksniai, bilateraliniai gyvūnai, turintys segmentuotą kūną. Segmentai gali susilieti (oligomerizacija) ir sudaryti atskiras kūno dalis. Išsivysto nariuotos galūnės, vyksta raumenų diferenciacija (išsivysto skersaruožiai raumenys): atsiranda išorinis chitininis skeletas (egzoskeletas), apsaugantis nuo įvairaus poveikio, be to, prie jo prisitvirtina raumenys. Tarpus tarp organų užpildo miksocelis, susidaręs susiliejus pirminei bei antrinei kūno ertmėms. Pasižymi tobulomis virškinimo, kvėpavimo, šalinimo, kraujotakos, nervų, endokrinine ir lytine sistemomis bei sudėtingu vystymusi (būdingos įvairios lervinės stadijos).
Nariuotakojų kūnas skirstomas į atskiras dalis: galvą, krūtinę ir pilvelį. Kai kurių nariuotakojų galvos bei krūtinės segmentai susilieja ir virsta galvakrūtine. Kūną dengia kieta kutikula, sudaryta iš artimo celiuliozei junginio – chitino.
Nariuotakojų kraujotakos sistema atvira. Nugarinėje kūno pusėje yra gerai išsivysčiusi širdis. Kraujas vadinamas hemolimfa, kuri teka ne tik kraujagyslėmis, bet ir kūno ertmėmis (sinusais, lakūnomis). Virškinimo sistema sudaryta iš priekinės, vidurinės ir užpakalinės žarnos. Į vidurinę žarną atsiveria virškinimo liaukos. Užpakalinė žarna užsibaigia analine anga. Kvėpavimo sistema labai įvairi ir priklauso nuo gyvenimo būdo. Dauguma smulkiųjų nariuotakojų kvėpavimo organų neturi ir kvėpuoja visu kūno paviršiumi per plonytę kutikulą. Vandenyje gyvenančios rūšys turi žiaunas ar jų funkcijas atliekančias galūnes, o sausumoje – vėduoklinius plaučius ar trachėjas. Šalinimo sistemą sudaro Malpigijaus vamzdeliai ar koksalinės liaukos, kurias turi žemesnieji nariuotakojai. Nervų sistemą sudaro pilvo nervų grandinėlė, aplinkryklinis žiedas ir trijų skyrių smegenys. Reguliavimo funkcijas atlieka ir vidaus sekrecijos liaukos. Lytinė sistema išsivysčiusi skirtingai. Dauginasi tik lytiniu būdu. Dažnas lytinis dimorfizmas.
Vėžiagyviai Voragyviai Vabzdžiai


Nariuotakojų filogenetiniai ryšiai su primityviosiomis daugiašerėmis nekelia abejonių. Šios grupės išsiskyrė gana seniai, kai polichetų protėviai dar turėjo kopėtinę nervų sistemą, būdingą oligomeriniams anelidams. Polichetų kutikulė tapo chitinizuotu šarvu, prie kurio geriau prisitvirtina nariuotakojų diferencijuoti raumenys. Be to, šarvai buvo patikimesnė apsaugos priemonė. Nariuotakojų protėviai buvo judrūs gyvūnai, todėl jiems būdinga priekinės dalies cefalizacija. Iš polichetų palpų susidarė geresni jutimo organai – antenulės. Vietoj paprastų akių išsivystė facetinės. Iš dalies priekinių galūnių formavosi burnos organai. Polichetų dviejų skiaučių parapodijos tapo dvišakomis galūnėmis. Šalia nariuotakojų galūnių išsivystė žiauniniai priedai. Primityvieji nariuotakojai dar turėjo daug segmentų. Judėjo gyvatiškais judesiais, būdingais polichetams.
Dar labiau pakito vidaus sandara. Vietoj celomo išsivystė miksocelis. Todėl kraujotakos sistema tapo atvira. Nugarinė kraujagyslė virto vamzdelio formos širdimi. Nefridijų skaičius labai sumažėjo. Vietoj homonominės metametrijos tarp nariuotakojų prasidėjo oligomerizacija. Todėl, sumažėjus segmentų skaičiui, išsivystė heteronominė metametrija, pakito vidaus organai (sumažėjo nefridijų skaičius).
Nariuotakojai skilo į dvi šakas: trilobitinius ir vėžiagyvius. Trilobitinių vystymosi pradžioje dvišakos galūnės tapo nediferencijuotos vienšakos, pradėjo formuotis galvos skydas. Rasta fosilija: spiriggina. Tai pereinamoji forma tarp trilobitų ir polichetų. Galūnės nediferencijuotos, išsidėsčiusios kūno šonuose, kaip polichetų parapodijos. Turėjo trilobitams būdingą galvos skydą iš 4 liemens segmentų, iš kurių vėliau susidarė trilobitų galvos atkarpa. Spirigina jau turėjo stambias, tikriausiai, facetines akis. Tuose pat sluoksniuose aptiktos spiriginai priskiriamos lervos, artimos trilobitų lervai. Trilobitai išmirė dar paleozojuje, bet iš jų vystėsi skorpionvėžiai, kurie jau buvo cheliceriniai. Jie prarado antenules. Pirmosios dvi kojų poros tapo cheliceromis ir pedipalpais.
Skorpionvėžiai tokie panašūs į skorpionus, kad kartais būdavo sunku atskirti, kuriems priklauso rastoji fosilija. Jos diferencijuojamos pagal radimvietę. Skorpionai – sausumos gyvūnai. Skorpionvėžių kojelės atliko žiaunų funkcijas. Skorpionų jos – pilvelio viduje tapo vėduokliniais plaučiais.
Cheliceriniams (kaip ir vėžiagyviams) evoliucionuojant vyko oligomerizacija. Jeigu trilobitai dar turėjo iki 44 segmentų, tai skorpionvėžiai – 19, o voragyviai nuo 19 iki 10. Mažiausiai segmentų turi erkės. Primityviausi yra skorpionai ir pseudoskorpionai. Dar gerokai nariuotus kūnus turi solpūgos ir šienpjoviai. Susijungus jų kūnas tapo nenariuotas. Koncentruojasi ir nervų sistema. Tarp erkių aptinkama ir primityvių segmentuotų formų, tai rodo, jog ši grupė seniai atsiskyrė nuo bendro voragyvių kamieno. Chelicerinių primityvūs kvėpavimo organai – vėduokliniai plaučiai – pakinta į tobulesnes, geriau pritaikytas sausumos sąlygoms – trachėjas. Primityvieji vorai dar turi vėduoklinius plaučius, pvz., skorpionai, bet greta jų yra jau ir trachėjos. Sudėtingesnės sandaros vorai ir erkės turi tik trachėjas. Taigi voragyviams evoliucionuojant, primityvūs kvėpavimo organai pakito į tobulesnius.
Nariuotakojai (Arthropoda) – pagal rūšių skaičių gausiausias gyvūnų karalystės tipas. Nariuotakojai – kosmopolitai, paplitę visame pasaulyje, išskyrus Antarktidos ledynus. Tai trisluoksniai, bilateraliniai gyvūnai, turintys segmentuotą kūną. Segmentai gali susilieti (oligomerizacija) ir sudaryti atskiras kūno dalis. Išsivysto nariuotos galūnės, vyksta raumenų diferenciacija (išsivysto skersaruožiai raumenys): atsiranda išorinis chitininis skeletas (egzoskeletas), apsaugantis nuo įvairaus poveikio, be to, prie jo prisitvirtina raumenys. Tarpus tarp organų užpildo miksocelis, susidaręs susiliejus pirminei bei antrinei kūno ertmėms. Pasižymi tobulomis virškinimo, kvėpavimo, šalinimo, kraujotakos, nervų, endokrinine ir lytine sistemomis bei sudėtingu vystymusi (būdingos įvairios lervinės stadijos).
Posmkāji (Arthropoda) (grieķu: ἀρθρον — locītava un πούς/ποδός — kāja) — pirmmutnieku dzīvnieku tips, kas iekļauj sevī kukaiņus, vēžveidīgos, zirnekļveidīgos un daudzkājus. Tas ir sugām visbagātākais un izplatītākais dzīvnieku tips. Šī tipa pārstāvjus raksturojošā pamatīpašība ir tos klājošais ārējais hitīna skelets. Posmkāji ir izplatīti praktiski pa visu zemeslodi un tiem ir liela bioloģiska un ekoloģiska nozīme. Kukaiņi, līdzās ar zīdītājiem un putniem ir dominējošā sauszemes dzīvnieku grupa. Uz Zemes ir vairāk kā 1 000 000 kukaiņu sugu. Zirnekļveidīgie un daudzkāji, kaut arī nav sasnieguši tādu daudzveidību, tomēr arī ir ļoti izplatīti. Vēžveidīgie apdzīvo praktiski visus pasaules okeāna platuma grādus un dziļumus. Dažos reģionos vēžveidīgie ir dominējošā planktona sastāvdaļa.
Posmkāju taksonomiskā hierarhija nav viennozīmīga. No vienas puses, tika izlemts, ka daži posmkāju apakštipi nav radniecīgi ar daudzsaru tārpiem (Annelida). Konkrēti, skaitās, ka traheātu (Uniramia) apakštips ir tuvāks onihoforām (Onychophora), nekā citiem posmkājiem, taču lielākā zinātnieku daļa tam nepiekrīt, kā arī tas neiet kopā ar ģenētiskajiem pētījumiem. Cits viedoklis ir tradicionāli apgalvot, ka daudzsaru tārpi ir tuvākie radinieki kā posmkājiem, tā arī onihoforām, pamatojoties uz segmentu vienādo skaitu, tomēr vēlāk sāka runāt par konverģentu evolūciju, jo posmkāji ir tuvāki nematodēm (Nematoda), jo abiem ir spēja uz ārējā apvalka maiņu. Divi iespējamie ģeneoloģiskie atzari arī nosaka divus augstākos taksonus, pie kuriem varētu pieskaitīt posmkājus: posmainie (Articulata) un apvalku metošie (Ecdysozoa).
Apakštaksonu klasifikācija posmkāju grupas iešpusē arī ir neviennozīmīga. Piecas galvenās apakšgrupas tiek piedēvētas vai nu apakštipiem vai klasēm. Bez šīm apakšgrupām ir virkne fosilo organismu, lielākoties no apakškembrija, kurus ir grūti pieskaitīt kādai no šīm grupām gan tāpēc, ka tie būtiski atšķiras no jau zināmajiem organismiem, gan arī aiz neskaidrām savstarpējām radniecīgajām saitēm. Agrāk daudzkājus un kukaiņus bieži vien apvienoja traheātos (Tracheata), bet beidzamie pētījumi rāda, ka daudzkāji nav tuvāki kukaiņiem kā vēžveidīgajiem.
Tips Posmkāji (Arthropoda)
(†) - izmirušu organismu grupa.
Posmkāji (Arthropoda) (grieķu: ἀρθρον — locītava un πούς/ποδός — kāja) — pirmmutnieku dzīvnieku tips, kas iekļauj sevī kukaiņus, vēžveidīgos, zirnekļveidīgos un daudzkājus. Tas ir sugām visbagātākais un izplatītākais dzīvnieku tips. Šī tipa pārstāvjus raksturojošā pamatīpašība ir tos klājošais ārējais hitīna skelets. Posmkāji ir izplatīti praktiski pa visu zemeslodi un tiem ir liela bioloģiska un ekoloģiska nozīme. Kukaiņi, līdzās ar zīdītājiem un putniem ir dominējošā sauszemes dzīvnieku grupa. Uz Zemes ir vairāk kā 1 000 000 kukaiņu sugu. Zirnekļveidīgie un daudzkāji, kaut arī nav sasnieguši tādu daudzveidību, tomēr arī ir ļoti izplatīti. Vēžveidīgie apdzīvo praktiski visus pasaules okeāna platuma grādus un dziļumus. Dažos reģionos vēžveidīgie ir dominējošā planktona sastāvdaļa.
Artropoda adolah filum nan paliang gadang dalam dunia binatang dan mencakup serangga, laba-laba, udang, lipan, jo binatang sajinih lainnyo. Artropoda biaso ditamui di lauik, aia tawa, darek, jo lingkuang udaro, tamasuak babagai bantuak simbiosis jo parasit. Kato artropoda barasa dari bahaso Yunani ἄρθρον árthron, "rueh, buku, atau segmen", jo πούς pous (podos), "kaki", nan kok disatuan baarati "kaki barueh-rueh"[2]. Artropoda dikana juo jo namo binatang barueh atau binatang babuku-buku.
Karakteristik nan khas sainggo artropoda babeda jo filum nan lain yaitu: tubuah basegmen, segmen biasonyo basatu manjadi duo atau tigo daerah nan jaleh, anggota tubuah basegmen bapasangan (asa panamoan Artropoda), simetribu bilateral, eksoskeleton (rangka lua) bakitin. Sacaro bakalo mangalia jo dipabaharui sabagai patumbuahan binatang, kanal alimentari sarupo pipa antaro muluik jo anus, sistem sirkulasi tabukak, hanyo pambuluah darah nan biasonyo babantuak sabuah struktur dorsal (balakang) co pipa manuju kanal alimentar jo bukaan lateral (sampiang) di daerah abdomen (paruik), rongga tubuh; sabuah rongga darah atau hemosol jo selom tareduksi.
Garih evolusi artropoda dapek ditalusuri dari periode Kambrium. Kalompok ko umumnyo dianggap sabagai monofiletik dan banyak analisis nan mandukung panampekan artropoda basamo jo cycloneuralia dalam superfilum Ecdysozoa. Tapi, hubuangan dasa Metazoa alun dikatahui sacaro pasti. Baitu pulo hubuangan di antaro sasamo kalompok artropoda masih acok dipadebatkan.
Artropoda mamiliki kontribusi pado suplai makanan manusia, baiak langsuang (sabagai makanan) maupun indak langsuang, sarupo mambantu panyerbukan. Sabagian spesies tatantu dikatahui marupokan vektor panyakik babahayo bagi manusia, binatang, jo tumbuahan.
Artropoda adolah filum nan paliang gadang dalam dunia binatang dan mencakup serangga, laba-laba, udang, lipan, jo binatang sajinih lainnyo. Artropoda biaso ditamui di lauik, aia tawa, darek, jo lingkuang udaro, tamasuak babagai bantuak simbiosis jo parasit. Kato artropoda barasa dari bahaso Yunani ἄρθρον árthron, "rueh, buku, atau segmen", jo πούς pous (podos), "kaki", nan kok disatuan baarati "kaki barueh-rueh". Artropoda dikana juo jo namo binatang barueh atau binatang babuku-buku.
Karakteristik nan khas sainggo artropoda babeda jo filum nan lain yaitu: tubuah basegmen, segmen biasonyo basatu manjadi duo atau tigo daerah nan jaleh, anggota tubuah basegmen bapasangan (asa panamoan Artropoda), simetribu bilateral, eksoskeleton (rangka lua) bakitin. Sacaro bakalo mangalia jo dipabaharui sabagai patumbuahan binatang, kanal alimentari sarupo pipa antaro muluik jo anus, sistem sirkulasi tabukak, hanyo pambuluah darah nan biasonyo babantuak sabuah struktur dorsal (balakang) co pipa manuju kanal alimentar jo bukaan lateral (sampiang) di daerah abdomen (paruik), rongga tubuh; sabuah rongga darah atau hemosol jo selom tareduksi.
Garih evolusi artropoda dapek ditalusuri dari periode Kambrium. Kalompok ko umumnyo dianggap sabagai monofiletik dan banyak analisis nan mandukung panampekan artropoda basamo jo cycloneuralia dalam superfilum Ecdysozoa. Tapi, hubuangan dasa Metazoa alun dikatahui sacaro pasti. Baitu pulo hubuangan di antaro sasamo kalompok artropoda masih acok dipadebatkan.
Artropoda mamiliki kontribusi pado suplai makanan manusia, baiak langsuang (sabagai makanan) maupun indak langsuang, sarupo mambantu panyerbukan. Sabagian spesies tatantu dikatahui marupokan vektor panyakik babahayo bagi manusia, binatang, jo tumbuahan.
Rujuakan Lua error in Modul:Citation/CS1 at line 3828: attempt to index field 'date_names' (a nil value). "Arthropoda". Online Etymology Dictionary. http://www.etymonline.com/index.php?term=Arthropoda&allowed_in_frame=0.
Artropod ialah haiwan invertebrat yang mempunyai rangka luar, badan bersegmen, dan anggota bersendi. Artropod ialah ahli dalam filum Arthropoda (daripada bahasa Greek ἄρθρον árthron, "sendi", dan ποδός podós "kaki", yang bersama-sama bermaksud "kaki bersendi"), dan termasuklah serangga, araknid, krustasea, dan lain-lain.
Artropod ialah haiwan invertebrat yang mempunyai rangka luar, badan bersegmen, dan anggota bersendi. Artropod ialah ahli dalam filum Arthropoda (daripada bahasa Greek ἄρθρον árthron, "sendi", dan ποδός podós "kaki", yang bersama-sama bermaksud "kaki bersendi"), dan termasuklah serangga, araknid, krustasea, dan lain-lain.
Geleedpotigen (Arthropoda) zijn koudbloedige dieren met een uitwendig skelet van chitine, eventueel versterkt met calciumcarbonaat, waarvan de poten een aantal gewrichten hebben. Hun lichaam is gesegmenteerd, waarbij verschillende ondergroeperingen kunnen zijn ontstaan (tagmosis, versmelting/samengroeiing van een aantal segmenten), bijvoorbeeld bij spinnen in een kopborststuk en achterlijf, bij insecten in een kop, borststuk en achterlijf. Ze vormen de grootste stam van het dierenrijk en komen in alle leefomgevingen (land, zoet water en zout water) voor. De insecten alleen al tellen, exclusief niet-ontdekte soorten, ongeveer 5-10 miljoen soorten,[1] en volgens sommige theorieën wel 30 miljoen, maar voor 1980 waren er slechts 200.000 soorten beschreven; in 2008 zijn dat er al 1.000.000.
Tot de Arthropoda behoren de zespotigen (voornamelijk insecten), de (achtpotige) spinachtigen, de (tien tot veertienpotige) kreeftachtigen, de duizendpotigen (duizend- en miljoenpoten) en de uitgestorven trilobieten.
Ongeveer vier op de vijf diersoorten op aarde zijn geleedpotigen.
De geleedpotigen worden in vier groepen (onderstammen) ingedeeld. De groepen onderscheiden zich in diverse morfologische kenmerken van elkaar. Belangrijke punten zijn de vergroeiingen van de segmenten (tagmosis) en de aan- of afwezigheid van bepaalde ledematen aan de segmenten. Een oersegment van een geleedpotige heeft 2 paar ledematen. Het bovenste paar heeft de functie van een kieuw en is geveerd, het onderste paar wordt gebruikt om op te lopen. In veel gevallen is slechts een van de twee paar ledematen per segment aanwezig. De ledematen kunnen ook van functie zijn veranderd tot voelsprieten (antennes), vleugels of monddelen.
Er zijn weleens voorstellen gedaan dat deze groepen polyfyletisch zijn en dus onafhankelijk van elkaar zijn ontstaan. De overwegende opinie bij onderzoekers is echter dat de geleedpotigen evolutionair gezien één groep vormen.
In de jaren 70 werden de uit het Cambrium afkomstige fossielen van de Burgess Shale nauwkeurig bestudeerd. Het meest voorkomende fossiel in de Burgess Shale is Marrella, dat eerder werd ingedeeld bij de trilobieten. Het bleek echter dat Marrella in geen van de vier bekende groepen kon worden ondergebracht: Marrella heeft twee paar uitsteeksels aan de kop, beide voor de mond geplaatst. In moderne indelingen van de Geleedpotigen wordt het daarom vaak met enkele verwante soorten in een eigen onderstam geplaatst, de Marrellomorpha.
Ook met toevoeging van de Marrellomorpha blijven er echter nog fossiele geleedpotigen uit de Burgess Shale en andere paleozoïsche vindplaatsen waarvan de indeling niet duidelijk is, zoals Yohoia, Burgessia en Branchiocaris.
Geleedpotigen (Arthropoda) zijn koudbloedige dieren met een uitwendig skelet van chitine, eventueel versterkt met calciumcarbonaat, waarvan de poten een aantal gewrichten hebben. Hun lichaam is gesegmenteerd, waarbij verschillende ondergroeperingen kunnen zijn ontstaan (tagmosis, versmelting/samengroeiing van een aantal segmenten), bijvoorbeeld bij spinnen in een kopborststuk en achterlijf, bij insecten in een kop, borststuk en achterlijf. Ze vormen de grootste stam van het dierenrijk en komen in alle leefomgevingen (land, zoet water en zout water) voor. De insecten alleen al tellen, exclusief niet-ontdekte soorten, ongeveer 5-10 miljoen soorten, en volgens sommige theorieën wel 30 miljoen, maar voor 1980 waren er slechts 200.000 soorten beschreven; in 2008 zijn dat er al 1.000.000.
Tot de Arthropoda behoren de zespotigen (voornamelijk insecten), de (achtpotige) spinachtigen, de (tien tot veertienpotige) kreeftachtigen, de duizendpotigen (duizend- en miljoenpoten) en de uitgestorven trilobieten.
Ongeveer vier op de vijf diersoorten op aarde zijn geleedpotigen.
Leddyra er den største dyregruppa på jorda, og omfattar over ein million kjende artar. Dei vert kjenneteikna ved å ha eit eksoskjelett og ein tydeleg tagmatisert kropp; det vil seie at kroppen er bygd opp av fleire ulike mindre segment som har fusjonert og spesialisert seg til ulike funksjonar.
Til leddyra høyrer m.a. insekta, edderkoppdyra, krepsdyra, skolopendrane og tusenbeina. Gruppa finst i alle slags habitat over heile kloten, og er påvist i over 500 millionar år gamle fossil.
Leddyra er protostome eucoelomatar med tilbakedanna kroppshole og ope blodkarsystem. Dei har segmentert kropp, kor fleire segment saman dannar større strukturar. Opphavleg trur ein desse kroppane har vore metameriske som hjå leddmakk - at segmenta har sett like ut - men hjå dagens arthropodar har dei i stor grad fått spesialiserte funksjonar. Dette er kjend som tagmose.
Ein leddyrkropp er dekt av eit ytre skjelett av kitin og vassavstøytande lipoprotein. Dette vert skild ut or hypodermis, som svarar til epidermis hjå menneske og er det ytste laget dyret har av levande cellevev. Fordelane med eit eksoskjelett er at det stabiliserer kroppen og gjev muskelfeste. Det syt òg for at dyret kan stå oppreist utan å bruka muskelkrefter. Baksida av medaljen er at alle dyr med dette må skifta skal undervegs i livsløpet for å kunne halda fram veksten sin. I tillegg er eksoskjelettet tungt; noko som gjer at leddyr aldri kan bli spesielt store med mindre dei lever i vatn.
Ein annan fordel leddyra har framfor mange andre virvellause dyr er dei ledda ekstremitetane deira. Desse vert brukt til rørsle, matinntak, forsvar og sansing; og sit i par på dei ulike kropssegmenta. Det er leddelinga i beina som er det viktigaste skiljet mellom Arthropoda og dei nærskylde rekkjene Onynchophora (fløyelsdyr) og Tardigrada (bjørnedyr).
Trilobittane levde for 540-250 millionar år sidan i paleozoikum-tida, og levde i havet. Dei fanst i eit breitt spekter av økologiske nisjar, og varierte i storleik frå nokre få millimeter til ein dryg meter. Det er denne dyregruppa me finn flest av i norske fossilførande lag.
Trilobittane hadde tredelt kropp beståande av hovud, thorax (mellomkropp) og pygidium (hale). Dei skilde seg frå moderne leddyr m.a. ved å ha eksoskjelettplater berre på oversida.
Cheliceratane, eller klosaksdyra, lever i alle habitat og omfattar dolkehalar, sjøskorpionar †, havedderkoppar og edderkoppdyr. Desse har todelt kropp delt i prosoma og opisthosoma, og er vanlegvis utrusta med fire beinpar.
Prosoma, eller framkroppen, består av åtte segment og eit presegmentalt acron. Dei to første segmenta er utan ekstremitetar, medan chelicerane eller klosaksane som har gjeve gruppa namnet sitt sit på tredje. Dette er skarpe kjevedelar som ved mange høve inneheldt gift. Fjerde til åttande segment bèr som regel beinpar eller pedipalpar.
Opisthosoma eller abdomen består av tolv segment og eit postsegmentalt telson. Telsonet er ein slags hale som er mest tydeleg hjå dolkhalar og skorpionar. Edderkoppane manglar dette, men har i staden spinnvorter som produserer silke her. Ekstremitetane på bakkroppen er som regel sterkt tilbakedanna eller redusert til gjellestrukturar.
Krepsdyra, Crustacea, lever i havet, i ferskvatn eller i fuktige terrestriske habitat. Desse har tredelt kropp beståande av hovud, thorax og abdomen eller pleon, men ved mange høve smeltar hovud og thorax saman til ein cephalothorax.
Hovudet ber to par antenner, eit par fasettaugo og tre par munndelar kalla maxillar og mandiblar. Thorax og pleon ber ei rekkje laterale ekstremitetar, inkludert gjellene, og ender i eit telson.
Krepsdyra opptrer i ei rekkje ulike former, men har det til felles at larvestadiet tek til med ei såkalla naupliuslarve. Dei er òg dei einaste nolevande leddyra med klart todelte (=birame) ekstremitetar.
Mangefotingane (Myriapoda) består m.a. av tusenbein og skolopendrar, og er dei sterkast metameriske leddyra. Kroppen deira består av eit hovud og ei lang rekkje like segment (15-375) med eit gitt tal på ekstremitetar. Alle artane lever på land, fordi trakéopningane deira ikkje kan lukkast, og gruppa omfattar både rovdyr og planteetarar.
Seksfotingane (Hexapoda) er seksbeinte leddyr med ein tredelt kropp som består av hovud, thorax og abdomen. Det er beskrive rundt 900 000 artar i denne underrekka, og om lag 3500 nye kjem til kvart år. Forskarar anslår at det finst mellom 5 og 80 millionar artar av dei.
Dei fleste seksfotingar er insekt.
Leddyra er den største dyregruppa på jorda, og omfattar over ein million kjende artar. Dei vert kjenneteikna ved å ha eit eksoskjelett og ein tydeleg tagmatisert kropp; det vil seie at kroppen er bygd opp av fleire ulike mindre segment som har fusjonert og spesialisert seg til ulike funksjonar.
Til leddyra høyrer m.a. insekta, edderkoppdyra, krepsdyra, skolopendrane og tusenbeina. Gruppa finst i alle slags habitat over heile kloten, og er påvist i over 500 millionar år gamle fossil.
Leddyr utgjør med sine mer enn 1 million kjente arter, om lag 80 % av alle dyrearter på jorda.[1] I tillegg antas det at det finnes flere millioner uoppdagede arter av leddyr. Leddyrene har inntatt nær sagt alle biotoper på jorda, inklusive havet. De fire store leddyrgruppene er insekter, edderkoppdyr og krepsdyr, foruten de utdødde trilobittene. Den økologiske utbredelsen fordeler seg med insekter og edderkoppdyr på land, og krepsdyr og havedderkopper i vann.[2] Siden insektene i regnskogen antakelig er svært tallrike, har det vært anslått at antall arter av leddyr kan være så høyt som 30 millioner.[3]
Enkelte forskere inkluderer leddyrenes nermeste slektninger - bjørnedyrene og fløyelsdyrene - og skiller da mellom leddyr i vid forstand (Arthropoda eller Panarthropoda) og egentlige leddyr (Euarthropoda eller Arthropoda).[4] Bjørnedyr og fløyelsdyr mangler imidlertid de typiske leddelte beina som kjennetegner leddyrene, og regnes vanligvis ikke med i leddyrene. Fløyelsdyr eller fløyelsormer (Onychophora) atskilte seg tidlig i evolusjonsforløpet og grenet av fra resten av leddyra.[5]
Det mest fremtredende fellestrekket for de egentlige leddyrene er at de har en segmentert (leddelt) kropp, og et hardt ytre skall. Hvert kroppsledd har opprinnelig hatt et lem, men dette kan siden gjennom evolusjonen være forsvunnet eller omdannet til antenner eller kjevedeler. Det er også normalt at leddyr er bilateralt symmetriske, og at hele kroppen inkludert beina og øynene er dekket med et ytre skjelett av stive skjelettplater. Skjelettplatene kalles ofte kutikulaen, og er primært bygget opp av kitin, som i ren form er meget mykt. For å styrke kutikulaen er den herdet (sklerotisert) gjennom avleiring av mineraler, proteiner, fenoler, og hos krepsdyr også kalk og kalsiumkarbonat. Hos gruppa Ichneumonoidea blant årevingene er det påvist forsterkning med metallene sink og mangan. Skjelettplatene er døde, de består ikke av levende celler slik som virveldyrenes knokler, får og fjær er.[6]
Når dyret vokser må skallet skiftes, ved at skjelettplatene løses opp innenfra mens det samtidig dannes nye som avleires fra huden. Når dyret kryper ut av sitt gamle skjell er det nye skjelettet fortsatt mykt, og i løpet av noen dager kan dyret vokse før det igjen dekkes av harde skallplater.[7] Hos mange grupper kan skallskiftet i forbindelse med en omdanning (metamorfose) av dyret fra et stadum til et annet.[8]
Noen av kroppens ledd ble i stamarten «sammenfattet» til større kroppsavsnitt (såkalte tagmata). Dette var opprinnelig hode (cephalon eller caput) og resten av kroppen. I mange delgrupper oppstår ytterligere tagmata, som hos insekter, krepsdyr og edderkoppdyr - som har tre tagmata: hode, bryststykke og bakkropp. Hodet (cehpalon, prosoma) kan da også være sammenvokst med brystet (thorax, mesosoma) til et sammenvokst segment - hodebryststykket (cephalothorax). Bakkroppen utgjør da det tredje segmentet (pygidium, metasoma); eller subsidiært det andre segmentet (abdomen) dersom hode og kropp er sammenvokst.
Både kroppen som helhet og beina må være leddet, dvs. at det må finnes bevegelige områder mellom skjelettplatene, for at dyret skal kunne bevege seg. Lemmene beveges ved hjelp av indre muskler, og beskyttelsen fra skjelettet gjør at dyrene kan være svært aktive. Noen av beina hos den opprinnelige stamarten brukes ikke lenger som gang- eller svømmebein: Det første (forhenværende) beinpar danner en antenne («følehorn») fremst på hodet, mens de neste beinpar bak er blitt til munndeler. Munndelene blir i mange delgrupper videre spesialisert til under- og overkjever. Utover antennene har hodet et par fasettøyne samt i mange arter to ytterligere par med mindre, såkalte medianøyne, dersom de ikke har blitt redusert bort under artens evolusjon.
Leddyrene har tverrstripede bøye- og strekkmuskler, og ikke hudmuskelsekker slik leddormene har. Musklene er hos insektene direkte festet til den indre overflaten av eksoskjelettet. Hjertet og tarmveggen hos leddyr har en glatt muskulatur. Nervesystemet er som regel godt utviklet i leddyrene, og artene i havet har et utviklet system av blodkar, selv om blodet også kan strømme fritt mellom cellene i selve vevet - og enkelte krepsdyr mangler sågar et hjerte. Men mange har et dorsalt plassert rørformet hjerte med muskelsekk som pumper blodet rundt, via arterier til et helt åpent blodsystem hvor blodet pumpes rundt til vevet og cellene vedd hjelp av nervestyrte muskelbevegelser. Åndedrettet blant krepsdyrene i vann skjer med gjeller, og en rekke arter kan foretå gassutveksling direkte gjennom huden, det kan skje gjennom bena eller antennene eller i sjeldne tilfeller gjennom hele kroppens overflate. Landlevende leddyr puster ved hjelp av ånderør (trakéer), som frakter oksygen direkte til vevet og cellene. Enkelte grupper har også boklunger, mens de fleste leddyr kvitter seg med fordøyd mat (ekskresjon) gjennom såkalte malpighiske rør.[9]
Hjernen er relativt sett velutviklet, og sitter på ryggsiden bak svelget. Den er festet via en dobbelt nervestreng til en nervering rundt svelget, og de ulike leddene har også utviklede nervetilknytninger. Leddyra har mest instinktstyrt atferd, men er også i stand til å lære. Mange grupper har sensoriske følehår på utsiden, men det er bare noen grupper av insekter som er i stand til å høre.[10] Leddyr orienterer seg ved hjelp av syn hvor dyret sanser lys. Leddyrene er utstyrt med fotoreseptorceller som ofte er plassert i eller ved fasettøynene. Hvert øye ser bare et utsnitt av omgivelsene, og disse utstnittene settes sammen i et mosaikksyn. Øynene er sensitive for bølgelengder fra UV-lys til rødt lys, og registrerer også polarisert lys. Fotoreseptorene er i stand til å skille mellom lys som kommer direkte fra lyskilden, lys som er reflektert fra andre objekter og lys som skinner gjennom et materiale (for eksempel blader). På denne måten kan insektene orientere seg etter farge, form, mønster og bevegelse. Enkelte insektlarver (som forskjellige sommerfugler og møll) er sensitive for UV-stråler, blått og grønt lys, og bruker disse egenskapene til å finne frem til skjulte områder under blader hvor de kan forpuppe seg i fred for omverdenen.
De fleste leddyr er særkjønnede, det vil si enten hann eller hunn, selv om noen arter også er tvekjønnede. Det skjer ofte en indre befruktning. Formeringen skjer ved at det voksne individet legger egg som klekkes etter noen dager. Noen arter kan legge og klekke ikke-befruktede egg, som en ukjønnet formeringsstrategi, men kjønnet befruktning er det normale. Det klekkede dyret lever da normalt i et stadium på noen dager eller inntil en måned som larve, hvoretter det omdannes til voksent, kjønnsmodent stadium som vanligvis omfatter resten av levealderen. Blant insekter er stadiene egg-larve-nymfe-voksen, og man skiller systematisk mellom de insektene som gjennomgår såkalt fullstendig forvandling med puppestadie, de som har vinger og gjennomgår en viss forvandling (egentlige insekter), og endelig de som har liten eller ingen forskjell mellom unge og voksne (urinsektene).[11]
Leddyr kan være planteetere, syreetere, eller altetende.
Systematiseringen av leddyrene er svært vanskelig og omstridt, ulike forfattere og forskningsmiljøer bruker ulike inndelinger. Innenfor fylogenien er det et prinsipp at man forsøker å skille ut naturlige grupper kalt klader, hvor artene har felles stamfar. Dersom gruppen består av undergrupper eller individer fra ulike klader, anses den ikke naturlig men parafyletisk, som eksempelvis gruppen Maxillopoda.Samtidig er mye av systematikken utviklet før molekylærbiologien skjøt fart, ofte på basis av morfologi, det vil si ytre kjennetegn som antall ledd, antall synlige ben, størrelse, osv. Noen regner leddyrene som helhet som en rekke, andre regner hver av hovedgruppene som rekker. Nedenfor angis en vanlig gruppering av leddyrene i grove trekk, ned til nivået klasse.
Leddyr utgjør med sine mer enn 1 million kjente arter, om lag 80 % av alle dyrearter på jorda. I tillegg antas det at det finnes flere millioner uoppdagede arter av leddyr. Leddyrene har inntatt nær sagt alle biotoper på jorda, inklusive havet. De fire store leddyrgruppene er insekter, edderkoppdyr og krepsdyr, foruten de utdødde trilobittene. Den økologiske utbredelsen fordeler seg med insekter og edderkoppdyr på land, og krepsdyr og havedderkopper i vann. Siden insektene i regnskogen antakelig er svært tallrike, har det vært anslått at antall arter av leddyr kan være så høyt som 30 millioner.
Enkelte forskere inkluderer leddyrenes nermeste slektninger - bjørnedyrene og fløyelsdyrene - og skiller da mellom leddyr i vid forstand (Arthropoda eller Panarthropoda) og egentlige leddyr (Euarthropoda eller Arthropoda). Bjørnedyr og fløyelsdyr mangler imidlertid de typiske leddelte beina som kjennetegner leddyrene, og regnes vanligvis ikke med i leddyrene. Fløyelsdyr eller fløyelsormer (Onychophora) atskilte seg tidlig i evolusjonsforløpet og grenet av fra resten av leddyra.

Stawonogi (Arthropoda, z gr. ἄρθρον arthron – staw + πούς, ποδός pous, podos – noga) – najliczniejszy w gatunki typ zwierząt na Ziemi. Dotychczas opisano ponad milion gatunków. Według danych IUCN opisano 950 000 gatunków owadów i 40 000 skorupiaków. Liczby w pozostałych grupach stawonogów wahają się – w zależności od źródła – od kilkunastu tysięcy (np. wije) do ponad 60 tysięcy gatunków pajęczaków, a z każdym rokiem przybywa ich coraz więcej.
Nazwę „stawonogi” zwierzęta te zawdzięczają charakterystycznym dla wszystkich przedstawicieli tego typu wyrostkom, których człony połączone są stawami.
Stawonogi zamieszkują wszystkie środowiska nadające się do życia (od głębin morskich po szczyty gór), co jest możliwe dzięki ich licznym przystosowaniom fizjologicznym, biologicznym, anatomicznym, jak i dużej rozrodczości.
Przypuszcza się, że najliczniejszym i najszerzej rozprzestrzenionym lądowym stawonogiem na świecie jest Oppiella nova, gatunek mechowca[2].
Wiele osobników ma zdolności do diapauzy (obniżenia poziomu metabolizmu), dzięki czemu mogą przetrwać niekorzystne warunki panujące w środowisku. Długość życia jest bardzo zróżnicowana, żyją od kilku dni do kilkudziesięciu lat, kraby nawet do 35 lat. Według niektórych badaczy japoński krab pacyficzny dożywa nawet wieku około 100 lat, jednak przypuszczenia te nie zostały nigdy potwierdzone.
Mięśnie występujące u stawonogów można podzielić na zginacze oraz prostowniki, zapewniające możliwość zmiany położenia elementów szkieletowych – a więc umożliwiające ruch. Podstawą działania odnóży składających się z członów jest ich połączenie za pomocą stawów.
Skorupiaki Pajęczaki Owady czułki (2 pary)Budowa układu oddechowego jest zależna od środowiska życia.
Skorupiakokształtne – skrzela, wymiana gazów całą powierzchnią ciała.
Pajęczaki – tchawki, płucotchawki, narządy oddechowe mogą również nie występować.
Tchawkowce – tchawki lub brak narządów.
Układ krążenia stawonogów jest otwarty, składa się z serca i naczyń. Płyn krążący w tym układzie nazywa się hemolimfą. Serce o podługowatym kształcie, z którego wychodzi kilka naczyń krwionośnych, położone jest po grzbietowej stronie ciała w worku osierdziowym. Serce posiada ostia, przez które hemolimfa jest wprowadzana do serca. Barwniki to niebieska hemocyjanina oraz czerwona hemoglobina.
Układ pokarmowy jest podobny do budowy pierścienic – zawsze składa się z trzech odcinków: jelita przedniego, środkowego i tylnego. Zarówno jelito przednie, jak i tylne wysłane są kutykulą. Dlatego nie mogą zachodzić tutaj żadne procesy trawienia i wchłaniania. Zachodzą tylko w jelicie środkowym. Otwór gębowy jest otoczony narządami gębowymi, umożliwiającymi m.in. pobieranie i mechaniczne rozdrabnianie pokarmu. Stawonogi odżywiające się pokarmem płynnym (np. pajęczaki, owady krwiopijne) mają umięśnioną gardziel ssącą lub żołądek ssący. Skorupiaki posiadają żołądek podzielony na dwie części - rozdrabniająca pokarm chitynowymi listewkami oraz trawiąca, do której ujście ma gruczoł wątrobowo-trzustkowy. U pajęczaków zachodzi trawienie zewnętrzne i późniejsze zasysanie nadtrawionych części ofiary. U owadów następuje duża modyfikacja aparatu gębowego.
U stawonogów występują:
Cewki Malpighiego występują u owadów, wijów i wyżej uorganizowanych pajęczaków.
Stawonogi wodne wydalają głównie silnie rozcieńczony amoniak. Lądowe zaś, które muszą oszczędzać wodę, przetwarzają amoniak w związki azotowe słabo rozpuszczalne w wodzie, jak guanina (pajęczaki) czy kwas moczowy (owady).
Typ: stawonogi (Arthropoda)
Współczesne analizy filogenetyczne wskazują, że stawonogi są monofiletyczną grupą wewnątrz kladu wylinkowców (Ecdysozoa). Ich najbliższymi krewnymi są prawdopodobnie pazurnice i niesporczaki, wraz ze stawonogami należące do grupy Panarthropoda (niektóre analizy genetyczne sugerowały jednak, że niesporczaki są bliżej spokrewnione z nicieniami). Do stawonogów zaliczano niektórych przedstawicieli fauny ediakarańskiej, takich jak rodzaje Spriggina i Parvancorina, jednak ich przynależność do stawonogów jest bardzo wątpliwa. Najstarsze znane skamieniałości stawonogów pochodzą z wczesnego kambru – są to odciski odnóży pozostawione przez przedstawiciela ichnorodzaju Monomorphichnus na obecnych terenach Nowej Fundlandii. Są one znacznie starsze od najstarszych znanych skamieniałości ciała należących bezsprzecznie do stawonogów[3].
Na podstawie:
Halanych et al. Evidence from 18S ribosomal DNA that the lophophorates are protostome animals. „Science”. 267 (5204), s. 1641–1643, 1995. DOI: 10.1126/science.7886451 (ang.).
Edgecombe et al. Higher-level metazoan relationships: recent progress and remaining questions. „Organisms Diversity and Evolution”. 11, s. 151–172, 2011. DOI: 10.1007/s13127-011-0044-4 (ang.).
Stawonogi (Arthropoda, z gr. ἄρθρον arthron – staw + πούς, ποδός pous, podos – noga) – najliczniejszy w gatunki typ zwierząt na Ziemi. Dotychczas opisano ponad milion gatunków. Według danych IUCN opisano 950 000 gatunków owadów i 40 000 skorupiaków. Liczby w pozostałych grupach stawonogów wahają się – w zależności od źródła – od kilkunastu tysięcy (np. wije) do ponad 60 tysięcy gatunków pajęczaków, a z każdym rokiem przybywa ich coraz więcej.
Nazwę „stawonogi” zwierzęta te zawdzięczają charakterystycznym dla wszystkich przedstawicieli tego typu wyrostkom, których człony połączone są stawami.
Stawonogi zamieszkują wszystkie środowiska nadające się do życia (od głębin morskich po szczyty gór), co jest możliwe dzięki ich licznym przystosowaniom fizjologicznym, biologicznym, anatomicznym, jak i dużej rozrodczości.
Przypuszcza się, że najliczniejszym i najszerzej rozprzestrzenionym lądowym stawonogiem na świecie jest Oppiella nova, gatunek mechowca.
Wiele osobników ma zdolności do diapauzy (obniżenia poziomu metabolizmu), dzięki czemu mogą przetrwać niekorzystne warunki panujące w środowisku. Długość życia jest bardzo zróżnicowana, żyją od kilku dni do kilkudziesięciu lat, kraby nawet do 35 lat. Według niektórych badaczy japoński krab pacyficzny dożywa nawet wieku około 100 lat, jednak przypuszczenia te nie zostały nigdy potwierdzone.
Artrópodes (Arthropoda, do grego arthros (ἄρθρον), articulado e podos (ποδός), pés) são um filo de animais invertebrados que possuem exoesqueleto rígido e vários pares de apêndices articulados, cujo número varia de acordo com a classe.
Compõem o maior filo de animais existentes, representados por animais como os gafanhotos (insetos), as aranhas (aracnídeos), os caranguejos (crustáceos), as centopeias (quilópodes) e os piolhos-de-cobra (diplópodes). Têm cerca de um milhão de espécies descritas, e estima-se que os representantes deste filo equivalem a cerca de 84% de todas as espécies de animais conhecidas pelo homem.[3] Possuem uma ampla gama de cores e formatos, e no que diz respeito ao tamanho, alguns vão desde as formas microscópicas, como no plâncton (com menos de 1/4 de milímetro), até crustáceos com mais de 3 metros de espessura.[4]
Sua existência é datada nos registros fósseis desde o período Cambriano (há cerca de 542 a 488 milhões de anos), onde criaturas como as Trilobitas eram encontradas em abundância nos oceanos.[5] Algumas teorias sobre a origem deste filo sustentam que os ancestrais dos artrópodes podem ter sido os anelídeos (vermes de corpo segmentado em anéis) ou de algum outro ancestral em comum.[6]
Os artrópodes habitam praticamente todos os tipos de ambientes no planeta, sejam eles aquáticos ou terrestres. Mesmo nos lugares mais inóspitos e sob temperaturas baixíssimas, como nas geleiras da Antártida, é possível encontrar a presença dos artrópodes.[7] Alguns dentre a classe dos insetos, representam os únicos invertebrados que possuem a capacidade de voar. Também se encontram alguns que são parasitas e outros que apresentam características simbióticas. Muitos destes animais estão diretamente ligados ao homem, seja por serem utilizados como alimento, como também por causarem prejuízos na saúde e na agricultura.
Os artrópodes têm apêndices articulados; o corpo segmentado, envolvido num exoesqueleto de quitina (números da imagem acima). Os apêndices estão especializados para a alimentação, para a percepção sensorial, para defesa e para locomoção. São estas "pernas articuladas" que dão o nome ao filo e que o separam dos filos mais próximos, os Onychophora e os Tardigrada.[4] O exoesqueleto é uma camada de cutícula quitinosa que reveste externamente todo o corpo dos artrópodes. Ele apresenta placas articuladas e contínuas. A presença de fenol na placa a deixa mais rígida e com a cor mais escura.
Eles são animais metamerizados, isto é, têm corpo segmentado, mas sua metameria não é tão evidente como a dos anelídeos; isso porque sua metameria heterônoma: os metâmeros (segmentos) diferenciam-se durante o seu desenvolvimento, alguns deles fundindo-se para a formação de tagmas que, como nos insetos, são tipicamente a cabeça, o tórax e o abdómen.
Entre as diferentes classes de artrópodes há casos em que dois ou mais tagmas se unem formando uma única peça como é o caso de certos grupos de crustáceos em que os tagmas cabeça e tórax se unem formando o cefalotórax e nos quilópodes e diplópodes em que o tórax se une com o abdômen formando o tronco. No subfilo Chelicerata os tagmas denominam-se prossoma (que corresponde ao cefalotórax) e opistossoma (que corresponde ao abdômen)
O primeiro segmento da cabeça é denominado ácron e normalmente suporta os olhos, que podem ser simples ou compostos. O último segmento do abdômen é terminado pelo télson. Cada segmento contém, pelo menos primitivamente, um par de apêndices.
Para poderem crescer, os artrópodes têm de se desfazer do exosqueleto "apertado" e formar um novo, num processo designado muda ou ecdise(é uma troca periódica do exoesqueleto, sendo o tempo e a quantidade de trocas variáveis de acordo com a espécie e as condições ambientais). O ponto de começo da ecdise é na linha de muda.
Por fazerem mudas, eles fazem parte do clado Ecdysozoa, que é um dos maiores grupos do reino animal, incluindo ainda os nematódeos, os Nematomorpha, os Tardigrada, os Onychophora, os Loricifera, os Priapulida e os Cephalorhyncha.
Diferentemente de anelídeos e moluscos, a excreção dos artrópodes é realizada por Túbulos de Malpighi, ou por glândulas especializadas, e não por nefrídeos.
Muitos grupos de artrópodes respiram por um sistema de traqueias, túbulos que abrem para o exterior através de poros na cutícula chamados espiráculos, e que se estendem por todo o corpo, promovendo a troca de gases. Nos aracnídeos, existe também as chamadas filotraqueias ou pulmões foliáceos. Crustáceos respiram por brânquias e insetos aquáticos têm brânquias ligadas ao sistema de traqueias.
O sistema circulatório dos artrópodes consiste numa bateria de corações que se dispõem ao longo do corpo e que bombeiam a hemolinfa (o "sangue" destes animais na maior parte das vezes não contém hemoglobina, baseada em ferro, mas sim hemocianina, baseada em cobre), que se encontra banhando os tecidos. Uma das diferenças da hemoglobina dos vertebrados para a hemocianina é o fato de esta ser um pigmento azulado,[8] pois em vez de ferro, possui cobre em seu princípio ativo, e ao reagir com o oxigênio, fica azulado, e quando livre do oxigênio, fica incolor. Nos insetos, os pigmentos respiratórios estão ausentes.
O sistema nervoso, de um modo geral, é como o dos anelídeos: gânglios cerebroides anterior dorsal, seguido de um par de nervos ventrais, com um par de gânglios por segmento. Porém nos artrópodes existe um alto grau de cefalização, o cérebro é formado pela fusão dos gânglios cerebroides, e tem três regiões distinguidas: o protocérebro, o deuterocérebro e o tritocérebro.
Os artrópodes são geralmente dioicos, com fecundação interna, utilizando de apêndices modificados para transferência de espermatozóides.
Embora alguns artrópodes tenham um padrão de desenvolvimento embrionário semelhante a dos anelídeos, na maior parte os ovos dos artrópodes são ricos em alimento e centrolécitos, a albummina, que fica no centro do ovo, deixando na periferia citoplasma sem albumina. Ocorre a clivagem superficial do ovo e o embrião desenvolve-se na periferia do ovo.
Os artrópodes são protostômios e possuem um celoma reduzido a um espaço à volta dos órgãos da reprodução e da excreção.
O Filo Arthropoda, são invertebrados protostomados do clado Ecdysozoa e constituem o filo mais biodiverso ocupando os uma vasta érea do nosso planeta, apresentando-se como um dos principais componentes de biomassa.[10] Assim como apresentam uma vasta gama de comportamentos alimentares. sua diversidade e grande distribuição levam naturalmente a interações significativas com o homem, com doenças vetoriais, pragas agrícolas, polinizadores, fontes de alimentos dentre outros[11]
Origem e evolução
O sucesso do Arthropoda é facilmente rastreado às vantagens significativas e diversas fornecidas por três características: apêndices articulados, um corpo metamérico (segmentado) geralmente dividido em segmentos fundidos (tagmata), e um exoesqueleto rígido composto principalmente por quitina.
Os dados evolutivos de Arthropoda já foram revistos por vezes.[12][13] Porém, com o advento de novas técnicas moleculares foi possível uma escala filogenômica mais completa.[12] Registros fosseis de Arthropoda, pertencente ao clado Ecdysozoa, são datados desde a radiação do terceiro estágio do Cambriano[13] que descrevem sequencias de eventos que levaram inclusive a biomineralização,[14] com de vestígios de Rusophycus e traços locomotores atribuídos a Diplichnitas,[14] porém a datação molecular sugere que os artrópodes tiveram sua origem e diversificação inicial no Ediacarano[12] e para Euarthropoda (Chelicerata + Mandibulata) sugere uma evolução inteiramente cambriana para este filo[14]
Estudos filogenéticos moleculares mostram a evolução da linhagem do dos artrópodes durante o Cambriano desde 518 milhões de anos atrás. A linhagem inclui grupos não biomineralizados, como Radiodonta que fornecem informações sobre a construção passo a passo da morfologia dos euartrópodes, incluindo o exoesqueleto, membros biramosos, segmentação e estruturas cefálicas.[14] e grande parte de sua diversidade foi originada de variações de um plano corporal como resultante de mudanças adaptativas no genoma[11]
As relações dos principais clados de artrópodes há muito são controversas, porém se sabe que os onicóforos ou os tardígrados são os parentes mais próximos dos artrópodes.[13] A base da árvore do panartrópode é ocupada por Onychophora, Tardigrada e uma série de lobópodes cambrianos.[14] Estudos paleontológicos dispõe panartrópoda (Onychophora, Tardigrada e Arthropoda) como sendo monofiléticos, já que compartilham apêndices segmentais e características morfológicas semelhantes,[15] e um RNAm que é sugerido como um caráter derivado compartilhado[9]
A linhagem de caule dos euartrópodes retrata um claro cenário de aquisição de caráter, desde a condição basal de um corpo anulado com membros lobópodes até níveis crescentes de artropodização[14]
Caráter especial para evolução
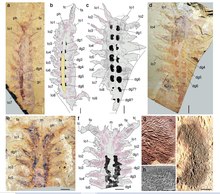
Diversos estudos focam seus esforços na evolução de caráteres morfológicos externos que foram importantes no processo de irradiação dos artrópodes. Porém, análises mais detalhadas evidenciou que os sistemas internos, como por exemplo o sistema digestório, podem ter contribuído para esse sucesso. Essas estruturas teriam aumentado a eficiência no processamento de alimentos e manteria o nível de energia que esses organismos demandavam para os seus estilos de vida ativo, móvel e predatório. Essa inovação pode ter contribuído para o sucesso evolutivo dos artrópodes, permitindo explorar mais recursos alimentares e modos de vida.[16]
Classificação filogenética
Atualmente há um consenso de que o filo Arthropoda é monofilético (com base em evidências morfológicas, de desenvolvimento e moleculares) e que deve ter existido um ancestral comum que originou os Arthropoda, Tardigrada e Onycophora. Esse grupo é chamado de Panarthropoda e suas principais sinapomorfias são a presença de um exoesqueleto contendo quitina, sem colágeno, que sofre mudas, perda de cílios e redução do celoma. Um grupo monofilético é caracterizado por incluir o ancestral em comum mais recente do grupo e todos os descendentes desse ancestral.
Além dessas características, a monofilia de Arthopoda vem da presença compartilhada de pernas compostas por podômeros esclerotizados separados por artrodiais membranas, músculos que se fixam nos tendões intersegmentais, olhos compostos nos quais novos elementos oculares são adicionados em uma zona de proliferação nas laterais do campo ocular em desenvolvimento, e a presença de dois neurópilos ópticos.
Onychophora é tradicionalmente reconhecido como o grupo irmão de Arthropoda. A evidência que apoia essa relação deriva mais obviamente do sistema circulatório hemocélico aberto, com um coração dorsal apresentando óstios segmentares em ambos os grupos. Artrópodes e onicóforos compartilham a musculatura segmentar da perna (musculatura que é distinta em tardígrados), possuem nefrídios ou derivados nefridiais que surgem das paredes das cavidades celômicas e os onicóforos assim como os artrópodes possuem hemocianina. Essa relação entre os dois grupos é fortemente apoiada em análises amplamente amostradas de marcas de sequência expressa.
Táxons
Mandibulata é um grupo de artrópodes com mandíbulas como parte principal da boca do adulto. Dentre os representantes desse grupo estão Myriapoda e Pancrustacea (Crustacea + Hexapoda). A monofilia de Mandibulata é sustentado por uma série de caracteres compartilhado.
Outros táxons fósseis exibem uma mistura de caracteres de crustáceos, miriápodes e hexápodes, em particular os Tanazios silurianos. Originalmente referido aos crustáceos, os apêndices da cabeça foram subsequentemente reinterpretados como incluindo filamentos frontais. A posição de Tanazios no Mandibulata é ambígua, mas sua combinação de caracteres de crustáceos e miriápodes (por exemplo, tagmosis em uma cabeça e tronco homônomo) sugere sua relevância para rastrear as divisões basais entre as linhagens de mandibulados.[13]
Uma hipótese mais antiga foi do grupo Paradoxopoda, que uniu Myriapoda com Chelicerata ao invés de com outros mandibulados (crustáceos e hexápodes). São várias as características compartilhadas entre os dois grupos que sustentam a hipótese, dentre elas o padrão de neurogênese observado[9]
Estudos recentes apoiam a monofilia de Chelicerata como o grupo irmão de Mandibulata, em linha com a evidência anatômica de artrópodes mandibulares como um grupo natural. Novos caracteres de mudanças genômicas raras adicionam mais suporte ao Mandibulata; miriápodes compartilham dois supostamente novos microRNAs (genes reguladores não codificadores) com crustáceos e hexápodes que não são compartilhados com queliceratos[15]
Chelicerata é um grupo de artrópodes que possuem como características um corpo dividido em cefalotórax (prossomo) e abdômen (opistossomo), um par de quelíceras, um par de pedipalpos e quatro pares de pernas locomotoras. Os quelicerados existentes foram tradicionalmente divididos em Pycnogonida, que se refere às aranhas-do-mar, e Euchelicerata que tem como integrantes organismos de ordens conhecidas: Aranae (aranhas), Scorpionida (escorpiões), Opiliones (opiliões) e Acari (ácaros e carrapatos). Além destes, Xiphosuria (carangueijos-ferradura), também estão incluídos nesse grupo[9]
Outra hipótese também merece atenção, que relaciona Pycnogonida como grupo irmão de todos os outros artrópodes e não um grupo irmão de Euchelicerata.[12] Essa hipótese, chamada Cormogonida (nome que se refere aos gonóporos situados no tronco, ao contrário das pernas), foi por bastante tempo aceita e defendida por diversos autores, porém hoje classificar por meio de análises morfológicas e moleculares Pycnogonida e Euchelicerata como grupo irmãos e integrantes de Chelicerata faz mais sentido Características como a presença de quelíceras sustentam esse posicionamento[12][13]
Características típicas, como olhos compostos, tronco e mebros artropodizados e apêndices da cabeça especializados, foram adquiridos gradualmente e, quando excluídos do conjunto de dados, junto com fósseis, a ideia de Cormogonida é novamente recuperada.[12]
O registro fóssil de Chelicerata se estende até o início do Cambriano, e é exemplificado por Wisangocaris barbarahardyae do Sul da Austrália e o estreitamente aliado Sanctacaris uncata do Burgess Shale. Em análises filogenéticas, eles estão mais intimamente ligados a Euchelicerata do que Pycnogonida. Muitos quelicerados do Paleozóico têm uma aparência superficial de caranguejo-ferradura e foram agrupados como Synziphosurina[9]
Os miriápodes são divididos em quatro grupos principais Chilopoda, Diplopoda, Symphyla e Pauropoda.[9] O registro fóssil de miriápodes não é amostrado até meados do Siluriano[13] possivelmente subaquático.[9] Miriápodes e os hexápodes já foram considerados paralifeticos, formando um clado conhecido como Atelocerata, porém, essa hipótese se baseava tão somente na morfologia,[10][13][15] sendo rejeitada diante de análises moleculares[13]
Atualmente, Myriapoda é monofilético, com seus dois clados principais sendo Chilopoda e Progoneata,[12][15] sendo sustentado por evidencias moleculares.[9] Todavia, as relações internas desse grupo ainda estão longe de serem resolvidas se tratando de um grande conflito entre a filogenia molecular e morfológica. A morfologia tem apoiado uma relação de grupo irmão entre milípedes e paurópodes (clado chamado Dignatha), e também sugeriram que os sínfilanos estão intimamente relacionados ao grupo diplópode-paurópode (esses três grupos juntos constituem Progoneata). Já a filogenia molecular, aponta vários conjuntos de dados moleculares contradizem o clado Dignatha, em favor de um grupo sinfílano-paurópode chamado Edafopoda[9]
Para Crustacea podem ser citadas como apomorfia, a fusão dos olhos naupliares e dois órgãos segmentares, as glândulas antenais e maxilares, colocando-os provavelmente como não monofilético.[13] A parafilia de crustáceos em relação a Hexapoda era frequentemente questionada e proposto como grupo irmão de Hexapoda, possivelmente, Branchiopoda, Copepoda ou Branchiopoda,[13] mas, com testes moleculares foi possível traçar o parentesco entre Hexapoda, Branchiopoda e Remipedia unido-os em um clado denominado Allotriocarida,[12] com Remipedia sendo o mais aceito até o momento.[15] Do ponto de vista da morfologia, a maioria dos argumentos a favor da monofilia do grupo Pancrustacea, provém de estudos do sistema nervoso[13] através de estudos com uma abordagem chamada de neuropilogenia[15]
Para Hexapoda, seu registro fóssil conhecido começa no Devoniano,[13] com haxápoda nidificando dentro do clado de Crustacea e sendo monofilético.[12] Estudos recentes sobre a anatomia e o desenvolvimento do sistema nervoso central (SNC) dos artrópodes identificaram muitos outros elementos neurológicos, que parecem singulares a Hexapoda + Crustacea. Nos hexápodes e nos crustáceos, o desenvolvimento do SNC começa com a delaminação das células aumentadas, conhecidas como neuroblastos, da ectoderme (neuroectoderme) de cada segmento.[10] A relação de parentesco entre Hexapoda e Crustacea se dá devidos aos remípedes.[12] Os indivíduos agrupados dentro de Pancrustácea apresentam a mais diversa disparidade nos planos corporais[9]
Estima-se que existam cerca de dois milhões de espécies de organismos já descritos de todos os habitats. Organismos que habitam o ambiente terrestre têm sido amostrados em maior quantidade do que os aquáticos. Entretanto, os mares são muito mais ricos em níveis taxonômicos.[17] Dentre os diversos grupos de organismos que vivem no ambiente marinho destacam-se os artrópodes.
Os artrópodes constituem cerca de 80% de todas as espécies de animais conhecidos. A enorme capacidade adaptativa desde grupo, fez com que estes conseguissem habitar em praticamente todos os ambientes, dentre eles o marinho.[18]
Os Trilobitomorpha foram artrópodes marinhos de grande abundância no paleozóico, hoje já extintos. Eles ocuparam uma diversidade muito grande de habitats, desde recifes até oceanos profundos, bentônicos ou livre-natantes. Possuíam diversos hábitos alimentares, com indivíduos herbívoros, filtradores, necrófagos e até predadores.[19] A diversidade gigantesca desse grupo pode ser bem exemplificada em uma espécie chamada Agnostus pisiformis. Neste o escudo cefálico e o pigídio eram utilizados de forma parecida a das valvas de um ostrácode. Além do mais, levando em conta os hábitos alimentares, já foi levantado que este poderia atuar como parasita.[20]
Os Hexapoda por sua vez compreendem o grupo com maior número de espécies. Entretanto, somente alguns poucos representantes são marinhos,[21] sendo que 20 ordens têm representantes marinhos. Existem representantes dos Coleoptera, Hemiptera e Diptera, mas os únicos espécimes de insetos que vivem em mar aberto são os do gênero Halobates, embora eles vivam na interface ar-mar. Esses insetos são conhecidos por se deslocarem por sobre a água. Das 46 espécies pertencentes a esse gênero somente 5 são oceânicas, ou seja, vivem em mar aberto, as demais vivem em proximidade ao litoral.[22] Os demais insetos marinhos são conhecidos assim por possuírem pelo menos um dos estágios de seu desenvolvimento em ambiente marinho. Entre os exemplos estão as ninfas de alguns gêneros de Odonata, popularmente conhecidas como libélulas, as lagartas de espécies de Lepidoptera, Orthoptera (gafanhotos) e algumas poucas espécies de Hymenoptera, formigas.[21]
O subfilo Chelicerata constitui um grupo que inclui as aranhas, escorpiões, ácaros, carrapatos, opiliões(Classe Arachnida), e ainda outras duas pequenas classes (Merostomata e Pycnogonida). Dentro da classe Arachnida, existe a ordem Acari, representado pelos ácaros. Estes apesar de viverem predominantemente em ambientes terrestres ocuparam com sucesso os ambientes aquáticos. Contudo, a classe Merostomata e Pycnogonida são mais representativos no ambiente marinho.[18]
Os Merostomata, conhecidos popularmente como caranguejos-ferradura ou límulos (da ordem Xiphosura, que inclui espécies viventes), são quelicerados aquáticos caracterizados por cinco ou seis pares de apêndices abdominais modificados como brânquias e um telson articulado em forma de esporão. Podem ser encontrados na costa noroeste do Atlântico, no Golfo do México, ao longo das costas asiáticas do Japão e Coréia até as Índias Orientais e as Filipinas. Estes vivem em águas rasas sobre fundos macios, migrando para a superfície, no período de acasalamento.[18] Os Euripterida, também pertencentes à classe Merostomata, são comumente conhecidos como escorpiões-marinhos, e já foram extintos. Eram bastante diversificados no Paleozoico, possuindo cerca de 200 espécies. Estes eram predominantemente aquáticos, atuando como grandes predadores de sua época, podendo atingir tamanhos próximos a 100 cm. São os maiores artrópodes já conhecidos, tendo exemplares que atingiram até 2,5 metros de comprimento.[23] Os Pycnogonida constituem um grupo de cerca de 1.000 espécies marinhas, conhecidos popularmente como aranhas-do-mar. Vivem estritamente nos oceanos, desde o ártico e a Antártida até os trópicos, em diversas formas litorâneas, como espécies que vivem em regiões abissais.[18]
O subfilo Crustacea, cuja etimologia vem do latim crusta = crosta, concha, superfície dura + aceus = pertence a, é um grupo representado por animais como caranguejos, siris e camarões. É o único subfilo de artrópodes cujos membros são primariamente aquáticos, sendo a maioria dos grupos marinho. Estudos mostram que muitas espécies são usadas como bioindicadores em vários sistemas aquáticos, pois tem a capacidade de bioacumulação de contaminantes metálicos; são sensíveis a variações de fatores físico-químicos e ambientais. Considerando os estágios de desenvolvimento dos crustáceos, normalmente possuem uma fase larval inicial e a fase adulta. A fase larval é planctônica livre-natante na maioria das espécies marinhas enquanto que na fase adulta são, em geral, bentônicos.[18]
Dentro do subfilo Crustacea, destaca-se a classe Malacostraca, sendo esta a que possui maior número de espécies conhecidas, totalizando 22.651. Entre a grande diversidade dos Malacostraca, os representantes mais comuns fazem parte da ordem Decapoda, com cerca de 10.000 espécies descritas. A maioria dos decápodos é marinha, porém alguns invadiram o ambiente de água doce.[18]
Os crustáceos sempre foram bastante diversificados, compondo grande parte da fauna marinha do período Ordoviciano e da era Mesozoica e Cenozoica, nestas últimas representaram a grande maioria da fauna de artrópodes. Sabe-se que dos 5800 gêneros de artrópodes marinhos destes períodos, 2600 destes são crustáceos, perdendo somente em número para os trilobitas. Os três grupos de artrópodes mais representativos no registro fóssil são: ostrácodes (o grupo com maior registro fóssil entre os crustáceos), cirripédios e malacóstracas (estes foram muito representativos no mesozoico, isso se deve a grande diversificação dos decápodes). Importantes grupos de artrópodes marinhos atuais, como por exemplo os copépodos, o grupo mais abundante em ecossistemas marinhos pelágicos - possuem um registro fóssil bastante pequeno.[24]
Na fauna dos solos são encontrados os Artrópodes do subfilo Hexapoda, que são os insetos, principalmente com as classes Coleoptera (besouros) e Hymenoptera (formigas), os do subfilo Myriapoda (Diplopoda, Chilopoda, Pauropoda e Symphyla) e os da classe Arachnida (aracnídeos).
São características a presença de peças bucais (mandíbulas, maxilas, quelíceras, pedipalpos) que estão adaptadas para a mastigação e a sucção. A respiração é feita através de traqueias ou filotraqueias, sistema nervoso ganglionar. Os órgãos dos sentidos são bastante especializados: Antenas que são quimiorreceptoras, olhos que podem ser simples ou compostos, alguns possuem órgãos gustativos e auditivos.
Estes animais são encontrados, em sua maioria, na camada mais externa do solo, conhecida como serapilheira ou liteira (espécies edáficas). As espécies que habitam os espaços dos horizontes minerais abaixo da camada de serapilheira são conhecidas como euedáficas.
Em 2010, foram encontradas quatro novas espécies de colêmbolos (pequenos artrópodes com seis pernas e parentes próximos dos insetos) na caverna mais profunda do mundo, com mais de dois quilômetros abaixo do solo, Krubera-Voronja (Cáucaso). O artrópode proveniente das maiores profundidades até hoje descrito foi coletado a mais de 1980 m da superfície do solo e nomeado Plutomurus ortobalaganensis. As outras três espécies encontradas são: Anurida stereoodorata, Deuteraphorura kruberaensis e Schaefferia profundísima.
Os quatro artrópodes descritos podem ajudar a perceber como os animais se adaptaram ao longo de milhares de anos, para se adaptar à vida subterrânea. Esses artrópodes não possuem pigmentação no corpo, nem nos olhos, e uma das espécies orienta-se no escuro por antenas capazes de captar informação sobre o seu meio ambiente. Eles se alimentam de um fungo que cresce no solo das cavernas, contribuindo para a reciclagem da matéria orgânica. [25]
No Pantanal mato-grossense foram feitas coletas na região de Pirizal por Figueiredo (2007), Oliveira-Silva (2007), Tissiani (2009) e Anjos (2009), e em áreas vizinhas por Pinho (2003), Castilho (2005) e Battirola (2007). Nessas áreas do Pantanal há uma ocorrência e desenvolvimento de migrações horizontais e verticais por parte dos artrópodes terrestres, que são estratégias de sobrevivência durante períodos de cheia. E nas coletas acontecidas, em 2007, foram coletados 29.321 artrópodes distribuídos entre insetos (Hexapoda), aracnídeos (Arachnida) e miriápodes (Myriapoda). Os insetos somaram 82,2% do total de artrópodes coletados, e os grupos predominantes, ou seja, que mais se locomoveram no solo foram os colêmbolos (Collembola), besouros e himenópteros (Hymenoptera), a maioria formigas, correspondendo, respectivamente, a 39,5%, 20,4% e 17,3% do total de insetos amostrados na grade do Pirizal. Os aracnídeos compreenderam 17, 6% dos artrópodes e foram representados por 15,8% de ácaros (Acari) e 0,9% de aranhas.
Alguns táxons de miriápodes comuns na Amazônia, como sínfilos (Symphyla) e paurópodes (Pauropoda), são pouco encontrados no Pantanal e outros como diplópodes (Diplopoda), conhecidos como piolhos-de-cobra ou centopeia, e quilópodes ou lacraias (Chilopoda), apresentaram baixa densidade nesta região.
Os artrópodes constituem o grupo mais abundante dos macroinvertebrados aquáticos. Os insetos e os crustáceos representam grande parte dos artrópodes encontrados em rios.Os principais problemas que os insetos aquáticos apresentam no ambiente aquático são o suprimento de O2 deficitário e o movimento da água dificulta permanência num só ponto e movimentar-se dentro da água é difícil. Os insetos tiveram que se adaptar ao meio aquático através de modificações no sistema respiratório como:
Os insetos também apresentaram outras adaptações para o ambiente aquático, como as adaptações lóticas, onde os insetos que vivem nestes sistemas tendem a ser dorsoveltralmente achatados, algumas vezes com pernas lateralmente projetadas. Insetos aquáticos podem ser bioindicadores no meio ambiente e conseguem se desenvolver em ambiente aquático temporário através da deposição de ovos resistentes a desidratação e a um ciclo de vida rápido.
Os aeglídeos (caranguejos) constituem um grupo ecológico de decapodos anomuros que vive embaixo de pedras e folhas do substrato dos cursos de águas continentais como rios, riachos, lagos e cavernas. São importantes nas cadeias tróficas dos cursos d’água servindo de alimento para peixes, aves e jacarés. São considerados eficazes predadores de larvas aquáticas de insetos, especialmente dos simulídeos hematófagos, popularmente conhecidos como borrachudos. Os Cladoceros ( pulgas d’água ) e Copepodos, importantes na composição da fauna de invertebrados aquáticos, podem ser encontrados em curso d’água mais lentos, geralmente nas margens associados a macrofitas aquáticas, algumas espécies podem ser parasitas de peixes.
Entre os Chelicerata, a ordem Acari (ácaros) tem o maior número de representantes associados a ambientes aquáticos. Os adultos atingem geralmente um tamanho de 0,5 mm até 2 mm. Ocorrem em todos os corpos d’água, em cavidades de arvores, nascentes quentes e cascatas.
Cladocera são os filópodes mais bem sucedidos; a maioria é de água doce, são de pequeno porte, conhecidos também como pulgas–d'água. A maioria das espécies vive preferencialmente na região litorânea de ecossistemas lacustres. Nem todos os cladóceros de água doce são planctônicos (por exemplo, a família Macrothricidae não tem representante no plâncton). Os cladóceros planctônicos mais frequentes em água doce pertencem às famílias: Sididae, Daphniidae, Bosminidae e Chydoridae, sendo que os representantes da última família frequentemente são encontrados fazendo parte dos bentos.
Copépodos formam com os cladóceros os organismos mais típicos do chamado plâncton de rede. Mesmo sem equipamento, podem ser facilmente reconhecidos pelo seu corpo alongado. Os copépodes habitam os mais diferentes ambientes aquáticos, tais como água doce, salobra, salgada e mesmo terras úmidas. As formas planctônicas são, na grande maioria, de pequeno tamanho. As principais ordens de copépodes com representantes no plâncton de água doce são: Calanoida, que são essencialmente planctônicas, Cyclopoida, que têm representantes planctônicos e bentônicos, e Harpacticoida cuja maioria é bentônica.
Anostraca (artêmias) são relativamente pequenos e habita regiões interiores, mas não necessariamente só ambientes de água doce. As 200 espécies conhecidas de anóstracos ocorrem em lagos salgados e lagoas e alagados temporários de ambientes úmidos.
Notostraca contém apenas 10 espécies bentônicas pertencentes aos gêneros Triops e Lepidurus, encontrados e águas interiores, calmas e livre de peixes.
Ostrácodes são microcrustáceos diminutos encontrados na maioria dos hábitats tanto de água doce como marinhas. Na água doce ocorrem em lagos, lagoas e fontes termais, são espécies cavadoras e intersticiais, podendo também viver em algas e plantas aquáticas.
Decápodes são muito frequentes em águas continentais brasileiras; são representados por várias espécies do gênero: Euryrhynchus, Macrobrachium, Palaernonetes e Pseudopalaemon (KENSLEY & WALKER, 1992); os decápodes representam os chamados camarões de água doce.
Os Misidáceos são peracáridos pequenos, encontrados em hábitats de água doce, salobro e marinho, têm maior importância em lagos de regiões temperadas, uma vez que habitam preferencialmente águas frias.
Anfípodes são pequenos frequentemente encontrados em grande número e alta diversidade em ambientes aquáticos; os Gammaridea são os únicos anfípodes em hábitats de água doce, nos quais são bentônicos e ecologicamente importantes, em especial na vegetação.
Os Isópodes são, na sua maioria, terrestres e marinhas e somente poucas espécies são de águas continentais. Muito pouco é conhecido sobre a ecologia e o papel destes organismos na dinâmica de águas continentais.
Os insetos planctônicos são muito raros. Somente alguns grupos de Diptera na fase larval têm representantes no plâncton, dentre estes a larva de Chaoborus (=Comethma) é a mais importante. O gênero Chaoborus da família Chaoboridae é encontrado em todos os tipos de ambientes lacustres, desde pequenas lagoas temporárias, até grandes lagos permanentes. Uma de suas características mais importantes é a alternância diária de hábitat: à noite são planctônicos e durante o dia são bentônicos. Grande número de insetos são aquáticos ou têm parte de seu ciclo de vida na água. Em ambos os casos, a grande maioria das larvas destes organismos é bentônica. Dentre os grupos de insetos que têm representantes nos bentos continentais (principalmente sob a forma larval), destacam-se os dípteros, ephemerópteros, plecópteros, odonata, hemípteros, coleópteros, neurópteros, tricópteros e lepidópteros. De todos estes grupos, os dípteros são os que têm maior importância, visto que os seus principais representantes aquáticos quironomídeos e caoborídeos, são encontrados, via de regra, em grande número.
Araneae – São encontradas na maioria dos ambientes terrestres, mas existe uma única espécie que vive em água doce, abaixo da superfície, a Argyroneta aquática (Argyronetidae) da Eurásia, é cursória; vive e caça abaixo da superfície da água.
Artrópodes (Arthropoda, do grego arthros (ἄρθρον), articulado e podos (ποδός), pés) são um filo de animais invertebrados que possuem exoesqueleto rígido e vários pares de apêndices articulados, cujo número varia de acordo com a classe.
Compõem o maior filo de animais existentes, representados por animais como os gafanhotos (insetos), as aranhas (aracnídeos), os caranguejos (crustáceos), as centopeias (quilópodes) e os piolhos-de-cobra (diplópodes). Têm cerca de um milhão de espécies descritas, e estima-se que os representantes deste filo equivalem a cerca de 84% de todas as espécies de animais conhecidas pelo homem. Possuem uma ampla gama de cores e formatos, e no que diz respeito ao tamanho, alguns vão desde as formas microscópicas, como no plâncton (com menos de 1/4 de milímetro), até crustáceos com mais de 3 metros de espessura.
Sua existência é datada nos registros fósseis desde o período Cambriano (há cerca de 542 a 488 milhões de anos), onde criaturas como as Trilobitas eram encontradas em abundância nos oceanos. Algumas teorias sobre a origem deste filo sustentam que os ancestrais dos artrópodes podem ter sido os anelídeos (vermes de corpo segmentado em anéis) ou de algum outro ancestral em comum.
Os artrópodes habitam praticamente todos os tipos de ambientes no planeta, sejam eles aquáticos ou terrestres. Mesmo nos lugares mais inóspitos e sob temperaturas baixíssimas, como nas geleiras da Antártida, é possível encontrar a presença dos artrópodes. Alguns dentre a classe dos insetos, representam os únicos invertebrados que possuem a capacidade de voar. Também se encontram alguns que são parasitas e outros que apresentam características simbióticas. Muitos destes animais estão diretamente ligados ao homem, seja por serem utilizados como alimento, como também por causarem prejuízos na saúde e na agricultura.
Artropodele (din greacă arthron, "articulație" și podos, "picior") sunt animale nevertebrate, ce au membrele articulate. Încrengătura include insecte, milipede, centipede, arahnide, crustacee etc. Artropodele sunt organisme cu un exoschelet (schelet extern), corp segmentat și membre (apendice) anexate de cap și torace. Artropodelor sunt caracterizate cuticulele chitinizate, la crustacee cuticulele sunt mineralizate cu carbonat de calciu. Cuticula este rigidă și nu se întinde, de aceea procesul de creștere e însoțit de năpârlirea periodică. Primele membre participă la apucarea hrănii și fărămițarea. Corpul este alcătuit din segmente, fiecare având câte o pereche de membre. În prezent artropodele constiuie cel mai variat grup de animale. Peste 80% din toate speciile descrise revin artropodelor. Spre deosebire de alte nevertebrate, organele de simț ale atropodelor sunt foarte dezvoltate, au inimă ce pompează sângele, iar cavitatea internă constituie hemocelul. Artropodele sunt unisexuate, se reproduc doar pe cale sexuată (sunt și excepții, unii se înmulțesc prin partenogeneză). Toate speciile sunt ovipare, numai scorpionii pot fi clasificați ca vivipari. Se dezvoltă prin metamorfoză incompletă, completă sau direct.
Ele populează cele mai diverse ecosisteme: terestre, marine, aeriene.
Artropodele au o mare importanță pentru om. Unele fac parte din dieta omului, sunt polenizatoare ale plantelor angiosperme. Altele ducând un mod de viață parazit, generează boli grave, sau aduc pagube agriculturii.
Artropodele sunt nevertebrate cu corpul segmentat și membre articulate[2]. Membrele la fel ca și corpul sunt acoperite cu cuticulă, care este alcătuită în principal din α-chitină, un derivat al glucozei[3]. Pe lângă aceste trăsături, artropodele se caraterizează prin simetrie bilaterală, posedă un creier dorsal de care e conectat lanțul nervos ventral ce străbate corpul longitudinal[4].
Toate artropodele în stadiul de embrion sunt segmentate, constituite dintr-o serie de module ce se repetă. Numărul de segmente variază în dependență de clasă și ordin. De exemplu la unele chilopode numărul lor ajunge până la 181, iar la unii acarieni corpul este nesegmentat[5]. Fiecărui segment corespunde câte o pereche de apendice (membre). La majoritatea artropodelor actuale segmentele sunt grupate în regiuni, numite și tagme[5]. De asemenea, și modul de repartizare a tagmelor este divers. La chelicerate corpul este divizat în două tagme: prosomă (cefalotorace) și opistosomă (abdomen). Corpul insectelor și crustaceelor cuprinde cap, torace și abdomen[6]. Iar la miriapode corpul este alcătuit din cap și trunchiul divizat în segmente aproximativ egale, fără vreo delimitare în torace și abdomen. În segmentarea corpului, artropodele au unele caracteristici, anterior gurii se află acronul și posterior anusului – telsonul[5]. Acestea nu fac parte din segmentarea descrisă anterior, ele sunt lipsite de membre. Pe acron sunt plasați ochii.
Membrele (apendice) artropodelor sunt biramate. Ramura superioară externă îndeplinește rolul branhiilor, iar cea inferioară internă servește drept membre locomotoare. Membrele artropodelor au structură deosebită și îndeplinesc funcții diferite: tactilă (antene)[6], fărămițarea hrănii (chelicere), apucarea și reținerea prăzii (pedipalpii)[7], sunt implicate în acuplare (gonopodele). Numărul membrelor locomotoare variază. Crustaceele au 5 perechi de picioare, arahnidele – 4 perechi, insectele – 3 perechi. Abdomenul, la cele mai multe dintre clase, este lipsit de membre.
Cuticula reprezintă exoscheletul acelular secretat de celulele epidermei[5]. Structura cuticulei lor variază în detalii, dar, în general, este compusă din trei straturi principale: epicuticulă – strat extern subțire și impermeabil pentru apă și care oferă o anumită protecție; exocuticulă – strat mediu mai gros, constă din chitină și proteine; endocuticulă – strat intern, la fel constă din chitină și proteine, dar este mai elastic[8]. Exocuticula și endocuticulă împreună se numesc procuticulă. Fiecare segment a corpului este învelită cu cuticulă rigidă. Regiunile de articulare a segmentelor și articolelor membrelor sunt acoperite cu o cuticulă flexibilă[5].
Cuticula crustaceelor acvatice este biomineralizat cu carbonatul de calciu extras din apă[9]. Unele crustacee terestre au capacitatea de a depozita substanțe minerale, deoarece în mediul terestru carbonatul de calciu dizolvat e greu de găsit. Împregnarea cuticulei cu carbonat de calciu are loc în exocuticulă și în stratul exterioar a endocuticulei[8]. Datorită biomineralizării cuticula crustaceelor este mai rezistentă la factorii mecanici externi[10].
Cuticula poate avea perișori aspri, excrescențe ale unor celule speciale din epidermă. Firele sunt de formă diferită și funcționează ca anexe. De exemplu, ele sunt folosite ca receptori pentru a detecta curenți de aer sau apă, sau ca senzori tactili, facilitează înotul, filtrează substanțele nutritive din apă etc[5].
Cuticula nu se poate întinde și împiedică creșterea animalului. Artropodele înlocuiesc periodic cu o cuticulă nouă pe cea veche, proces numit năpârlire. Năpârlirea are loc pe tot parcursul dezvoltării, până când artropodul ajunge la dimensiunea completă[11].
Debarasarea de vechea cuticulă are loc simultan cu formarea cuticulei noi. În faza inițială a năpârlirii, animalul încetează să se alimenteze și epiderma elaborează un lichid special pentru năpârlire, un amestec de enzime care ajută la dizolvarea endocuticulei (stratul intern al cuticulei). În acest fel, cuticula veche se va putea desprinde de corp. Această etapă începe atunci când epiderma a secretat noua epicuticulă, pentru a proteja cuticula nouă de enzime. Epicuticula nu poate fi dizolvată de enzime. Sub epicuticulă se realizează producerea celorlalte straturi noi: exocuticula și endocuticula. Când această etapă este completă, animalul își umflă corpul inspirând o cantitate mare de apă sau aer. Din cauza presiunii exercitate din interior, apare o fisură în cuticula veche de-a lungul punctelor slabe ale exocuticulei vechi[11]. Este nevoie de câteva minute pentru ca artropodul să scape de cuticula veche. După aceasta, cuticula nouă este moale, având nevoie de ceva timp ca să se întărească. În acest moment, când cuticula încă nu s-a întărit, are loc creșterea în dimensiuni a corpului. Unele artropode pompează apă sau aer, pentru ca la întărire cuticula să capete un aspect mai voluminos. Multe artropode consumă vechea cuticulă pentru a readuce în organism substanțele necesare[11].
Deoarece în timpul năpârlirii cuticula nouă este moale, artropodele sunt practic neprotejate și aproape imobilizate până la întărirea ei. Din numărul total de omoruri a artropodelor de către alți prădători, perioadei de năpârlire îi revine 80-90%[11].
Organele interne ale artropodelor, la fel, prezintă o anumită metamerie (dispunere segmentară). Cavitatea internă a corpului este ocupată de hemocel, în care se revarsă hemolimfa[12]. Celomul înconjoară inima și glandele sexuale.
Modul de trai al artropodelor este strâns legat de dezvoltarea sistemului nervos, de tip scalariform. El este reprezentat de creier și lanțul nervos ventral. Creierul este concentrat în cefalotorace sau cap, deasupra esofagului. El este constituit din 3 regiuni: protocerebron – inervează ochii, deutocerebron – inervează antenele și tritocerebron – inervează chelicerele sau mandibulele. Lanțul nervos ventral se află sub intestin, de-a lungul abdomenului. În fiecare segment al corpului lanțul nervos formează câte un ganglion. Ganglionii sunt conectați prin comisuri (fascicule, relativ groase, de nervi) și conective. Ganglionii toracici reprezintă o masă unică sub- sau cricumesofagiană. Numărul ganglionilor la diverse artropode este diferit[13].
Aparatul circulator al artropodelor este de tip deschis. Inima tubulară este situată în partea dorsală a abdomenului și opistosomei. Contracția inimii se realizează din posterior spre anterior, împingând hemolimfa spre cap. Inima se sprijină pe cuticulă cu ajutorul unor mușchi mici[12]. De la inimă pleacă una sau două ?aorte? ce se ramifică spre toate organele și țesuturile. Lichidul intern care circulă prin vase se numește hemolimfă. La chelicerate și unele crustacee oxigenul e transportat prin pigmenți, iar la insecte și miriapode prin trahei puternic ramificate. Hemolimfa se întoarce în inimă prin osteole laterale. Osteolele permit hemolimfei să intre în inimă, dar împiedică revenirea ei în corp. Pigmentul respirator, care transportă oxigenul este hemocianina la arahnide, centipede și la unele crustacee sau hemoglobina la insecte și unele crustacee. Hemocianina conține atomi de cupru de care se leagă oxigenul, iar hemoglobina atomi de fier. Spre deosebire de vertebrate, pigmenții sunt dizolvați în hemolimfă, și nu sunt incluși în componența unor celule speciale cum ar fi eritrocitele[12].
Organele respiratorii ale artropodelor sunt diverse. Crustaceele și unele chelicerate acvatice respiră prin branhii, altele prin plămâni (saci pulmonari) și trahei. Miriapodele și hexapodele respirația e realizată de trahei ramificate prin tot corpul, oxigenând direct țesuturile. Artropodele de dimensiuni foarte mici, în special cele acvatice, efectuează schimbul de gaze prin tegument, datorită difuziei[14].
Artropodele marine realizează excreția prin glande antenale, cele terestre au mai multe moduri de a elimina metaboliții, produsele metabolismului. Tuburile lui Malpighi sunt unice printre animale. Ele reprezintă tuburi care plutesc în cavitatea corpului și se deschid la granița dintre intestinul mediu și posterior. Majoritatea artropodelor terestre și marine au glande coxale, de asemenea niște tuburi. Pe lângă acestea, mai sunt și nefrocite, celule specializate în colectarea produselor excretoare. La unele, metaboliții se acumulează în cuticulă și sunt eliminați la năpârlire, împreună cu cuticula veche.
Artropodele se clasifică în:
Artropode cu chelicere
Reprezentanți: paianjeni; scorpioni; căpușe;
Artropode cu antene
Reprezentanți: crustacee; miriapode; insecte;
|date= (ajutor) |pages= și |page= (ajutor); |access-date= necesită |url= (ajutor)
Artropodele (din greacă arthron, "articulație" și podos, "picior") sunt animale nevertebrate, ce au membrele articulate. Încrengătura include insecte, milipede, centipede, arahnide, crustacee etc. Artropodele sunt organisme cu un exoschelet (schelet extern), corp segmentat și membre (apendice) anexate de cap și torace. Artropodelor sunt caracterizate cuticulele chitinizate, la crustacee cuticulele sunt mineralizate cu carbonat de calciu. Cuticula este rigidă și nu se întinde, de aceea procesul de creștere e însoțit de năpârlirea periodică. Primele membre participă la apucarea hrănii și fărămițarea. Corpul este alcătuit din segmente, fiecare având câte o pereche de membre. În prezent artropodele constiuie cel mai variat grup de animale. Peste 80% din toate speciile descrise revin artropodelor. Spre deosebire de alte nevertebrate, organele de simț ale atropodelor sunt foarte dezvoltate, au inimă ce pompează sângele, iar cavitatea internă constituie hemocelul. Artropodele sunt unisexuate, se reproduc doar pe cale sexuată (sunt și excepții, unii se înmulțesc prin partenogeneză). Toate speciile sunt ovipare, numai scorpionii pot fi clasificați ca vivipari. Se dezvoltă prin metamorfoză incompletă, completă sau direct.
Ele populează cele mai diverse ecosisteme: terestre, marine, aeriene.
Artropodele au o mare importanță pentru om. Unele fac parte din dieta omului, sunt polenizatoare ale plantelor angiosperme. Altele ducând un mod de viață parazit, generează boli grave, sau aduc pagube agriculturii.
Článkonožce (staršie: článkonohé[1]; lat. Arthropoda) sú najpočetnejší živočíšny kmeň. Je to obrovská, dosť starobylá a vnútorne veľmi diferencovaná skupina článkovaných živočíchov, ktoré fylogeneticky nadväzujú na Polychaeta. V procese fylogenézy však získali znaky, ktoré im umožnili osídliť všetky suchozemské i sladkovodné ekosystémy a vrátiť sa aj naspäť do mora, z ktorého vzišli. Všade, kde sa vyvíjali, dosiahli mimoriadnu mnohorakosť foriem a prekvapujúcu diverzitu (na súši tvoria podľa niektorých odhadov dokonca až 95 % živočíšnych druhov).
Telo sa skladá v najjednoduchších prípadoch (Myriapoda) z hlavy a takmer rovnakých článkov (segmentov), u väčšiny je však segmentácia výrazne heteronómna (články sa od seba líšia stavbou). Stavbu tela a telových príveskov odvodzujeme z hypotetického východiskového modelu, ktorý mal na každom segmente (okrem prvého článku akronu a posledného pygidia) 1 pár článkovaných, s takmer stopercentnou istotou dvojvetvových končatín. Tomuto modelu sú najbližšie trilobity, ale nie sú mu príliš vzdialené ani primitívne kôrovce Crustacea a viacnôžky Myriapoda; druhá skupina však už má končatiny druhotne jednovetvové. Telo vyšších článkonožcov sa primárne člení na tri oddiely: hlavu (caput), hruď (thorax) a bruško (abdomen). Končatiny sú u podstatnej väčšiny vyvinuté len na hrudi (sú to primárne kráčavé nohy), na hlave z nich vznikajú špecializované prívesky, a na brušku zanikajú alebo prekonávajú zložitú zmenu tvaru a funkcie – modifikáciu.
Telo je kryté chitínovou kutikulou, ktorú produkuje epidermis. Kutikula sa skladá z troch častí:
Hrúbka a mikroštruktúra týchto vrstiev a niektoré prímesové látky ovplyvňujú konečnú tvrdosť kutikuly – táto môže byť relatívne mäkká (Arachnoidea), ale aj pevná a tvrdá (mnohé skupiny hmyzu). Má viaceré veľmi výhodné vlastnosti – je veľmi odolná voči kyselinám i zásadám, zabraňuje dehydratácii a aj u silne „opancierovaných“ druhov je vždy dostatočne pružná. Má iba jednu nevýhodu a to že zabraňuje v raste. Preto sa článkonožce periodicky zvliekajú – väčšina len v larválnom štádiu, no niektoré po celý život. Proces zvliekania riadi hormón ekdyzón, ktorý produkujú párové predohrudné žľazy (glandulae protothoracales). Kutikula vždy obsahuje istú prímes CaCO3, čo ju robí pevnejšou. Článkonožce s mäkkou kutikulou ho majú prítomný len v stopách, kým u iných skupín pozorujeme silnú (väčšina Diplopoda) alebo veľmi silnú (väčšina Crustacea) inkrustáciu (prestúpenie) CaCO3. Sfarbenie kutikuly je predovšetkým pigmentové (chemické), u mnohých skupín sa však vyskytuje aj sfarbenie fyzikálne – vzniká interferenciou svetla na jemných, ale často zložitých mikroskulptúrach epikutikuly. Týmto spôsobom vznikajú predovšetkým rôzne, často veľmi nápadné kovové lesky hmyzu, zriedka i pavúkov. Pretože kutikula napriek svojej pružnosti umožňuje len veľmi obmedzený pohyb, musel proces sklerotizácie už od začiatku prebiehať paralelne so vznikom segmentácie. Výsledkom týchto fylogeneticky veľmi úspešných procesov je kutikulárny ektoskelet, zložený z mnohých obrúčok a štítkov, ktoré sú spoločne nazývané sklerity. Ektoskelet každého somitu má u fylogeneticky pôvodných typov tvar obrúčky – táto sa skladá z chrbtovej časti (tergitu) a brušnej časti (sternitu), ktoré sú spojené bočným (pleurálnym) blanitým väzom. Takúto stavbu však pozorujeme pomerne zriedka (Myriapoda, časť Crustacea), alebo iba na niektorých častiach tela (abdomen u šesťnožcov Hexapoda). Naopak, u mnohých skupín vznikajú zložité odvodené sklerity (najmä v ektoskelete hrudi).
Hlava vzniká zrastom prvých šiestich telových segmentov. Proces vzniku hlavy sa nazýva cefalizácia. Pre aktívne pochopenie vzťahov medzi troma hlavnými recentnými skupinami článkonožcov (Chelicerata, Crustacea a Tracheata) je nevyhnutné venovať hlavovým segmentom a ich príveskom aspoň stručný výklad:
Hruď (thorax) je u pôvodnejších viacnôžok ešte neodlíšená od bruška, ale už u dvojkrídlovcov sú 3 hrudné somity naznačené tým, že majú len po jednom páre nôh (bruškové somity zrastajú po dva a vytvárajú diplosomity). Zvyšné vzdušnicovce (teda šesťnožce) majú hruď mohutnú, vždy trojčlánkovú. Hruď kôrovcov sa skladá z väčšieho počtu článkov (obyčajne 7 – 8, no niekedy až 11). U primitívnych foriem ju tvoria voľné články, väčšinou však pozorujeme zrastanie jedného, dvoch, alebo všetkých článkov hrudi s hlavou – vzniká tak pereion, nazývaný aj prosoma alebo cephalothorax. U klepietkavcov zrastá hlava s hruďou na pravú hlavohruď (cephalothorax), vzácnou výnimkou sú nohatky (Pantopoda) s voľnými hrudnými článkami Nohatky sa však už v súčasnosti vyčlenujú ako sesterský kmeň klepietkavcov, čiže doň nepatria. U mnohých roztočov (Acarina) ostáva hlava voľná, ale hruď zrastá s bruškom – vzniká tzv. idiosoma, a niekedy sa dokonca všetky 3 telové oddiely spoja do 1 celku (corpus, omniosoma).
Bruško (abdomen) má končatiny len u Myriapoda, Merostomata a sčasti u Crustacea, u ostatných pozorujeme úplnú redukciu končatín. Niekedy sú však bruškové končatiny modifikované na snovacie bradavky (čeľaď Araneae) a skoro vždy z nich vzniká pomocný kopulačný aparát samcov.
Končatiny sú tiež článkované, primárne dvojvetvové. U väčšiny článkonožcov vonkajšia vetva (exopodit) zaniká, takže končatinu tvorí len vnútorná vetva (endopodit). Niekedy je končatina tiež dvojvetvová, ale jej vetvenie je odvodené, posunuté apikálne (bulbus u pavúkov, klepetá u rakov a niektorých pavúkovcov). Končatina sa primárne skladá z týchto častí:
U jednotlivých skupín však pozorujeme množstvo odchyliek – napr. koleno u vzdušnicovcov úplne redukuje, niekedy zaniká i trochanter, a pod. V stavbe nôh a zvlášť modifikovaných končatín sa u článkonožcov prejavuje mimoriadna variabilita.
Svalstvo článkonožcov je priečne pruhované a upína sa na kutikulárny ektoskelet zvnútra. U viacnôžok je ešte pomerne jednoduché a pripomína fylogeneticky východiskový stav – svalový vak mnohoštetinavcov. U vyšších článkonožcov pozorujeme rozpad svalového vaku v špecializované svalové skupiny – najmä svaly v končatinách sú dosť zložité a ťažko homologizovateľné so svalmi stavovcov (Vertebrata).
Telová dutina článkonožcov je mixocoel – vzniká splynutím embryonálne zakladaných coelomových dutiniek so zvyškom primárnej telovej dutiny (blastocoelu). Intersegmentálne disepimenty zanikajú, no ich rudimentom zostáva systém lakún – blanitých priepustí medzi sinusmi (dutinkami) otvorenej obehovej sústavy; pričom je zrejmé, že sinusy sú zvyškami coelomových dutiniek. Obehová sústava sa však na rozvode dýchacích plynov podieľa len u kôrovcov a primitívnych pavúkovcov, u vyšších pavúkovcov iba čiastočne (Araneae) alebo vôbec nie (ostatné vyššie Arachnoidea a všetky Tracheata). Pokiaľ hemolymfa plní aj dýchaciu (respiračnú) funkciu, je dýchacím pigmentom hemocyanín, niekedy hemoglobín – časť Crustacea a niektoré vodné larvy (Nematocera). Dýchacie pigmenty sú voľne rozpustené v hemolymfe, nie sú sústredené v erytrocytoch. Hemolymfu uvádza do pohybu trubicovité srdce na chrbtovej strane bruška. Po bokoch má štrbinovité ostie, ktoré majú funkciu chlopní. Pri systole (zmrštení srdca) je hemolymfa vypudená aortou smerom k hlave, potom sa rozlieva v systéme sinusov, až sa dostane opäť do abdomenu, odkiaľ je pri diastole (roztiahnutí srdca) nasávaná do perikardiálneho sinusu a ostiami do srdca. U drobných kôrovcov je niekedy srdce bez aorty (Cladocera), alebo môže úplne chýbať (Ostracoda a väčšina Copepoda). Hemolymfa je vtedy uvádzaná do pohybu peristaltikou čreva a pohybom svalov v tielku.
Stavba dýchacej sústavy závisí od spôsobu života. Primárne vodné článkonožce (kôrovce a Merostomata) majú žiabre, ktoré sú pôvodne na žiabrových príveskoch (epipoditoch) pri báze hrudných i bruškových končatín; druhotne môžu mať inú polohu – napr. u Decapoda visia do dvoch dutín medzi bočným rozšírením karapaxu a stenou tela. U mnohých drobných druhov žiabre chýbajú a výmena plynov prebieha cez povrchovú jemnú kutikulu (Cladocera, Ostracoda, Copepoda). Dýchacie orgány suchozemských článkonožcov sú dvojakého typu: Pľúcne dutinky a vzdušnice
Pľúcne dutinky sú fylogeneticky primitívnejším typom dýchacích orgánov. Sú umiestnené na spodnej strane bruška v počte 1 pár (väčšina Araneae) až 4 páry (Scorpionidea). Vznikli druhotným ponorením žiabrových epipoditov pod úroveň povrchu tela a tu, v uzavretých dutinkách (samozrejme vo vlhku) prebieha výmena plynov. Pľúcne dutinky komunikujú s vonkajškom len cez štrbinovité otvory, ktoré sú chránené viečkom (operculum).
Vzdušnice (trachey) sú vývojovo vyšším typom dýchacieho ústrojenstva. Sú to vlastne ektodermálne vliačeniny, spevnené chitinóznou špirálovitou výstužou. Tracheálna sústava vyúsťuje na povrch nenápadnými otvormi (stigmami) po bokoch bruška, niekedy i hrude. Vetvia sa v systém vzdušničiek (tracheol), ktoré privádzajú kyslík priamo k bunkám. Takýto mechanizmus dýchania je veľmi efektívny – vylučuje totiž akékoľvek zdržanie v zásobovaní tkanív kyslíkom, spôsobené u iných skupín živočíchov zložitým prenosom O2 a CO2 dýchacími pigmentami. Znamená to, že svaly Tracheata nikdy nepracujú na kyslíkový dlh, tak častý napr. pri mäkkýšoch, no i inde, vrátane stavovcov. Vzdušnice sú však výhodné len pre telo menších rozmerov. Veľkosť tela vzdušnicovcov má prirodzenú fyziologickú limitu asi 20 – 25 cm, väčšie telo by už tracheálny systém nedokázal efektívne zásobovať kyslíkom. U niektorých lariev vodného hmyzu vznikajú tracheálne žiabre. Sú to vlastne druhotné vyliačeniny kutikuly obyčajne na konci alebo po bokoch bruška, do ktorých vnikajú trachey. K výmene plynov dochádza na jemnej, lamelárne zriasenej kutikule (najády Ephemeroptera, Zygoptera a Plecoptera). Niektoré vodné larvy dýchajú rektálnou dutinou vystlanou tracheolami (Anisoptera), alebo primitívnymi kutikulárnymi žiabrami (Simuliidae). Nakoniec, u suchozemských alebo amfibických článkonožcov platí to isté ako u vodných – u niektorých drobných foriem špecializované dýchacie orgány zanikajú, takže výmena plynov prebieha iba povrchom tela (napr. veľmi drobné roztoče).
Nervová sústava je u primitívnejších skupín (viacnôžky, nižšie kôrovce) ešte v podstate rebríčková, väčšinou (u odvodených typov) je už gangliová. Centrom nervovej sústavy je cerebrálne ganglium, alebo mozog. Vzniká zrastom troch párových ganglií neurocephalonu. V hlave je okrem toho aj podhltanová uzlina, ktorá vznikla zrastom troch párov ganglií gnathocephalonu. Inervuje ústne orgány. Z hlavy vedú smerom do tela dva hlavné nervové povrazce, no tieto obyčajne integrujú v jeden povrazec, ktorý v každom somite vytvára uzlinu. Zvlášť veľké sú uzliny v hrudných somitoch, ktoré inervujú zložité svalstvo končatín, a u hmyzu aj lietacie svaly. U odvodených typov článkonožcov pozorujeme časté zrasty ganglií, ktoré korešpondujú s tagmatizáciou (zrastaním hlavných telových oddielov) – napr. u hmyzu s kompaktnou hruďou je len jedna mohutná hrudná uzlina – ganglion thoracale.
Článkonožce majú veľmi rozvinuté zmysly. Receptory sa obyčajne rozdeľujú do troch kategórií:
Mechanoreceptory reagujú na mechanické podnety: vibrácie, tlak, zemskú gravitáciu, no zrejme i na teplotu a pod. Sú umiestnené na celom povrchu tela, no hlavne na exponovaných častiach: na hlave, končatinách a tykadlách. Sú to tzv. hmatové senzily – poznáme ich v zásade 3 typy:
Posledný typ majú hlavne pavúky a bolo dokázané, že tieto veľmi jemné senzily registrujú aj vibrácie vzduchu – pavúky teda istým spôsobom „počujú“, i keď, samozrejme, na inej úrovni ako cicavce. Diferencovaný sluchový zmysel je u článkonožcov zriedkavý, ale vyskytuje sa. Sú to tzv. tympanálne orgány, ktoré sú vyvinuté u aktívne stridulujúceho hmyzu (Saltatoria, Auchenorrhyncha). Ďalším mechanoreceptorom sú statocysty. U článkonožcov sú vždy v hlave, druhotne môžu splynúť v jeden orgán. U Decapoda sa nachádzajú v bazálnych článkoch prvého páru tykadiel. Zvláštne mechanoreceptory sú i vnútri tela, najmä u hmyzu, a nazývajú sa chordotonálne orgány. Registrujú napätie v jednotlivých častiach tela, odpor prostredia, a tým aj rýchlosť prúdenia vzduchu, vody a pod. Najznámejší je tzv. pedicelový (Johnstonov) orgán, ktorý sa nachádza v druhom článku tykadla ako dôležitý znak triedy Ectognatha. U niektorých skupín (Diptera) môže registrovať aj vibrácie vzduchu, teda zvuk.
Chemoreceptory reagujú na látky rozpustené vo vzduchu a vo vode – predstavujú teda čuchový a chuťový zmysel, u článkonožcov sú tieto vnemy často spojené. Chemoreceptory sú opäť predovšetkým na hlave a sú to dosť jednoduché senzorické jamky alebo papily. Chuťové senzily sú hlavne v ústach a na ústnych príveskoch, čuchové sú najmä na tykadlách. Niektoré skupiny hmyzu, kde samec vyhľadáva samicu na diaľku pomocou feromónov, nemajú v ostrosti čuchu medzi živočíchmi konkurenciu.
Fotoreceptory registrujú ľudským okom viditeľné, ale často i neviditeľné elektromagnetické žiarenie. Sú to oči niekoľkých typov:
Tráviaca sústava môže byť rozlične stavaná. Začína sa ústami s ústnymi skleritmi a príveskami, ktoré vznikli modifikáciou hlavových končatín. Nasleduje hltan (pharynx), plynule prechádzajúci do pažeráka (oesophagus), ktorý sa často rozširuje v hrvoľ (ingluvies). Hrvoľ slúži v niektorých prípadoch ako zásobník na predtrávenú alebo rôzne upravenú potravu pre larvy (napr. včely). Tráviaca sústava pokračuje svalnatým alebo žuvacím žalúdkom (proventriculus) a prechádza do žľaznatého žalúdka (ventriculus, mesentheron), ktorý je ako jediný v tráviacej sústave endodermálneho pôvodu. Na rozhraní žuvacieho a žľaznatého žalúdka je často systém slepých výbežkov (coeca), ktoré obsahujú symbiotické mikroorganizmy, u xylofágnych druhov nevyhnutné na trávenie celulózy. Ventriculus má u pôvodných typov (Crustacea, Chelicerata) hepatopankreatickú žľazu, ktorá môže byť tvorená viacerými lalokmi. Okrem sekrečnej funkcie priamo v hepatopankrease prebiehajú tráviace procesy. U drobných foriem (Cladocera, Ostracoda, mnohé Acarina) môže byť hepatopankreas redukovaný. U Tracheata produkuje tráviace enzýmy priamo žľaznatý epitel žalúdka. Žalúdok pokračuje črevom (ileum), ktoré sa končí rektálnou dutinou s análnym otvorom.
Vylučovaciu sústavu tvoria orgány dvojakého charakteru:
S exkrečným systémom súvisí aj tukové teleso (corpus adiposum), prítomné hlavne u lariev. Má primárne zásobnú funkciu, ale hromadia sa v ňom aj niektoré metabolity, ktoré zostali nevylúčené. Nakoniec, k exkrečným mechanizmom patrí aj zvliekanie kutikuly (u lariev, a niekedy i dospelcov). Vo vrstvičke tekutiny, ktorá oddeľuje starú kutikulu od novej, nájdeme tiež určité látky, ktorých sa organizmus potrebuje zbaviť.
Sú okrem vzácnych výnimiek (parazitické kôrovce) gonochoristy. Oplodnenie je vnútorné, pomocou penisu alebo modifikovaných končatín. Vajíčko má chitinózny obal, často s charakteristickými skulptúrami. V hornej časti vajíčka je 1 alebo viac otvorov (mikropyle), ktorými sa spermia dostáva k jadru. Väčšina druhov sa vyvíja z oplodneného vajíčka, no pre niektoré skupiny je typická partenogenéza. Poznáme v zásade 3 typy partenogenézy:
Vajíčko obsahuje dosť žĺtka a jeho brázdenie je superficiálne, gastrula vzniká epiboliou. Vývin môže byť priamy i nepriamy. Prebieha v mnohých variantoch. Typy premeny sú však veľmi dôležité pre systematické triedenie kmeňa.
U Arthropoda, keďže ide o článkované živočíchy, je priamy vývin nazývaný často epiméria; terminologicky jasnejšie by bolo asi označenie juvenilná izoméria. Pri epimérii už 1. instar po vyliahnutí z vajíčka vyzerá ako „zmenšenina“ dospelého živočícha a má hneď plný počet somitov. Vyskytuje sa u väčšiny Arachnoidea, ďalej u Cladocera, Isopoda, Amphipoda, Astacidae, Scolopendromorpha, Geophilomorpha, Thysanura a u triedy Entognatha s výnimkou Protura.
Na hranici priameho a nepriameho vývinu je anaméria alebo juvenilná anizoméria. Pri anamérii juvenilný jedinec vyzerá skoro ako dospelý, líši sa len menším počtom somitov, a niekedy len menším počtom končatín. Definitívny počet somitov a končatín získava medzi jednotlivými zvliekaniami. Typicky anamérne sú napr. Pseudoscorpionidea, Opilionidea, Acarina, všetky Diplopoda, Lithobiomorpha a Protura. U niektorých skupín je na začiatku vývinu larva, ktorá prekonáva premenu na subadultné štádium, ktoré sa mení na dospelého jedinca anamérnym spôsobom (Copepoda, Merostomata, Pantopoda).
Nepriamy vývin prebieha cez sekundárne larvy; u jednotlivých skupín Arthropoda prichádza do úvahy vlastne 5 variánt:
1. premena väčšiny nižších kôrovcov cez nauplius;
2. premena väčšiny vyšších kôrovcov (Malacostraca) cez zoeu;
3. premena hrotnáčov (Merostomata) cez tzv. trilobitoidnú larvu;
4. premena nohatiek (Pantopoda) cez tzv. phoxichilídium;
V prípadoch 1 až 4 prebieha premena subadultného štádia na dospelého jedinca v podstate anamérnym spôsobom. (Napr. nauplius u Copepoda prekonáva premenu na subadultné, tzv. kopepoditové štádium, ktoré sa mení na dospelého jedinca vlastne anamérnym procesom. Niekedy oddeľuje tieto vývinové štádiá ďalší instar, tzv. metanauplius);
5. premena hmyzu (Ectognatha = Insecta) cez rôzne utvárané sekundárne larvy. Pri hmyze hovoríme viac-menej historicky o dvoch hlavných typoch premeny:
Dodnes je popísaných asi 1 200 000 druhov článkonožcov, no skutočný počet bude ešte vyšší: minimalistické odhady sú asi 3 milióny, maximalistické až 10 miliónov. Tieto údaje nie sú nadnesené – z asi 80-tich radov článkonožcov sú dôkladne preskúmané vlastne iba motýle a chrobáky, a aj to len vo vyspelejších krajinách sveta. Celé veľké oblasti (sub)trópov, ale i morí ešte len čakajú na systematický výskum väčšiny radov a čeľadí článkonožcov.
Systematika článkonožcov v súčasnosti patrí k najspornejším v celej živočíšnej systematike. O rôznych variantoch systematiky článkonožcov pozri aj články bezhryzadlovce, hryzadlovce a vzdušnicovce.
Systematika primárne používaná v tejto wikipédii je takáto [3][4][5][6][7][8]:
kmeň: článkonožce (Arthropoda)
Poznámky:
Niektorí autori (najmä staršie) do článkonožcov (Arthropoda) zaraďujú aj pomalky (Tardigrada) alebo pazúrikavce (Onychophora) alebo tak pomalky ako aj pazúrikavce. V súčasných systémoch väčšinou Arthropoda nezahŕňajúce pomalky či pazúrikavce nazývame na upresnenie aj Euarthropoda (i keď tento názov sa niekedy používa aj v trochu užších významoch [6]); zoskupenie Euarthropoda+Tardigrada+Onychophora nazývame Panarthropoda (iné názvy: Gnathopoda, Ailopoda, Podophora, Antennopoda); a zoskupenie Euarthropoda+Tardigrada nazývame Tactopoda. [9][10][11][12]
Článkonožce (staršie: článkonohé; lat. Arthropoda) sú najpočetnejší živočíšny kmeň. Je to obrovská, dosť starobylá a vnútorne veľmi diferencovaná skupina článkovaných živočíchov, ktoré fylogeneticky nadväzujú na Polychaeta. V procese fylogenézy však získali znaky, ktoré im umožnili osídliť všetky suchozemské i sladkovodné ekosystémy a vrátiť sa aj naspäť do mora, z ktorého vzišli. Všade, kde sa vyvíjali, dosiahli mimoriadnu mnohorakosť foriem a prekvapujúcu diverzitu (na súši tvoria podľa niektorých odhadov dokonca až 95 % živočíšnych druhov).
Členonožci (znanstveno ime Arthropoda, iz grščine ἄρθρον arthron - »sklep« in ποδός podos - »stopalo«, dobesedno stopala s sklepi) so najštevilčnejše deblo živali. Znanih je preko milijon vrst, kar znaša tri četrtine vseh znanih živalskih vrst,[1] po eni od novejših ocen pa naj bi bilo vseh vrst med 5 in 10 milijonov.[2]
Sodeč po telesni zgradbi in drugih značilnosti so členonožci sorodni kolobarnikom. Najočitnejša podobnost je, da imajo oboji členjeno telo in enako osnovno zgradbo živčevja z možgani na hrbtni strani sprednjega dela telesa in trebušnjačo - živčno povezavo na trebušnem delu z zadebelitvami (gangliji) v vsakem členu. Členi so med seboj različni (t. i. heteronomno členjenje) in tvorijo jasno definirane telesne regije, največkrat dve ali tri - glavoprsje in zadek ali glavo, oprsje in zadek. Zanje je značilno tudi, da jih obdaja trden, večplasten zunanji skelet, ki ga izloča zunanja plast kože. Deli se v tanko, zunanjo plast prokutikulo in mnogo debelejšo epikutikulo spodaj. Prokutikula je iz beljakovin in včasih tudi vodoodpornih voskov, epikutikula pa je ponovno dvoplastna, sestavljata jo eksokutikula na zunanji in endokutikula na notranji strani. Obe gradijo beljakovine in polisaharid hitin. Zgradba je v eksokutikuli ojačana s procesom, podobnim strojenju: molekule so križno povezane med seboj, kar da trdno, neprožno strukturo, ki je značilna za členonožce. V pregibih med členi eksokutikula manjka, zato so upogljivi. Zaradi trdnega zunanjega ogrodja se morajo vsi členonožci leviti - periodično odvreči pretesno hitinjačo, da lahko rastejo. Njihova obarvanost je najpogosteje posledica prisotnosti melaninskih pigmentov v skeletu, ki dajejo rjavo, rumeno, rdečo ali oranžno barvo. Nekatere vrste pa so živopisanih, iridescenčnih barv, ki nastanejo zaradi loma svetlobe na fini rebrasti strukturi vrhnje plasti.
Ime so dobili po značilnosti, da so poleg telesa členjene tudi okončine. Razen pregibov so toge in omišičene tako, da se lahko hitro premikajo po principu vzvoda. Izrastki zunanjega skeleta v notranjosti okončin, na katere se pripenjajo mišice, omogočajo natančnejši nadzor nad gibanjem. Členjene okončine so ena od bistvenih prednosti napram kolobarnikom, ki je verjetno omogočila takšen razmah členonožcev. Poleg tega so se pri raznih podskupinah okončine spremenile za opravljanje drugih nalog poleg lokomocije. Krilate žuželke so med členonožci posebnost, saj imajo poleg nog še dodaten par ali dva kril, ki so se razvila povsem ločeno.
Tudi notranja zgradba členonožcev je členjena. Imajo odprto krvožilje, kjer hemolimfa obliva notranje organe v skupni telesni votlini. Poganja jo bolj ali manj izraženo cevasto srce, sodeluje pa tudi skeletno mišičje kadar je žival aktivna. Zgradba dihal in izločal je raznolika, glede na taksonomski položaj in okolje, v katerem določena skupina členonožcev živi. Čutila za kemične in mehanske dražljaje so večinoma v obliki izrastkov skeleta - čutilnih dlak, poleg tega pa mnogi členonožci tudi vidijo s katerim od tipov očes. Ta so bodisi enostavna očesca, bodisi sestavljena očesa.
Členonožci imajo z izjemo nekaj ozko specializiranih skupin ločena spola in se razmnožujejo spolno. Oploditev je po navadi notranja, bodisi neposredno, bodisi posredno, kadar samec odloži paket semenčic in ga samica nato pobere s svojo spolno odprtino. Zunanje oploditve se poslužujejo samo nekateri vodni predstavniki. Praktično vsi členonožci ležejo jajčeca, v katerih se razvijajo zarodki. Navidezna izjema so ščipalci, kjer se ličinke izležejo iz jajčec že v materinem telesu.
Pravkar izlegla ličinka je lahko miniaturna kopija odrasle živali brez spolnih organov, ali pa se od nje tako skrajno razlikuje, da je bila še do pred nedavnim opisana kot ločena vrsta (npr. pri nekaterih rakih). Zaporedje levitev, skozi katere gre preden odraste, razdeli življenje členonožca v jasno razmejene faze (stadije). Levitve se lahko nadaljujejo tudi skozi odraslo obdobje do konca življenja. Prehodu med ličinko in odraslo živaljo pravimo preobrazba. Ta pri členonožcih, ki se kot odrasli ne levijo več, sovpada z zadnjo levitvijo.
Členonožci živijo v skoraj vseh naravnih okoljih na Zemlji, od visokogorij do morskih globin in ekstremnih habitatov. Poleg amniotov so edina skupina živali, ki uspeva v suhih okoljih. To jim omogoča zunanji skelet, ki je lahko prevlečen z voskasto plastjo in učinkovito ščiti pred izsušitvijo.
Členonožce navadno razvrščamo v pet poddebel, od katerih so predstavniki enega v celoti izumrli:
Vendar pa je klasifikacija na tem nivoju še vedno predmet mnogih debat in rednega prilagajanja razvrstitve novim odkritjem. Nedvoumno razvrščanje otežujejo tudi nekateri fosilni členonožci, ki bodisi niso podobni nobeni od uveljavljenih skupin, bodisi kažejo podobnost z več kot eno. Rezultati molekularnih raziskav med drugim porajajo dvom v veljavnost skupine rakov.[3]
Členonožci (znanstveno ime Arthropoda, iz grščine ἄρθρον arthron - »sklep« in ποδός podos - »stopalo«, dobesedno stopala s sklepi) so najštevilčnejše deblo živali. Znanih je preko milijon vrst, kar znaša tri četrtine vseh znanih živalskih vrst, po eni od novejših ocen pa naj bi bilo vseh vrst med 5 in 10 milijonov.
Leddjur (Arthropoda) är en stam inom djurriket som omfattar så varierande djurgrupper som insekter, spindeldjur, mångfotingar och kräftdjur. Leddjuren är jordens artrikaste stam, omkring 80 procent av alla beskrivna djurarter beräknas vara leddjur. Sammantaget finns över en miljon kända arter av leddjur, av vilka de flesta är insekter. I Sverige finns det 29 060 kända arter.[1]
Benämningen "leddjur" kommer sig av att gruppen delar det gemensamma, grundläggande draget att kroppen, och benen, är ledade. Dessa leders funktion möjliggörs genom att leddjuren har ett så kallat exoskelett av kitin, ett yttre förhårdnat "hudskal" som inte bara tjänar som skydd, utan också utgör fäste för musklerna. Leddjurens muskler är fästade för att uppnå en hävstångseffekt (att böja en arm, med triceps- och bicepsmuskler, vore omöjligt för ett däggdjur, om dessa muskler ej vore fästa på överarmsbenet; däggdjur har med andra ord ett endoskelett). Dessa grundläggande anatomiska drag ger leddjuren stor rörlighet och leddjur har utvecklat förflyttning både genom att hoppa, simma, gå och flyga.
Leddjuren var en av de tidigaste grupper som lämnade havet och gick upp på land. De kan spåras 550 miljoner år tillbaka, men det är fortfarande inte säkerställt i vilka primitiva djurgrupper de har sitt ursprung. För cirka 350 miljoner år sedan intog de också luften, i form av de första flygande insekterna, och det skulle dröja ytterligare 100 miljoner år innan djur från någon annan grupp gjorde dem sällskap. Till utdöda grupper av leddjur hör trilobiter och havsskorpioner.
Gemensamma anatomiska drag för alla leddjur är att kroppen är indelad i segment från vilka de ledade, parvist ordnade benen utgår. Kärlsystemet är öppet och för kroppsvätskornas cirkulation finns ett hjärta. För andningen finns antingen gälar, boklungor eller ett trakésystem. Nervsystemet hos leddjur består av många ganglier som hålls samman med två nervtrådar, som går genom kroppens segment och är förbundna med hjärnan.
Olika grupper av leddjur har utvecklats så att segmentens funktion allt mer specialiserats på olika vis, genom att segmenten har vuxit samman och bildat särskilda funktionella enheter, som ett huvud, eller bildat särskilda funktionella bihang, som antenner.
Hos enkelfotingarna, som hör till de primitivare leddjuren, finns bara ett huvud och en kropp uppdelad i segment som till sin funktion inte skiljer sig så starkt åt. Ett par gångben utgår från vart och ett av kroppens segment. Dubbelfotingarna har en liknande enkel byggnad med uppdelning i ett huvud och en kropp, men hos dessa har två på varandra följande ursprungliga segment smält samman till ett dubbelt segment. Två par gångben utgår därför från vart och ett av kroppens synliga segment.
Spindlarna har en kroppsbyggnad som är uppdelad i två grundläggande enheter, cephalothorax som består av det sammanvuxna huvudet och mellankroppen, och bakkroppen. De har åtta gångben ordnade i fyra benpar. Hos insekterna har kroppen tre grundläggande enheter, huvud, mellankopp och bakkropp. Insekterna har sex gångben ordnade i tre benpar. Kräftdjuren är en mycket varierad grupp, de har huvud, mellankropp och bakkropp, även om bakkroppen hos vissa grupper är starkt reducerad. De kan ha fler ben, till exempel har de tiofotade kräftdjuren tio gångben ordnade i fem benpar.
Leddjur förekommer över hela världen, i många olika miljöer. Insekterna dominerar på land, medan kräftdjuren dominerar i havet.
Att beräkna det totala antalet av levande arter är oerhört svårt då det ofta har som utgångspunkt en serie av antaganden för att räkna upp antalet från specifika platser för att beräkna hela världen. En studie från 1992 beräknade att det i Costa Rica allena fanns cirka 500 000 arter, av vilka 365 000 var leddjur.[2] Andra uppskattningar slår fast att leddjuren har cirka 1 170 000 beskrivna arter vilket utgör cirka 80 procent av alla kända levande varelser.[3] Leddjuren är viktiga medlemmar i marina ekosystem, samt i ekosystem i färskvatten, och på land- och i luften. Det är bara leddjuren och amnioterna, vars levande medlemmar är reptiler, fåglar och däggdjuren, som har anpassat sig för ett liv i torra miljöer.[4]
Leddjur uppvisar som den mångformiga grupp de är stor variation i levnadssätt. Utformningen på mundelarna och benen är karaktärer som visar på deras födoval. Det finns såväl predatorer som växtätare och sådana som livnär sig på dött organiskt material och de som är filtrerare. I näringskedjor är leddjuren själva ofta föda för många andra djur. I ekosystemen har de en viktig roll, till exempel som nedbrytare och pollinatörer.
Många leddjur har en fortplantning med inre befruktning men det finns också exempel på yttre befruktning, främst bland leddjur som lever i havet. En del leddjur, bland annat vissa insekter, kan också föröka sig genom partenogenes. Hos leddjur är hermafrodism ett sällsynt undantag, men det förekommer till exempel hos rankfotingar. De flesta leddjur lägger ägg, men skorpioner uppvisar vivipari, det vill säga att de föder levande ungar.
De unga individerna av en del leddjur, som spindlar och skorpioner, liknar de fullvuxna djuren, medan unga individer av andra leddjur, som kräftdjur och många insekter inte alls liknar de fullvuxna djuren och därför vanligen benämns larver.
Omsorgen om avkomman är varierande, det finns leddjur som uppvisar både mycket liten och mycket stor omsorg om avkomman. En del marina leddjurs ägg och larver driver till exempel med strömmarna, medan många samhällsbildande insekter lägger ner mycket tid på vård av äggen och uppfödningen av larverna.
Ett gemensamt drag för flertalet leddjur är att deras exoskelett begränsar tillväxten och måste bytas ut. Så är fallet för kräftdjur, mångfotingar och spindlar. Insekter utvecklas genom metamorfos och byter med ett fåtal undantag inte hud som fullvuxna. När ett leddjur byter hud är dess kropp som mest oskyddad och de är då extra sårbara för fiender.
Kräftdjur, som krabbor, humrar, kräftor och räkor har länge varit en del av människans mat och odlas nu i stor, kommersiell skala. Insekter och deras larver är minst lika näringsrika som kött och äts både råa och kokas i många icke-europeiska kulturer. Kokt tarantel anses vara en delikatess i Kambodja. Det största bidraget av leddjur till människornas livsmedelsförsörjning är av pollinering. En studie 2008 undersökte 100 grödor som enligt FAO odlades för användning som livsmedel och beräknade pollineringens ekonomiska värde till 153 miljarder euro, vilket är 9,5 procent av världens jordbruksproduktion för livsmedel under 2005. Bin producerar också honung som ligger till grund för en snabbt växande industri och internationell handel.
Trots att leddjuren är den största stammen på jorden och trots att tusentals arter är giftiga, står de för relativt få bett och sting på människor.[5] Följder av sjukdomar som överförs av blodsugande insekter är mycket allvarligare (det finns omkring 2000 myggsorter i Sverige). Andra blodsugande insekter infekterar kreatur och dödar därigenom många djur och minskar kraftigt nyttan av andra.
Den relativa enkelheten hos leddjurens kroppsplan, som låter dem gå på olika ytor både på land och i vatten, har gjort dem användbara som modeller för robotik. Överflödet av segment tillåter leddjur liksom biomimetiska robotar att röra sig normalt även med skadade eller förlorade tillbehör (som exempelvis kroppsdelar).[6][7]
Traditionellt har leddjuren, segmenterade som de är, sammanförts med de likaledes segmenterade ringmaskarna i en grupp som kallas Articulata[8], men molekylära data pekar klart på att denna grupp är polyfyletisk, och att leddjuren egentligen inte är närmare släkt med ringmaskar.
Som leddjurens närmaste släktingar idag räknas istället trögkrypare och klomaskar, och därutöver bildar de tillsammans med rundmaskar och flera andra smågrupper den högre gruppen Ecdysozoa.[9][10]
Till leddjuren räknas alla kräftdjur, sexfotingar (insekter m.m.), mångfotingar och palpkäkar (spindlar m.m.). Släktskapet mellan dessa fyra grupper är en omstridd fråga. Traditionellt har sexfotingar och mångfotingar betraktats som varandras närmaste släktingar. Tillsammans med kräftdjuren bildade dessa gruppen Mandibulata. Molekylära data kastar dock tvivel över den indelningen, och pekar snarare mot att sexfotingar och kräftdjur hör ihop[11]. En del studier finner till och med att sexfotingarna är en undergrupp inom kräftdjuren[12]. Se diskussion av detta under Pancrustacea
Leddjur (Arthropoda) är en stam inom djurriket som omfattar så varierande djurgrupper som insekter, spindeldjur, mångfotingar och kräftdjur. Leddjuren är jordens artrikaste stam, omkring 80 procent av alla beskrivna djurarter beräknas vara leddjur. Sammantaget finns över en miljon kända arter av leddjur, av vilka de flesta är insekter. I Sverige finns det 29 060 kända arter.
Benämningen "leddjur" kommer sig av att gruppen delar det gemensamma, grundläggande draget att kroppen, och benen, är ledade. Dessa leders funktion möjliggörs genom att leddjuren har ett så kallat exoskelett av kitin, ett yttre förhårdnat "hudskal" som inte bara tjänar som skydd, utan också utgör fäste för musklerna. Leddjurens muskler är fästade för att uppnå en hävstångseffekt (att böja en arm, med triceps- och bicepsmuskler, vore omöjligt för ett däggdjur, om dessa muskler ej vore fästa på överarmsbenet; däggdjur har med andra ord ett endoskelett). Dessa grundläggande anatomiska drag ger leddjuren stor rörlighet och leddjur har utvecklat förflyttning både genom att hoppa, simma, gå och flyga.
Leddjuren var en av de tidigaste grupper som lämnade havet och gick upp på land. De kan spåras 550 miljoner år tillbaka, men det är fortfarande inte säkerställt i vilka primitiva djurgrupper de har sitt ursprung. För cirka 350 miljoner år sedan intog de också luften, i form av de första flygande insekterna, och det skulle dröja ytterligare 100 miljoner år innan djur från någon annan grupp gjorde dem sällskap. Till utdöda grupper av leddjur hör trilobiter och havsskorpioner.
Eklem bacaklılar ya da Arthropoda, omurgasızların en büyük şubesidir.
Arthropoda (arthros: eklem, podos: ayak), "eklemli ayaklılar" anlamına gelmektedir. Vücutları; baş (caput veya cephalo), göğüs (thorax) ve karın (abdomen) olmak üzere üç bölgeden meydana gelir. Her bölge çeşitli sayıda segmentten (parçadan) ibarettir. Vücudun her bir segmenti esnek bir deriyle birbirine bağlanmıştır. Vücut yüzeyi, sert bir madde olan kitinden meydana gelen bir dış iskeletle örtülüdür. Kitin, dış epitelin salgısıdır ve hayvanların büyümesi için zaman zaman değiştirilerek atılır. Ekstremiteler (bacak, kanat, duyarga ve ağız parçaları gibi hareket edebilen çıkıntılar) daima eklemli parçalardan meydana gelmiştir. Sindirim sistemi ön, orta ve arka bağırsak olmak üzere üç bölümden ve bunların çeşitli kalınlıktaki kısımlarından meydana gelir. Kan dolaşımı açıktır. Kan oksijen taşımayıp, besin maddeleri, artık maddeler ve hormonları taşır. Dolaşım sistemi, sırtta bir yürek ve ana damardan meydana gelir. Bazılarında sırt damarından kollar da çıkar. Sinir sistemi, birbirine bağlantısı bulunan gonglion adı verilen boğumlardan ibarettir.
Ayrı eşeylidirler. Karada, havada ve suda yaşarlar. İnce derililer vücut yüzeyiyle, sudakiler solungaçla, karadakiler de trakealarla (borular sistemi) solunum yaparlar.
Yaklaşık 700.000 - 1.000.000 kadar türü bulunmaktadır.
Eklem bacaklılar şubesinin altı bacaklılar (Hexapoda), çok bacaklılar (Myriapoda), kabuklular (Crustacea) ve soyu tükenmiş trilobitler (Trilobitomorpha) alt şubelerinde görülen, duyu organı olarak gelişmiş hareketli baş uzantısı olan duyargalar, diğer alt şubesi olan ve akrep, kene ve örümceklerin yer aldığı keliserliler (Chelicerata) alt şubesinde yoktur. Kabuklularda iki çift, diğer gruplarda bir çift duyarga bulunur.
Eklem bacaklılar ya da Arthropoda, omurgasızların en büyük şubesidir.
Arthropoda (arthros: eklem, podos: ayak), "eklemli ayaklılar" anlamına gelmektedir. Vücutları; baş (caput veya cephalo), göğüs (thorax) ve karın (abdomen) olmak üzere üç bölgeden meydana gelir. Her bölge çeşitli sayıda segmentten (parçadan) ibarettir. Vücudun her bir segmenti esnek bir deriyle birbirine bağlanmıştır. Vücut yüzeyi, sert bir madde olan kitinden meydana gelen bir dış iskeletle örtülüdür. Kitin, dış epitelin salgısıdır ve hayvanların büyümesi için zaman zaman değiştirilerek atılır. Ekstremiteler (bacak, kanat, duyarga ve ağız parçaları gibi hareket edebilen çıkıntılar) daima eklemli parçalardan meydana gelmiştir. Sindirim sistemi ön, orta ve arka bağırsak olmak üzere üç bölümden ve bunların çeşitli kalınlıktaki kısımlarından meydana gelir. Kan dolaşımı açıktır. Kan oksijen taşımayıp, besin maddeleri, artık maddeler ve hormonları taşır. Dolaşım sistemi, sırtta bir yürek ve ana damardan meydana gelir. Bazılarında sırt damarından kollar da çıkar. Sinir sistemi, birbirine bağlantısı bulunan gonglion adı verilen boğumlardan ibarettir.
Ayrı eşeylidirler. Karada, havada ve suda yaşarlar. İnce derililer vücut yüzeyiyle, sudakiler solungaçla, karadakiler de trakealarla (borular sistemi) solunum yaparlar.
Yaklaşık 700.000 - 1.000.000 kadar türü bulunmaktadır.
Членистоногі мають багато спільного з кільчастими червами, особливо з поліхетами. Проте через ряд ароморфозів вони досягли вищого ступеня диференціації тканин та органів, перш за все нервової системи й органів чуття; деякі групи мають складну поведінку, яка виявляється в піклуванні про потомство, здатності до складної будівельної діяльності, суспільному житті й навіть мові (наприклад, у вищих перетинчастокрилих).
Як і кільчаки, членистоногі — метамерні тварини. Їхнє тіло складається з ряду сегментів, проте метамерія членистоногих різко гетерономна. Групи подібних сегментів об'єднуються у відділи тіла — тагми. Найчастіше виділяються три тагми: голова, груди, черевце. У павукоподібних голова злита з грудьми в єдину тагму — головогруди; у трилобітів і багатоніжок є всього дві тагми — голова і тулуб. Особливе значення має процес цефалізації — утворення голови — тагми, де сконцентровані органи чуття (вусики, очі) та первинної переробки їжі (ротовий апарат).
Голова складається з головної лопаті, або акрона, та кількох передніх сегментів. На думку більшості вчених, акрон гомологічний простоміуму кільчастих червів. Голова членистоногих має 6 сегментів. Кількість сегментів, котрі входять до складу грудей і черевця, яке закінчується анальною лопаттю, або тельсоном (гомолог пігідія), різко коливається, проте в межах типу спостерігається тенденція до стабілізації та зменшення їх кількості.
У результаті тагматизації об'єднується і внутрішній вміст сегментів; одні органи розростаються за рахунок інших рівноцінних органів. Зокрема, спостерігається концентрація гангліїв нервової системи в головний мозок.
Характерною ознакою членистоногих, від якої походить назва типу, є будова їхніх кінцівок. Вони складаються з окремих члеників, що рухомо з'єднані між собою суглобами, утворюючи багатоколінні важелі, здатні до складних і точних рухів. Відповідно до тагматизації тіла членистоногих відбулася й спеціалізація їхніх кінцівок. На голові знаходяться кінцівки, що виконують чутливу функцію — антени (одна або дві пари), решта кілька пар кінцівок перетворилися на ротові органи, що беруть участь у захопленні, утриманні та подрібненні їжі (верхні та нижні щелепи або хеліцери та частково педипальпи). Кінцівки грудної тагми виконують в основному локомоторну функцію (плавання, повзання, ходіння, бігання, стрибання), хоча до цих функцій іноді додаються й інші (дихальна, чутлива). Кінцівки черевця у багатьох груп редуковані. Лише у трилобітів і деяких ракоподібних (клас Malacostraca) вони призначені для плавання; одна-дві пари черевних кінцівок часто спеціалізуються як статеві, що призначені для запліднення та розмноження.
У найповнішому вигляді кінцівка членистоногого складається з основної частини (протоподит), від якої відходять дві гілки: зовнішня (екзоподит) та внутрішня (ендоподит), крім того, у водяних форм є зябровий відросток (епіподит). У первинно водяних членистоногих (наприклад, ракоподібних) деякі кінцівки зберігають двогіллясту будову; в більшості ж кінцівок одна з гілок редукується, і вони стають одногіллястими (наприклад, ноги у Шестиногих та Багатоніжок).
Важливою особливістю членистоногих, яка відрізняє їх від кільчастих червів і обумовлює основні риси їх організації, є наявність твердої кутикули, що виконує не тільки захисну функцію, а є також зовнішнім скелетом (екзоскелетом), до якого прикріплюються м'язи.
До складу кутикули входять білки, амінокислоти, ліпіди, глікопротеїди, феноли, пігменти, вода (до 40%). Проте найхарактернішим компонентом кутикули членистоногих є хітин. Це високомолекулярний полісахарид, мономером якого є глюкоза. Він нагадує глікоген або целюлозу, але на відміну від цих сполук кожна молекула мономера через групу NH зв'язана з ацетильною групою О=С—СН3. Його хімічна назва — полі-N-ацетил-d-глюкозамін. У членистоногих хітин представлений не в чистому вигляді, а як сполука з білком артроподином (насправді це кілька білків), який з хітином утворює нерозчинний комплекс.
У кутикулі членистоногих є ділянки, вкриті товстою, твердою кутикулою, нездатною до розтягнення (склерити), і м'якою, еластичною, розтяжною (мембрани). Чергування цих ділянок і зумовлює рухливість тіла та його придатків. Найчастіше в кожному сегменті (крім головних) є чотири склерити: спинний (тергіт), черевний (стерніт) та пара бічних (плеврити), з'єднані рухомо еластичними мембранами. У кінцівках членики вкриті твердою кутикулою, а зчленування — мембраною, що дозволяє їм рухатися один відносно іншого. Існує ще й внутрішній скелет — вирости склеритів усередину тіла. До них кріпляться м'язи. Кутикула не тільки вкриває все тіло членистоногого, а й вистилає передню та задню кишки, а також трахеї у трахейнодишних.
Мускулатура членистоногих не утворює суцільного м'язового мішка, а представлена окремими пучками, що з'єднують між собою рухливі ділянки кутикулярного скелету — склерити, або членики кінцівок. Мускульні пучки прикріплюються до внутрішніх виростів кутикули. У членистоногих виникає новий тип руху — з опорою на зовнішній скелет, а не на шкірно-м'язовий мішок, як у червів. Майже вся мускулатура членистоногих, за винятком деяких м'язів внутрішніх органів, поперечносмугаста. Це надає їм певної переваги, оскільки поперечносмугасті м'язи скорочуються швидше, ніж гладенькі. Завдяки такій будові локомоторного апарату членистоногі здатні до дуже швидких (відносно їхніх розмірів) і різноманітних рухів, а деякі з них (більшість комах) — і до польоту.
Порожнина тіла членистоногих змішана. Під час ембріонального розвитку в них, як і в кільчастих червів, закладаються парні целомічні мішки, що мають метамерну будову. Пізніше стінки целомічних мішків руйнуються, розпадаючись на окремі клітини, а целомічні порожнини зливаються із залишками первинної порожнини тіла, утворюючи змішану порожнину тіла — міксоцель, що не має власної клітинної вистилки. Це система лакунарних або щілиноподібних порожнин між внутрішніми органами. Із мезодермальних клітин стінок целомічних мішків згодом утворюються мускулатура, клітини крові, жирове тіло та інші мезодермальні утвори. У міксоцелі циркулює рідина, що зветься гемолімфою. Вона є одночасно і порожнинною рідиною, і кров'ю.
Травна система членистоногих складається з трьох відділів: ектодермальної передньої, ентодермальної середньої та ектодермальної задньої кишки. Кожен із цих відділів, у свою чергу, диференціюється залежно від типу живлення. Характерною рисою травного тракту членистоногих, яка відрізняє їх від інших типів тварин, є перетворення кінцівок передніх сегментів тіла на ротові, призначені для утримання й механічної переробки їжі. У ряді випадків додатково до ротового апарата в передній кишці є особливий відділ для механічної обробки їжі (жувальний шлунок Malacostraca або м'язовий шлунок комах). Часто передня кишка служить і для тимчасового зберігання їжі — воло (анатомія) метеликів, бджіл. У більшості наземних членистоногих до передньої кишки відкриваються слинні залози. У середній кишці з'являються різноманітні вирости, що збільшують її поверхню (печінкові вирости ракоподібних та павукоподібних, пілоричні придатки комах). У середній кишці та її придатках відбуваються основні процеси травлення та всмоктування. Задня кишка, особливо в наземних членистоногих, також диференціюється на відділи, що виконують різні функції; найважливішою серед них є всмоктування води з екскрементів і повернення її в гемолімфу. Це необхідно для збереження води в організмі. Функцією кишечнику наземних членистоногих є також осморегуляція та видалення з організму продуктів дисиміляції, для чого призначені спеціальні трубчасті вирости задньої ділянки середньої кишки (у павукоподібних) або передньої частини задньої кишки (у комах та багатоніжок) — мальпігієві судини.
Видільна система первинноводяних форм (ракоподібні, мечохвости) представлена видозміненими парними целомодуктами, які мають різні назви залежно від їх розміщення (антенальні, максилярні, коксальні залози). У наземних членистоногих замість них цю функцію виконують мальпігієві судини разом із заднім відділом кишечника, в який вони відкриваються. Мальпігієві судини всмоктують розчинені у воді продукти обміну, переробляючи їх у нерозчинні речовини, а в задній кишці відбувається зворотне всмоктування води і деяких корисних речовин та повернення їх у порожнинну рідину.
Кровоносна система членистоногих незамкнена і частково редукована. У ній залишаються лише головні судини — спинна, іноді черевна та деякі бічні, капілярів немає. З'являється серце. Усі артерії відкриваються в порожнину тіла. Функції крові виконує гемолімфа.
Органи дихання членистоногих різноманітні. Дуже дрібні членистоногі, які мають тонкі покриви й живуть у воді або в дуже вологих місцях, можуть дихати всією поверхнею тіла. У великих водяних членистоногих органами дихання є зябра — це видозмінені кінцівки або їхні частини (епіподити). У наземних членистоногих органами дихання є легеневі мішки, які також вважають видозміненими кінцівками (у павукоподібних), та трахеї (у частини павукоподібних, багатоніжок і комах). Легеневі мішки — це глибокі мішкоподібні вгини зі складчастими стінками, що відкриваються назовні вузькими щілинами. Трахеї — тонкі, найчастіше розгалужені трубочки, що відкриваються назовні маленькими отворами, а всередині тіла обплітають усі внутрішні органи, доносячи кисень навіть до окремих клітин.
Нервова система членистоногих складається з надглоткового ганглія, або головного мозку, навкологлоткових конектив і черевного нервового ланцюжка. Будова надглоткового ганглія дуже складна. Він має три відділи: передній (протоцеребрум), середній (дейтоцеребрум) та задній мозок (тритоцеребрум). Найскладнішу будову має протоцеребрум. У черевному нервовому ланцюжку часто спостерігається концентрація гангліїв і утворення складніших гангліїв. У деяких випадках усі ганглії зливаються в єдиний синганглій, як у краба або кімнатної мухи.
Більшість членистоногих мають добре розвинені органи чуття (дотику, хімічного чуття, рівноваги, зору). Очі членистоногих бувають двох типів — прості, що мають одну лінзу, й складні, або фасеткові, до складу яких входить велика кількість, інколи кілька тисяч, вічок, або оматидіїв, що щільно прилягають одне до одного. Кожен оматидій сприймає тільки одну точку предмета, що знаходиться перед ним, у результаті чого фасеткове око дає зображення, що складається з безлічі окремих точок, тобто мозаїчне. Фасеткові очі характерні для мечохвостів, більшості ракоподібних і комах, а також для викопних трилобітів.
Членистоногі розмножуються лише статевим шляхом. Більшість їх — роздільностатеві, хоча відомі й гермафродити. Часто присутній виразний статевий диморфізм — зовнішня відмінність самиць і самців. Запліднення внутрішнє або сперматофорне — за допомогою одягнених оболонкою пакетів сперми (сперматофорів), які самець вводить у статеві отвори самиці чи підвішує до них.
Яйця дуже багаті на жовток, який оточує ядро яйцеклітини. Тому дробіння в них поверхневе й неповне. Воно зосереджене на певній ділянці поверхні яйця, що зветься зародковою смужкою. Членистоногим характерна ембріонізація розвитку, ранні етапи його проходять під захистом оболонки яйця. З яйця виходить личинка, тіло якої вже складається з кількох сегментів. Якщо після цього продовжується утворення нових сегментів, то такий тип розвитку зветься анаморфозом; якщо ж тварина вилуплюється з яйця з повним числом сегментів — епіморфозом. У деяких членистоногих розвиток прямий: з яйця виходить цілком сформована особина, але меншого розміру.
Ріст і зміна форми тіла членистоногих утруднюються через щільну нерозтяжну кутикулу, тому їх постембріональний розвиток супроводжується періодичними линяннями, коли стара кутикула скидається і замінюється новою. У той короткий період, коли нова кутикула м'яка, збільшуються розміри тіла. Линяння здійснюються під контролем нейроендокринної системи.
Động vật Chân khớp hay Động vật Chân đốt là những động vật không có xương sống, có một bộ xương ngoài (bộ xương vĩnh viễn), một cơ thể chia đốt và những đốt phụ. Động vật Chân khớp thuộc Ngành Chân khớp (Arthropoda từ tiếng Hy Lạp ἄρθρον arthron, "khớp", và ποδός podos "chân", nghĩa là "chân khớp"). Có hơn 1 triệu loài chân khớp được mô tả, khiến chúng chiếm trên 80% tất cả các sinh vật được tìm thấy, và là một trong hai nhóm động vật thực sự sinh sống được ở môi trường khô – nhóm khác là động vật có màng ối. Chúng có kích thước từ rất nhỏ như sinh vật phù du cho đến chiều dài vài mét.
Ước tính có khoảng 1.170.000 loài động vật chân khớp đã được miêu tả, và chúng chiếm hơn 80% tất cả các loài động vật còn sống đã được biết đến.[3] Một nghiên cứu khác ước tính rằng có khoảng từ 5 đến 10 triệu loài còn tồn tại, bao gồm đã miêu tả và chưa miêu tả.[4] Ước tính tổng số loài còn sống là cực kỳ khó khăn do nó thường phụ thuộc vào một loại các giả thiết để mở rộng quy mô tính toán ở từng khu vực đặc biệt nhân lên cho toàn thế giới. Một nghiên cứu năm 1992 ước tính có 500.000 loài động vật và thực vật ở Costa Rica, trong đó có 365.000 loài là động vật chân khớp.
Chúng là các sinh vật quan trọng trong các hệ sinh thái biển, nước ngọt, đất liền và trong không khí, và là một trong hai nhóm động vật chính đã thích nghi trong các môi trường khô; nhóm còn lại là amniote, là những loài còn sống là bò sát, chim và thú.[5] Một phân nhóm động vật chân khớp là côn trùng, đây là nhóm có nhiều loài nhất trong tất cả các hệ sinh thái trên cạn và nước ngọt.[6] Loài côn trùng nhẹ nhất có khối lượng nhỏ hơn 25 microgram,[7] trong khi loài nặng nhất hơn 70 gram.[8] Một số loài giáp xác thì có kích thước lớn hơn nhiều; như chân của các loài cua nhện Nhật Bản có thể dài đến 4 mét.[7]
Người ta cho rằng các động vật Ediacaran Parvancorina và Spriggina cách đây khoảng 555 triệu năm là các động vật chân khớp.[9][10][11] Các động vật chân khớp nhỏ có vỏ sống như động vật hai mảnh vỏ đã được tìm thấy trong các lớp hóa thạch tuổi Cambri sớm từ 542 đến 540 triệu năm ở Trung Quốc.[12][13] Các hóa thạch bọ ba thùy tuổi Cambri sớm nhất là 530 triệu năm, nhưng lớp này đã từng khác đa dạng và phân bố toàn cầu.[14] Các cuộc kiểm tra lại các hóa thạch trong đá phiến sét Burgess thập niên 1970 có tuổi 505 triệu năm đã xác định có một số loài động vật chân đốt, một số loài trong có không thể xếp vào bất kỳ nhóm nào đã từng được nhận dạng, và do đó đã nổ ra các cuộc tranh luận về sự bùng nổ Cambri.[15][16][17] Hóa thạch của Marrella trong đá phiến sét Burgess Shale đã cung cấp bằng chứng rõ ràng sớm nhất về sự lột xác.[18]
Các hóa thạch giáp xác sớm nhất có tuổi khoảng 514 triệu năm trong kỷ Cambri,[19] và tôm hóa thạch có tuổi khoảng 500 triệu năm.[20] Hóa thạch giáp xác thường được tìm thấy từ kỷ Ordovic trở về sau.[21] Chúng hầu hết sống hoàn toàn trong nước, có thể do chúng chưa bao giờ phát triển các hệ bài tiết that conserve water.[22]
Euarthropod đặc biệt được phân thành 5 phân ngành, một trong số đó đã tuyệt chủng:[24]
Bên cạnh các nhóm chính này, cũng còn một số nhóm hóa thạch, hầu hết từ Cambri sớm, chúng rất khó phân loại, hoặc vào nhóm thiếu quan hệ rõ ràng với các nhóm chính hoặc có quan hệ rõ ràng với nhiều nhóm chính. Marrella là nhóm đầu tiên được nhận dạng có sự khác biệt rõ ràng với các nhóm nổi tiếng.[25]
Phát sinh loài của các nhóm động vật chân khớp còn tồn tại là một lĩnh vực đang được quan tâm và có nhiều tranh cãi.[26] Các nghiên cứu gần đây nhất có khuynh hướng rằng giáp xác là cận ngành với các nhóm sáu chân khác được lồng trong nó. Myriapoda được nhóm cùng với Chelicerata trong một số nghiên cứu gần đây (tạo thành Myriochelata),[23][27] và với Pancrustacea trong các nghiên cứu khác (tạo thành Mandibulata).[28] Việc xếp các trilobite tuyệt chủng cũng là chủ đề tranh cãi.[29]
Vì mã quốc tế về danh mục động vật học công nhận không có sự ưu tiên trên cấp họ, một số cấp phân loại cao hơn có thể được xem xét theo nhiều tên gọi khác nhau.[30]
|tên 3= thiếu |họ 3= trong Authors list (trợ giúp) |tựa đề= trống hay bị thiếu (trợ giúp); |url= trống hay bị thiếu (trợ giúp); ||ngày truy cập= cần |url= (trợ giúp) |coauthors= bị phản đối (trợ giúp) |issn= (trợ giúp), doi:10.1144/SP286.10 |month= bị phản đối (trợ giúp) |coauthors= bị phản đối (trợ giúp) |month= bị phản đối (trợ giúp) |coauthors= bị phản đối (trợ giúp) Động vật Chân khớp hay Động vật Chân đốt là những động vật không có xương sống, có một bộ xương ngoài (bộ xương vĩnh viễn), một cơ thể chia đốt và những đốt phụ. Động vật Chân khớp thuộc Ngành Chân khớp (Arthropoda từ tiếng Hy Lạp ἄρθρον arthron, "khớp", và ποδός podos "chân", nghĩa là "chân khớp"). Có hơn 1 triệu loài chân khớp được mô tả, khiến chúng chiếm trên 80% tất cả các sinh vật được tìm thấy, và là một trong hai nhóm động vật thực sự sinh sống được ở môi trường khô – nhóm khác là động vật có màng ối. Chúng có kích thước từ rất nhỏ như sinh vật phù du cho đến chiều dài vài mét.
Согласно одной из гипотез, членистоногие возникли ещё до кембрийского взрыва[10]. Сторонники этой гипотезы считают первыми известными членистоногими представителей эдиакарской биоты Parvancorina и Spriggina, живших около 555 млн лет назад[11][12][13]. Уже в раннекембрийских (539—541 млн лет) слоях в Китае обнаружены явные мелкие членистоногие с раковинами примерно как у двустворчатых моллюсков[14][15]. Самые ранние найденные окаменелости трилобитов датируются ранним кембрием (530 млн лет назад), но большое разнообразие видов и всемирное распространение даже ранних трилобитов приводит к выводу, что они к тому времени существовали уже давно[16]. Повторное исследование окаменелостей фауны сланцев Бёрджес (возрастом около 505 млн лет) выявило много членистоногих, которых невозможно отнести ни к одной из известных групп. Это положило начало новому витку дебатов по поводу кембрийского взрыва[17][18][19]. Окаменелость Marrella из той же фауны предоставила исследователям первое ясное свидетельство линьки[20].
Самая ранняя окаменелость ракообразного датируется кембрием, 513 млн лет назад[21], а чуть позже, 500 млн лет назад, уже обнаруживаются десятиногие ракообразные, напоминающие креветок[22]. С ордовикского периода и далее ископаемые ракообразные встречаются достаточно часто[23].
К типу членистоногих относятся самые ранние известные наземные животные. Они датируются поздним силуром, около 419 млн лет назад. Вероятно, животные этого типа оставили и наземные следы возрастом около 450 млн лет[24]. У членистоногих имелось несколько преадаптаций для освоения суши, в том числе суставчатый экзоскелет предоставляющий защиту от иссыхания и силы тяжести, а также способы передвижения, независимые от наличия воды[25]. В то же время гигантские пресноводные ракоскорпионы стали рекордсменами по размеру среди членистоногих, достигая длины в 2,5 м[26].
Самый древний известный представитель паукообразных, Palaeotarbus jerami, жил около 420 млн лет назад (силурийский период)[27][прим. 2]. Attercopus fimbriunguis, живший 386 млн лет назад (девонский период), обладал первыми известными органами для выделения паутиноподобных нитей, но паутинных бородавок у него не было, и поэтому его нельзя причислить к истинным паукам[29] (которые впервые появляются в позднекарбоновых отложениях, 299 млн лет назад[30]). Из юрских и меловых пород известно уже много окаменелостей пауков, включая представителей многих современных семейств[31]. Первые известные скорпионы с лёгочным дыханием датируются началом каменноугольного периода[32].
Древнейшее известное насекомое, Rhyniognatha hirsti, и одно из двух древнейших известных шестиногих (наряду с Rhyniella praecursor) было обнаружено в райниевом кремне в 1919 году. Окаменелости датируются ранним девонским периодом, около 410 млн лет назад. Rhyniognatha питалась, предположительно, спорофиллами сосудистых растений или же могла быть хищником. Возможно это было одно из первых членистоногих, обладающих крыльями[33].
Положение группы членистоногих в таксономической иерархии неоднозначно. С одной стороны, некоторые авторы (например Сидни Ментон) предложили версию, что разные подтипы членистоногих независимо произошли от червеподобных предков (другими словами, предполагалась полифилия членистоногих). В частности, эти авторы считали, что подтип трахейные (Uniramia) ближе к онихофорам (Onychophora), чем к остальным членистоногим. Однако большинство учёных не приняло эту точку зрения. Это противоречит и генетическим исследованиям[34].
По более традиционной версии, кольчатые черви (Annelida) — ближайшие родственники как членистоногих, так и онихофор. Позднее некоторые авторы стали говорить о конвергентной эволюции, считая, что членистоногие ближе к нематодам (имея общие с ними способности к линьке), чем к аннелидам. В любом случае предполагается монофилия членистоногих как типа[35][36]. Две вероятные родословные определяют два вышестоящих таксона, к которым можно было бы причислить членистоногих: членистые (Articulata) и линяющие (Ecdysozoa)[37].
Классификация подтаксонов внутри группы членистоногих также неоднозначна. 4 основные группы (ракообразные, хелицеровые, трахейнодышащие и вымершие трилобитообразные) имеют ранг подтипов[38]. Кроме них, есть и ряд ископаемых групп высокого ранга (в большинстве своём из нижнего кембрия), которые трудно отнести к какой-либо подгруппе либо по причине несхожести с известными группами, либо из-за неясности их родственных взаимоотношений.
Многоножек и насекомых часто объединяют в одну группу неполноусых. Некоторые последние исследования, однако свидетельствуют, что многоножки ближе к ракообразным, чем к насекомым.
Здесь приведено несколько наиболее известных классификаций, в которых указанные основные группы различным образом объединяются в подтипы (или типы) и надклассы.
Основана на гипотезе о полифилетическом происхождении членистоногих. В связи с этим, тип членистоногих здесь разбивается на три самостоятельных типа[39].
Тип Хелицеровые
Тип Ракообразные
Тип Трахейные
Основана на гипотезе о монофилетическом происхождении членистоногих. При этом тип членистоногих объединяется с онихофорами и тихоходками в группу Lobopoda[8][40].
Подтип Трилобитообразные (Trilobitomorpha — парафилетический таксон)
Подтип Хелицеровые (Chelicerata)
Подтип Жвалоносные (Mandibulata)
Подтип Неполноусые (Atelocerata) или трахейные
До недавнего времени вышеприведённая система была более или менее общепризнанной. Сейчас она подвергается интенсивному пересмотру. Так, всё большее подтверждение получает концепция «Pancrustacea», основанная на том, что ракообразные и насекомые (вместе образующие Pancrustacea (англ.)русск.) — более близкие родственники, чем многоножки и насекомые (объединяемые в других классификациях в группу Atelocerata)[38]:
EuarthropodaChelicerata (Хелицеровые)
Myriapoda (Многоножки)
Cirripedia (Усоногие)
Remipedia (Ремипедии, слепые ракообразные)
Collembola (Коллемболы, или ногохвостки)
Branchiopoda (Жаброногие)
Cephalocarida (Цефалокариды)
Malacostraca (Высшие раки)
Insecta (Насекомые)
Членистоногих содержат в научных целях для проведения лабораторных исследований и в качестве декоративных домашних животных. Многие виды пауков, насекомых, ракообразных и многоножек успешно размножаются в неволе. Размножение некоторых эффектных видов жуков и бабочек поставлено на коммерческую основу[41]. Многие виды раков и креветок разводят в качестве источника пищи[42].
Согласно одной из гипотез, членистоногие возникли ещё до кембрийского взрыва. Сторонники этой гипотезы считают первыми известными членистоногими представителей эдиакарской биоты Parvancorina и Spriggina, живших около 555 млн лет назад. Уже в раннекембрийских (539—541 млн лет) слоях в Китае обнаружены явные мелкие членистоногие с раковинами примерно как у двустворчатых моллюсков. Самые ранние найденные окаменелости трилобитов датируются ранним кембрием (530 млн лет назад), но большое разнообразие видов и всемирное распространение даже ранних трилобитов приводит к выводу, что они к тому времени существовали уже давно. Повторное исследование окаменелостей фауны сланцев Бёрджес (возрастом около 505 млн лет) выявило много членистоногих, которых невозможно отнести ни к одной из известных групп. Это положило начало новому витку дебатов по поводу кембрийского взрыва. Окаменелость Marrella из той же фауны предоставила исследователям первое ясное свидетельство линьки.
Самая ранняя окаменелость ракообразного датируется кембрием, 513 млн лет назад, а чуть позже, 500 млн лет назад, уже обнаруживаются десятиногие ракообразные, напоминающие креветок. С ордовикского периода и далее ископаемые ракообразные встречаются достаточно часто.
К типу членистоногих относятся самые ранние известные наземные животные. Они датируются поздним силуром, около 419 млн лет назад. Вероятно, животные этого типа оставили и наземные следы возрастом около 450 млн лет. У членистоногих имелось несколько преадаптаций для освоения суши, в том числе суставчатый экзоскелет предоставляющий защиту от иссыхания и силы тяжести, а также способы передвижения, независимые от наличия воды. В то же время гигантские пресноводные ракоскорпионы стали рекордсменами по размеру среди членистоногих, достигая длины в 2,5 м.
Самый древний известный представитель паукообразных, Palaeotarbus jerami, жил около 420 млн лет назад (силурийский период). Attercopus fimbriunguis, живший 386 млн лет назад (девонский период), обладал первыми известными органами для выделения паутиноподобных нитей, но паутинных бородавок у него не было, и поэтому его нельзя причислить к истинным паукам (которые впервые появляются в позднекарбоновых отложениях, 299 млн лет назад). Из юрских и меловых пород известно уже много окаменелостей пауков, включая представителей многих современных семейств. Первые известные скорпионы с лёгочным дыханием датируются началом каменноугольного периода.
Древнейшее известное насекомое, Rhyniognatha hirsti, и одно из двух древнейших известных шестиногих (наряду с Rhyniella praecursor) было обнаружено в райниевом кремне в 1919 году. Окаменелости датируются ранним девонским периодом, около 410 млн лет назад. Rhyniognatha питалась, предположительно, спорофиллами сосудистых растений или же могла быть хищником. Возможно это было одно из первых членистоногих, обладающих крыльями.
节肢動物是動物的一类/门,由昆虫纲、甲壳纲、蛛形纲等外骨骼動物組成被稱为节肢动物门(学名:Arthropoda)的分類單位。在動物界中所屬物種最多的一門,已被人類命名的昆蟲類就有超過75萬種[1] [2] [3]。除昆蟲外,常見的蝦、蟹、蜘蛛、蜈蚣及已滅絕的三葉蟲都屬於节肢動物。
节肢動物的特點為其分節的肢體,以及主要成份為α-甲殼素的角質層。甲壳生物的角質層中也包括了碳酸鈣,是生物礦化(英语:biomineralization)的產物。
其身體表面有幾丁質組成的外骨骼;隨着身體的長大,需要經歷蛻皮的階段,將舊的外骨骼脫掉。
身體兩側對稱,身體分節,像環節類動物,但部分體節融合成特別部位,如頭部及胸部。有些節肢動物,例如蜘蛛,頭部及胸部進一步融合成頭胸部。每個特化部位的機能亦作相應的變化。
身體,取體節的反復構造,被外骨骼包圍而形成身體表面的體節單位。體節之間關節可動。根據種類的不同,也有如果體節裡面特定的東西組合能相合一下繼續的外骨骼被覆蓋等,外表上某在構成機能上的單位。譬如,有分成頭部,胸部,腹部3部分,或頭胸部,腹部的2部分的身體稱呼,這個作為每節肢動物的各分類群的特徵採用。
身體的附肢,例如足部、觸角、口器等都分節。 附肢的功能亦高度特化,負責感覺、行走、游泳、進食、以至生殖等。
從各體節,有腳出。腳同樣用外骨骼覆蓋,腳上也有關節。
一部分的分類群,成為由蛋孵化的幼體性成熟的成體的過程,有進行顯著地形態變化的生物變態。甲殼類和昆蟲類等。
节肢动物体内有腹神经索,并且在每个體節中,连着一对神经节,感觉神经和运动神经从神经节连接到身体其他部分。
節肢動物通常分為5亞門,其中一個已滅絕:
其中六足亚门和多足亚门又统称单支类(Uniramia(英语:Uniramia))。
像螃蟹、海螯蝦、淡水龙虾、蝦等甲壳亚门生物長久以來就已是人類料理的一部份,目前也有不少已經商業飼養再販售[5]。昆蟲的營養價值至少和肉類相當,在歐洲、印度以外的許多國家(伊斯蘭文化例外)會生食或是熟食昆蟲[6][7]。烹調過的捕鳥蛛(會去掉其防御用的高度刺激性觸毛)是柬埔寨的一項美味[8][9][10],也是委内瑞拉南部Piaroa印第安人的食物。人類也會無意間食用了其他食物中的昆蟲[11],因此食品安全法規會訂定不同食物中允許被昆蟲污染的程度。為了供人類食用而刻意飼養节肢动物或是其他小動物(稱為minilivestock),已開始出現在畜牧业中,而且是對生態友善的概念[12]。商业蝴蝶繁殖(英语:Commercial butterfly breeding)提供鳞翅目給蝴蝶温室、学校及研究機構,做為教育展览或是研究用。
不過节肢动物對人類食物最大的貢獻是幫助植物传粉:2008的研究調查了100項联合国粮食及农业组织認定作為食物的植物,估計传粉的經濟價值為1530億歐元,約是2005年農業食物生產產值的9.5%[13]。除了授粉外,蜂族會製作蜂蜜,這也是快速興起的產業及國際貿易項目[14]。
紅色的胭脂虫生長在中美洲,對阿茲特克及瑪雅文明有重要的經濟價值[15],當中美洲在被西班牙殖民的時候,胭脂虫是墨西哥第二大的出口[16],目前又開始受到重視[17]。馬蹄蟹的血液內含有凝血劑鱟血液細胞溶解產物(英语:Limulus Amebocyte Lysate),現在用來檢查抗生素以及洗腎機器中是否有危險的细菌,也用來檢測脊髓性腦膜炎以及一些癌症[18]。法醫昆蟲學利用节肢动物的相關證據判斷死者的死亡時間,死亡地點,有時甚至也包括死因[19]。近年來昆蟲也用來做為藥品或是其他醫療用物質的潛在來源[20]。
節肢動物結構相對簡單,而且可以在陸地上及水中活動,因此有些机器人設計開始考慮模仿節肢動物的結構。分節提供的冗餘性可以讓節肢動物及仿生學机器人在部份肢體受損或是移除時仍會正常移動[21][22]
|last=值左起第5位存在換行符 (帮助) 节肢動物是動物的一类/门,由昆虫纲、甲壳纲、蛛形纲等外骨骼動物組成被稱为节肢动物门(学名:Arthropoda)的分類單位。在動物界中所屬物種最多的一門,已被人類命名的昆蟲類就有超過75萬種 。除昆蟲外,常見的蝦、蟹、蜘蛛、蜈蚣及已滅絕的三葉蟲都屬於节肢動物。
节肢動物的特點為其分節的肢體,以及主要成份為α-甲殼素的角質層。甲壳生物的角質層中也包括了碳酸鈣,是生物礦化(英语:biomineralization)的產物。

節足動物(せっそくどうぶつ)とは、動物界最大の分類群で、昆虫類、甲殻類、クモ類、ムカデ類など、外骨格と関節を持つグループである。種多様性は非常に高く、陸・海・空・土中・寄生などあらゆる場所に進出して様々な生態系と深く関わり、現生種は約110万種と名前を持つ全動物種の85%以上を占める[2] 。なお、いわゆる「虫」の範疇に入る動物は当動物門のものが多い [3]。
節足動物の多様性は高く、分類によって様々な外見を持つ。体の表面はクチクラ(キチン質とタンパク質等)でできた外骨格でおおわれる。成長に伴い体のサイズが大きくなるときには、脱皮により古い外骨格は脱ぎ捨てられ、新しい外骨格が形成される。体は体節の繰り返し構造、つまり体節制をとり、体の表面を被う外骨格も体節単位になっている。各体節からは、それぞれ一対の関節肢と呼ばれる付属肢が出ている。
関節肢も体と同様に外骨格で覆われ、途中に関節がある。関節肢は分類によって歩脚・遊泳脚・鋏・鎌・顎・触角・鰓など様々な功能に応じて色んな形に特化する。 関節肢は内肢と外肢をもつ二叉型が基本形であるが、現生節足動物の中にこの性質を明瞭に保留するのは甲殻類だけで、ほとんどの分類群は内肢しか見られない単枝型である。また、絶滅した三葉虫とマーレラなど、カンブリア紀の殆どの節足動物の歩脚は二叉型である。
体節の間は関節状に可動であることが多い。ただし、複数体節の融合や分化など、いわゆる異規体節制がある程度以上発達し、例えば頭部や頭胸部はそれぞれの群で独特の複数体節が融合(合体節)してできたものであり、顎や触角などは頭部に含まれる体節の付属肢に由来する。こうして種類によって体節のうちの特定のものが組み合わされてひとつづきの外骨格で覆われる場合が多く見られ、外観上あるいは機能上の単位を構成する。例えば、体を頭部、胸部、腹部の3部、または頭胸部、腹部の2部に分けて呼ぶ場合があり、これは節足動物の各分類群ごとの特徴として用いられる。
多くの左右相称動物と同様、節足動物は体腔を持ち、消化管は体の前後に貫通し、いわゆる口と肛門という2つ開け口を持つ。体節に貫通する紐状の心臓と中枢神経はそれぞれ体の背面と腹面に付く。
循環系は開放血管系であり、細胞外液、リンパ液や血液という区別は存在せず、リンパ液と血液の役割を兼ねている血リンパが心臓と組織の間隙(血体腔)に流れる。心臓の伸縮や体の運動によって血リンパは心臓の動脈から体の静脈と呼吸器などの器官を通り、心門を通じて再び心臓に戻る。リンパ液の中に免疫系に関わる血球がある。
神経系として、1対の神経索が体の前後に走り、体節に合わせて分節し、左右の連絡でつながるはしご形神経系がある。頭部ないし頭胸部に備える脳は前大脳・中大脳・後大脳という3つの神経節からなる。この脳は体節と共に数対の神経節から融合した結果であり、そこから食道を囲んだ食道神経環は体の神経節に繋がる。クモ綱や昆虫の様に、各部位の体節の融合が強く、神経が集中してはしご形が不明瞭な場合もある。
節足動物は様々の方法を通じて周りの環境を感知する。体表は常に剛毛(感覚毛)をもち、種によっては触覚・振動・音・水流や気流・温度・匂いや味・化学物質など視力以外の感覚を持つ。 鋏角類以外の節足動物は頭部に触角という関節肢をもち、ほとんどの場合は重要な感覚器官である。触角のない鋏角類の中でも、ウデムシやサソリモドキの様に一部の歩脚が特化して感覚器官として使うことがある。
他にも昆虫の口器にある顎肢は嗅覚や味覚に関わり、サソリの櫛状板・ヒヨケムシのラケット器官・一部の昆虫と甲殻類の後端にある尾毛も感覚器官であり、コオロギやキリギリスの前足・バッタの胸部と腹部の間に両側・カマキリの後胸部腹面には聴覚に関わる特殊な器官を持つ。
節足動物は、単眼(ocelli)と複眼(compound eye)という2つの種類の眼を持ことが基本形と思われる。分類によって両方を備え、片方しか持たず、複眼の単眼化、稀に視力が失なわれた種類もある。また、単眼の中に元々単眼だったものは背単眼、複眼から退化したものは側単眼と呼ばれる。
複眼は図形認識能力を備え、多数の個眼という構成単位からモザイク画の様な視覚を産生する。単眼は主に明暗を感知する能力を持つが、一部のクモの単眼は図形認識能力を持ち、中でもハエトリグモの視力は非常に発達し、内部の網膜は自由に動かすことができる。
六足類と甲殻類は基本的に単眼と複眼を両方備える。多足類は背単眼を持たず、中にゲジは複眼、他のものは複眼由来の側単眼を持つ。鋏角類の中でカブトガニ綱は単眼と複眼を両方備え、クモ綱は複眼を持たず、背単眼と側単眼(中眼と側眼)を混在し、或いは片方のみを持つ。また、カンブリア紀の古生物から始め、三葉虫類や節足動物と思われるアノマロカリス類も発達した複眼を持つ。
関節は柔らかい膜で繋がり、2つの関節の外骨格内側に付着する筋肉で運動する。胴体の関節は様々な方向で湾曲できることが多いが、関節肢の関節はほとんど1対の蝶番に固定されて一つの平面上でしか動かない。そのため節足動物の関節肢、特に基部は多数の節に分かれて様々な動きに対応する。また、外骨格は体の内側へ延長し、いわゆる内突起(apodeme)で筋肉に付着面を提供することも多い。例えばカニの鋏の中では可動指の関節に繋がり、大量の筋肉が付着する板状の内突起が見られる。
また、体中のリンパ液の水圧を変えて関節肢を動かすこともある。この様な特徴はクモ綱に多く見られ、原始的な運動方法と思われる。
節足動物は様々な生息環境に進出し、それに応じて多様な呼吸様式を持つ。主なパターンとして陸生のものは気管や書肺、水生のものは鰓。稀に呼吸器官を持たず、体表でガス交換を行う種類もある。
求愛行動・交尾或いは交接・メイトガード・護卵など、節足動物は分類によって様々な繁殖行動を持つ。原則として有性生殖を行う卵生動物であるが、サソリやアブラムシなどから単為生殖・卵胎生の例も知られる。
成長に伴い古い外骨格の下で新しい外骨格を形成し、脱皮を通じて古い外骨格から抜け出して成長する。新しい外骨格は柔らかく、固くなるのも時間が掛かり、脱皮直後の節足動物は無防備である。そのため節足動物の脱皮は常に隠れ場所で行うことが多い。中でも古い外骨格を摂食して栄養を回収する種類や、一部のクモは脱皮直後のメスを狙って交接することが知られる。
節足動物の幼生は基本的に成体と似たような外見を持つだが、甲殻類のノープリウス幼生や昆虫の幼虫など、成長の過程で著しく形態が変化する変態を行う分類群も少なくない。昆虫の成虫になる脱皮は羽化と呼ばれる。多足類は成体になるまで脱皮に伴って体節と脚の対数を増やす。
ハエトリグモの求愛ダンス
幼生を背負するサソリの雌
羽化直後のアブラゼミ成虫
ヨーロッパミヤマクワガタの生活環
鰓曳動物門など
節足動物門
節足動物がどのような祖先から生まれて来たかについては、長く議論されていた。 以前は、前口動物であること、体節制を持つことなどの共通点から、環形動物に近縁であると考えられ、体節動物(Articulata)としてまとめられた。しかしその後では、環形動物はトロコフォア幼生を共有する軟体動物と類縁があると考えられており(冠輪動物)、体節制という共通点は単なる原始的な共通先祖性質、または収斂進化の結果と思われる。
側節足動物(Parathropoda)という便宜的なまとめをされたこともある舌形動物、有爪動物、緩歩動物との類縁関係を含め、この3つの動物門がヘッケル派の系統樹では前口動物の上の方に置かれるという考えあった。しかし、分岐分類学による検討や分子遺伝学による情報から、舌形動物はウオジラミに近縁の節足動物だと解明され、緩歩動物も他の動物門より節足動物に近縁とも思われ、側節足動物は多系統となり、否定された。
現在は、節足動物・有爪動物・緩歩動物の単系統性が定説となり、まとめて汎節足動物(Panarthropoda)になる。これらは更に鰓曳動物と線形動物などに加えて脱皮動物(Ecdysozoa)というグループになる。
アノマロカリスとオパビニアをはじめとして、カンブリア紀の化石からは、節足動物であるとも思われるものの、現在の分類では所属場所が見あたらない、最初期の節足動物の系統発生を示唆するものがある。その中でも、アノマロカリス類と節足動物の類縁関係が注目されている(アノマロカリス類#分類及び恐蟹綱#系統関係を参照)。同時期の節足動物の化石記録のうち、三葉虫類は最も早期で、おそよ5億2,100万年前に出現するが、5億3,700万年前にも遡る節足動物の生痕化石(Rusophycusなど)が見つかっており、更なる早期な起源を示唆する[1]。
現生の節足動物は、鋏角亜門(クモ綱、カブトガニなど)・多足亜門(ムカデ、ヤスデなど)・甲殻亜門(甲殻類)・六脚亜門(昆虫など)という4つの亜門に分類されている。化石種まで範囲を広げれば、三葉虫(三葉虫形亜門)という過去の大グループや幾つの分類が明らかになっていない種も存在した。
特に注目されるのは、前端体節の融合による頭部(鋏角類の場合は前体)の付属肢である、その構造は亜門によって以下のパターンがある。
系統について、かつては形態に従って多足類と六脚類は近縁とされてきた。しかし、分子遺伝学的情報は多足類と六脚類の直接の関連を支持せず、むしろ六脚類は側系統である甲殻類から分岐したという結果が出ている。また、節足動物全体の中ではウミグモ類の位置が問題となっている。鋏角類に含めるのが従来の判断であるが、これをそれ以外の節足動物全部と対置すべきとの説もある。また、三葉虫類をはじめてとして、幾つの絶滅した分類群と現存分類群の類縁関係については、未だに定説がない。アノマロカリス類やオパビニアなど恐蟹綱(Dinocaridida)のものは、節足動物全般の初期脇道系統(ステムグループ節足動物)として考えられる。
†Megacheira綱(側系統群?)
†三葉虫形亜門
ヒゲエビ亜綱
分子系統学、分岐分類学以前には、形態に基づく以下の分類が使用されていた。流通している書籍と文献にもこの分類にしたがっているものも多い。よって参考・比較のため、また生物学史上の意義もあり、以下に併記する。
節足動物(せっそくどうぶつ)とは、動物界最大の分類群で、昆虫類、甲殻類、クモ類、ムカデ類など、外骨格と関節を持つグループである。種多様性は非常に高く、陸・海・空・土中・寄生などあらゆる場所に進出して様々な生態系と深く関わり、現生種は約110万種と名前を持つ全動物種の85%以上を占める 。なお、いわゆる「虫」の範疇に入る動物は当動物門のものが多い 。
절지동물(節肢動物, 문화어: 마디다리동물, 영어: arthropod)은 절지동물문(節肢動物門, Arthropoda)에 속하는 동물의 총칭으로 곤충과 거미, 갑각류 등을 포함한다. 절족동물(節足動物)이라고도 한다. 현존하는 동물의 80% 이상이 절지동물문에 포함된다. 현재까지 알려진 절지동물의 수는 약 100만 종 이상이다. 또한 캄브리아기 초기부터 절지동물의 화석으로 발견된다.
호흡기·운동용 부속지·입틀·감각기의 발달 정도에 따라 생활하는 방식에 많은 차이가 있다. 바다에서는 새우나 게로 대별되는 갑각류가 주류를 이루며, 육지에서는 거미류나 곤충류가, 하늘에서는 곤충류만이 살고 있다. 생식 기간 외에는 단독 생활을 하는 종류가 많다. 그러나 많은 개체들은 먹이를 얻기 위하여 일시적 또는 영속적으로 모이기도 하는데, 특히 개미나 벌 집단에서 이러한 것이 발달되어 완벽한 사회생활의 형태를 보여준다. 이들 집단은 먹이를 얻기 위해 먼 곳까지 이동하기도 하지만, 대부분 일정한 지역에서 먹이를 구하며, 그 중에는 분포 지역에 세력권을 형성하는 경우도 있다.
먹이를 얻는 방법으로, 거미류에서는 그물과 같은 포획 장치를 이용하며, 또 매우 재미있는 예로 개미 등의 사회생활을 하는 곤충들은 버섯류를 재배하거나 다른 동물을 기르기도 한다. 천적이 많은 반면 방어 방법도 발달하여, 분비물로 관을 만들어 그 속에 살거나 튼튼한 집 또는 고치를 만드는 종류도 있다. 또한 독샘이나 독침, 부속지가 변형된 집게발이나 발톱 등의 무기로 적극적인 공격을 하기도 한다. 그리고 동식물·먼지·바위와 비슷한 형태나 색깔의 의태 또는 몸을 보호하기 위한 보호색이 발달된 종류도 있다.^^
대부분 암수딴몸으로서 체내 수정을 한다. 곤충류나 갑각류 중에는 양성 생식과 단위 생식을 교대하는 종류가 알려져 있는데, 이 밖에 수컷이 드물거나 없기 때문에 단위 생식을 하는 종류도 있다.
한편, 개미나 벌 등 사회 생활을 하는 곤충에서는 번식 능력을 갖는 암수컷의 개체 외에도 생식 능력이 없이 특정한 일만을 계속하는 개체도 있다. 절지동물은 생식 습성이 매우 다양한데, 특히 독립적으로 생활하는 종에서는 배우자를 얻기 위해 경쟁·유인·잠복 등 성의 본능적인 행동이 발달되어 있다. 대부분 난생이지만, 일부 난태생인 것도 있다. 특히 기생성의 무리에서는 유생 생식·유체 성숙, 다배 현상과 같은 특수한 예도 알려져 있다. 알의 대부분은 중황란으로서, 표면의 세포질만이 난할이 진행되는 표할을 한다.
이들은 각기 다른 보육 형태를 엿보인다. 즉, 알을 낳은 뒤 무책임하게 방치해 두는 원시형이 있는가 하면, 어미가 알을 품는 것, 유충이 부화하여 곧 먹이를 얻을 수 있도록 적당한 장소에 알을 낳는 것, 유충이 먹을 수 있도록 정교한 집을 지어 먹이를 저장하는 것, 또는 유충에게 먹이를 공급해 주는 것 등 여러 모습을 보인다. 유충의 발달 형식은 일반적으로 무변태·불완전 변태·완전 변태로 나뉘는데, 이 밖에 몸의 생김새가 조금씩 변화하는 개형 변태, 몸의 길이는 변화하나 몸의 생김새는 변화하지 않는 정형 변태 등으로 구별된다.잘 기억해라!!
체제는 환형동물에서 볼 수 있는 체절 구조가 기본을 이루나, 체표에 키틴질의 외골격이 있다는 점이 다르다. 또 좌우 대칭으로 대부분 머리·가슴·배의 세 부분, 또는 머리가슴과 배의 두 부분으로 나뉘어 있다. 부속지는 각 체절마다 한 쌍씩 존재하나 몸의 부분에 따라 모양과 기능이 다르다. 모든 종에서 공통적으로 나타나는 체제는 몸 앞쪽 끝에 입, 뒤쪽 끝에 항문, 배쪽에 부속지, 내부에서는 등쪽에 심장, 중축에 소화관 등이 위치한다는 점이다. 부속지의 대부분은 2교차형이 기본이다. 물 속에 사는 절지동물은 아가미로 산소를 받아들이는 반면, 육지에 사는 것들은 숨관 또는 책허파(서폐)라는 특수한 호흡기가 발달되어 있어 그것으로 산소를 얻는다. 아가미는 호흡의 표면적을 넓히기 위해 실 모양, 잎사귀 모양, 나뭇가지 모양으로 되어 있으며, 숨관은 각 기관 사이에 그물 모양으로 분포되어 있다.
순환계는 개방혈관계로서, 혈액은 심장에서 각 대동맥을 거쳐 기관 사이사이로 운반된다. 그리하여 온몸을 순환한 정맥피는 아가미를 가진 종류에서는 아가미 혈관에 의하여 아가미로 보내져 동맥피가 된 뒤 심장으로 들어간다. 그러나 숨관을 가진 종류에서는 정맥피가 숨관 속의 공기와 직접 가스교환을 일으켜 동맥으로 전환된다.














.jpg)
.png)


















_2.jpg)
















Ventral.jpg)


.jpg)













Ventral.jpg)


.jpg)












_3.jpg)























.png)

.png)
.jpg)























.png)
.jpg)







_micrograph.jpg)



.jpg)
.jpg)












































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}