The Rotifera (="wheel bearers") are a group of tiny animals first observed by early microscopists in the late 1600s. Around 2,000 rotifer species have been described (Segers 2008). Most species are in the range of 0.1 to 1 mm in length, although a few species may reach 2 to 3 mm. Different rotifer species display a striking variety of body forms and the morphology of individuals may be further altered (e.g, by growth of spines) in response to ecological cues indicating the presence in the environment of particular types of prey or predators (Wallace and Snell 2001 and references therein). Most rotifers are solitary, but there are a small number of colonial (mostly sessile, i.e. attached to the substrate) species (reviewed in Wallace 1987). Although there are both solitary and colonial sessile rotifers, most rotifers are motile and very active. (Wallace and Snell 2001 and references therein; Wallace 2002; Brusca and Brusca 2003)
The anterior end of a rotifer bears a ciliary organ known as the corona. In action, the movements of the coronal cilia can give the impression of a pair of rotating wheels, giving this phylum its name. The ventral appendage known as the "foot" secretes a sticky cement for temporarily attaching the rotifer to a substrate. In larvae of sessile rotifers, the cement forms a bond with the substrate that is not easily broken and if sessile rotifers are dislodged they do not reattach (Wallace 1980). Most rotifer species display eutely, i.e., the adults of a given species all have the same fixed number of cells (in the case of rotifers, around a thousand cells, about a quarter of which make up the nervous system). (Wallace and Snell 2001 and references therein; Wallace 2002; Brusca and Brusca 2003)
Rotifers are best known from freshwater, but some live in damp soil or moss and there are many marine species, although only a fraction of these are exclusively marine (Wallace and Snell 2001 and references therein).
Parthenogenesis is very common among rotifers in general and is the exclusive mode of reproduction among the bdelloid rotifers. The bdelloid rotifers, which account for around a quarter of the roughly 2000 described rotifers (Segers 2008), have attracted particular attention from researchers because the bdelloid clade is the most ancient group of animals known in which males are unknown and reproduction is strictly parthenogenetic (i.e., eggs develop without fertilization). (Wallace and Snell 2001 and references therein; Wallace 2002; Brusca and Brusca 2003)
In addition to their striking lack of males, bdelloids have attracted attention for the fact that most of them can undergo anhydrobiosis, slowly drying out until they resemble a wrinkled barrel, with the head and foot retracted into the animals's trunk (Wallace and Snell 2001). Desiccated bdelloids may be revived after many years in this state. Wilson and Sherman (2010) proposed that the ability of bdelloids to dry up, as well as to be transported long distances by air currents in their desiccated state, may allow them to escape fungal parasites in both time and space. An important and well supported hypothesis for the widespread evolutionary maintenance of sex in organisms in general is that sexual recombination creates genetic variation that allows organisms to stay ahead of their parasites in a never ending coevolutionary arms race. Wilson and Sherman suggest that the alternative mechanism they have documented for bdelloid rotifers to escape their parasites may help explain how this clade could persist for tens of millions of years in the absence of sex.
The Rotifera were formerly believed to be sister to the endparasitic Acanthocephala. A variety of analyses now strongly suggest that the Acanthocephala are in fact a clade of rotifers, most likely sister to the bdelloids (Sørensen and Giribet 2006).
The Rotifera (="wheel bearers") are a group of tiny animals first observed by early microscopists in the late 1600s. Around 2,000 rotifer species have been described (Segers 2008). Most species are in the range of 100 to 1000 µm (=0.1 to 1 mm) in length, although a few species may reach 2 to 3 mm. Different rotifer species display a striking variety of body forms and the morphology of individuals may be further altered (e.g, by growth of spines) in response to ecological cues indicating the presence in the environment of particular types of prey or predators (Wallace and Snell 2001 and references therein). Most rotifers are solitary, but there are a small number of colonial (mostly sessile, i.e., attached to the substrate) species (reviewed in Wallace 1987). Depending on the species, colonies may range in size from fewer than 5 individuals to large colonies of 50 to 200 individuals or, in the case of a few species, more than a thousand individuals. Although there are both solitary and colonial sessile rotifers, most rotifers are motile and very active. (Wallace and Snell 2001 and references therein; Wallace 2002; Brusca and Brusca 2003)
The anterior end of a rotifer bears a ciliary organ known as the corona. In action, the movements of the coronal cilia can give the impression of a pair of rotating wheels, giving this phylum its name. The ventral appendage known as the "foot" usually has 2 toes (range 0 to 4) and pedal glands, with ducts exiting near the toes, that secrete a sticky cement for temporarily attaching the rotifer to substrates. In larvae of sessile rotifers, the cement forms a bond with the substrate that is not easily broken and if sessile rotifers are dislodged they do not reattach (Wallace 1980). Most rotifer species display eutely, i.e., the adults of a given species all have the same fixed number of cells (in the case of rotifers, around a thousand cells, about a quarter of which make up the nervous system). The rotifer's pharynx is modified as a "mastax" consisting of sets of internal jaws. (Wallace and Snell 2001 and references therein; Wallace 2002; Brusca and Brusca 2003)
Rotifers are best known from freshwater, but some live in damp soil or moss and there are many marine species, although only a fraction of these are exclusively marine (see review of saltwater rotifers by Fontaneto et al. 2006). Densities in freshwater generally range up to around 1000 rotifers per liter, but with abundant food densities can exceed 5000 individuals per liter and, reportedly, in some water bodies at certain times densities may exceed 100,000 individuals per liter (Wallace and Snell 2001 and references therein).
Rotifers vary in their feeding habits. Some rotifers are ciliary suspension feeders; others grasp prey (mainly small animals, but some plant material as well) with protrusible, pincer-like mastax jaws; and still others draw prey into a funnel-shaped trap formed by spines or setae on the corona (Brusca and Brusca 2003). A few rotifers have evolved symbiotic relationships with other species, for example living on the gills of crustacean arthropods or on snail egg masses (see May 1989 for a review of the literature on commensal and parasitic relationships between rotifers and other organisms). Although parasitism is not common among rotifers in general, there is one major exception: The Acanthocephala, until recently generally treated as a phylum but now believed to be a clade of parasitic rotifers, most likely sister to the free-living bdelloids (Garcia-Varela and Nadler 2006; Min and Park 2009; Witek et al. 2009), are obligate intestinal parasites of vertebrates as adults, with arthropod intermediate hosts.
Most rotifers are oviparous, releasing eggs outside their bodies, but a a few are ovoviviparous, retaining the embryo inside the body until it hatches. Although most rotifers other than the bdelloids are dioecious (i.e., have both males and females), with the exception of the genus Seison males are generally reduced in abundance, size, and complexity. Parthenogenesis is very common among rotifers in general and is the exclusive mode of reproduction among the bdelloid rotifers. The bdelloid rotifers have attracted particular attention from researchers because the bdelloid clade is the most ancient group of animals known in which males are unknown and reproduction is strictly parthenogenetic (i.e., eggs develop without fertilization). Bdelloids account for nearly a quarter of the roughly 2,000 described rotifers (Segers 2008). The life cycle of some rotifer species may include both a parthenogenetic (amictic) phase and a mictic phase in which haploid eggs are produced, some of which may parthenogenetically produce haploid males, which can then fertilize haploid eggs with their mitotically produced haploid sperm. (Wallace and Snell 2001 and references therein; Wallace 2002; Brusca and Brusca 2003)
In addition to their striking lack of males, bdelloids have attracted attention for the fact that most of them can undergo anhydrobiosis, slowly drying out until they resemble a wrinkled barrel, with the head and foot retracted into the animals's trunk (Wallace and Snell 2001). Desiccated bdelloids may be revived after many years in this state. Wilson and Sherman (2010) proposed that the ability of bdelloids to dry up, as well as to be transported long distances by air currents in their desiccated state, may allow them to escape fungal parasites in both time and space. An important and well supported hypothesis for the widespread evolutionary maintenance of sex in organisms in general is that sexual recombination creates genetic variation that allows organisms to stay ahead of their parasites in a never ending coevolutionary arms race. Wilson and Sherman suggest that the alternative mechanism they have documented for bdelloid rotifers to escape their parasites may help explain how this clade could persist for tens of millions of years in the absence of sex.
The Rotifera were formerly believed to be sister to the endparasitic Acanthocephala, with the two groups together forming a clade that has been referred to as the Syndermata. A variety of analyses now strongly suggest that the Acanthocephala are in fact a clade of rotifers, most likely sister to the bdelloids, making the name Syndermata a junior synonym of Rotifera (Sørensen and Giribet 2006). Based on a large-scale analysis of molecular data (amino acid sequences), Witek et al. (2009) concluded that Syndermata and Gnathostomulida together comprise a monophyletic clade known as the Gnathifera. The Gnathifera was originally proposed as a monophyletic clade based on jaw morphology and may also include the Micrognathozoa and, conceivably, the Cycliophora (Funch et al. 2005; Sørensen and Giribet 2006).
Wallace and Snell (2001) provide a thorough review of rotifer biology, including a dichotomous key to families. Smith (2001) provides a dichotomous key to the freshwater genera of the United States. Segers (2008) reviews the global diversity of rotifers.
Rotifera, of die raderdiertjies, is ’n filum van mikroskopiese en byna-mikroskopiese diertjies met ’n pseudoseloom (skynbuikholte). Hulle is vir die eerste keer in 1696 beskryf, deur eerw. John Harris. Ander vorme is later, in 1703, deur Antonie van Leeuwenhoek beskryf.[1]
Die meeste raderdiertjies is sowat 0,1–0,5 mm lank, hoewel hul lengte kan wissel van 50 μm tot meer as 2 mm.[2] Hulle kom wêreldwyd algemeen in varswateromgewings voor. Daar is ’n paar soutwaterspesies, byvoorbeeld die genus Synchaeta.
Sommige is vryswemmend en eintlik plankton, terwyl ander op ’n substrata voortkruip of sittend is binne byvoorbeeld buise wat aan ’n substrata vas is. Sowat 25 spesies is koloniaal, soos Sinantherina semibullata. Raderdiertjies is ’n belangrike deel van die varswatersoöplankton: hulle is ’n groot voedselbron en baie spesies dra by tot die ontbinding van organiese materiaal.[3] Die meeste kom globaal voor, maar daar is sommige inheemse spesies, soos Cephalodella vittata in die Baikalmeer.[4]
|work= (help) Rotifera, of die raderdiertjies, is ’n filum van mikroskopiese en byna-mikroskopiese diertjies met ’n pseudoseloom (skynbuikholte). Hulle is vir die eerste keer in 1696 beskryf, deur eerw. John Harris. Ander vorme is later, in 1703, deur Antonie van Leeuwenhoek beskryf.
Rotatorilər (lat. Rotifera və ya lat. Rotatoria) — 1500-ə qədər növü olub, əksəriyyəti şirin sularda, bir qismi isə dənizlərdə yaşayır. Bədən ölçüləri 0.04-2.5 mm-ə qədərdir.
Rotatorilər qabarıq, yumurtaşəkilli və ya iyşəkilli bədən formasına malikdir. Plankton formaların bədən örtüyü üzərində müxtəlif çıxıntılar vardır.Bəzi rotatorilərin örtüyü həlməşikşəkilli maddədən ibarətdir.[1] Bu canlıların əksəriyyəti sərbəst hərəkətlidir, bəziləri isə oturaq həyat tərzi keçirir.
Rotatorilərin bədəni baş, gövdə, quyruq şöbə və ya ayaqdan ibarətdir. Ağız dəliyi baş şöbənin qarın hissəsində yerləşir. Ağızın önündə və arxa tərəfində iki kirpik həlqəsi yerləşir. Bəzi rotatorilərin baş şöbəsində topa şəklində hissetmə qılları, bir və ya iki ədəd piqmentli gözcük olur. Gövdə qalın kutikula ilə örtülmüşdür. Bir çox rotatorilərdə kutikula müxtəlif törəmələrlə təchiz olunmuş möhkəm zireh əmələ gətirir. Zireh formasına, quruluşuna və şəklinə görə müxtəlif olur. Boru rotatorilərin müdafiə olunmasına xidmət edir. Quyruq şöbə və ya ayaq bədənin çıxıntısı olub, əyilmək qabiliyyətinə malikdir. Bu hissə çəngəl şəklində iki yerə haçalanır. Haçaların qurtaracağı yapışqanlı maddə ifraz edən vəzilərlə təchiz olunmuşdur. Bu orqanlar vasitəsilə oturaq həyat tərzi keçirən rotatorilər müəyyən subsrata yapışa bilirlər.Rotatorilərdə dəri-əzələ kisəsi yoxdur.
Həzm sistemi üç hissədən ibarətdir. Ağız ən uca yaxın fırlanan aparatın kirpiki tacları arasında, qarın tərəfdə yerləşir. O ağız boşluğuna, ağız boşluğu isə getdikcə genişlənib əzələvi udlağa keçir. Udlağın divarlarında yerləşən iki cüt möhkəm dişcikli xitin lövhədən ibarət çeynəyici aparatı vardır. O bir cüt çəkiccik və bir cüt zindancıqdan ibarətdir. Rotatorilərin qidasını əsasən qamçılılar və birhüceyrəli yosunlar təşkil edir. Rotatorilərin mədəsində qida çox az müddətdə qalır. Qidanın tutulduğu andan defekasiyaya qədər 2-dən 20 dəqiqəyə qədər vaxt keçir. Sinir sistemi - nisbətən sadədir. Bir udlaqüstü sinir düyünündən və ondan hər tərəfə gedən sinirlərdən ibarətdir. Digər sinirlərdən xeyli böyük olan iki sinir beyindən arxaya doğru, bağırsağın yan tərəfləri ilə bədən boyu ayağa qədər uzanır. Rotatorilərdə hiss orqanları rolunu çıxıntılar yerinə yetirir Onlardan ikisi qarında, biri isə beldə tək olur. Əksər rotatorilərdə bir cüt və ya yalnız bir göz olur. Gözlər bilavasitə udlaqüstü düyündə və ya onun üstündə yerləşir. Göz bir olduqda o, piqment qədəhlərinin qabarıq tərəfləri ilə qarşı-qarşıya duran iki gözcüyün birləşməsidir. Rotatorilərdə ''qan-damar'' və tənəffüs sistemi yoxdur. İfrazat sistemi protonefridial tipdədir. Bağırsağın yanlarında, bədən boşluğunda iki zəif şaxələnmiş kanal vardır. Onlar kloakaya açılır. Kanallardakı şaxələrin ucları qapalı olur. Şaxələrin sayı 4-dən 50-yə qədər olur. Cinsi sistem - Rotatorilər ayrıcinslidir və cinsi dimorfizmlə fərqlənirlər. Dişilər erkəklərdən daha çox təsadüf olunur. Yumurtalıq iki hissəyə bölünmüşdür. Erkəklər dişilərdən nisbətən kiçikdir, onların bağırsağı reduksiya etmişdir. Erkəklər mayalanmadan dərhal sonra ölür. Erkək cinsi sistem yeganə toxumluqdan ibarətdir.[2]
Rotatorilərin həyat dövriyyəsi əsl cinsi nəslin partogenetik nəsil ilə növbələşməsindən ibarətdir. Heteroqoniya xarakteri daşıyır. Qışlamış yumurtadan yazda partogenetik dişilər çıxır, onlar isə partogenetik dişilərə başlanğıc verir. Partogenetik dişi nəsillərindən biri indiyə qədər qoyulmuş yumurtalardan həcmcə 2-3 dəfə kiçik yumurtalar qoymağa başlayır. Yumurtalardan tezliklə dişi nəsil çıxır və tezliklə cinsi nəslin dişiləri ilə cütləşir. Partogenetik yol ilə erkəkləri törədən dişilər cütləşmədən sonra xüsusi mayalanmış yumurtalar əmələ gətirir. Onlar iridir, bir neçə xüsusi möhkəm təbəqə ilə örtülüdür və sakitlik dövrü keçirir. Mayalanmış yumurtalar gələcəkdə partogenetik dişilərin ilk nəslini verir. Rotatorilərin dövrülüyü bir çox növlərdə, çoxalmanın xarakteri ilə əlaqədar olmayan mövsümi səbəblər üzündən dəyişilir. Rotatorilərin erkək fərdlərində regenerasiya qabiliyyəti vardır.
Rotatorilər kosmopolit heyvanlardır.Əksər rotatorilər şirin sularda yaşayır və onlar dib və plankton formalara bölünür. Dibdə yaşayan rotatorilər əksər hallarda suyun dibində sürünür və ya üzür, oturaq növləri də vardır. Tipik plankton rotatorilərdə heyvanın bədən səthini böyüdən müxtəlif tipli çıxıntılar vardır. Şirinsu rotatoriləri daha iri şirinsu orqanizmləri üçün qida mənbəyi kimi mühüm rol oynayır. Dəniz rotatoriləri nisbətən azdır. Quru, eləcə də bəzi su rotatoriləri məhv olmadan tamamilə quruyana qədər dözürlər. Bu zaman onlar anabiotik vəziyyətə düşür, lakin ehtiyatla islatdıqda dirilirlər. Rotatorilərin dirilməsi ilk dəfə 1701-ci ildə Levenhukk tərəfindən kəşf edilmişdir. Bundan əlavə, qurumuş rotatorilər 4 saat ərzində -270 °C-yə qədər aşağı temperatura və 5 dəqiqə ərzində +100 °C-yə qədər yuxarı temperatura dözə bilir.
Rotatorilərin əmələ gəlməsi haqqında müxtəlif fərziyyələr mövcud olmuşdur. Uzun müddət Rotatoria-ların Polychaeta-ların sürfəsi olan troxofor oxşarlığı əsasında olan həlqəvi qurdların neotenik sürfələri hesab edirdilər. Rotatoria qrupunun təkamül yolu isə düzbağırsaqlı turbellarilərdən başlanır. Rotatoria-ların turbellarilərlə yaxınlığının onlarda protonefridinin primitiv quruluşu və ağızın qarın tərəfdə yerləşməsi sübut edir.
Onlardan Xəzər dənizində 67 növü qeyd edilmişdir, əsasən Volqa çayının dənizə tökülən hissəsində rast gəlinir. Onları dənizə çay axını gətirir. Platyas quadricornis, Platyas patulus və s. əsasən şirin sularda, bəzi növləri isə Xəzərin dayaz və duzluluğu az olan yerlərində yaşayırlar. Şortəhər sularda yaşayan Trichocersa caspisa kimi növlər Xəzər dənizinin endemik rotatoriləridir.
Araz su anbarının zooplanktonunda 38 növ rotatori yayılmışdır1[3]. Növmüxtəlifliyinin zənginliyi bir tərəfdən sututarın ekoloji-coğrafi vəziyyəti, digər tərəfdən isə rotatorilər faunasının ətraflı öyrənilməsi ilə bağlıdır. Son məlumatlara görə, Mingəçevir su anbarında cəmi 15 növ rotatorinin yayıldığı müəyyən edilmişdir[4].
Rotatorilər (lat. Rotifera və ya lat. Rotatoria) — 1500-ə qədər növü olub, əksəriyyəti şirin sularda, bir qismi isə dənizlərdə yaşayır. Bədən ölçüləri 0.04-2.5 mm-ə qədərdir.
Els rotífers (Rotifera del llatí rota,roda, i ferre, dur) constitueixen un embrancament d'animals pseudocelomats pràcticament microscòpics. Foren descrits per primera vegada per John Harris l'any 1696 (Hudson i Gosse, 1886). S'atribueix erròniament a Leeuwenhoek la primera descripció d'aquests animals però Harris ja havia realitzat esbossos l'any 1703.
La majoria de rotífers mesuren entorn els 0,1-0,5 mm de longitud i són comuns en aigua salabrosa tot i que algunes espècies habiten en aigües salades.
El nom es va posar en referència al seu aparell succionador, la corona, que recorda una corona reial o una serra dentada circular. La corona està composta de cilis que, al moure's, creen petits corrents que atreuen les partícules de l'entorn cap a ella. De la corona passen a una faringe i d'aquesta a l'aparell mastegador denominat màstax, on es trituren les partícules succionades, per a després ser digerides a l'estómac i finalment expulsats els residus per la cloaca. La corona també la utilitzen per a desplaçar-se per l'aigua, a manera d'hèlice. En l'extrem oposat a la corona es distingeix un peu (generalment un parell de peus) amb el qual poden aferrar-se al substrat.
Existeixen rotífers amb una cutícula relativament gruixuda i rígida que donen a l'animal una aparença capsular; uns altres són flexibles i d'aspecte i moviment vermiforme. Encara que la gran majoria de rotífers són nedadors, alguns són sèssils i viuen en l'interior de tubs o càpsules gelatinoses. Al voltant de 25 espècies són colonials (com Sinantherina semibullata).
En la majoria d'espècies els mascles són escassos, de manera que la reproducció per partenogènesi és bastant comuna. A més existeix un marcat dimorfisme sexual, ja que els mascles existents solen tenir l'aparell digestiu atrofiat, i la seva existència es redueix a la producció d'esperma. La vida d'una femella sol ser d'una o dues setmanes.
Algunes espècies són xerobionts, és a dir, els seus ous fertilitzats s'enquisten i creen formes de resistència capaces de sobreviure als períodes de sequera i dessecació en els quals l'hàbitat aquàtic desapareix per motius estacionals. Al ser rehidratats emergeixen ràpidament (en menys de dues hores) com organismes amb capacitat natatòria. Algunes espècies produeixen un derivat del sucre trehalosa amb el qual creen una substància gelatinosa que ajuda a resistir la falta d'aigua ambiental.
El següent cladograma, adaptat de [1], mostra les afinitats dels rotífers:
GnathiferaRotifera
Els rotífers (Rotifera del llatí rota,roda, i ferre, dur) constitueixen un embrancament d'animals pseudocelomats pràcticament microscòpics. Foren descrits per primera vegada per John Harris l'any 1696 (Hudson i Gosse, 1886). S'atribueix erròniament a Leeuwenhoek la primera descripció d'aquests animals però Harris ja havia realitzat esbossos l'any 1703.
La majoria de rotífers mesuren entorn els 0,1-0,5 mm de longitud i són comuns en aigua salabrosa tot i que algunes espècies habiten en aigües salades.
Vířníci (Rotifera, Rotatoria) je kmen převážně sladkovodních, mikroskopických živočichů. Některé druhy žijí i v půdě nebo v moři. Jsou důležitou součástí planktonu. Je známo okolo 2000 druhů, v Česku žije asi 600 druhů.
Při zběžném pohledu se vířníci podobají velkým nálevníkům, ale jsou na rozdíl od nich mnohobuněční. Většina vířníků je menší než 0,5 mm, největší jsou velcí asi 2 mm. Nejvýznačnějším znakem vířníků je zatažitelný vířivý aparát na přední části těla. Dva věnce brv neustále kmitají a přihánějí potravu k ústnímu otvoru. Slouží také jako hnací orgán při plavání. Dalším charakteristickým znakem je žvýkací hltan (mastax).
Tělní dutina je pseudocoel, vylučovací soustava je tvořena protonefridiemi. Vířníci mají stálý a neměnný počet buněk (eutelie), podobně jako hlísti. Povrch těla kryje intracelulární kutikula (syncytiální epidermis). Některé druhy vytvářejí na povrchu svého těla pevné krunýře z desek a ostnů. Většina vířníků má na opačném konci těla nohu ukončenou vidličkou. Pomocí ní lezou píďalkovitě po dně nebo se odrážejí a skáčou. Do vidličky ústí lepkavé žlázy, jimiž se vířníci dočasně přilepují na předměty ve vodě. Někteří se přichytávají na různé předměty ve vodě, jiní se ve vodě volně vznášejí. Dostanou-li se do nepříznivých podmínek, vytvoří si na povrchu těla pevnou schránku, ve které přežívají (stav anabiózy).
Jsou to gonochoristé, ale samci jsou menší než samice a někdy zcela (u pijavenek) chybějí. Populace se pak rozmnožuje partenogenezí. Při páření samec vychlípitelným penisem vstříkne sperma přímo do tělní dutiny samice. U některých vířníků se setkáváme s heterogonií, střídání generací s haploidními vajíčky a partenogenetických generací s diploidními vajíčky.
Vířníci (Rotifera, Rotatoria) je kmen převážně sladkovodních, mikroskopických živočichů. Některé druhy žijí i v půdě nebo v moři. Jsou důležitou součástí planktonu. Je známo okolo 2000 druhů, v Česku žije asi 600 druhů.
Hjuldyr er en række af mikroskopiske dyr, der lever i vand og fugtig jord.
Der findes omkring 2000 arter af hjuldyr, fordelt på tre klasser.
Række: Rotifera
Hjuldyr er en række af mikroskopiske dyr, der lever i vand og fugtig jord.
Der findes omkring 2000 arter af hjuldyr, fordelt på tre klasser.
Rädertierchen (Rotifera, veraltet Rotatoria) sind 0,1 bis 0,5 Millimeter (Acanthocephala bis 70 Zentimeter) lange vielzellige Tiere mit genetisch festgelegter, gleich bleibender Anzahl von Zellen (Eutelie). Am Kopf befinden sich bewegliche Wimpernkränze, das Räderorgan. Bislang sind weltweit etwa 2000 teilweise sehr verschiedene Arten beschrieben, von denen etwa 550 in Deutschland vorkommen.
Rädertiere treten in vielen Lebensräumen auf. Auf dem Land, in Bäumen, in feuchtem Moos oder zwischen Bodenpartikeln sind sie ebenso zu Hause wie im Meer oder im Süßwasser. Dabei macht ihnen die Kälte der Antarktis ebenso wenig etwas aus wie die Hitze von Thermalquellen.
Die verschiedenen Rädertiergattungen leben entweder dauerhaft an Pflanzen festsitzend oder freischwebend im Wasser oder Detritus.
Das Aussehen der Rädertiere ist sehr vielgestaltig, dennoch lässt sich der Körper grob in drei Abschnitte gliedern:
Rädertiere haben, bedingt durch ihre Körperform, verschiedene Möglichkeiten der Fortbewegung: Gleiten, schwimmen, spannerartig kriechend, mit den Wimpern des Kopfes laufend oder strudelnd.
Um Trockenzeiten überstehen zu können, geben bdelloide Rädertiere einen Teil ihrer Körperflüssigkeit ab und schrumpfen zu einer kugelförmigen Gestalt zusammen. In diesem sehr widerstandsfähigen Dauerstadium, auch Trockenstarre genannt, können sie bis zu vier Jahre überleben.
Am Rumpf oder Fuß können einzelne Eier oder Eipakete hängen, die eine ähnlich hohe Widerstandskraft gegen Umwelteinflüsse haben wie die ausgewachsenen Tiere.
Der für die Rotifera typische Kaumagen (Mastax, Pl. Mastaces) zeigt kieferartige, komplexe Gerüste aus einzelnen stäbchen-, schild- oder plattenförmigen, chitinhaltigen Hartteilen, den sogenannten Trophi (Sg. Trophus). Sie können gegeneinander bewegt werden und können je nach Form verschiedene Funktionen, wie Einsaugen, Zermahlen, Zerquetschen oder Ergreifen von Beute wahrnehmen. Bei den Monogononta sind die Trophi je nach Art stark unterschiedlich geformt und bilden ein wichtiges Merkmal zur genauen taxonomischen Bestimmung einzelner Arten. Bei den Arten der Bdelloida zeigen sie unter den einzelnen Arten eine eher gleichartige Struktur, es wird auch von den typischen ramaten Mastaces oder den ramaten Trophi der Bdelloida gesprochen (englisch ramate trophi)[1].
Die meisten Arten ernähren sich von Algen oder Detritus. Brachionus calyciflorus beispielsweise ernährt sich von einzelligen Algen und Bakterien. Es strudelt diese mit seinem Räderorgan herbei. Lindia torulosa ernährt sich von Blaualgen, deren Fäden sie mit dem Kauapparat abkneifen. Es gibt aber auch räuberische Rädertierchen, wie die Floskularien (Floscularia), die sehr kleine Lebewesen und Partikel aus dem durch ihre Ruderorgane aufgewirbelten Wasser fangen, oder die Cephalodella die ebenfalls räuberisch leben. Pleurotrocha petromyzon ernährt sich wiederum aasfressend von toten Wasserflöhen[2]. Die Collotheca, Cupelopagis vorax aber auch Stephanoceros fimbriatus haben eine andere Jagdweise. Sie fangen ihre Nahrung mittels weit aufgesperrten Trichtern am oberen Körperende. Die beiden Arten der urtümlichen Gattung Seison leben im Meer als Parasiten auf Arten der Krebsgattung Nebalia. Auch Süßwasserformen wie Proales werden als Parasiten angesehen, da sie in Algen wie Volvox und Vaucheria leben und sich von diesen ernähren.
Die Beziehungen zwischen Räuber und Beute können komplex sein. Das Rädertierchen Asplanchna brightwellii ernährt sich räuberisch von Plankton, darunter auch anderen Rädertierchen der Gattung Brachionus. In Wasser, das Asplanchna enthält, entwickelt Brachionus lange Stacheln als Körperfortsätze, die den Räuber beim Fangen behindern, die Entwicklung dieser Stacheln wird nur durch den Räuber induziert und unterbleibt sonst. Als auslösender Faktor konnte eine chemische Substanz (vermutlich ein Peptid) identifiziert werden, die Asplanchna in das Wasser abgibt. Solche Substanzen mit hormonähnlicher Wirkung auf eine andere Art werden Kairomone genannt.[3]
Die verschiedenen Gattungen der Rädertiere nehmen unterschiedliche Möglichkeiten der Fortpflanzung wahr. Unter günstigen Bedingungen (meist in den Sommermonaten) erfolgt eine ungeschlechtliche Vermehrung (siehe auch: Parthenogenese), unter ungünstigen Bedingungen (meist im Herbst) findet die geschlechtliche Fortpflanzung statt.
Einige Arten wie etwa Adineta vaga sind in der Lage, genetisches Material anderer Lebewesen in ihr Erbgut aufzunehmen, was die Nachteile ausgleicht, die sich aus ungeschlechtlicher Fortpflanzung ergeben. Diese Fortpflanzungsstrategie wurde bei Rädertierchen erstmals durch Eugene Gladyshev nachgewiesen[4] und war zuvor nur bei Bakterien bekannt. Darüber hinaus verzichtet A. vaga auf sexuelle Fortpflanzung und Meiose.[5]
Rädertiere haben unterschiedliche Lebenserwartungen. Der Durchschnitt liegt bei etwa einer Woche.
Im sibirischen Permafrostboden wurden Individuen der Gattung Adineta entdeckt, die seit 24.000 Jahren in Kryptobiose überlebt hatten und sich nach dem Auftauen durch Parthenogenese vermehrten. Ihre Fähigkeiten, ihre Zellen und Organe abzuschirmen und eine lange kryogene Konservierung zu überleben, sind von großem Interesse für Wissenschaftler. Vor dieser Entdeckung waren die einzigen anderen Tiere, von denen man wusste, dass sie so lange überleben können, Spulwürmer, die nach 42.000 Jahren wiedererweckt werden konnten.[6]
Als erster wissenschaftlicher Beobachter der Rädertierchen gilt Antoni van Leeuwenhoek, der die Rädertiere mit seinem selbstgebauten Mikroskop beobachtete. Da dessen Vergrößerung nicht sehr stark war, konnte er die flimmernde Mundöffnung nur ungenau beobachten, beschrieb jedoch ihr räderförmiges Aussehen.
Heute werden die Rädertierchen zwar weiterhin als Tierstamm akzeptiert, nach phylogenetischen Untersuchungen sowohl der Morphologie als auch anhand von molekularbiologischen Vergleichen müssen jedoch die früher ebenfalls als Tierstamm betrachteten Kratzwürmer (Acanthocephala) als Schwestergruppe der Bdelloida innerhalb der Rädertierchen angesehen werden.[7][8] Die umfassende Gruppe (Rotifera i. e. S., Seisonacea, Acanthocephala) wird gelegentlich "Syndermata" genannt, die meisten Bearbeiter bevorzugen aber, den Namen Rotifera im erweiterten Sinne für diese Gruppierung zu gebrauchen. Die Ordnung Seisonacea (mit der einzigen Familie Seisonidae, 2 Arten) wird von einigen Systematikern als einzige Ordnung (monotypisch) in eine eigene Klasse "Pararotatoria" eingeordnet.
Innerhalb der Rädertierchen werden folgende Verwandtschaftsverhältnisse als eine der aktuellen Hypothesen angenommen:
RädertierchenBdelloida und Kratzwürmer (Acanthocephala)
Früher wurden die Rädertierchen in eine Gruppe der „Schlauchwürmer“ oder auch Rundwürmer (Nemathelminthes) gestellt, ein Verlegenheits-Taxon ohne klare Abgrenzung, das in neueren Systematiken als nicht monophyletisch erkannt worden ist. Neuere Untersuchungen auf morphologischer, vor allem aber auf genetischer Basis[9] haben nun übereinstimmend ergeben, dass die Rädertierchen mit zwei kleinen, wenig bekannten Gruppen, den Kiefermündchen (Gnathostomulida) und den Micrognathozoa (mit der einzigen Art Limnognathia maerski) eine Klade bilden, die nach einem morphologischen Merkmal, dem ähnlichen Feinbau des Kieferapparats Gnathifera („Kieferträger“) genannt worden ist. Nächstverwandt wären die lange in ihrer Stellung rätselhaften Pfeilwürmer (Chaetognatha). Inzwischen wurde die fossile Art Amiskwia sagittiformis Walcott 1911 aus dem mittelkambrischen kanadischen Burgess-Schiefer als mögliche morphologische Zwischenform ausgemacht.[10] Die Zusammengehörigkeit der Gruppen wird auch durch den übereinstimmenden Bau der für die Entwicklung wichtigen Hox-Gene unterstützt.[11]
Der Stamm der Rotifera umfasst etwa 2030 Arten, von denen 1570 Arten auf die Unterklasse der Monogononta entfallen. Die weitere Untergliederung ist nach heutigen Kenntnisstand wie folgt:[12][13]
Das einzige bekannte Fossil aus dem Stamm Rotifera stellt der Fund eines Vertreters der Klasse Bdelloidea (Ordnung Bdelloida) in tertiärem Dominikanischen Bernstein dar. Zugleich liefert dieser Fund den Beweis, dass Parthenogenese seit mindestens 25 bis 40 Millionen Jahren existiert.[18]
Rädertierchen (Rotifera, veraltet Rotatoria) sind 0,1 bis 0,5 Millimeter (Acanthocephala bis 70 Zentimeter) lange vielzellige Tiere mit genetisch festgelegter, gleich bleibender Anzahl von Zellen (Eutelie). Am Kopf befinden sich bewegliche Wimpernkränze, das Räderorgan. Bislang sind weltweit etwa 2000 teilweise sehr verschiedene Arten beschrieben, von denen etwa 550 in Deutschland vorkommen.
De rajerdere (Letien: Rotifera) vörmen 'ne stam van klein mieëcellige organisme. Ze behuuere toet 't óngerriek Eumetazoa en häör lingdje bereik zelje 2 mm. De menkes zeen klènder en einveljiger geboedj es de vruikes. 'Ruum 2200 saorte zeen weitesjappelik besjreve.
Rajerdere waere gekènmirk door e träöt- toet bolvörmig lief, döks doorzichtig wie glaas, en dragen ane kop meistes 'nen dóbbele krans van trilhaor, die waere gebroek bieje vergaoring van voor enne veurtbewaeging. De ventraal móndjäöpening lèdj nao de farynx dae deens duit es 'ne kawmaag. De meiste rajerdere laeven in water. E klein aantaal laef inne baom.
Ónger druuegdje kónne rajerderen euvergaon nao 'n toestandj van sjiendoead: cryptobioos. In daen röstoestandj kónne rajerdere temperature van 270 graoje Celsius gedoerendje 5 menuut en 100 graoje gedoerendje inkel oer verdrage. Rajerdere vermeinigvöljige zich inkel generaties parthenogenetisch (door deiling) en ei maol de jaor geslechtelik.
De rajerdere (Letien: Rotifera) vörmen 'ne stam van klein mieëcellige organisme. Ze behuuere toet 't óngerriek Eumetazoa en häör lingdje bereik zelje 2 mm. De menkes zeen klènder en einveljiger geboedj es de vruikes. 'Ruum 2200 saorte zeen weitesjappelik besjreve.
Rajerdere waere gekènmirk door e träöt- toet bolvörmig lief, döks doorzichtig wie glaas, en dragen ane kop meistes 'nen dóbbele krans van trilhaor, die waere gebroek bieje vergaoring van voor enne veurtbewaeging. De ventraal móndjäöpening lèdj nao de farynx dae deens duit es 'ne kawmaag. De meiste rajerdere laeven in water. E klein aantaal laef inne baom.
Ónger druuegdje kónne rajerderen euvergaon nao 'n toestandj van sjiendoead: cryptobioos. In daen röstoestandj kónne rajerdere temperature van 270 graoje Celsius gedoerendje 5 menuut en 100 graoje gedoerendje inkel oer verdrage. Rajerdere vermeinigvöljige zich inkel generaties parthenogenetisch (door deiling) en ei maol de jaor geslechtelik.
Rotifera (lat. rota = kolo + -fer = nosač), obično nazivane životinje s kolima ili kolnjaci ili kolaste životinjice,[1] čine koljeno Rotifera, bosanski obično rotifere. To su organizmi, beskičmenjaci, Pseudocelomata mikroskopseke ili submikroskopskih dimenzija.
Prvi ih je opisao John Harris , 1696., a ostale oblike Antonie van Leeuwenhoek, 1703.[2] Većina rotifera le dugs od 0.1–0.5 mm, iako se njihova veličina može kretati od 50 µm do preko 2 mm,[1] i uobičajene su u slatkovodnom okruženju širom svijeta s nekoliko vrsta u slanim vodama.
Neke rotifere su slobodnoplivajuće planktonske, drugi se kreću vrpcom duž supstrata, a neki su sesilne, koji žive unutar cijevi ili želatinozni organizmi koji su pričvršćeni na supstrat. Postoji oko 25 kolonijalnih vrsta (npr. Sinantherina semibullata), bilo sjedeće ili planktonske. Rotiferi su važan dio slatke vode zooplanktona, glavni su izvor hrane i sa mnogim vrstama također doprinose razgradnji organske tvari u tlu.[3] Većina vrsta je kosmopolitska, ali postoje i neke endemske vrste, poput Cephalodella vittata u Bajkalskom jezeru.[4] No, nedavni dokazi putem barkodiranja sugeriraju da su neke 'kosmopolitske' vrste, poput Brachionus plicatilis, Brachionus calyciflorus, Lecane bulla, između ostalih, zapravo su kompleks vrsta.[5][6] U nekim novijim tretmanima, rotiere se postavljaju s Acanthocepha u većem kladusu zvanom Syndermata.
Građa rotifera je vrlo različita, ali se ugrubo na tijelu mogu uočiti tri dijela, pri čemu vrste koje žive na biljkama potsjećaju na crve, a vrste u vodi su kesaste.
Zavisno od tjelesne građe, kolnjaci se mogu kretati na više načina: klizanjem, plivanjem, puzanjem (skupljanjem i istezanjem) ili se uz pomoć treplji na roratornom organu. Tokom suše, rotifere imaju sposobnost anabioze. Iz tijela izbacuju tjelesnu tečnost i zbiju se u lopticu. U takvom obliku, zvanom i "suha ukočenost", mogu preživjeti i do četiri godine. Na trupu ili stopalu mogu ostati pričvršćeni paketi ili pojedinačna jaja koja imaju istu sposobnost reagiranja na loše ekološke uvjete kao i odrasle životinje.[7][8]
Rotiferai su vrlo otporne životinje i mogu preživjeti u vrlo različitim sredinama. Na kopnu, žive na stablima, u vlažnim mahovinama, kao i u pijesku ili između zrnaca tla, ali jednako tako nastanjuju i mora i slatkovodne sredine, gdje su važan izvor hrane većim životinjama. Dobro podnose hladnoćeu Antarktika i Arktika, kao ni visoke temperature termalnih izvora. Različiti rodovi kolnjaka žive ili trajno pričvršćeni na biljkama, slobodno plutaju vodom ili u detritusu.
Većina vrsta hrani se algama ili detritusom. Neke vrste, pomoću treplji i strujom vode uzimaju alge ili bakterije. Međutim, postoje i grabežljiva rotifere koje u vodenoj strujii love vrlo malehne jednočelijske životinje ili nihove dijelove. Vrsta Asplanchna birghtwelli, velika samo oko 1 mm također je grabljivac, tako što u vodu izlučuje osobit peptid. Druge vrste imaju drugačije načine lova, a neke parazitiraju na drugim životinjama.
Rotifere pojedinih rodova imaju različite načine razmnožavanja. U povoljnim okolnostima (uglavnom u ljetnim mjesecima) razmnožavaju se nespolno (partenogenezom), a kad su uvjeti nepovoljni (uglavnom u jesen), razmnožavanje je spolno.[9] Očekivani životni vijek pojedinih vrsta je različit, a u prosjeku je to oko jedne sedmice.
Utvrđeno je da veličina genoma vrste Adineta vaga iznosi oko 244 Mb.[10] Čini se da su genomi monogonota znatno manji od onih kod Bdelloida. Sadržaj nuklearne DNK (2C) u Monogononta u osam različitih vrsta četiri različita roda kretao se gotovo četverostruko, od 0,12 do 0,46 pg..[11] Veličina haploidnog, "1C" genoms vrsta roda Brachionus kreće se u rasponu od 0,056 do 0,416 pg.[12]
John Harris prvi je opisao rotifere (posebno bdeloidnne) 1696. godine kao "životinju poput velikog magota koja bi se mogla uvući u sfernu figuru, a zatim se ponovo ispružiti; kraj repa se pojavio kliješta poput onih kof uholadže".[2] 1702. Antonie van Leeuwenhoek dao je detaljan opis Rotifer vulgaris i kasnije opisano Melicerta ringens i druge vrste.[13] Također je bio prvi koji je objavio zapažanja o oživljavanju određenih vrsta nakon sušenja. Ostali promatrači opisali su druge oblike, ali tek kada je 1838. godine objavljena knjiga Christiana Gottfrieda Ehrenberga „Die Infusionsthierchen als vollkommene Organismen“, rotiferi su prepoznati kao višećelijske životinje.[13] Opisano je oko 2200 vrsta rotifera. U toku je rasprava o njihovoj taksonomiji. Jedani ih smještaju koljeno Rotifera, s tri razreda: Seisonidea, Bdelloidea i Monogononta.[14] Najveća skupina je Monogononta, sa oko 1500 vrsta, a slijedi je Bdelloidea, sa oko 350 vrsta. Postoje samo dva poznata roda sa tri vrste Seisonidea.[15]
Za Acanthocephala, koje su se prethodno smatrale zasebnim koljenom, dokazano je da su modificirane rotifere. Tačan odnos prema ostalim članovima koljene još nije riješen.[16] Jedna od mogućnosti je da su Acanthocephala bliže Bdelloidea i Monogononta nego Seisonidea; odgovarajuća imena i odnosi prikazani su u kladogramu ispod.
Rotifera su, strogo govoreći, ograničene na Bdelloidea i Monogonontu. Rotifera, Acanthocephala i Seisonida čine kladus zvani Syndermata.[17]
Iako se kolnjaci i danas smatraju koljenom, prema filogenetskim, morfološkim i molekularnobiološkim usporedbama danas ih se više ne smatra prirodnom skupinom (monofilija) jer se još jedno koljeno, Acanthocephala, smatra sestrinskom s redom Bdelloida iz koljena kolnjaka[18][19].
Rotifera Cuvier 1817 (2 classas, 8 òrdres, 33 familhas, 112 genres, 1816 espècias)
Los rotifèrs son d'organismes pichons (al maximum 3 mm), que lor cap e lor partida posteriora son pas cobèrts per la cuticula. Lor boca es enrodada de cilhas filtrant las particulas de noiridura que circulan dins l'aiga. An una multiplicacion asexuada, mas tanben se pòdon multiplicar de manièra sexuada, sustot dins de condicions de vida marridas.
Rotifera (lat. rota = kolo + -fer = nosač), obično nazivane životinje s kolima ili kolnjaci ili kolaste životinjice, čine koljeno Rotifera, bosanski obično rotifere. To su organizmi, beskičmenjaci, Pseudocelomata mikroskopseke ili submikroskopskih dimenzija.
Prvi ih je opisao John Harris , 1696., a ostale oblike Antonie van Leeuwenhoek, 1703. Većina rotifera le dugs od 0.1–0.5 mm, iako se njihova veličina može kretati od 50 µm do preko 2 mm, i uobičajene su u slatkovodnom okruženju širom svijeta s nekoliko vrsta u slanim vodama.
Neke rotifere su slobodnoplivajuće planktonske, drugi se kreću vrpcom duž supstrata, a neki su sesilne, koji žive unutar cijevi ili želatinozni organizmi koji su pričvršćeni na supstrat. Postoji oko 25 kolonijalnih vrsta (npr. Sinantherina semibullata), bilo sjedeće ili planktonske. Rotiferi su važan dio slatke vode zooplanktona, glavni su izvor hrane i sa mnogim vrstama također doprinose razgradnji organske tvari u tlu. Većina vrsta je kosmopolitska, ali postoje i neke endemske vrste, poput Cephalodella vittata u Bajkalskom jezeru. No, nedavni dokazi putem barkodiranja sugeriraju da su neke 'kosmopolitske' vrste, poput Brachionus plicatilis, Brachionus calyciflorus, Lecane bulla, između ostalih, zapravo su kompleks vrsta. U nekim novijim tretmanima, rotiere se postavljaju s Acanthocepha u većem kladusu zvanom Syndermata.

Un rotifero es un membro de un filo de animales multe peti, comun en lagos.
└─o Sindermato o Rotifero ├─o Monogononto │ ├─o │ │ ├─o Colotecaseo │ │ └─o Flosculariaseo │ └─o Ploimido └─o Lemniseo ├─o Bdeloideo └─o ├─o Sesonideo └─o Acantocefalo ├─o Arciacantosephalo │ ├─o Apororincido │ ├─o Moniliformido │ ├─o Oligacantorincido │ └─o Jigantorincido ├─o Eoacantosefalo │ ├─o Jiracantosefalo o Cuadrijirido │ └─o Neoecinorincido └─o Paleacantosefalo ├─o Polimorfido └─o Ecinorincido
Un rotifero es un membro de un filo de animales multe peti, comun en lagos.
Weldiarten (Rotifera of iar uk: Rotatoria) san en stam faan diarten mä amanbi 2000 slacher.
Weldiarten (Rotifera of iar uk: Rotatoria) san en stam faan diarten mä amanbi 2000 slacher.
Τα τροχοφόρα και ενίοτε τροχόζωα είναι μικροσκοπικά ζώα (40 μm – 3 mm), κοσμοπολίτικα με πάνω από 1.800 είδη, που ζουν κυρίως στα γλυκά νερά, ελεύθερα ή προσκολλημένα. Το διαφανές σώμα τους καλύπτεται από εφυμενίδα και στο πρόσθιο τμήμα φέρει ένα βλεφαριδοφόρο τροχοφόρο όργανο, που ουσιαστικά είναι βλεφαρίδες, των οποίων η συντονισμένη κίνηση δίνει την εντύπωση περιστρεφόμενου τροχού (rotifera).
Βδελοειδές τροχοφόρο
Brachionus quadridentatus
Τα τροχοφόρα και ενίοτε τροχόζωα είναι μικροσκοπικά ζώα (40 μm – 3 mm), κοσμοπολίτικα με πάνω από 1.800 είδη, που ζουν κυρίως στα γλυκά νερά, ελεύθερα ή προσκολλημένα. Το διαφανές σώμα τους καλύπτεται από εφυμενίδα και στο πρόσθιο τμήμα φέρει ένα βλεφαριδοφόρο τροχοφόρο όργανο, που ουσιαστικά είναι βλεφαρίδες, των οποίων η συντονισμένη κίνηση δίνει την εντύπωση περιστρεφόμενου τροχού (rotifera).
Βδελοειδές τροχοφόρο
Brachionus quadridentatus
Ротиферите (Rotifera, Rotatoria) се едни од најситните многуклеточни животни (метазои). Тоа се животни со карактеристичен сложен виличен апарат и цилијарен (трепчест) ротаторен орган кој се користи при локомоцијата и исхраната. Ротиферите припаѓаат во наттипот Platyzoa, што значи дека тоа се животни со сплескана форма на телото. Исто така, според веќе спомнатиот виличен апарат, тие се вбројуваат во групата на Gnathifera (виличести метазои).
Ротиферите се најчесто седентарни животни (некои повремено, а други постојано). По својата морфологија тие се најразлични.
Традиционално, ротиферите се сметаа за дел од типот наречен Aschelminthes (Nemathelminthes) кој се состоеше од животни со псевдоцелом. Современите филогенетски анализи не ја поддржуваат групата на Nemathelminthes и укажуваат на блиска врска помеѓу ротиферите, боцкоглавците (Acanthocephala), виличестите црви (Gnathostomulida), како и со неодамна опишаниот тип Micrognathozoa, притоа соединувајќи ги во групата Gnathifera (виличести животни). Обично боцкоглавците се сметаат за сестринска група на ротиферите, но ова не е сосема поддржано од молекуларните анализи, кои укажуваат на тоа дека боцкоглавците се високо организирани ротифери. Сепак, на оваа хипотеза и треба поддршка и од морфолошки податоци. Филогенетската позиција на гнатиферите останува несигурна, но најскорешните филогенетски анализи сугерираат дека тие се или основна група во Spiralia или монофилетска група заедно со гастротрихите и сплесканите црви.
Од 2003, во Rotifera постојат околу 1817 видови распоредени во пет реда и 34 фамилии. Тие се поделени во три класи: Seisonidea, Bdelloidea и Monogononta. Последната содржи околу 80% од сите познати видови и има најголем морфолошки диверзитет.
По големина, ротиферите може да варираат од помалку од 100 µm до 2,500 µm, но повеќето видови се големи од 150–500 µm. Според ова, ротиферите (заедно со гастротрихите) се најмалите метазои.
Најчесто телото е поделено на глава, труп и стапало, но овој основен градбен план може силно да варира. Најзабележлив орган во главениот регион е короната или ротаторниот апарат, кој/а се состои од метахроно трепкачки цилии поредени во различни снопчиња. Генерално, короната сочинува големо букално поле кое ја обиколува устата и циркумапикален сноп кој го обвива ацилијарниот апикален главин регион. Короната многу варира во својата градба. На пример, кај класата Bdelloidea, циркумапикалниот сноп е поделен медијално така што формира два трохални диска. При пливањето или исхраната, трепкањето дава илузија како да животното да носи два мали, ротирачки тркала (rota – тркало, fero – носи). Короната се користи за движење и исхрана. Трепкањето ги носи честичките од храна кон усниот отвор кој секогаш е лоциран (повеќе или помалку) вентрално. Трупот може да варира по форма – од многу елонгирана форма (некогаш со телескопски и ретракторни псевдосегменти) до поглобуларна форма. Стапалото се состои од еден до неколку псевдосегменти и често има две терминални завршоци (т.н. прсти) со отвори на адхезивни (цементни) жлезди на нивниот врв. Стапалото и прстите може да се редуцирани кај неколку родови и видови. Интегументот на ротиферите е од синцицијален тип, што значи дека епидермалните клетки не се одделени со клеточни мембрани. Надворешна кутикула отсуствува (која е присутна кај повеќето други инвертебрати). Наместо кутикула, ротиферите имаат внатрешноклеточна кончеста ламина. Дебелината на ламината е најразлична, а кај некои таксони, делови од неа се толку дебели што формираат масивен телесен оклоп. Таквите видови се лорикатни, за разлика од тие со потенка ламина кои се илорикатни.
Усниот отвор е сместен вентрално или апикално во короната. Тој води до голтка која содржи сложен мастикаторен (џвакачки) апарат кој се нарекува мастакс. Тој се состои од цврсти вилични делови трофи и од мали мускули кои ги поврзуваат виличните елементи. Трофите се состојат од четири главни елементи: парни рами, парни унци, парни манубриуми и непарен фулкрум. Исто така може да има и различни придружни елементи кои со заедничко име се нарекуваат епифарингеални елементи. Рамите се централни елементи во трофите и често се опремени со заби или дентикули. Тие се каудално споени со флексибилен лигамент, така што се способни да се отвораат и затвораат, со што ја раздробуваат храната. Единствениот непарен елемент – фулкрумот – се протега каудално од артикулациската точка на рамите. Унците се наоѓаат рострално или вентрално од рамите. Тие може да се стапчевидни со единечен остар заб или штитовидни со неколку цврсти заби. Унците се користат да ја заграбуваат храната и да манипулираат со неа или (кај некои предаторски ротифери) да го пенетрираат интегументот на пленот. Унците проксимално ги поврзуваат манубриумите. Тие најчесто се стапчевидни со добро развиена глава и се одговорни главно за движењето на унците. Морфолошките карактеристики на четирите главни елементи доста се разликуваат. Затоа, ротиферните трофи се поделени на девет различни типови, во зависност од големината и формата на главните склерити. Трофите се многу значајни за таксономијата на ротиферите – неколку фамилии и родови можат да се препознаат само по типот на вилици. Од голтката, краток хранопровод води до желудникот. Дигестивните ензими се произведуваат во синцициелни гастрични жлезди кои се отвораат во самиот желудник. По желудникот доаѓа цревото, кое се завршува со дорзална клоака.
Парови од протонефридии ја контролираат екскрецијата и одржувањето на осмотската рамнотежа. Секоја протонефридија се состои од еден или повеќе мултицилијарни терминални клетки и мултицилијарни канални клетки кои водат до собирачките цевчиња, кои, пак, ги носат екскретите до мочниот меур.
Женскиот репродуктивен систем се состои од еден или два синцициелни гермовителарии (составени од герминален регион и жолточник), опколени со фоликуларен слој. Класите Seisonidea и Bdelloidea имаат парни гонади, додека Monogononta поседуваат само еден гермовителариум. Може да постои и јајцевод кој се формира од фоликуларниот слој и ги поврзува гермовителариумот и клоаката. Машките репродуктивни органи сочинуваат непарен семеник и пенис.
Ротиферите се забележани во сите делови од светот. Тие се или вистински космополити, или, пак, ендемични видови. Конкретно, бделоидните родови Philodina и Rotaria и неколку моногононтски родови (Brachionus, Keratella, Lecane и Lepadella) ги содржат некои од најраспространетите видови забележани на повеќе места во светот. И другите видови се чести но имаат поограничено распространување, на пример, на евроазискиот континент или Холарктикот и пантропските региони. Веќе спомнавме дека некои видови се ендемични. На пример, не повеќе од 11 видови од родот Notholca се ендемични за Бајкалското Езеро. Меѓу клучните фактори за распространетоста на ротиферите се нивните криптобиотички способности, како и продукцијата на т.н. заспани јајце клетки. На тој начин, ротиферите се распространуваат на големи растојанија со помош на ветерот или водата, избегнувајќи ги неповолните животни услови.
Ротиферите (Rotifera, Rotatoria) се едни од најситните многуклеточни животни (метазои). Тоа се животни со карактеристичен сложен виличен апарат и цилијарен (трепчест) ротаторен орган кој се користи при локомоцијата и исхраната. Ротиферите припаѓаат во наттипот Platyzoa, што значи дека тоа се животни со сплескана форма на телото. Исто така, според веќе спомнатиот виличен апарат, тие се вбројуваат во групата на Gnathifera (виличести метазои).
Ротиферите се најчесто седентарни животни (некои повремено, а други постојано). По својата морфологија тие се најразлични.
Өйрөлмәктәр (лат. Rotifera, рус. Коловратки) — күп күҙәнәкле умыртҡаһыҙҙар тибы. Төп айырмаһы — кәүҙәнең башы өлөшөндә бырау һымаҡ әйләнеүсе керпексәләрҙән торған аппарат, ул туҡланыу һәм хәрәкәт итеү өсөн ҡулланыла.
Бөтә Ер шарында таралған. Бер нисә отряды, яҡынса 2 мең төрө билдәле, Башҡортостанда яҡынса 100 төрө бар. Кәүҙәһе башлыса оҙонса (оҙонлоғо 0,04—2,3 мм), баштан, кәүҙәнән һәм аяҡтан (ҡайһы берҙә редукцияға бирелгән) тора. Башында — аҙыҡ табыу һәм хәрәкәт итеү өсөн хеҙмәт иткән өйөрөлмәк аппарат (2 керпек тажынан тора). Күптәре панцирлы. Тән ҡыуышлығында мускулдар, аш һеңдереү, бүлеп сығарыу, нервы һәм енес системалары урынлашҡан. Аяғында цемент биҙҙәре булған сәтәше бар, ул хәрәкәт итеү һәм субстратҡа йәбешеү ағзаһы булып тора. Айырым енесле хайуан (ата заттар ваҡ). Йомортҡа һала, һирәгерәк тере бала тыуҙыра. Ҡайһы берҙәре бер енесле үрсеү юлын ике енесле менән берләштереү хас булһа, икенселәре тик бер енесле юл менән генә үрсей. Башлыса һыу хайуаны; бер күҙәнәкле ылымыҡтар, микроорганизмдар менән туҡлана. Һыу ятҡылыҡтарының үҙ аллы таҙарыу процестарында ҡатнаша.
Өйрөлмәктәр (лат. Rotifera, рус. Коловратки) — күп күҙәнәкле умыртҡаһыҙҙар тибы. Төп айырмаһы — кәүҙәнең башы өлөшөндә бырау һымаҡ әйләнеүсе керпексәләрҙән торған аппарат, ул туҡланыу һәм хәрәкәт итеү өсөн ҡулланыла.
Бөтә Ер шарында таралған. Бер нисә отряды, яҡынса 2 мең төрө билдәле, Башҡортостанда яҡынса 100 төрө бар. Кәүҙәһе башлыса оҙонса (оҙонлоғо 0,04—2,3 мм), баштан, кәүҙәнән һәм аяҡтан (ҡайһы берҙә редукцияға бирелгән) тора. Башында — аҙыҡ табыу һәм хәрәкәт итеү өсөн хеҙмәт иткән өйөрөлмәк аппарат (2 керпек тажынан тора). Күптәре панцирлы. Тән ҡыуышлығында мускулдар, аш һеңдереү, бүлеп сығарыу, нервы һәм енес системалары урынлашҡан. Аяғында цемент биҙҙәре булған сәтәше бар, ул хәрәкәт итеү һәм субстратҡа йәбешеү ағзаһы булып тора. Айырым енесле хайуан (ата заттар ваҡ). Йомортҡа һала, һирәгерәк тере бала тыуҙыра. Ҡайһы берҙәре бер енесле үрсеү юлын ике енесле менән берләштереү хас булһа, икенселәре тик бер енесле юл менән генә үрсей. Башлыса һыу хайуаны; бер күҙәнәкле ылымыҡтар, микроорганизмдар менән туҡлана. Һыу ятҡылыҡтарының үҙ аллы таҙарыу процестарында ҡатнаша.
किरीटी (रोटिफ़ेरा, Rotifera) स्वतंत्र रूप से रहनेवाले छोटे-छोटे प्राणी हैं। इनके शरीर के अगले भाग में एक रोमाभ (Ciliary) अंग होता है, जिसके रोमाभ इस तरह गति करते हैं कि देखनेवाले को शरीर के आगे चक्र (पहिया) चलता मालूम पड़ता है। इसीलिये इन्हें पहिएदार जंतु (ह्वील ऐनीमलक्यूल, Wheel animalcule) कहते हैं। अंग्रेजी नाम रोटिफ़ेरा का यही तात्पर्य हैं। इसीलिए इस वर्ग का नाम रोटिफ़ेरा या रोटेटोरिया रखा गया है।
किरीट अधिकतर साधारण स्वच्छ (अलवण) जल में रहते हैं। कुछ खारे पानी में रहते हैं और कुछ समुद्र में भी पाए जाते हैं। कुछ पृथ्वी पर नम स्थानों पर रहते हैं और कुछ काई (Moss) के पौधे की पत्तियों के अक्ष में रहते हैं। कुछ किरीटी परोपजीवी भी होते हैं। एक जाति घोंघा (Snail) के अंडों पर परोपजीवी होती है। इस तरह किरीटी ने विभिन्न प्रकार के निवासस्थान अपना रखे हैं। इनका वितरण भी विस्तृत है। ये संसार के सभी कोनों में पाए जाते हैं।
किरीटी मेटाज़ोआ में काफी छोटे जंतु हैं। इनकी लंबाई .04 से 2 मिलीमीटर तक होती हैं, परंतु अधिकतर किरीटी .5 मिलीमीटर से लंबे नहीं होते। ये प्रोटोज़ोआ से बड़े नहीं होते, इसलिए प्रारंभ में लोग इनको भी प्रोटोज़ोआ मान बैठे थे। इतने छोटे होते हुए भी इनके शरीर के भीतर अनेक जटिल इंद्रियतंत्र होते हैं, जिन्हें बिना सूक्ष्मदर्शी यंत्र से नहीं देखा जा सकता।
किरीटी का शरीर लंबाकार होता है। अध्ययन के लिए उसे तीन भागों में विभाजित किया जाता हैं। पहला, आगे का चौड़ा भाग हैं जिसपर रोमाभ अंग होता हैं। इस भाग को सर कहते है और रोमाभ अंग को मुकुट (कॉरोना, Corona)। सर के बाद का लंबा भाग धड़ कहलाता है और तीसरे भाग को दुम (या फुट भी) कहते हैं। साधारणत: किरीटी ऐसे ही होते हैं, परंतु कुछ ऐसे भी हैं जिन्होंने विशेष रूप धारण कर लिए हैं। कुछ थैली के आकार के होते हैं, कुछ गोल होते हैं (जैसे ट्रोकोस्फ़ियरा, Trochosphaera) और कुछ चौड़े होते हैं (जैसे ब्रैकियांसी, Brachioncy) और कुछ लंबे पतले होते हैं, जैसे (रोटेलिया, Rotalia)। कुछ किरीटियों का प्याले जैसा शरीर एक लंबे डंठल द्वारा पृथ्वी से जुड़ा रहता हैं। यदि किरीटी का शरीर आड़ा काटकर देखा जाए तो प्राय: गोल दिखाई पड़ेगा, परंतु कुछ किरीटियों में पार्श्वीय तथा कुछ में प्रतिपृष्ठीय दीवारें चिपटी होती हैं। अधिकतर किरीटी द्विपार्श्व सममिति (Bialateral Synmetry) वाले होते हैं, परंतु कुछेक बाहरी अंगों के कारण असममित मालूम पड़ने लगते हैं। उदाहरण के लिये किसी में पैर की दो अंगुलियों में से एक लंबी और एक छोटी होती हैं। कुछ किरीटियों का शरीर प्रतिपृष्ठीय दीवार की ओर झुका रहता है और किसी में पूरा शरीर सर्पिल होता हैं।
शरीर हलके पीले रंग के आवरण, बाह्यत्वक् या क्यूटिकिल (Cuticle) से ढका रहता है। बाह्यत्वक कुछ कड़ा होता है, इसीलिये शरीर का इधर-उधर मुड़ना संभव नहीं होता। इसीलिए कोशिकाभित्ति में प्राय: वलय होते हैं। कभी कभी वलय इतने गहरे होते हैं कि शरीर खंडदार मालूम होने लगता है। कुछ किरीटियों के धड़ का बाह्यत्वक् विशेषकर अधिक मोटा और कड़ा हो जाता है। इसको लौरिका कहते हैं। विपत्ति के समय शरीर का आगे का भाग लौरिका के भीतर समा जाता हैं। लौरिका का बाह्यत्वक् सादा होता है या उसपर षट्कोणीय अथवा अन्य नमूने बने रहते हैं।
किरीटी के शरीर के आगे के भाग को केवल सुविधा के लिए सर कहा जाता है। यह चौड़ा होता है और सामने चपटा। कभी कभी सामने का बीच का भाग उभड़ा रहता हैं। इसके चारों ओर रोमाभ होते हैं। रोमाभयुक्त भाग को मुकुट कहते हैं और उसके मध्य के रोमाभविहिन भाग को ऐपिकल फील्ड (Apical field)। ऐपिकल फ़ील्ड पर अनेक उभड़े अंग दिखलाई देते हैं। इनमें से कुछ ऐसे होते है जिनपर नीचे स्थित ग्रंथियों की नलिकाएँ खुलती हैं और कुछ संवेदक होते हैं जिनपर कड़े बाल होते हैं। अधिक किरीटियों में मुकुट गोलाकार होता है। कुछ जंतुओं में यह दो पिंडकों (लोब्स, lobes) में बँटा रहता है। डंठल से पृथ्वी पर अनुरक्त रहनेवाले किरीटियों में मुकुट प्याले की शक्ल का होता है और उसका स्वतंत्र भाग कई पिंडकों में विभाजित रहता हैं। डेलायड में द्विपिंडकी (बाइ लोब्ड, bilobed) मुकुट के बीच में एक प्रमुख उभाड़ होता है, जिसका उपयोग वह पृथ्वी या पौधे आदि की सतह पर चलनें में करता हैं। इस उभाड़ को रोस्ट्रम कहते हैं।
कारोना के रोमाभ एक साथ इस प्रकार गति करते हैं कि सामने पानी की लहरें बन जाती हैं। यह जल की लहरें खाद्य पदार्थ के जल में तैरते हुए टुकड़े मुँह तक ले आती हैं और खाद्य पदार्थ या तो मुँह में चला जाता हैं या उसे मुखांग पकड़ लेते हैं। अनेक किरीटियों में रोमाभ भोजन प्राप्त करने के मुख्य साधन होते हैं और अन्य सभी किरीटियों में ये भोजनप्राप्ति में सहायता देते हैं। मुकुट द्वारा पैदा की गई जल की लहरों से अन्य लाभ भी हैं। ये जानवरों के चारों ओर का पानी बदलती रहती हैं जिससे जानवर को ताजा आक्सिजन मिलता रहता हैं। स्वतंत्र रूप से तैरनेवाले किरीटियों में रोमाभ उन्हें तैरने में सहायता देते हैं। जल की लहरें शरीर के निकट एकत्र हुए उत्सर्जित (एक्स्क्रीटरी, excretory) पदार्थ बहा ले जाती हैं।
मुँह मुकुट के मध्य में प्रतिपृष्ठीय रेखा की ओर होता है। इसके नीचे का ऐपिकल फील्ड का भाग कुछ उठा रहता है, मानो वह निचला ओठ हो। किसी किसी किरीटी के ऐनिकल फील्ड में आँखें भी होती हैं। अधिकतर किरीटियों में आँखें मस्तिष्क पर स्थित होती हैं। आँखें या तो दो होती हैं या एक। किसी किरीटि में आँख तुंड (रोस्ट्रम, rostrum) पर भी स्थित होती है। आँख देखने में छोटे लाल चिह्न की भाँति होती हैं।
धड़ लंबाकार होता है या अनेक प्रकार से चपटा। यह सादा होता है या वर्मिका (Lorica) युक्त। वर्मिका सादी होती है या उसपर अनेक नमूने बने रहते हैं। किसी किसी में वर्मिका पर काँटे भी होते हैं। पैडलिया नामक किरीटी पर बड़े बड़े काँटे होते हैं जो शरीर के चलायमान पिंडकों पर स्थित रहते हैं। धड़ पर कुछ विशेष स्पर्शांग होते हैं। इनमें एक जोड़ा शरीर के दोनों बगल में होता हैं। इसे पार्श्वीय श्रृंगिका पृष्ठीय (लैटरल ऐंटेनी, lateral antennae) कहते हैं। एक श्रृंगिका पृष्ठीय तल पर होती हैं। इसे पृष्ठीय श्रृंगिका (डॉरसल ऐंटेना, dorsal antenna) कहते हैं। जिस स्थान पर धड़ और दुम मिलते हैं वहाँ मध्यपृष्ठीय (मिड-डॉर्सल, mid-dorsal) रेखा पर मलद्वार या गुदा स्थित हैं। धड़ पीछे की ओर पतला होता जाता हैं और दुम में मिल जाता है। कुछ किरीटियों में, विशेषकर मुकुटयुक्त किरीटियों में, धड़ और दुम बिल्कुल अलग मालूम पड़ते हैं।
कुछ किरीटियों में दुम छोटी और कुछ में बड़ी होती हैं। दुम के बाह्यत्वक् पर गहरे वलय होते हैं जिससे वह कई खंडों की बनी हुई मालूम पड़ती है। किरीटी दुम की सहायता से तल पर रेंगते हैं और तैरते समय दुम पतवार का कार्य करती हैं। पृथ्वी से जुड़े रहनेवाले किरीटी में दुम लंब डंठलाकार हो जाती है और जंतु को पृथ्वी से जोड़े रहती हैं। दुम के अंत में एक से चार तक नन्हें नन्हें चलायमान अंग होते हैं। जिन्हें अंगुली या टो (toes) कहते हैं। ये छोटे, तिकोने होते हैं, या पतले, लंबे काँटे जैसे। अंगुलियों के सिरों पर दुम के भीतर स्थित ग्रंथियों की नलिकाएँ खुलती हैं। ये ग्रंथियाँ चिपचिपा पदार्थ पैदा करती हैं, जो चलते (रेंगते) समय अंगुलियों को सतह से चिपकाने करता हैं।
किरीटी प्राय: पारदर्शी होते हैं। कोई कोई कुछ हलके पीले लगते हैं। यह इसलिए कि बाह्यत्वक् या बाहरी आवरण का रंग पीला-सा होता है। भूरे, लाल या नारंगी रंग के किरीटी भी मिलते हैं। यह रंग खाए हुए भोजन का होता है जो पारदर्शी से झलकता हैं।
किरीटी में लैंगिक द्विपरूता (सेक्सुअल डाइमॉर्फ़िज्म, Sexual dimorphism) भी मिलती हैं। केवल दो वर्गों (प्लायमा और सीयसोनेशिया) में नर तथा नारी दोनों एक जैसी होती हैं। शेष सब में नर छोटा और नारी बड़ी होती हैं। नर की बनावट भी साधारण होती हैं। डेलायड नामक एक गण (ऑर्डर) के किरीटियों में नर मिलते ही नहीं। केवल नारियाँ पाई जाती हैं और इनमें अनिषेकजनन (पारथिनाजेनिसस, Parthenogenesis) द्वारा बच्चे पैदा होते हैं।
शरीर की दीवार तथा आंतरंगों के बीच के स्थान को स्यूडोसीलोम (Pseudo-coelome) कहते हैं। केंचुए या मेंढक जैसे जानवरों में इस स्थान का सीलोम (Coelome) कहते हैं। सीलोम में शरीर की दीवार के अंदर की ओर मध्यजनस्तर (मीसोडर्म, Mesoderm) की एक परत होती है और उसी की एक परत आंतरगों पर। इस तरह सीलोम मध्यजनस्तर के बीच की गुहा है और स्यूडोसीलोम में मध्यजनस्तर की परतें नहीं होती। स्यूडोसीलोम एक तरल पदार्थ से भरी रहती हैं। इसमें कुछ बड़ी-बड़ी शाखादार कोशिकाएँ (सेल) भी होती हैं। इसमें शाखाएँ एक पतला जाल-सा बना डालती है। ये कोशिकाएँ कदाचित् कीटाणुओं को खा डालती हैं। इसलिए इनको फ़ैगोसाइट (Phagocyte) कहते हैं। कुछ लोगों का यह भी विचार है कि यह उत्सर्जन में सहायता देती हैं।
मुंह से प्रारंभ होकर आहारनाल गुदा पर बाहर खुलती है। मुँह पतले मुख नाल (बकल ट्यूब, Buccal tube) में खुलता है और मुखनाल ग्रसनी (फ़ैरिन्स, Pharynx) में। किरीटी की ग्रसनी सारे जंतुजगत् में विलक्षण है। यह बड़ी मांसल थैली होती है। इसके भीतर का अस्तर, जो बाह्यचर्म का बना होता है, भोजन चबाने का एक जटिल उपकरण है। इसके मैस्टैक्स कहते हैं। इस उपकरण के सात भाग होते हैं जिन्हें ट्राफ़ाई (Trophi) कहते हैं। जीवित अवस्था में ट्रोफ़ाई प्राय: सदा गति करते हैं और पृष्ठी प्राणियों के दिल (हृदय) की भाँति मालूम पड़ते हैं। साधारण व्यक्ति इसके हृदय समझ बैठते हैं। ट्रोफ़ाई जबड़ों का कार्य करते हैं। भोज्य पदार्थ, जो करते हुए जबड़ों की लहरों के साथ आकर आहारनाल में पहुँच पिस जाते हैं। ग्रसनी की दीवार से लारग्रंथियाँ संबंधित होती हैं। यह पाचक लार को आहारनाल में पहुँचाती हैं। यह रस पिसते हुए भोज्य पदार्थ से मिलकर पाचन क्रिया पूरा करता हैं। ग्रसनी ग्रासनली (ईसोफ़ेगस, Oesophagus) में खुलती है। इसकी लंबाई भिन्न-भिन्न किरीटियों में भिन्न-भिन्न होती है। ग्रासनली आमाशय में खुलती है। यह चौड़ी थैली की भाँति होती है। आमाशय पीछे की ओर पतला होता जाता है और आंत्र (इंटेस्टाइन, Intestine) में परिवर्तित हो जाता है। आंत्र के अंतिम भाग को प्राय: अवस्कर (Cloaca) कहते हैं। इसलिए कि इसमें उत्सर्गी तंत्र की नलिकाएँ और अंडवाहिनी (ओविडक्ट, Oviduct) खुलती हैं।
श्वसन के लिए किरीटी में विशेष अंग नहीं होते। शरीर के चारों ओर जल रहता हैं। इसी जल में घुले हुए आक्सिजन का शरीर की दीवार की कोशिकाओं में विसरण (डिफ्यूजन, Diffusion) हो जाता हैं।
नाइट्रोजन-युक्त मल को बाहर निकालने के लिए किरीटी में उत्सर्जन तंत्र होता है। दो मुख्य उत्सर्जन नलिकाएँ होती हैं, जो शरीर के पार्श्वीय भागों में होती हैं। आगे से एक दूसरी से जुड़ी रहती हैं। इन उत्सर्जन नलिकाओं को प्रोटोनेफ्रडियल, (Pretonephridal) नलिका कहते हैं। प्रत्येक प्रोटोनेफ्रडियल नलिका में दो से क. स्टेफ़ैनोसिरॉस (Stephanoceros) नामक किरीटी। यह एक स्कंध द्वारा पृथ्वी में चिपका रहता है और स्कंध पैर की ग्रंथियों से निकले हुए रस की बनी थैली से ढका रहता है। ख. टाइगुए, स्कंधवाला किरीटी। इसके स्कंध पर एक अंडा चिपका हुआ है। ग. लिमनियास (Limnias) इसका वलय (Corona) द्विपिंडकीय है। घ. अनेक लिमनियास एक दूसरे के साथ समूह में। ङ. पेडैलिया (Pedalia) इसका शरीर कई चलायमान पिंडकों से बना होता है। इन पिंडकों पर लंबे-लंबे काँटे होते हैं। च. डेलायड (Bdelloid) का आगे का भाग (बगल से)। इसमें तुंड (रोस्ट्रम) स्पष्ट है। ज. शरीर की दीवार की काट (सेक्शन)। झ. पैर या दुम। इसके अंदर ग्रंथियाँ हैं, जिनकी नलिकाएँ बाहर की ओर खुलती हैं; अंगुलियाँ भी स्पष्ट हैं। आठ तक फ्लेम बल्ब नामक अंग खुलते हैं। लट्टू जैसे ये अंग स्यूडोयीलोम के तरल पदार्थ से नाइट्रोजन युक्त पदार्थ सोख लेते हैं और उसे प्रोटोनेफ्रडियल नलिका द्वारा बाहर निकाल देते हैं। दोनों तरह की नलिकाओं से मिलकर एक नली बनती हैं, जो क्लोएका में खुलती है और क्लाएक बाहर खुलती है।
मस्तिष्क की प्रतिनिधि एक बड़ी द्विपिंडकीय गुच्छिका बाइलोब्ड गैंग्लिऑन (Bilobed ganglion) है, जो ग्रसनी (मैस्टैक्स, Mastax) के पृष्ठीय ओर रहती है। इससे अनेक तंत्रिकाएँ निकलती हैं, जो शरीर के विभिन्न भागों में संबंध स्थापित करती है। तंत्रिका तंत्र शरीर की गति तथा अन्य क्रियाओं और अभिक्रियाओं पर नियंत्रण रखता है।
किरीटी के शरीर में अनेक प्रकार की ज्ञानेंद्रियाँ होती हैं। इनमें आँखें प्रमुख हैं जिनका उल्लेख पहले हो चुका है। इनका कार्य प्रकाश बोध। लगभग सभी किरीटियों में पार्श्वीय श्रृंगिकाएँ होती हैं। इसी तरह पृष्ठीय तल पर मस्तिष्क के ऊपर एक, या एक जोड़ी श्रृंगिका होती है। इसे पृष्ठीय श्रृंगिका कहते हैं। मुकुट (कारोना) पर भी अनेक ज्ञानेंद्रियाँ होती हैं, विशेषकर हाइडेटाइना (Hydatina) और सिनचीटा आदि में।
नर और नारी अलग अलग होते हैं। अधिक संख्या में नारियाँ दिखलाई देती हैं। नर केवल प्रजनन काल में ही दिखलाई पड़ते हैं। नर मादा से 1। 10 छोटे होते हैं। मादा का जननपिंड एक अंडाशय है। इससे एक नली, अंडावाहिनी, निकलकर क्लाएका में खुलती है। किसी किसी डेलायड में अंडाशय का एक जोड़ा होता है। नर जननपिंड एक बड़ी थैली जैसा वृषण (टेस्टिस, Testes) होता है। इससे एक नली बाहर खुलती है। इस नली को श्रुकवाहिनी कहते हैं। शुक्रवाहिनी की नली में अंदर रोमाभ होते हैं। उसमें एक जोड़ा (या अधिक) प्रोस्टेट ग्रंथियाँ खुलती हैं। शुक्रवाहिनी का अंतिम भाग ऐसा होता है कि वह उलटकर बाहर निकल आता है और मैथुन के लिए सिर्रस (Cirrus) का कार्य करता हैं।
मैथुन के समय सिर्रस नारी के क्लोएका में डाल दिया जाता है और शुक्राणु वहाँ छोड़ दिए जाते हैं। किरीटी में इस यथाक्रम ढंग का उपयोग कम जंतु करते हैं। अधिक संख्या में किरीटी सिर्रस को शरीर की दीवार फाड़कर भीतर डाल देते हैं और स्यूडोसील में शुक्राणु छोड़ते हैं। इस क्रिया को हाइपोडर्मिक इप्रेग्नेशन कहते है।
शुक्राणु अंडे के परिपक्व होने के पहले उसमें प्रवेश कर जाते हैं। उसके बाद अंड का आवरण कड़ा हो जाता है और प्राय: काँटेदार, दानेदार या अन्य नमूनेवाला हो जाता है। संसेचन के अनंतर परिवर्धन प्रारंभ होता हैं। कुछ समय उपरांत नन्हें बच्चे निकलते हैं। स्वतंत्र रूप से तैरनेवाले किरीटियों में बच्चे रूप रंग एवं आकार में वयस्कों जैसे होते हैं। वे कुछ ही दिनों में परिपक्व हो जाते हैं। नर जन्म के समय ही परिपक्व होते हैं, इसीलिए जितने बड़े इस समय होते है जीवन भर उतने ही बड़े रहते हैं। डंठल से जुड़े रहनेवाले किरीटियों के बच्चे भी स्वतंत्र रूप से तैरनेवाले होते हैं। कुछ समय बाद अपने पाद (फुट) की सहायता से वे तल से लग जाते हैं और पाद लंबा होकर डंठल बना देता हैं।
किरीटी या रोटिफेरा वर्ग (क्लास, class) के जीवों को तीन गणों में विभाजित किया गया है। इनके नाम हैं सीयसोनिडा, डेलॉयडिया और मॉनोगोनौंटा। इनमें से अंतिम गण में सबसे अधिक किरीटी हैं। इनमें सबसे कम विकसित सीयसोनिडा समुद्र में रहनेवाले किरीटी का छोटा गण है। डेलॉयडिया अधिकतर देखने में आते हैं। इनका मुकुट परावर्ती (रिट्रैक्टाइल, Retractile) होता है और दो पिंडकों में विभाजित रहता हैं। इनमें पर नहीं होते, केवल नारियाँ मिलती हैं। इनमें प्रजनन अनिषेकजनन क्रिया द्वारा होता हैं, अर्थात् परिवर्धन, के लिये अंडे को संसेचन की आवश्यकता नहीं होती। शेष सब तैरनेवाले, अर्थात् डंठल द्वारा पृथ्वी से जुड़े रहनेवाले, किरीटी मॉनोगोनौंटा गण में हैं। इनमें नर छोटे होते हैं और उनके एक वृषण होता हैं। यह गण तीन उपकरणों में विभाजित हैं। इनके नाम हैं:
(क) प्लायमा, अर्थात् तैरनेवाले प्राणी;
(ख) फ्लोर-कूलेरियेमिया, तैरनेवाले या पृथ्वी से जुड़े किरीटी; और
(ग) कौलोथिकेशिया, अधिकतर पृथ्वी से जुड़े रहनेवाले किरीटी, जिनका अगला भाग प्याले के आकार का होता है, केंद्रीय मुँह होता है तथा प्राय: रोमाभ के स्थान पर कॉरोना में बड़े-बड़े अचलायमान काँटे होते हैं।
किरीटी (रोटिफ़ेरा, Rotifera) स्वतंत्र रूप से रहनेवाले छोटे-छोटे प्राणी हैं। इनके शरीर के अगले भाग में एक रोमाभ (Ciliary) अंग होता है, जिसके रोमाभ इस तरह गति करते हैं कि देखनेवाले को शरीर के आगे चक्र (पहिया) चलता मालूम पड़ता है। इसीलिये इन्हें पहिएदार जंतु (ह्वील ऐनीमलक्यूल, Wheel animalcule) कहते हैं। अंग्रेजी नाम रोटिफ़ेरा का यही तात्पर्य हैं। इसीलिए इस वर्ग का नाम रोटिफ़ेरा या रोटेटोरिया रखा गया है।
किरीट अधिकतर साधारण स्वच्छ (अलवण) जल में रहते हैं। कुछ खारे पानी में रहते हैं और कुछ समुद्र में भी पाए जाते हैं। कुछ पृथ्वी पर नम स्थानों पर रहते हैं और कुछ काई (Moss) के पौधे की पत्तियों के अक्ष में रहते हैं। कुछ किरीटी परोपजीवी भी होते हैं। एक जाति घोंघा (Snail) के अंडों पर परोपजीवी होती है। इस तरह किरीटी ने विभिन्न प्रकार के निवासस्थान अपना रखे हैं। इनका वितरण भी विस्तृत है। ये संसार के सभी कोनों में पाए जाते हैं।
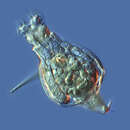
The rotifers (/ˈroʊtɪfərz/, from the Latin rota, "wheel", and -fer, "bearing"), commonly called wheel animals or wheel animalcules,[1] make up a phylum (Rotifera /roʊˈtɪfərə/) of microscopic and near-microscopic pseudocoelomate animals.
They were first described by Rev. John Harris in 1696, and other forms were described by Antonie van Leeuwenhoek in 1703.[2] Most rotifers are around 0.1–0.5 mm (0.0039–0.0197 in) long (although their size can range from 50 μm (0.0020 in) to over 2 mm (0.079 in)),[1] and are common in freshwater environments throughout the world with a few saltwater species.
Some rotifers are free swimming and truly planktonic, others move by inchworming along a substrate, and some are sessile, living inside tubes or gelatinous holdfasts that are attached to a substrate. About 25 species are colonial (e.g., Sinantherina semibullata), either sessile or planktonic. Rotifers are an important part of the freshwater zooplankton, being a major foodsource and with many species also contributing to the decomposition of soil organic matter.[3] Most species of the rotifers are cosmopolitan, but there are also some endemic species, like Cephalodella vittata to Lake Baikal.[4] Recent barcoding evidence, however, suggests that some 'cosmopolitan' species, such as Brachionus plicatilis, B. calyciflorus, Lecane bulla, among others, are actually species complexes.[5][6] In some recent treatments, rotifers are placed with acanthocephalans in a larger clade called Syndermata.
In June 2021, biologists reported the restoration of bdelloid rotifers after being frozen for 24,000 years in the Siberian permafrost.[7][8]
Rev. John Harris first described the rotifers (in particular a bdelloid rotifer) in 1696 as "an animal like a large maggot which could contract itself into a spherical figure and then stretch itself out again; the end of its tail appeared with a forceps like that of an earwig".[2] In 1702, Antonie van Leeuwenhoek gave a detailed description of Rotifer vulgaris and subsequently described Melicerta ringens and other species.[9] He was also the first to publish observations of the revivification of certain species after drying. Other forms were described by other observers, but it was not until the publication of Christian Gottfried Ehrenberg's Die Infusionsthierchen als vollkommene Organismen in 1838 that the rotifers were recognized as being multicellular animals.[9]
About 2,200 species of rotifers have been described. Their taxonomy is currently in a state of flux. One treatment places them in the phylum Rotifera, with three classes: Seisonidea, Bdelloidea and Monogononta.[10] The largest group is the Monogononta, with about 1,500 species, followed by the Bdelloidea, with about 350 species. There are only two known genera with three species of Seisonidea.[11]
The Acanthocephala, previously considered to be a separate phylum, have been demonstrated to be modified rotifers. The exact relationship to other members of the phylum has not yet been resolved.[12] One possibility is that the Acanthocephala are closer to the Bdelloidea and Monogononta than to the Seisonidea; the corresponding names and relationships are shown in the cladogram below.
Syndermata EurotatoriaThe Rotifera, strictly speaking, are confined to the Bdelloidea and the Monogononta. Rotifera, Acanthocephala and Seisonida make up a clade called Syndermata.[13]
The word rotifer is derived from a Neo-Latin word meaning "wheel-bearer",[14] due to the corona around the mouth that in concerted sequential motion resembles a wheel (though the organ does not actually rotate).
Rotifers have bilateral symmetry and a variety of different shapes. The body of a rotifer is divided into a head, trunk, and foot, and is typically somewhat cylindrical. There is a well-developed cuticle, which may be thick and rigid, giving the animal a box-like shape, or flexible, giving the animal a worm-like shape; such rotifers are respectively called loricate and illoricate. Rigid cuticles are often composed of multiple plates, and may bear spines, ridges, or other ornamentation. Their cuticle is nonchitinous and is formed from sclerotized proteins.
The two most distinctive features of rotifers (in females of all species) are the presence of corona on the head, a structure ciliated in all genera except Cupelopagis and presence of mastax. In the more primitive species, the corona forms a simple ring of cilia around the mouth from which an additional band of cilia stretches over the back of the head. In the great majority of rotifers, however, this has evolved into a more complex structure.
Modifications to the basic plan of the corona include alteration of the cilia into bristles or large tufts, and either expansion or loss of the ciliated band around the head. In genera such as Collotheca, the corona is modified to form a funnel surrounding the mouth. In many species, such as those in the genus Testudinella, the cilia around the mouth have disappeared, leaving just two small circular bands on the head. In the bdelloids, this plan is further modified, with the upper band splitting into two rotating wheels, raised up on a pedestal projecting from the upper surface of the head.[15]
The trunk forms the major part of the body, and encloses most of the internal organs. The foot projects from the rear of the trunk, and is usually much narrower, giving the appearance of a tail. The cuticle over the foot often forms rings, making it appear segmented, although the internal structure is uniform. Many rotifers can retract the foot partially or wholly into the trunk. The foot ends in from one to four toes, which, in sessile and crawling species, contain adhesive glands to attach the animal to the substratum. In many free-swimming species, the foot as a whole is reduced in size, and may even be absent.[15]


The coronal cilia create a current that sweeps food into the mouth. The mouth opens into a characteristic chewing pharynx (called the mastax), sometimes via a ciliated tube, and sometimes directly. The pharynx has a powerful muscular wall and contains tiny, calcified, jaw-like structures called trophi, which are the only fossilizable parts of a rotifer. The shape of the trophi varies between different species, depending partly on the nature of their diet. In suspension feeders, the trophi are covered in grinding ridges, while in more actively carnivorous species, they may be shaped like forceps to help bite into prey. In some ectoparasitic rotifers, the mastax is adapted to grip onto the host, although, in others, the foot performs this function instead.[15]
Behind the mastax lies an oesophagus, which opens into a stomach where most of the digestion and absorption occurs. The stomach opens into a short intestine that terminates in a cloaca on the posterior dorsal surface of the animal. Up to seven salivary glands are present in some species, emptying to the mouth in front of the oesophagus, while the stomach is associated with two gastric glands that produce digestive enzymes.[15]
A pair of protonephridia open into a bladder that drains into the cloaca. These organs expel water from the body, helping to maintain osmotic balance.[15]
Rotifers have a small brain, located just above the mastax, from which a number of nerves extend throughout the body. The number of nerves varies among species, although the nervous system usually has a simple layout. Close to the brain lies a retrocerebral organ, consisting of two glands either side of a medial sac. The sac drains into a duct that divides into two before opening through pores on the uppermost part of the head. The function of the retrocerebral organ is unclear.[15][16]
The nervous system comprises about 25% of the roughly 1,000 cells in a rotifer.[16]
Rotifers typically possess one or two pairs of short antennae and up to five eyes. The eyes are simple in structure, sometimes with just a single photoreceptor cell. In addition, the bristles of the corona are sensitive to touch, and there are also a pair of tiny sensory pits lined by cilia in the head region.[15]
The coronal cilia pull the animal, when unattached, through the water.
Like many other microscopic animals, adult rotifers frequently exhibit eutely—they have a fixed number of cells within a species, usually on the order of 1,000.
Bdelloid rotifer genomes contain two or more divergent copies of each gene, suggesting a long-term asexual evolutionary history.[17] For example, four copies of hsp82 are found. Each is different and found on a different chromosome excluding the possibility of homozygous sexual reproduction.
Rotifers eat particulate organic detritus, dead bacteria, algae, and protozoans. They eat particles up to 10 micrometres in size. Like crustaceans, rotifers contribute to nutrient recycling. For this reason, they are used in fish tanks to help clean the water, to prevent clouds of waste matter. Rotifers affect the species composition of algae in ecosystems through their choice in grazing. Rotifers may compete with cladocera and copepods for planktonic food sources.
Rotifers are dioecious and reproduce sexually or parthenogenetically. They are sexually dimorphic, with the females always being larger than the males. In some species, this is relatively mild, but in others the female may be up to ten times the size of the male. In parthenogenetic species, males may be present only at certain times of the year, or absent altogether.[15]
The female reproductive system consists of one or two ovaries, each with a vitellarium gland that supplies the eggs with yolk. Together, each ovary and vitellarium form a single syncitial structure in the anterior part of the animal, opening through an oviduct into the cloaca.[15]
Males do not usually have a functional digestive system, and are therefore short-lived, often being sexually fertile at birth. They have a single testicle and sperm duct, associated with a pair of glandular structures referred to as prostates (unrelated to the vertebrate prostate). The sperm duct opens into a gonopore at the posterior end of the animal, which is usually modified to form a penis. The gonopore is homologous to the cloaca of females, but in most species has no connection to the vestigial digestive system, which lacks an anus.[15]
The phylum Rotifera encloses three classes that reproduce by three different mechanisms: Seisonidea only reproduce sexually; Bdelloidea reproduce exclusively by asexual parthenogenesis; Monogononta reproduce alternating these two mechanisms ("cyclical parthenogenesis" or "heterogony").[18] Parthenogenesis (amictic phase) dominates the monogonont life cycle, promoting fast population growth and colonization. In this phase males are absent and amictic females produce diploid eggs by mitosis which develop parthenogenetically into females that are clones of their mothers.[18] Some amictic females can generate mictic females that will produce haploid eggs by meiosis. Mixis (meiosis) is induced by different types of stimulus depending on species. Haploid eggs develop into haploid dwarf males if they are not fertilized and into diploid "resting eggs" (or "diapausing eggs") if they are fertilized by males.
Fertilization is internal. The male either inserts his penis into the female's cloaca or uses it to penetrate her skin, injecting the sperm into the body cavity. The egg secretes a shell, and is attached either to the substratum, nearby plants, or the female's own body. A few species, such as members of the Rotaria, are ovoviviparous, retaining the eggs inside their body until they hatch.[15]
Most species hatch as miniature versions of the adult. Sessile species, however, are born as free-swimming larvae, which closely resemble the adults of related free-swimming species. Females grow rapidly, reaching their adult size within a few days, while males typically do not grow in size at all.[15]
The life span of monogonont females varies from two days to about three weeks.
'Ancient asexuals': Bdelloid rotifers are assumed to have reproduced without sex for many millions of years. Males are absent within the species, and females reproduce only by parthenogenesis.
However, a new study provided evidence for interindividual genetic exchange and recombination in Adineta vaga, a species previously thought to be anciently asexual.[19]
Recent transitions: Loss of sexual reproduction can be inherited in a simple Mendelian fashion in the monogonont rotifer Brachionus calyciflorus: This species can normally switch between sexual and asexual reproduction (cyclical parthenogenesis), but occasionally gives rise to purely asexual lineages (obligate parthenogens). These lineages are unable to reproduce sexually due to being homozygous for a recessive allele.[20]
Resting eggs enclose an embryo encysted in a three-layered shell that protects it from external stressors.[21][22] They are able to remain dormant for several decades and can resist adverse periods (e.g., pond desiccation or presence of antagonists).[23][24] When favourable conditions return and after an obligatory period of diapause which varies among species, resting eggs hatch releasing diploid amictic females that enter into the asexual phase of the life cycle.[18][25]
Bdelloid rotifer females cannot produce resting eggs, but many can survive prolonged periods of adverse conditions after desiccation. This facility is termed anhydrobiosis, and organisms with these capabilities are termed anhydrobionts. Under drought conditions, bdelloid rotifers contract into an inert form and lose almost all body water; when rehydrated they resume activity within a few hours. Bdelloids can survive the dry state for long periods, with the longest well-documented dormancy being nine years. While in other anhydrobionts, such as the brine shrimp, this desiccation tolerance is thought to be linked to the production of trehalose, a non-reducing disaccharide (sugar), bdelloids apparently cannot synthesise trehalose. In bdelloids, a major cause of the resistance to desiccation, as well as resistance to ionizing radiation, is a highly efficient mechanism for repairing the DNA double-strand breaks induced by these agents.[26] This repair mechanism likely involves mitotic recombination between homologous DNA regions.[26]
Rotifers fall prey to many animals, such as copepods, fish (e.g. herring, salmon), bryozoa, comb jellies, jellyfish, starfish, and tardigrades.[27]
The genome size of a bdelloid rotifer, Adineta vaga, was reported to be around 244 Mb.[28] The genomes of Monogononts seem to be significantly smaller than those of Bdelloids. In Monogononta the nuclear DNA content (2C) in eight different species of four different genera ranged almost fourfold, from 0.12 to 0.46 pg.[29] Haploid "1C" genome sizes in Brachionus species range at least from 0.056 to 0.416 pg.[30]

Pair of Lepadella rotifers from pond water

Locula of the rotifer Keratella cochlearis
The rotifers (/ˈroʊtɪfərz/, from the Latin rota, "wheel", and -fer, "bearing"), commonly called wheel animals or wheel animalcules, make up a phylum (Rotifera /roʊˈtɪfərə/) of microscopic and near-microscopic pseudocoelomate animals.
They were first described by Rev. John Harris in 1696, and other forms were described by Antonie van Leeuwenhoek in 1703. Most rotifers are around 0.1–0.5 mm (0.0039–0.0197 in) long (although their size can range from 50 μm (0.0020 in) to over 2 mm (0.079 in)), and are common in freshwater environments throughout the world with a few saltwater species.
Some rotifers are free swimming and truly planktonic, others move by inchworming along a substrate, and some are sessile, living inside tubes or gelatinous holdfasts that are attached to a substrate. About 25 species are colonial (e.g., Sinantherina semibullata), either sessile or planktonic. Rotifers are an important part of the freshwater zooplankton, being a major foodsource and with many species also contributing to the decomposition of soil organic matter. Most species of the rotifers are cosmopolitan, but there are also some endemic species, like Cephalodella vittata to Lake Baikal. Recent barcoding evidence, however, suggests that some 'cosmopolitan' species, such as Brachionus plicatilis, B. calyciflorus, Lecane bulla, among others, are actually species complexes. In some recent treatments, rotifers are placed with acanthocephalans in a larger clade called Syndermata.
In June 2021, biologists reported the restoration of bdelloid rotifers after being frozen for 24,000 years in the Siberian permafrost.
Rotiferoj (Rotifera, ofte nomataj radanimaloj) formas filumon de mikroskopaj kaj preskaŭ-mikroskopaj fals-kelomataj animaloj.
Ili estis unuafoje priskribita de John Harris en 1696, kaj aliaj formoj estis priskribitaj de Antoni van Leeuwenhoek en 1703.[1] Plej rotiferoj estas ĉirkaŭ 0.1–0.5 mm longaj (kvankam ties grando povas gami el 50 μm al ĉirkaŭ 2 mm),[2] kaj oftas en nesalakvaj medioj tra la tuha mondo kun kelkaj salakvaj specioj; ekzemple, tiuj de la genro Synchaeta.
Kelkaj rotiferoj estas naĝantoj kaj vere planktonaj, dum aliaj moviĝas vermece laŭlonge de subtavolo, kaj kelkaj estas nemovaj, loĝantaj ene de tuboj aŭ gelatenaj fiksuloj kiuj estas ligitaj al subtavolo. Ĉirkaŭ 25 specioj estas koloniaj (ekz., Sinantherina semibullata), ĉu senmovaj aŭ planktonaj. Rotiferoj estas grava parto de nesalakva zooplanktono, kaj grava manĝofonto kaj kun multaj specioj ankaŭ kontribuas al la malkompono de grundorganika materialo.[3] Plej specioj de rotiferoj estas tutmondaj, sed estas ankaŭ kelkaj endemiaj specioj, kiaj Cephalodella vittata el la Lago Bajkalo.[4] Ĵusa strikoda pruvaro, tamen, sugestas ke kelkaj 'tutmondaj' specioj, kiaj Brachionus plicatilis, B. calyciflorus, Lecane bulla, inter aliaj, estas fakte specikompleksoj.[5][6]
En kelkaj ĵusaj traktaĵoj, rotiferoj eatas lokigitaj kun Acanthocephala en pli granda klado nomata Syndermata.
Rotiferoj (Rotifera, ofte nomataj radanimaloj) formas filumon de mikroskopaj kaj preskaŭ-mikroskopaj fals-kelomataj animaloj.
Ili estis unuafoje priskribita de John Harris en 1696, kaj aliaj formoj estis priskribitaj de Antoni van Leeuwenhoek en 1703. Plej rotiferoj estas ĉirkaŭ 0.1–0.5 mm longaj (kvankam ties grando povas gami el 50 μm al ĉirkaŭ 2 mm), kaj oftas en nesalakvaj medioj tra la tuha mondo kun kelkaj salakvaj specioj; ekzemple, tiuj de la genro Synchaeta.
Kelkaj rotiferoj estas naĝantoj kaj vere planktonaj, dum aliaj moviĝas vermece laŭlonge de subtavolo, kaj kelkaj estas nemovaj, loĝantaj ene de tuboj aŭ gelatenaj fiksuloj kiuj estas ligitaj al subtavolo. Ĉirkaŭ 25 specioj estas koloniaj (ekz., Sinantherina semibullata), ĉu senmovaj aŭ planktonaj. Rotiferoj estas grava parto de nesalakva zooplanktono, kaj grava manĝofonto kaj kun multaj specioj ankaŭ kontribuas al la malkompono de grundorganika materialo. Plej specioj de rotiferoj estas tutmondaj, sed estas ankaŭ kelkaj endemiaj specioj, kiaj Cephalodella vittata el la Lago Bajkalo. Ĵusa strikoda pruvaro, tamen, sugestas ke kelkaj 'tutmondaj' specioj, kiaj Brachionus plicatilis, B. calyciflorus, Lecane bulla, inter aliaj, estas fakte specikompleksoj.
En kelkaj ĵusaj traktaĵoj, rotiferoj eatas lokigitaj kun Acanthocephala en pli granda klado nomata Syndermata.
Los rotíferos (Rotifera, del latín rota, "rueda" y fera, "los que llevan") constituyen un filo de animales pseudocelomados microscópicos (entre 0,1 y 0,5 mm) con unas 2200 especies[2] que habitan en aguas dulces, tierra húmeda, musgos, líquenes, hongos, e incluso agua salada. Fueron descubiertos por primera vez por John Harris en 1696 (Hudson and Gosse, 1886), de quien se conservan dibujos fechados en 1703, a pesar de que el hallazgo suele atribuirse equivocadamente a Leeuwenhoek.
También incluye al exfilo Acanthocephala el cual se clasifica actualmente en Rotifera debido a que algunos rotíferos se encuentran más emparentados con Acanthocephala que con otros rotíferos, haciendo que Rotifera sea un grupo monofilético.[1]
La boca está situada en la zona ventral de la región cefálica, y puede estar rodeada por bandas ciliadas del aparato rotador que crean pequeñas corrientes que atraen las partículas de alimento del entorno. A continuación se encuentra la faringe, muy musculada, a la que se accede directamente a través de la boca o mediante un delgado tubo ciliado (como ocurre en algunas especies suspensívoras) situado entre ambas estructuras. La faringe presenta siete piezas duras y articuladas conocidas como trophi (que actúan a modo de dientes), y forman en conjunto el mástax (que actúa como el aparato masticador). Los trophi se distribuyen de la siguiente manera:
Los trophi (y mástax) son muy variables, y dependen principalmente del régimen alimentario de cada especie, por lo que es un buen identificador taxonómico. Situado en la parte posterior de la faringe encontramos glándulas salivares que vierten al mástax diversas enzimas digestivas. Por detrás de la faringe se encuentra un esófago que desemboca a un amplio estómago, situándose entre ambos un par de glándulas gástricas. Después del estómago aparece un intestino que desemboca en una cloaca a la que también vierten el sistema excretor y reproductor.
Los rotíferos tienen sexos separados. Se reproducen sexual y asexualmente. Los machos son más pequeños que las hembras, hasta el punto que, en algunas ocasiones, la hembra es diez veces más grande que el macho. También hay algunas especies en las que no existen individuos machos, ya que las hembras se reproducen por partenogénesis.
La cópula se produce por impregnación hipodérmica, es decir, el macho penetra a la hembra en cualquier parte del cuerpo. La inyección de los espermatozoides no es fácil ya que los rotíferos poseen una cutícula y una loriga que los protege. Por esta razón los machos suelen penetrar a la hembra por la región de la corona, donde la cutícula es más blanda.
Los huevos son fijados al sustrato o bien se adhieren a la hembra, pero también hay especies vivíparas. Algunos rotíferos producen dos tipos de huevos: los huevos amícticos son los huevos de verano, tienen la cáscara delgada y son diploides, por lo tanto no pueden ser fecundados. De estos huevos sale siempre una hembra por partenogénesis. El otro tipo de huevos son los huevos mícticos, huevos que ponen en invierno; tienen la cáscara dura y son haploides. Si no son fecundados producen un macho por partenogénesis, pero si son fecundados producen una hembra.
Generalmente los rotíferos no tienen larvas. Del huevo salen individuos sexualmente maduros o casi maduros. Únicamente las especies sésiles tienen un estadio larvario, que es bastante breve.
Los rotíferos se alimentan de detritos órganicos particulados, bacterias, algas unicelulares y protozoos. Comen partículas de hasta 10 micrómetros de tamaño. Al igual que los crustáceos, los rotíferos contribuyen al reciclaje de nutrientes.
Existen rotíferos con una cutícula relativamente gruesa y rígida que dan al animal una apariencia capsular; otros son flexibles y de aspecto y movimiento vermiforme. Aunque la gran mayoría de rotíferos son nadadores, algunos son sésiles y viven en el interior de tubos o cápsulas gelatinosas. Alrededor de 25 especies son coloniales (como Sinantherina semibullata). En la mayoría de especies los machos son escasos, de forma que la reproducción por partenogénesis es bastante común. Además existe un marcado dimorfismo sexual ya que los machos existentes suelen tener el aparato digestivo atrofiado, y su existencia se reduce a la producción de esperma. La vida de una hembra suele ser de una o dos semanas.
Algunas especies son xerobiontes, es decir, sus huevos fertilizados se enquistan y crean formas de resistencia capaces de sobrevivir a los períodos de sequía y desecación en los que el hábitat acuático desaparece por motivos estacionales. Al ser rehidratados emergen rápidamente (en menos de dos horas) como organismos con capacidad natatoria. Algunas especies producen un derivado del azúcar trehalosa con el que crean una sustancia gelatinosa que ayuda a resistir a la falta de agua ambiental.
El análisis ultrestructural encontró que Rotifera sería un grupo parafilético con respecto a Acanthocephala.[3] El análisis filogenómico ha encontrado los siguientes resultados:[4]
Rotifera Hemirotifera PararotatoriaLa posibilidad alternativa es clasificar a Acanthocephala como un subtaxón dentro de Rotifera (Rotifera sensu lato). Por tanto ambos grupos podrían considerarse rotíferos[1] y el término "Syndermata" se convertiría en un sinónimo cladístico.[4]
Los rotíferos (Rotifera, del latín rota, "rueda" y fera, "los que llevan") constituyen un filo de animales pseudocelomados microscópicos (entre 0,1 y 0,5 mm) con unas 2200 especies que habitan en aguas dulces, tierra húmeda, musgos, líquenes, hongos, e incluso agua salada. Fueron descubiertos por primera vez por John Harris en 1696 (Hudson and Gosse, 1886), de quien se conservan dibujos fechados en 1703, a pesar de que el hallazgo suele atribuirse equivocadamente a Leeuwenhoek.
También incluye al exfilo Acanthocephala el cual se clasifica actualmente en Rotifera debido a que algunos rotíferos se encuentran más emparentados con Acanthocephala que con otros rotíferos, haciendo que Rotifera sea un grupo monofilético.
Keriloomad ehk rotatoorid (Rotifera ehk Rotatoria) on kosmopoliitne alamate loomade hõimkond, kuhu kuuluvad mikroskoopilised pseudotsöloomiga loomad.
Keriloomade morfoloogia ja bioloogia on väga mitmekesine. Mageveekogudes, eriti eutroofsetes, moodustavad nad tihti suure osa zooplanktonist.
Enamik keriloomi on umbes 0,1–0,5 mm pikad ning elavad valdavalt mageveelistes või vähese soolsusega veekogudes.
Keriloomad võivad olla planktilised, aeglaselt substraadil liikuvad või sessiilsed ehk elavad substraadile kinnitunult. Keriloomad on virustajad ehk aktiivsed biofiltraatorid.
Keriloomade nimetus tuleb sellest, et nende peaotsas asub ripsmetest koosnev keriaparaat ehk mälumik, mille abil nad toiduosakesi suhu toimetavad ja mis neil vajadusel liikuda aitab.
Esimesena kirjeldas neid 1696. aastal John Harris.
Keriloomad ehk rotatoorid (Rotifera ehk Rotatoria) on kosmopoliitne alamate loomade hõimkond, kuhu kuuluvad mikroskoopilised pseudotsöloomiga loomad.
Keriloomade morfoloogia ja bioloogia on väga mitmekesine. Mageveekogudes, eriti eutroofsetes, moodustavad nad tihti suure osa zooplanktonist.
Enamik keriloomi on umbes 0,1–0,5 mm pikad ning elavad valdavalt mageveelistes või vähese soolsusega veekogudes.
Keriloomad võivad olla planktilised, aeglaselt substraadil liikuvad või sessiilsed ehk elavad substraadile kinnitunult. Keriloomad on virustajad ehk aktiivsed biofiltraatorid.
Keriloomade nimetus tuleb sellest, et nende peaotsas asub ripsmetest koosnev keriaparaat ehk mälumik, mille abil nad toiduosakesi suhu toimetavad ja mis neil vajadusel liikuda aitab.
Esimesena kirjeldas neid 1696. aastal John Harris.
Rotifera animalia sasizelomatu mikroskopikoz osatutako filum bat da. Lehenengo aldiz John Harrisek deskribatu zituen 1696an eta Anton van Leuuwenhoekek ere beste forma batzuk aipatu zituen 1703an. Gehienak 0,1 eta 0,5 milimetro arteko tamaina dute, nahi zeta 50 μm eta 2 milimetro arteko tamaina izan dezaketen. Normalki ur gezatan bizi dira mundu osoan zehar, itsastarrak diren espezie batzuekin batera. Rotifera batzuk igerilariak dira eta, beraz, guztiz planktoniko. Beste batzuk substratuan zehar mugitzen dira eta beste batzuk sesilak dira, tutu batzuen barruan bizi direlarik. 25 espezie kolonial dira (adibidez Sinantherina semibullata). Ur gezako zooplanktonaren parte garrantzitsua dira.
Rataseläimet (Rotifera) on eläinkunnan pääjakso. Rataseläimet ovat tavallisemmin alle 2 mm:n pituisia ja läpikuultavia. Niitä elää kaikkialla maapallolla. Lajeja tunnetaan Suomesta yli 400.[1]
Rataseläinten ihon pinnalla on kiinteä panssari ja ruumiin takapäässä jalka, jolla eläin kiinnittyy alustaan. Suun ympärillä on kaksiosainen levy, jonka ripsikiehkurat ohjaavat ravintohiukkasia suuhun ja toimivat myös liikuntaelimenä. Rataseläimet ovat yksineuvoisia ja lisääntyvät munimalla. Niille on tyypillistä sukupolvenvuorottelu partenogeneettisesti lisääntyvän ja hedelmöittyvän sukupolven välillä.
Rataseläimet elävät sammalikoissa, maassa ja suolattomassa vedessä. Osa rataseläimistä on myös loisia. Pikkumatojen ja äyriäisten ravintona niillä on merkitystä sisävesien planktonissa. Kuivuneina rataseläimet kestävät äärimmäisiä olosuhteita ja voivat tällöin pysyä elossa 3–4 vuotta ja levitä tuulen mukana paikasta toiseen. Rataseläimiä on myös tavattu keinotekoisissa ympäristöissä, kuten akvaarioissa ja piha-altaissa.
Rataseläimet (Rotifera) on eläinkunnan pääjakso. Rataseläimet ovat tavallisemmin alle 2 mm:n pituisia ja läpikuultavia. Niitä elää kaikkialla maapallolla. Lajeja tunnetaan Suomesta yli 400.
Rataseläinten ihon pinnalla on kiinteä panssari ja ruumiin takapäässä jalka, jolla eläin kiinnittyy alustaan. Suun ympärillä on kaksiosainen levy, jonka ripsikiehkurat ohjaavat ravintohiukkasia suuhun ja toimivat myös liikuntaelimenä. Rataseläimet ovat yksineuvoisia ja lisääntyvät munimalla. Niille on tyypillistä sukupolvenvuorottelu partenogeneettisesti lisääntyvän ja hedelmöittyvän sukupolven välillä.
Les Rotifères (Rotifera) constituent un embranchement du règne animal. Ce sont de petits organismes bilatériens (i.e. triploblastiques) protostomiens syndermés mesurant entre 50 µm et 3 mm qui ont souvent une forme de trompette, cylindrique ou sphérique. Ils possèdent deux couronnes de cils autour de leur bouche ainsi qu'un système organique spécialisé avec notamment un tube digestif[1].
On sait désormais que l'embranchement des Acanthocephala est en fait un groupe de Rotifères devenus parasites[2]. La notion actuelle de Rotifère (s.l.) est donc élargie aux Acanthocéphales et devient ainsi identique au clade des Syndermata[3]. Avec les Micrognathozoa et les Gnathostomulida ils appartiennent au clade des Gnathifera (au sein duquel ils sont groupe frère avec les Micrognathozoa), partageant avec eux la présence de mâchoires organisées de manière similaire[4],[5],[6].
Ces Pseudocœlomates ont un corps en trois parties bien distinctes: la tête (appareil rotateur) le tronc et le pied terminé par deux orteils. La tête et la partie postérieure ne sont pas couvertes par la cuticule[7],[8].
Le terme Rotifère (du latin rota, « roue ») leur vient des deux couronnes de cils entourant leur bouche, dans la région antérieure, qui tourbillonnent en sens contraire pour faire entrer l'eau et les particules de nourriture qui l'accompagnent. Ces cils peuvent aussi servir à la locomotion chez certaines espèces. Au fond du pharynx musculeux se situe un appareil masticateur caractéristique, le mastax, constitué de sept pièces dures et mobiles servant à broyer la nourriture.
Postérieur, leur pied possède souvent une glande adhésive qui permet à certaines espèces de se fixer au substrat. L'anus est en position postérieure et le système excréteur est formé de deux protonéphridies à l'intérieur du pseudocœlome. Le pseudocœlome renferme les organes internes. Cette cavité corporelle est en partie tapissée de mésoderme. Le liquide du pseudocœlome sert de squelette hydraulique, ce sont les mouvements de l'organisme qui assurent la répartition du liquide dans tout le corps afin de permettre la diffusion des nutriments et des déchets.
Le système nerveux est formé d'un ganglion cérébral dorsal antérieur et d'un nombre variable de nerfs. Le système sensoriel est composé d'organes photorécepteurs rudimentaires et de cils. Il n'y a pas d'appareil respiratoire ou circulatoire et le dioxygène diffuse à travers les tissus. La paroi du corps comporte une cuticule souple, la lorica[9]. L'animal est souvent transparent mais sa couleur peut être verte, orange, rouge ou brune selon la nourriture ingérée.
La plupart du temps, les rotifères ont une multiplication asexuée. Mais ils peuvent aussi se multiplier de manière sexuée, notamment quand leurs conditions de vie se dégradent.
Certaines espèces produisent deux sortes d'œufs, la première sorte donne des femelles et la seconde donne des mâles simplifiés incapables de se nourrir. Ces mâles produisent cependant des spermatozoïdes qui iront féconder les ovules femelles.
Les zygotes ont la particularité de pouvoir survivre même si leur milieu de vie s'assèche. Lorsque les conditions redeviennent plus favorables, ils sortent de leur léthargie et deviennent de nouvelles femelles qui se reproduisent par parthénogenèse.
Il semble qu'au sein des rotifères, toute la classe des Bdelloïdés se reproduit par parthénogenèse depuis plus de 35 millions d'années, sans qu'aucune trace de l'existence de mâle (ni même de méiose) n'ait à ce jour été découverte. Leur triple capacité de parthénogenèse, de capacité à protéger et réparer leur ADN et de résistance à la dessiccation pourrait constituer un avantage compétitif décisif dans le milieu soumis à de régulières dessiccations[10].
Dans les années 2000, plusieurs anomalies ou curiosités génétiques avaient été repérées chez ces Bdelloïdés[11], mais on ne comprenait pas encore comment ces espèces avaient pu se passer de sexualité depuis aussi longtemps, avant qu'en juillet 2013 une étude publiée dans la revue Nature apporte de premières explications[12]. Les chercheurs ont réussi à séquencer le génome d’Adineta vaga et ont montré qu'il n'existe pas dans cette espèce de similarités structurales complètes entre les deux chromosomes de chaque paire. Un chromosome donné peut même porter les deux versions d'un même gène[13].
L'espèce de rotifère A. vaga est connue pour être particulièrement résistante ; elle est capable d'anhydrobiose, avec une tolérance exceptionnelle à la dessiccation[14], mais aussi de réparer son ADN y compris après une forte irradiation (capacité supérieure à celles de Deinococcus radiodurans[15]). Récemment, une étude a confirmé que lors de périodes de sécheresse la dessiccation entraînait l'accumulation de cassures double brin d'ADN dans le génome d'A. vaga. Malgré l'accumulation d'un nombre important de ces cassures, ces individus sont capables de se réhydrater, survivre et réparer complètement leurs génomes[16].
Elle intriguait les scientifiques, car, selon les observations biologiques et paléontologiques disponibles, elle ne se reproduit qu’asexuellement, par parthénogenèse thélytoque, et ce depuis des millions d'années ; ce qui défie les lois de la sélection naturelle. On n'a jamais observé que des femelles de cette espèce pourtant assez courante. Sa survie en l'absence de mâle est une caractéristique insolite dans le monde animal ; la plupart des animaux (et des végétaux) se reproduisent en effet par voie sexuée au moins périodiquement, ce qui permet la diversification génétique, évite l'accumulation de mutations nuisibles dans leur génome, et assure à ces espèces une pérennisation réputée bien meilleure que celle des animaux asexuées.
Pour tenter de comprendre ce phénomène, un consortium international (codirigé par le CEA-Genoscope et l’Université de Namur, avec le CNRS (France) et l'INRA (France)[17]) a analysé son génome.
Les résultats publiés mi-2013 confirment l'asexualité de cette espèce (son génome présente une structure incompatible avec la reproduction sexuée, il dispose de chromosomes homologues, mais les gènes y sont disposés dans des ordres différents, et parfois même sur un seul chromosome, ce qui empêcherait en cas de reproduction sexuée la production normale de gamètes).
Cette étude a aussi expliqué la survie de l'espèce : le génome d’A. vaga dispose de mécanismes contournant les effets négatifs d'une longue période de reproduction asexuée[18] : il opère de fréquentes « conversions géniques » à l'intérieur de son génome, via une sorte de « copier-coller » de gènes, un mécanisme qui pourrait selon les auteurs de l'étude atténuer voire supprimer l'accumulation de mutations délétères. Grâce à cette étude, la science dispose maintenant d'un nouvel « indice génétique » pour détecter d'éventuelles autres espèces animales capables d'aussi durablement se reproduire sans mâle. Ces conversions géniques pourraient avoir lieu lorsque A. vaga répare son génome après dessiccation. Par ce même mécanisme il lui serait possible de capturer des gènes en provenance d'autres organismes [Comment ?] comme l'ont prouvé les données récentes du séquençage d'A. vaga[18],[16].
Un rotifère bdelloïde est revenu à la vie après avoir été congelé pendant 24 000 ans en Sibérie. Il est ensuite parvenu à se cloner, ont annoncé lundi 7 juin 2021 des scientifiques russes. L'animal était en cryptobiose : un état dans lequel le métabolisme est presque complètement à l'arrêt. Son âge est estimé entre 23 960 et 24 485 ans. Stas Malavin qui a séquencé son génome indique que celui-ci est proche d'Adineta vaga, il s'agirait d'une nouvelle espèce. Une fois décongelé, l'animal a pu se reproduire seul par parthénogenèse[19].
Ils vivent principalement en eau douce mais quelques espèces occupent les eaux salées ainsi que les milieux humides. Ils se nourrissent essentiellement de microorganismes en suspension dans l'eau[20]. Certains Rotifères sont des parasites de crustacés, de mollusques et d'annélides. Ils composent une grande partie du zooplancton d'eau douce et constituent une source de nourriture importante dans les écosystèmes d'eau douce. En milieu terrestre, ils interviennent dans la décomposition des matières organiques dans le sol. Ils sont utilisés dans des études écotoxicologiques[21].
Certaines espèces de rotifères comme Brachionus plicatilis sont utilisées en aquaculture et aquariophilie pour l'élevage des larves de poisson[22]. Elles constituent souvent la première nourriture distribuée aux larves avant les artemias.
La culture des rotifères se fait dans des cuves munies d'une bonne aération (diffuseur d'air ou d'oxygène) et la nourriture est constituée d'algues (Nannochloris, Isochrisis...), de levure de boulanger ou d'autres produits spécifiques à la culture ou l'enrichissement des rotifères.
L'embranchement des Rotifères contient 4 classes, environ 33 familles, 112 genres pour 1816 espèces. Le plus ancien fossile connu, appartenant à Keratella sp, a été découvert à la base de l'Éocène moyen d'Australie et remonte à −45 Ma [23].
Les Rotifères se décomposent en :
et désormais :
Selon World Register of Marine Species (7 mars 2016)[25] :
La position exacte des Rotifères parmi les Lophotrochozoaire n'est pas résolue, mais on les place fréquemment dans le groupe des Gnathifères et eux-mêmes parmi les Platyzoaires.
Philodina sp., un Philodinidae
Polyarthra sp., un Synchaetidae
Les Rotifères (Rotifera) constituent un embranchement du règne animal. Ce sont de petits organismes bilatériens (i.e. triploblastiques) protostomiens syndermés mesurant entre 50 µm et 3 mm qui ont souvent une forme de trompette, cylindrique ou sphérique. Ils possèdent deux couronnes de cils autour de leur bouche ainsi qu'un système organique spécialisé avec notamment un tube digestif.
On sait désormais que l'embranchement des Acanthocephala est en fait un groupe de Rotifères devenus parasites. La notion actuelle de Rotifère (s.l.) est donc élargie aux Acanthocéphales et devient ainsi identique au clade des Syndermata. Avec les Micrognathozoa et les Gnathostomulida ils appartiennent au clade des Gnathifera (au sein duquel ils sont groupe frère avec les Micrognathozoa), partageant avec eux la présence de mâchoires organisées de manière similaire,,.
Ainmhí micreascópach uisciúil le colainn nach bhfuil roinnte i dteascáin, clúdaithe de ghnáth le ciseal ceiritineach a fhéadfaidh a bheith ramhraithe i bplátaí. Tá balla matánach in easnamh ar an gcolainn. Snámhann sé le fáinne struchtúr cosúil le ribí gruaige a bhuaileann mar a bheadh roth ag casadh iontu. Ar a laghad 1,800 speiceas ann.

Philodina alimentándose
Clasificación científica Reino: Animalia Subreino: Eumetazoa Superfilo: Platyzoa Filo: Rotifera ClasesOs Rotíferos ou Rotifera constitúen un filo de animais pseudocelomados microscópicos (entre 0,1 e 0,5 mm) acuáticos. O nome de rotífero provén do latín rota (roda) e ferre (levar), é dicir, “portadores de rodas”, en referencia ao seu aparato succionador, a coroa, que lembra unha coroa real ou unha serra dentada circular.
Foron descubertos por John Harris en 1696 (Hudson and Gosse, 1886), de quen se conservan debuxos datados en 1703, a pesar de que o achado adoita atribuírse de xeito equivocado a Leeuwenhoek.
Os rotíferos presentan simetría bilateral e corpo posúe unha cabeza, un tronco e unha cola ou pé. Os machos son raros e nalgunhas especies non se coñecen. Pódense diferenciar unhas especies de outras polo tipo de coroa que aparece na cabeza, pola forma do corpo e pola cuberta cuticular externa.
O centro da cabeza non posúe cilios e leva o nome de rexión apical e está rodeada pola coroa de cilios. Na cabeza tamén ten unhas prolongacións dos órganos retrocerebrais.
O tronco é cilíndrico ou aplanado é presenta unha cuberta ou cutícula que varía dunhas especies a outras. Nalgúns casos é como unha caixa e noutros aparece segmentada en aneis o que lle permite cambiar a forma e lonxitude do corpo. No tronco poden aparecer unha ou dúas antenas dorsais e outras antenas laterais.
O pé está cuberto de aneis cuticulares que remata ou nuns dedos que se moven nas especies nadadoras ou nun disco adhesivo nas especies fixas ou sésiles.
Adoitan ter un tubo dixestivo moi completo, con boca, farinxe moi diferenciada con moitas pezas móbiles que fan o papel de mandíbulas. A coroa está composta de cilios que, ao moverse, crean pequenas correntes que atraen as partículas do contorno cara ela. Da coroa pasan a unha farinxe e desta ao aparato mastigador denominado mástax, onde se trituran as partículas succionadas, para despois ser dixeridas no estómago e finalmente excretados os residuos pola cloaca. A coroa tamén a utilizan para desprazarse pola auga, a modo de turbohélice. No extremo oposto á coroa se distingue un pé (xeralmente un par de pés) co cal poden aferrarse ao substrato.
Existen rotíferos cunha cutícula relativamente grosa e ríxida que dan ao animal un aspecto capsular; outros son flexibles e de aspecto e movemento vermiforme. Aínda que a gran maioría de rotíferos son nadadores, algúns son sésiles e viven no interior de tubos ou cápsulas xelatinosas. Arredor de 25 especies son coloniais (como Sinantherina semibullata).
Na maioría de especies os machos son escasos, de forma que a reprodución por partenoxénese é bastante común. Ademais existe un marcado dimorfismo sexual xa que os machos existentes acostuman ter o aparato dixestivo atrofiado, e a súa existencia limítase á produción de esperma. A vida dunha femia oscila entre as 1-2 semanas. Algunhas especies son xerobiontes, é dicir, os seus ovos fertilizados se enquistan e crean formas de resistencia capaces de sobrevivir aos períodos de seca nos que o hábitat acuático desaparece por motivos estacionais. Ó ser rehidratados emerxen rapidamente (en menos de dúas horas) como organismos con capacidade natatoria. Algunhas especies producen un derivado do azucre trehalosa co que crean unha substancia xelatinosa que axuda a resistir á falta de auga ambiental.
En toda a superficie do corpo hai moitas terminacións nerviosas. As cerdas teñen células sensoriais no seu pé. Na rexión apical teñen fosetas ciliadas con función quimiorreceptora, e tamén aparecen pelos sensoriais nalgúns casos. Os ollos poden aparecer en varios lugares como no cerebro, na coroa ou laterais. Moitos rotíferos presentan fototactismo positivo, é dicir nadan cara a luz.
Actualmente coñécense unhas 1.500-2.000 especies de rotíferos repartidos polas augas doces de todo o mundo, e incluso hai rotíferos de auga salgada ou de hábitats semiterrestres, entre os musgos e liques.
Os Rotíferos ou Rotifera constitúen un filo de animais pseudocelomados microscópicos (entre 0,1 e 0,5 mm) acuáticos. O nome de rotífero provén do latín rota (roda) e ferre (levar), é dicir, “portadores de rodas”, en referencia ao seu aparato succionador, a coroa, que lembra unha coroa real ou unha serra dentada circular.
Foron descubertos por John Harris en 1696 (Hudson and Gosse, 1886), de quen se conservan debuxos datados en 1703, a pesar de que o achado adoita atribuírse de xeito equivocado a Leeuwenhoek.
Kolnjaci (Rotatoria, Rotifera) su koljeno vrlo malenih životinja koje pripadaju natkoljenu oblenjaka.[1] Najveće životinje dosežu dužinu od 3 mm, s genetski utvrđenim brojem stanica (eutelija). U ovo koljeno svrstane životinje spadaju u skupinu najmanjih višestaničnih životinja (od 0,1 do najviše nepuna 3 milimetra). pokretljivi prednji i stražnji dijelovi izlaze iz hitinskog oklopa kojim je zaštićeno valjkasto tijelo, a na glavi imaju vijenac treplji (rotatorni aparat). Treplje neprekidno trepere i tako pokreću tijelo, a stvaranjem struje vode, prinosi hranu ustima. U svijetu živi oko 2.000 dijelom vrlo različitih vrsta.
Kolnjaci su vrlo otporne životinje i mogu preživjeti u vrlo različitim sredinama. Na kopnu, žive na stablima, u vlažnoj mahovini kao i u pijesku ili između mrvica tla, no jednako tako nastanjuju i mora i slatkovodne sredine, gdje su važan izvor hrane većim životinjama. Pri tome, ne smetaju im hladnoće Antarktike odnosno Arktika, kao ni visoke temperature termalnih izvora.
Različiti rodovi kolnjaka žive ili trajno pričvršćeni na biljkama ili slobodno plutaju vodom ili u detritusu.
Izgled kolnjaka je vrlo različit, no grubo se može podijeliti na tri dijela, s tim da vrste koje žive na biljkama oblikom potsjećaju na crve a vrste u vodi izgledaju vrećasto.
Obzirom na građu tijela, kolnjaci se mogu kretati na više načina: kliziti, plivati, skupljanjem i istezanjem puzati ili se kretati uz pomoć trepetljika na roratornom organu.
U vrijeme suše, kolnjaci koriste sposobnost anabioze. Iz tijela izbace tjelesnu tekućinu i skvrče se u kuglasti oblik. U takvom obliku, nazvanom i "suhom ukočenosti", mogu preživjeti do četiri godine.
Na trupu ili stopalu mogu ostati pričvršćeni paketi ili pojedinačna jaja koja imaju istu sposobnost reagiranja na loše uvjete u okolišu kao i odrasle životinje.
Većina vrsta hrani se algama ili detritusom. Neke vrste trepetljikama i strujom vode hvataju alge ili bakterije. No, postoje i grabežljivi kolnjaci koji iz vode koja struji love vrlo male jednostanične životinje ili dijelove. Vrsta Asplanchna birghtwelli koja je velika samo oko 1 mm je također grabežljivac. U vodu izlučuje peptid. Druge vrste imaju drugačije načine lova, a neke vrste parazitiraju na drugim životinjama.
Pojedini rodovi kolnjaka imaju različite načine razmnožavanja. U povoljnim uvjetima (uglavnom u ljetnim mjesecima) razmnožavaju se nespolno (partenogeneza), a kad su uvjeti nepovoljni (uglavnom u jesen), razmnožavanje je spolno.
Očekivani životni vijek pojedinih vrsta kolnjaka je različit, no u prosjeku je to oko jednog tjedna.
Iako se kolnjaci i danas smatraju koljenom, prema filogenetskim, morfološkim i molekularnobiološkim usporedbama danas ih se više ne smatra prirodnom skupinom (monofilija) jer se još jedno koljeno, Acanthocephala, smatra sestrinskom s redom Bdelloida iz koljena kolnjaka[2][3].
Danas se kao aktualna hipoteza prihvaćaju sljedeći srodnički odnosi unutar kolnjaka:
-- Kolnjaci (Rotatoria) |-- Seisonida '-- N.N. |-- Bdelloida i Acanthocephala '-- Monogononta
koljeno Rotifera
Kolnjaci (Rotatoria, Rotifera) su koljeno vrlo malenih životinja koje pripadaju natkoljenu oblenjaka. Najveće životinje dosežu dužinu od 3 mm, s genetski utvrđenim brojem stanica (eutelija). U ovo koljeno svrstane životinje spadaju u skupinu najmanjih višestaničnih životinja (od 0,1 do najviše nepuna 3 milimetra). pokretljivi prednji i stražnji dijelovi izlaze iz hitinskog oklopa kojim je zaštićeno valjkasto tijelo, a na glavi imaju vijenac treplji (rotatorni aparat). Treplje neprekidno trepere i tako pokreću tijelo, a stvaranjem struje vode, prinosi hranu ustima. U svijetu živi oko 2.000 dijelom vrlo različitih vrsta.
Rotifera atau rotifer (biasa disebut hewan roda) membuat sebuah filum dari hewan pseudoselomata mikroskopis dan dekat mikroskopis. Mereka pertama kali dijelaskan oleh Rev. John Harris pada 1696, dan bentuk lain yang digambarkan oleh Anton van Leeuwenhoek pada 1703.[1] Sebagian rotifera panjangnya sekitar 0,1-0,5 mm (meskipun ukuran mereka dapat berkisar dari 50 μm sampai 2 mm lebih),[2] dan yang umum di lingkungan air tawar di seluruh dunia dengan beberapa spesies air asin; misalnya, genus Synchaeta. Beberapa rotifera berenang bebas dan benar-benar planktonik, yang lain bergerak dengan inchworming sepanjang substrat, dan beberapa sessile, hidup di dalam tabung atau holdfasts agar-agar yang melekat pada substrat. Sekitar 25 spesies kolonial (misalnya, Sinantherina semibullata), baik sesil atau plankton. Rotifera adalah bagian penting dari zooplankton air tawar, menjadi sumber makanan besar dan dengan banyak spesies juga berkontribusi terhadap dekomposisi bahan organik tanah.[3] Sebagian besar spesies rotifera adalah kosmopolitan, tetapi ada juga beberapa spesies endemik, seperti Cphalodella vittata Danau Baikal.[4] Bukti barcode terbaru, bagaimanapun, menunjukkan. bahwa beberapa spesies kosmopolitan, seperti Brachionus plicatilis, B. calyciflorus, Lecane bula, antara lain, sebenarnya kompleks spesies.[5]
Dalam beberapa perlakuan terkini,, rotifer ditempatkan bersama acanthocephala dalam klad yang disebut Syndermata.
Rotifera atau rotifer (biasa disebut hewan roda) membuat sebuah filum dari hewan pseudoselomata mikroskopis dan dekat mikroskopis. Mereka pertama kali dijelaskan oleh Rev. John Harris pada 1696, dan bentuk lain yang digambarkan oleh Anton van Leeuwenhoek pada 1703. Sebagian rotifera panjangnya sekitar 0,1-0,5 mm (meskipun ukuran mereka dapat berkisar dari 50 μm sampai 2 mm lebih), dan yang umum di lingkungan air tawar di seluruh dunia dengan beberapa spesies air asin; misalnya, genus Synchaeta. Beberapa rotifera berenang bebas dan benar-benar planktonik, yang lain bergerak dengan inchworming sepanjang substrat, dan beberapa sessile, hidup di dalam tabung atau holdfasts agar-agar yang melekat pada substrat. Sekitar 25 spesies kolonial (misalnya, Sinantherina semibullata), baik sesil atau plankton. Rotifera adalah bagian penting dari zooplankton air tawar, menjadi sumber makanan besar dan dengan banyak spesies juga berkontribusi terhadap dekomposisi bahan organik tanah. Sebagian besar spesies rotifera adalah kosmopolitan, tetapi ada juga beberapa spesies endemik, seperti Cphalodella vittata Danau Baikal. Bukti barcode terbaru, bagaimanapun, menunjukkan. bahwa beberapa spesies kosmopolitan, seperti Brachionus plicatilis, B. calyciflorus, Lecane bula, antara lain, sebenarnya kompleks spesies.
Dalam beberapa perlakuan terkini,, rotifer ditempatkan bersama acanthocephala dalam klad yang disebut Syndermata.
I Rotiferi (Rotifera Cuvier, 1798) (dal latino: rota, "ruota" e ferō, "porto") costituiscono un phylum di animali pseudocelomati microscopici. La loro lunghezza tipica è tra 0,1 e 0,5 mm. Sono comuni nelle acque dolci di tutto il mondo, poche sono invece le specie marine. Si spostano nell’acqua contraendosi e allungandosi, facendo leva su due peduncoli posteriori. [1]
In alcune specie i maschi sono di ridotte dimensioni e in alcune sono scomparsi: in questi casi la riproduzione avviene unicamente per partenogenesi.
In alcune specie la riproduzione è principalmente partenogenetica ma ogni tanto vengono prodotti maschi di dimensioni ridotte, spesso neppure in grado di nutrirsi, e unicamente adibiti alla fecondazione. In queste specie gli zigoti ottenuti da fecondazione costituiscono forme di resistenza nei periodi di essiccamento dei corpi idrici che abitano: essi riprendono lo sviluppo al ripristino delle condizioni adatte alla sopravvivenza.
Nei rotiferi, come in alcuni crostacei (cladoceri, ostracodi), si realizza un'alternanza irregolare, non ciclica, tra generazioni anfigoniche e generazioni partenogenetiche. In particolare i rotiferi presentano un ciclo biologico molto complesso in risposta al loro adattamento ad ambienti che variano nel tempo (acque astatiche, ecc.)
Nelle forme dicogononti dulciacquicole quasi tutti gli individui sono femmine che producono uova per mitosi; queste uova si sviluppano partenogeneticamente e le rispettive popolazioni sono rappresentate da cloni geneticamente indipendenti.
Nelle forme monogononti si riscontrano, invece, maschi aploidi e due tipi diversi di femmine che possono generare due cicli riproduttivi distinti: ciclo amittico (femmine amittiche o virginopore) e ciclo mittico (femmine mittiche o sessuopore).
Le femmine amittiche producono uova diploidi con guscio sottile che si sviluppano partenogeneticamente; queste uova vengono anche dette "uova subitanee o estive" in quanto vengono prodotte d'estate, quando le condizioni sono favorevoli e si sviluppano in breve tempo. In tal modo si producono, sempre per partenogenesi, più generazioni di femmine amittiche, con conseguente rapido accrescimento della popolazione, già depauperata dalle precedenti condizioni sfavorevoli. Nello stesso periodo alcune femmine amittiche producono una o più generazioni di femmine mittiche le quali, a seguito di meiosi, producono uova aploidi che evolvono in individui maschi. Questi ultimi cominciano a fecondare le uova aploidi presenti ora nella popolazione; queste uova fecondate ("uova durature o invernali") sviluppano un guscio e sono in grado di resistere alle basse temperature e al disseccamento, rimanendo in uno stato di vita (sviluppo) latente sino a quando non si ristabiliscono le condizioni favorevoli, allorquando accelerano il loro sviluppo ed evolvono in femmine amittiche.
Ci sono circa 2 000 specie, raggruppate in tre classi:
Gli Acantocefali, il phylum più strettamente affine ai Rotiferi, sono a volte considerati una quarta classe. Anche gli Gnatostomulidi dovrebbero avere una stretta affinità filogenetica con i Rotiferi, evidenziata da un organo molto simile al mastax. Questi phyla costituiscono un gruppo forse correlato con i Trocozoi, come suggerito dalla presenza di una larva simile ad una trocofora in alcuni, e forse anche agli Ecdisozoi.
Esempi di Bdelloidea
Esempio di Monogononta: Brachionus quadridentatus
I Rotiferi (Rotifera Cuvier, 1798) (dal latino: rota, "ruota" e ferō, "porto") costituiscono un phylum di animali pseudocelomati microscopici. La loro lunghezza tipica è tra 0,1 e 0,5 mm. Sono comuni nelle acque dolci di tutto il mondo, poche sono invece le specie marine. Si spostano nell’acqua contraendosi e allungandosi, facendo leva su due peduncoli posteriori.
Rotifera sunt phylum pseudocoelomatorum animalium microscopicorum et paene microscopicorum. Quae primum a Reverendo Ioanne Harris anno 1696 descripta sunt, variaeque formae aliae ab Antonio van Leeuwenhoek anno 1703 reperta sunt.[1] Plurima Rotifera sunt circa 0.1–0.5 mm longa (sed eorum magnitudo a 50 μm ad plus quam 2 mm variat),[2] et latissime in circumiectis naturalibus aquae dulcis ubique in mundo reperiuntur, nonnullis speciebus aquae salsae exceptis, inter quas insigniter species generis Synchaetae.
Rotifera sunt phylum pseudocoelomatorum animalium microscopicorum et paene microscopicorum. Quae primum a Reverendo Ioanne Harris anno 1696 descripta sunt, variaeque formae aliae ab Antonio van Leeuwenhoek anno 1703 reperta sunt. Plurima Rotifera sunt circa 0.1–0.5 mm longa (sed eorum magnitudo a 50 μm ad plus quam 2 mm variat), et latissime in circumiectis naturalibus aquae dulcis ubique in mundo reperiuntur, nonnullis speciebus aquae salsae exceptis, inter quas insigniter species generis Synchaetae.
Verpetės (lot. Rotifera) – mažyčiai (0,1-0,5 mm ilgio) mikroskopiniai bestuburiai gyvūnai, savo dydžiu galintys prilygti stambiems pirmuonims (Protozoa). Pirmą kartą aprašė John Harris 1696 m., kitas formas 1703 m. aprašė Anton van Leeuwenhoek
Verpetės yra daugialąsčiai gyvūnai, turintys determinuotą ląstelių skaičių (iki 1000). Dauguma verpečių yra apie 0,1–0,5 mm ilgio (nors jų dydis gali varijuoti nuo 50μm iki 2 mm); paplitusios visame pasaulyje gėlo vandens baseinuose - upėse, ežeruose, balose ir grioviuose, tik nedidelė dalis gyvena jūros vandenyje, pvz., Synchaeta gentis.
Minta pirmuonimis ir kitais mikroskopiniais organizmais, kuriuos suvilgo seilių liaukų sekretu. Sausuoju metų laiku verpetės dažniausiai būna anabiozės būsenoje. Taip jos gali išbūti keletą metų. Vikiteka
Virpotāji (Rotifera) ir pirmmutnieku dzīvnieku tips, kas senākās klasifikācijās tiek uzkatīts par veltņtārpu tipa klasi. Šī tipa pārstāvju raksturīga īpatnība ir rotācijas jeb virpotājaparāts ķermeņa priekšējā galā, kas sastāv no skropstiņu gredzeniem. Tas kalpo gan kā barošanās, gan arī kā pārvietošanās līdzeklis. Virpotāji ir sīka izmēra dzīvnieki, kas nepārsniedz 2,5 mm garumu. Virpotāju pārstāvis Ascomorpha minima ir pats sīkākais daudzšūnu organisms, tā garums sastāda tikai ap 40 μm. Sakarā ar virpotāju mazajiem izmēriem to audi bieži vien iegūst sincitiālu uzbūvi. Šim tipam ir raksturīga eitēlija, un tas nozīmē, ka visiem vienas sugas pieaugušajiem īpatņiem ir vienāds šūnu skaits. Dažas virpotāju sugas spēj pāriet kriptobiozes stāvoklī.
Ķermenim pārsvarā ir garena, šķērsgriezumā apaļa vai plakana forma. Dažas formas var būt lodveidīgas (piem. Trochosphaera). Izmēri no 0,04 līdz 2,5 mm. Uz virpotāja ķermeņa virsmas bieži vien ir saskatāms posmveidīgs reljefs. Tomēr tam nav sakara ar ķermeņa metamērismu. Tas parasti ir ārējās segas regulārs garenisks krokojums, kas saistīts ar virpotāju spēju savilkt ķermeni, turklāt, krokas ieiet viena otrā līdzīgi tālskata posmiem. Parasti ķermenim ir labi izšķirama galvas daļa ar virpotājaparātu, viduklis un astes nodalījums jeb kāja.
Galva nav krasi norobežota no vidukļa. Galvas daļa ar īpašu muskuļu - retraktoru palīdzību spēj ievilkties viduklī. Virpotājaparāts galvas daļā vienkāršākā veidā sastāv diviem skropstiņu gredzeniem (priekšējā un aizmugurējā), kā arī skropstiņu laukuma starp tiem. Ja virpotājs brīvi peld ūdenī, tad virpotājaparātam, kas darbojas līdzīgi dzenskrūvei, pārsvarā ir peldēšanas (kustības) orgāna funkcija. Bet, ja virpotājs ir piestiprinājies nekustīgs, tad virpotājaparātam ir galvenokārt laupījuma pievilināšanas funkcija. Tam darbojoties, ūdens plūsma strauji virzās uz virpotāja galvas galu un aizrauj līdzi dažādus sīkus organismus, kuri pēc tam tiek satverti ar muti. Pie galvas ir redzami taustekļi, 1 vai 2 acis un mutes atvere.
Vidukli bieži vien ietver hitīnam līdzīgas vielas čaula. Tā lejas daļā virs kājas pamatnes atrodas anālā atvere. Dažu ģinšu pārstāvjiem pie vidukļa ir izaugumi, kas atvieglo peldēšanu. Viduklī atrodas lielākā iekšējo orgānu daļa.
Kāja ir muskuļots ķermeņa izaugums, kas virpotājam dod iespēju rāpot. Kāja parasti nobeidzas ar divzaru dakšu jeb t.s. pirkstiem. Dakšu galos ir izvietoti dziedzeri, kas izdala lipīgas vielas. Tas ir piestiprināšanās (fiksācijas) orgāns, ar kura palīdzību virpotājs piestiprinas pie substrāta. Kāja ir attīstīta galvenokārt rāpojošām formām.
Ķermeni sedz vienslāņains epitēlijs, kas uz ārpusi izdala kutikulu. Ādas muskuļu slāņa virpotājiem nav. Gludo muskulatūru daudzos gadījumos ir aizvietojusi šķērssvītrotā muskulatūra. Caurspīdīgām formām var saskatīt atsevišķus muskuļus, kas darbina dažādas ķermeņa daļas. Primārais ķermeņa dobums ir labi attīstīts, un sākas uzreiz zem epitēlija.
Virpotājiem ir labi attīstīts kutikulas slānis, kas var būt biezs un stingrs (bruņveidīgs), piedodot dzīvniekam karkasveidīgu formu, vai arī elastīgs, piemērots rāpojošām tārpveida formām. Bruņveidīgās kutikulas bieži vien sastāv no vairākām plātnēm un tām var būt dzelkņi, valnīši u.c. ornamentējums. Kutikulas izaugumi visvairāk ir raksturīgi planktoniskajām formām; šādi dzelkņi palielina ķermeņa virsmu un palīdz noturēties ūdens masā. Sēdošām formām epitēlijs izdala caurulītes "mājiņas" veida apvalkus, kuros sēd dzīvnieks. Caurulīte ir aizsardzības pielāgojums, kur virpotājs var ievilkties tajā.
Nervu sistēmu veido virsrīkles jeb smadzeņu ganglijs, no kura atiet nervu stiegras. Var būt arī zemrīkles un kāju gangliji. Nervi, kas iet uz ķermeņa ārējo segu, ir saistīti ar maņu orgāniem. Pie tiem pieder actiņas un it sevišķi taustes orgāni. Taustes orgāni ir galvas taustekļi, kas ir koniski izaugumi ar sariņu kūlīti virsotnē. Parasti šādi taustekļi ir trīs: ventrālo taustekļu pāris un dorsālais nepāra tausteklis. Actiņas atrodas virsrīkles ganglija rajonā, un tās ir 1 vai 2 invertēta tipa fotoreceptori, kuru uzbūve ir diezgan primitīva. Tikai nedaudzām sugām actiņā ir kaut kas līdzīgs stiklveida ķermenim.
Gremošanas sistēma sākas ar muti (stoma) ķermeņa priekšējā galā ventrālajā pusē, kuru parasti ieskauj skropstiņu aplis (vainags). Aiz mutes dobuma atrodas rīkle (pharynx), kurā ir izvietots barības sasmalcināšanas aparāts - mastakss (mastax). Mastaksa hitīna plātnīšu darbību var novērot dzīviem Rotaria ģints pārstāvjiem. Rīklē atveras siekalu dziedzeri. Aiz rīkles seko īss barības vads (oesophagus), bet aiz tā plašs kuņģis (ventriculus), kurā barība tiek ķīmiski pārstrādāta un rezorbēta. Kuņģī atveras parasti 2 (retāk vairāki) kuņģā dziedzeri. Aiz kuņģa atrodas īsa viduszarna, izklāta šūnām ar garām skropstiņām. Tālāk tā pāriet kloākā vidukļa aizmugurē.
Elpošana notiek ar visu ķermeņa segu. Acīmredzot, ūdens skābeklis tiek ieelpots arī caur zarnu traktu un izvadorgāniem; ūdenim plūstot caur tiem tiek atdots skābeklis.
Izvadsistēmu un osmoregulācijas sistēmu veido protonefrīdiji (parasti divi sānu izvadkanāli), kas beidzas ar kontraktilu urīnpūslī. Upīnpūslis atveras kloākā.
Dzimumorgānu sistēma mātītēm sastāv no olnīcas, dzeltenumdziedzeriem un īsa olvada. Olvads atveras kloākā. Tēviņu dzimumsistēmu veido viens sēklinieks, sēklvads un kopulācijas aparāts, kas nobeidzas kloākā.
Virpotajiem ir novērojams dzimumu dimorfisms. Tēviņi parasti ir mazāki par mātītēm un tiem ir vienkāršāka uzbūve. Olu nogatavošanās (ovoģenēze) vienmēr notiek mātītes organismā, bet dīgļa embrionālā attīstība notiek vai nu ārējā vidē (oldējējiem virpotājiem), vai arī mātītes organismā (dzīvdzemdētājiem). Drostalošanās ir pilnīga, pārveidotā spirāliskā tipa.
Lielākajai virpotāju daļai dzimumvairošanās mijas ar partenoģenētiskā vairošanās. Turklāt partenoģenētiskajām mātītēm attīstās divējādas olas: amiktiskās un miktiskās. Pirmajām ir divkāršs (diploīds) hromosomu komplekts, bet otrajām - vienkāršs (haploīds). Amiktiskās olas attīstās tikai partenoģenētiski. No tām izšķiļas tikai mātītes. Savukārt, no miktiskajām olām, kas attīstās dzimumceļā, izšķiļas tēviņi. Tēviņiem izšķiļoties miktiskās olas tiek apaugļotas un, iegūstot diploīdu hromosomu skaitu, pārvēršas par miera stadijas miktiskajām olām. Šādām olām ir daudz blīvāks apvalks. Pēc olu apaugļošanas tēviņi iet bojā. Attiecības starp šīm divām olu kategorijām ir saistītas ar heterogoniju, kas noris noteikta cikla laikā. Cikls norisinās šādi: no miera stadijas miktiskajām olām attīstās tikai mātītes. Šīs mātītes producē amiktiskās olas, kas atkal attīstās par mātītēm. Tādējādi noteikta perioda laikā rodas vairākas amiktisko mātīšu paaudzes. Šis ir partenoģenētiskās attīstības periods. Pēc tam iestājas periods, kad no amiktiskajām olām sāk attīstīties miktiskās mātītes, kurās attīstās miktiskās olas. No šīm olām partenoģenētiski rodas tēviņi, kas apaugļo miktiskās mātītes. No tām, attiecīgi, attīstās miera stadijas miktiskās olas, un cikls sākas no jauna. Tātad, viena cikla laikā partenoģenētisko paaudzi nomaina paaudze, kas attīstījusies no apaugļotām olām, kuru, savukārt, atkal nomaina partenoģenētiskā paaudze. Dažādām virpotāju sugām, kā arī atkarībā no ūdensbaseina apstākļiem šādu ciklu skaits gada laikā var būt atšķirīgs. Dažām sugām ir viens cikls (monocikliskas formas), citām divi cikli (dicikliskas formas) vai vairāki cikli (policikliskas formas).
Virpotāji ir plaši izplatīti pa visu zemeslodi. Tas lielā mērā ir saistīts ar virpotāju niecīgajiem izmēriem, un vēl vairāk ar miera stadijas olām, kuras ūdensputni vai ūdens dzīvnieki var pārnēsāt lielos attālumos. Tikpat viegli var izplatīties arī augsnes virpotāji, kas izžūstot pāriet anabiozes stāvoklī un ar vēju var tikt pārnesti uz jaunām dzīves vietām. Tāpēc liela daļa virpotāju ir kosmopolīti. Tomēr virpotāju kosmopolītisms nav tik neierobežots, kā varētu likties. Ir zināmas sugas, kuras ir pielāgojušās tikai aukstajiem, tikai tropiskajiem vai tikai mērenajiem apgabaliem. Lielākā virpotaju daļa ir saldūdens formas, bet ir zināmas arī jūras formas. Diezgan daudz virpotāju ir pielāgojušies dzīvei augsnē. Tie apdzīvo mežu augsnes trūdvielu slāni, sūnas u.c. sauszemes ekoloģiskas nišas.
Ūdens virpotāji sastāda būtisku planktona daļu. Šīm planktoniskajām formām parasti ir labi attīstīts virpotājaparāts, vai arī ir gari kutikulas izaugumi, kas palīdz tām notureties ūdens slānī. Savukārt kāja tām ir vāji attīstīta, vai arī vispār tās nav.
Bentiskajām un augsnes formām raksturīga ir rāpošana. Šiem virpotājiem ir garš tārpveidīgs ķermenis un kāja. Virpotāju augsnes formas spēj izžūt un tādā veidā saglabāt dzīvību anabiozes stāvoklī, no kura iziet, nokļūstot mitrā vidē.
Trešo ekoloģisko nišu veido sēdošās formas. Tās piestiprinās pie substrāta ar kāju, turklāt dzīvnieks sēd vai nu plānā gļotainā caurulītē, vai arī blīvākās caurulītēs, kuras virpotājs uzbūvē no dažādu materiālu piciņām, kuras pievelk ar savu virpotājaparātu.
Virkne formu ir pielāgojušās parazītismam. Par piemēru tāds ir Diglena volvocicola, kas parazitē Volvox aureus un virknē citu formu. Parazītisko virpotāju saimnieki ir daži oligohēti, sūneņi, augi u.t.t.
Virpotāji barojas ar dažādiem mikroorganismiem, būdami mikroskopiski plēsoņas. Savairodamies lielā daudzumā virpotaji veicina ūdensbaseinu bioloģisko pašattīrīšanos. Iznīcinādami milzu daudzumu vienšūņu organismu, tie novērš ūdens pārsātināšanu ar organiskajiem produktiem. Turklāt, masveidā savairojušies, virpotāji ir laba barības bāze citiem daudz lielākiem organismiem, piemēram, dažādu zivju mazuļiem. Daudzas virpotāju sugas tiek izmantotas par bioloģiskajiem indikatoriem ūdeņu tīrības pakāpes raksturošanai.
Virpotāji samērā nesen ir izdalīti, ka atsevišķs tips. Iepriekš tie skaitījās veltņtārpu tipa klase. Šis tips tiek dalīts 3 klasēs un 5 kārtās. Pēc dažādiem avotiem ir zināmas 1500-2500 virpotāju sugas. Latvijā ir konstatētas ap 400 virpotāju sugu.
Virpotāji (Rotifera) ir pirmmutnieku dzīvnieku tips, kas senākās klasifikācijās tiek uzkatīts par veltņtārpu tipa klasi. Šī tipa pārstāvju raksturīga īpatnība ir rotācijas jeb virpotājaparāts ķermeņa priekšējā galā, kas sastāv no skropstiņu gredzeniem. Tas kalpo gan kā barošanās, gan arī kā pārvietošanās līdzeklis. Virpotāji ir sīka izmēra dzīvnieki, kas nepārsniedz 2,5 mm garumu. Virpotāju pārstāvis Ascomorpha minima ir pats sīkākais daudzšūnu organisms, tā garums sastāda tikai ap 40 μm. Sakarā ar virpotāju mazajiem izmēriem to audi bieži vien iegūst sincitiālu uzbūvi. Šim tipam ir raksturīga eitēlija, un tas nozīmē, ka visiem vienas sugas pieaugušajiem īpatņiem ir vienāds šūnu skaits. Dažas virpotāju sugas spēj pāriet kriptobiozes stāvoklī.
De raderdieren (Rotifera) zijn een stam van kleine meercellige organismen. Ze behoren tot het onderrijk Eumetazoa en hun lengte bereikt zelden 2 mm. De mannetjes zijn kleiner en eenvoudiger gebouwd dan de vrouwtjes. Er zijn er circa 2180 soorten wetenschappelijk beschreven.[1]
Deze organismen worden gekenmerkt door een trompet- tot bolvormig lichaam, vaak doorzichtig als glas en draagt aan de kop meestal een dubbele krans van trilharen, die worden gebruikt bij de voedselvergaring en voortbeweging. De ventrale mondopening leidt naar de farynx die als kauwmaag dienstdoet. De meeste raderdiertjes leven in water. Een gering aantal leeft in de bodem.
Bij droogte kunnen raderdiertjes in een toestand van schijndood overgaan: cryptobiose. In rusttoestand kunnen raderdiertjes temperaturen van 270 graden Celsius gedurende 5 minuten en 100 graden gedurende enkele uren verdragen. Raderdiertjes vermenigvuldigen zich enkele generaties parthenogenetisch en één maal per jaar geslachtelijk.
De raderdieren (Rotifera) zijn een stam van kleine meercellige organismen. Ze behoren tot het onderrijk Eumetazoa en hun lengte bereikt zelden 2 mm. De mannetjes zijn kleiner en eenvoudiger gebouwd dan de vrouwtjes. Er zijn er circa 2180 soorten wetenschappelijk beschreven.
Hjuldyr er ei rekke protostome pseudocoelomate dyr som lever i vatn over heile verda. Gruppa vart først beskriven i 1696 av John Harris, sjølv om mange kjelder nemner Anton van Leeuwenhoek. Hjuldyra er vanlege i ferskvatn over heile verda, men det finst òg nokre få saltvassartar. Nokre av dei sym fritt i vatnet og er planktoniske, andre kryp rundt på botnen og atter andre er fastsitjande dyr som anten lever i røyr eller heldt seg fast. Om lag 25 artar er koloniale, desse er anten fastsitjande eller planktoniske. Dei fleste hjuldyr er 0,1-0,5 mm lange og såleis berre synlege i mikroskop.
Det finst om lag 2000 artar hjuldyr, fordelt på fem ordenar i to klassar.
Hjuldyra har fått namnet sitt frå coronaen, som sit rundt munnen og er dekt med flimmerhår. Rørslene til desse liknar eit hjul, og skapar ein virvelstraum som syg mat ned i munnen til dyret. Her vert maten toggen opp av ein karakteristisk pharynx med små kjevar. Coronaen driv òg dyret gjennom vatnet når det ikkje sit fast.
Dei fleste frittlevande hjuldyr har par med tær nedst på kroppen som dei brukar til å ankra seg fast medan dei et. Hjuldyr er bilateralsymmetriske og kjem i fleire ulike former. Rundt kroppen har dei eit velutvikla eksoskjelett som anten kan vera tjukt og rørslehemmande eller tynnare og fleksibelt.
Vaksne hjuldyr er euteliske - kvar art har eit bestemt tal på celler per individ, vanlegvis rundt 1000.
Forplantningsmønsteret til hjuldyr varierer etter klasse og miljøtilhøve. Det skjer både kjønna og ukjønna formeiring; i klassa Monogononta tilmed begge delar frå tid til annan. Her er nesten alle individa hoer; hannar opptrer berre under særskilde tilhøve hjå somme artar. Når dei ikkje er til stades skjer reproduksjonen via partenogenese og resulterer i avkom som er genetisk identisk med forelderen.
Hjå nokre artar kan det dannast to ulike slag partenogenetiske egg; den eine sorten utviklar seg til normale partenogenetiske hoer, medan den andre dukkar opp ved endra miljøtilhøve og utviklar seg til ein hann som har eit komplett sett kjønnsorgan men manglar tarm. Desse hannane befruktar hoer i den korte levetida si, og er såleis med på å produsera såkalla kvile-egg. Kvile-egga utviklar seg til zygotar som er i stand til å overleva særs harde miljøtilhøve av slaget som kan inntreffa om vinteren eller under tørketider. Når tilhøva betrar seg att utviklar desse egga seg til ein ny generasjon hoer som lever i mellom eit par dagar til omkring tre veker.
Hjuldyr som høyrer til ordenen Bdelloidea kan ikkje produsera kvile-egg, men mange av dei kan overleva lange periodar med dårlege tilhøve etter uttørking. Dette vert omtalt som anhydrobiose, og er ein kryptobiotisk tilstand. Under tørkeperiodar trekk bdelloidar seg saman til ein urørleg klump og mistar nesten all kroppsvæske; men når dei får vatn i seg att heldt kroppsfunksjonane fram etter få timar. Bdelloidar kan overleva tørre tilhøve over lang tid, den lengste dokumenterte perioden har vore ni år.
Bdelloid-genom inneheldt to eller fleire ulike kopiar av kvart gen, noko som kan tyda på ei lang ukjønna evolusjonær historie. Til dømes har ein funne fire versjonar av hsp82, alle forskjellige og funne på ulike kromosom. Dette umogleggjer homozygotisk kjønna formeiring.
Hjuldyr er ei rekke protostome pseudocoelomate dyr som lever i vatn over heile verda. Gruppa vart først beskriven i 1696 av John Harris, sjølv om mange kjelder nemner Anton van Leeuwenhoek. Hjuldyra er vanlege i ferskvatn over heile verda, men det finst òg nokre få saltvassartar. Nokre av dei sym fritt i vatnet og er planktoniske, andre kryp rundt på botnen og atter andre er fastsitjande dyr som anten lever i røyr eller heldt seg fast. Om lag 25 artar er koloniale, desse er anten fastsitjande eller planktoniske. Dei fleste hjuldyr er 0,1-0,5 mm lange og såleis berre synlege i mikroskop.
Det finst om lag 2000 artar hjuldyr, fordelt på fem ordenar i to klassar.
Hjuldyr er en gruppe mikroskopiske dyr.
Navnet kommer av det såkalte hjulorganet som vifter føde inn i munnen, og som brukes til forflytning. Disse små organismene blir sjelden større enn en millimeter, og lever av døde og levende planter, bakterier og dyr.
Hunnene bærer egg som befruktes og siden klekkes. I noen tilfeller kan det forekomme jomfrufødsel (partenogenese), når ubefruktede egg klekkes.
De lever i 1-2 uker. Naturlige fiender er større dyreplankton og andre vanndyr som lever av mikroskopiske dyr.
Hjuldyr er en gruppe mikroskopiske dyr.
Navnet kommer av det såkalte hjulorganet som vifter føde inn i munnen, og som brukes til forflytning. Disse små organismene blir sjelden større enn en millimeter, og lever av døde og levende planter, bakterier og dyr.
Hunnene bærer egg som befruktes og siden klekkes. I noen tilfeller kan det forekomme jomfrufødsel (partenogenese), når ubefruktede egg klekkes.
De lever i 1-2 uker. Naturlige fiender er større dyreplankton og andre vanndyr som lever av mikroskopiske dyr.
Soa arprodussion as dësrola për mojen d'euv nen fecondà ch'a dan sempe dle fumele. An efet, le popolassion ëd rotìfer a son ëd sòlit formà da 'd fumele ch'as arproduvo sensa la presensa dij mas-cc. Cand ch'a-i rivo dle condission ambientaj dësfavorèivoj ch'a mnasso ëd fé dëscomparì la popolassion, a comparisso antlora ij mas-cc e le fumele a fan dj'euv fecondà dai mas-cc.
Wrotki (Rotifera, zwane też wcześniej jako Rotatoria) – typ, małych (50-2000 µm), przezroczystych zwierząt bezkręgowych z charakterystycznym wieńcem rzęsków (aparatem wrotnym) od których pochodzi nazwa typu i o stałej dla gatunku liczbie komórek ciała (z tego względu, wzrost osobniczy odbywa się nie przez przyrost liczby komórek, tylko przez zwiększenie rozmiarów komórek ciała)[2]. Zwierzęta te zamieszkują przede wszystkim wody słodkie, ale znane są też formy morskie, słonowodne i lądowe, żyjące na wilgotnych mchach, w glebie kielichach roślin i dziuplach z wodą, czy na torfowiskach lub w piasku w wodzie interstycjalnej[3]. Żywią się martwą materią organiczną, bakteriami, glonami i pierwotniakami. W Polsce występują 554 gatunki[4], na świecie ponad 2000[2].
Ciało wrotka podzielone jest na trzy odcinki: głowę, tułów i nogę. Mają najczęściej kształt wydłużony z szerokim przednim końcem, tzw. aparatem wrotnym. Aparat wrotny lub rzęskowy ma postać tarczy z rozmieszczonymi na niej rzęskami. Tylny koniec ciała zwęża się tworząc krótszą lub dłuższą nogę zakończoną dwoma palcami. Ciało pokryte jest cienką kutikulą. Warstwa okrywająca ciało jest na tyle przejrzysta, że widoczne są przez nią narządy wewnętrzne wrotka. Po stronie grzbietowej głowy oraz po bokach tułowia znajdują się brodawki czuciowe zakończone szczecinkami. Otwór gębowy położony jest na tarczy aparatu, otwór odbytowy u nasady nogi. Aparat wrotny składa się z dwóch nieciągłych pasm rzęsek schodzących do zagłębienia otworu gębowego oraz kilku rzędów membranelli położonych bliżej strony grzbietowej.
Bezpośrednio pod cienką hypodermą znajduje się jama ciała wypełniona płynem, w której położone są wszystkie narządy wewnętrzne oraz pęczki mięśni. Mięśnie wrotka utworzone są przeważnie przez pojedyncze wielkie komórki.
Układ pokarmowy rozpoczyna się otworem gębowym leżącym na brzusznej stronie aparatu wrotnego. Tuż za otworem znajduje się silnie umięśniona gardziel (mastaks). W gardzieli widoczny jest wyraźnie aparat zastawkowy, składający się z trzech chitynowych zębów. Wszystkie części aparatu żującego podłączone są ze sobą i z gardzielą licznymi mięśniami. Gardziel przechodzi w krótki, urzęsiony przełyk, łączący się z dużym, również urzęsionym żołądkiem. Do przedniej części żołądka otwiera się jeden gruczoł zwany gruczołem trzustkowo-wątrobowym. Krótkie urzęsione jelito uchodzi do szerokiej kloaki, otwierającej się odbytem u nasady nogi.
Układ wydalniczy składa się z dwóch rurkowatych protonefrydiów leżących po bokach przewodu pokarmowego. Rurki te na wysokości żołądka skręcają się w pętlę i uchodzą do pęcherza moczowego. Pęcherz otwiera się do kloaki. System protonefridialny oprócz utrzymania równowagi wodnej, eliminuje też azotany, amoniak i fosforany, choć teoretycznie membrana powinna zadbać o zatrzymanie ich. Zjawisko to określane jako wydzielanie (w odróżnieniu od wydalania) jest znane też jako regeneracja fosforu i azotu, i wrotki mogą w ten sposób przywrócić do obiegu około 10-20% fosforu i 20-50% azotu w środowisku wodnym, skutecznie konkurując w tym względzie z bakteriami[3].
Rozmnażanie i układ rozrodczy. Większość wrotków jest rozdzielnopłciowa i występuje u nich dymorfizm płciowy. Grupa Bdelloidea rozmnaża się aseksualnie i samce w tej grupie nie są znane, natomiast Sensonidea rozmnażają się wyłącznie biseksualnie[3]. U pozostałych wrotków samce mają uwstecznione narządy wewnętrzne, u większości gatunków przewód nasienny kończy się prąciem. Jajniki mogą być pojedyncze lub parzyste, jajowody otwierają się do kloaki. Wrotki w warunkach niekorzystnych przechodzą w stan anabiozy lub encystacji, lub znoszą jaja przetrwalne, które mogą przetrwać setki lat w niekorzystnych warunkach, by podjąć rozwój w warunkach sprzyjających[3]. Wrotki mają także zdolność do rozmnażania się w drodze partenogenezy przez wiele pokoleń, bez udziału samców.
Żeński układ rozrodczy składa się z jajnika i żółtka, połączonych ze sobą, leżących po brzusznej stronie żołądka. Na tle jelita widoczny jest krótki jajowód uchodzący do kloaki. Samce są znacznie mniejsze i prościej zbudowane od samic. U wielu gatunków i w grupie Bdelloidea samce nie zostały dotychczas odkryte. Samce, jeżeli występują, pojawiają się tylko co pewien okres i to bardzo nieliczne.
Tradycyjnie wrotki zaliczane były (w randze gromady lub podtypu) do typu obleńców (Nemathelminthes). Aktualnie zazwyczaj traktowane są jako osobny typ, w którym wyróżniono dwie gromady:
Gromada Eurotatoria (parafiletyczna)
Gromada Pararotatoria
Czasem wyróżnia się też formy kuliste Sphaeroida, ale gromada ta zawiera tylko jeden gatunek – trochosferę.
Molekularne dane wskazują, że Rotifera jest grupą parafiletyczną. Podgrupę wrotków bowiem tworzą kolcogłowy (Acanthocephala). Razem z kolcogłowami wrotki tworzą grupę monofiletyczną (klad), zwany na przykład „Syndermata”.
Jedna z nowszych hipotez zakłada wspólne pochodzenie wrotków, kolcogłowów i szczękogębych. Zwierzęta te tworzą klad Gnathifera[5].
Na podstawie:
Halanych et al. Evidence from 18S ribosomal DNA that the lophophorates are protostome animals. „Science”. 267 (5204), s. 1641–1643, 1995. DOI: 10.1126/science.7886451 (ang.).
Edgecombe et al. Higher-level metazoan relationships: recent progress and remaining questions. „Organisms Diversity and Evolution”. 11, s. 151–172, 2011. DOI: 10.1007/s13127-011-0044-4 (ang.).
Wrotki (Rotifera, zwane też wcześniej jako Rotatoria) – typ, małych (50-2000 µm), przezroczystych zwierząt bezkręgowych z charakterystycznym wieńcem rzęsków (aparatem wrotnym) od których pochodzi nazwa typu i o stałej dla gatunku liczbie komórek ciała (z tego względu, wzrost osobniczy odbywa się nie przez przyrost liczby komórek, tylko przez zwiększenie rozmiarów komórek ciała). Zwierzęta te zamieszkują przede wszystkim wody słodkie, ale znane są też formy morskie, słonowodne i lądowe, żyjące na wilgotnych mchach, w glebie kielichach roślin i dziuplach z wodą, czy na torfowiskach lub w piasku w wodzie interstycjalnej. Żywią się martwą materią organiczną, bakteriami, glonami i pierwotniakami. W Polsce występują 554 gatunki, na świecie ponad 2000.
Os Rotifera ou rotíferos (do latim rota, roda + fera, do grego "phoros", aqueles que possuem) são um filo de animais aquáticos e microscópicos. O seu nome deriva do latim para “roda”, com referência à coroa de cílios que rodeiam a boca destes animais e que se movem rapidamente, para captar as partículas de alimento, parecendo uma roda a girar.
Por causa do seu pequeno tamanho e falta de peças duras, os rotíferos não fossilizam facilmente e o mais antigo que se conhece foi encontrado em âmbar da República Dominicana e data do período Eoceno, mas são comuns fósseis da espécie Habrotrocha angusticollis em turfa com 6000 anos (período Pleistoceno) em Ontário, no Canadá.
Conhecem-se cerca de 1700 espécies de rotíferos de vida livre, que vivem na maior parte das massas de água doce, incluindo pequenas poças de chuva, no solo húmido e também se encontram em musgos e líquenes que crescem em troncos de árvores e pedras, ou mesmo sobre fungos, crustáceos ou larvas aquáticas de insectos. Algumas espécies nadam livremente na água, mas outras são sésseis, agarrando-se a qualquer substrato, seja ele fixo ou flutuante.Os rotíferos são encontrados em todos os biomas terrestres, desde poças de água de chuva até grandes lagos de água fresca, também no meio de grãos de areia, e poucos em água salgada. São animais protostômios que são incluídos na meiofauna por serem multicelulares. A maioria das espécies é planctônica e podem contribuir com até 30% da biomassa total do plâncton. Em conseqüência disso, o conhecimento sobre a biologia dos rotíferos é baseado nas espécies planctônicas (RICCI; BALSAMO, 2000).
Eles estão em praticamente todos os lagos, lagoas e rios, em pedaços de musgos na floresta úmida, nas regiões tropicais e até mesmo em terrenos gelados do Ártico. Um lugar praticamente garantido para encontrar alguns é na beira de uma lagoa ou lago. Há rotíferos marinhos, mas a grande maioria ocorrem em água doce. Rotíferos possuem cerca de 0,1-0,5 mm de comprimento (embora o seu tamanho pode variar de 50 μm a mais de 2 milímetros).[1] Alguns rotíferos são de natação livre e verdadeiramente planctônicas, outros se movem ao longo de um substrato, e alguns são sésseis, vivendo dentro de tubos ou superfícies gelatinosas que estão ligados a um substrato. Cerca de 25 espécies são coloniais (por exemplo, Sinantherina semibullata), sésseis ou planctônicas. Rotíferos são uma parte importante do zooplâncton de água doce, sendo um dos principais e com muitas espécies também contribuindo para a decomposição da matéria orgânica do solo.[2] Em alguns estudos mais recentes, os rotíferos são classificados com acantocéfalos em um clado maior chamado Syndermata. Os membros da classe Bdelloidea encontram-se ocasionalmente em água salobra ou marinha. Estes rotíferos são capazes de sobreviver à dessecação, através do processo chamado criptobiose (ou anidrobiose), assim como os seus ovos – de facto, os embriões mais velhos têm mais possibilidade de vingar.
Possui o corpo alongado, cilíndrico ou sacular e é, na maioria das espécies, dividido em uma curta região anterior da cabeça, um estreito pescoço, grande tronco que constitui a maior parte do corpo e um pé terminal . Seu corpo é frequentemente dividido em uma série de anéis transversais que subtraem em diâmetro anterior e posteriormente a partir do maior anel no centro do tronco. Os músculos longitudinais ocasionam encaixe dos anéis do pé uns nos outros e no tronco, para encurtar o corpo. Um par de músculos retratores puxa a cabeça para o tronco[3] A Corona ciliada é como um anel epidérmico celular ciliado que percorre ao longo da cabeça. De um modo geral, a corona apresenta a parte ventral do anel , chamada campo bucal, que circunda a boca. A faixa circum-apical dorsal se estende do campo bucal até o topo da cabeça, e a extremidade anterior ciliada da cabeça, rodeada pela corona, é campo apical.[4] Uma estrutura que também é característica do grupo é o mástax, que constitui a faringe muscular do animal e é composto por sete peças duras chamadas trofos. Estas peças são modificadas de acordo com o tipo de nutrição do animal. A função do mástax é a de triturar os alimentos para posterior digestão . O mástax possui muitos músculos que controlam um complexo conjunto de mandíbulas, que consiste em proteínas endurecidas, ligeiramente flexível, como cutícula.O aparelho mandíbula é chamado de trophi e tritura a comida trazida para baixo pela ação ciliar. O trophi consiste de uma série de estruturas que mostram variações consideráveis. Em algumas espécies essas estruturas podem estar arranjadas em formatos complexos . Existem sulcos pequenos, travessas, e saliências sobre as pequenas projecções dos componentes que podem ser usados para identificar espécies, mas em muitos casos são distintos . (RUPERT; BARNES, 1996, WELCH, 2000).
Os olhos, as antenas e as aberturas das glândulas cerebrais estão situados no campo apical. As células epiteliais da corona são multiciliadas. Há sete variedades no padrão básico da corona, a coroa de cílios pode estar reduzida, aumentada, modificada e até mesmo ausente em alguns rotíferos. Quando ausente a espécie se alimenta por suspensão através de uma fileira de grandes cílios compostos denominada troco.[5] A cabeça de algumas espécies rotíferas possui uma projeção mediano-dorsal chamada rostro. O rostro é como uma pequena projeção formada por cílios e cerdas sensoriais em sua extremidade e também é aderente .[6].[7]
Abaixo da cutícula há uma epiderme sincicial, que segrega a cutícula, e bandas de músculos subepidérmicos, alguns circular, alguns longitudinal, e alguns correndo através da pseudocele para os órgãos viscerais (Fig. 09).A cutícula verdadeira é uma secreção extracelular da epiderme que, em geral, funciona como um esqueleto para proteção e ligação dos músculos,[8] A cutícula dos rotíferos, não contem colágeno ou quitina e é uma fina camada glicoproteica sem significado esquelético.[9] Em alguns rotíferos, a epiderme produz uma lâmina intra-sincicial, essa lâmina funciona como esqueleto, dando proteção e um lugar para a ligação 26 muscular. Esta lâmina é chamada lorica. A lorica pode ser dividida em placas distintas e seções aneliformes e costuma ser ornamentada com sulcos, espinhos ou apêndices articulados.[5]. Abaixo da epiderme há uma musculatura corporal muito desenvolvida composta por fibras transversais estriadas e lisas, as fibras se estendem de uma unidade do esqueleto epidérmico ate a outra e se ligam por tonofilamentos que passam pela epiderme, aos hemidesmossomos nas laminas intra-sincicial[10]
Os rotíferos movem-se por rastejamento, pelo batimento ciliar da corona ou saltando pela água usando apêndices especializados, pois a grande maioria habita o fundo da água ou vive em vegetação submersa .A corona é retraída quando o animal rasteja e o pé se adere ao substrato, usando a secreção adesiva das glândulas podais, então estende o corpo, fixa o rostro, libera o pé para se mover para frente e novamente se prende ao substrato com o pé, o resultando em uma locomoção por mede-palmo, como uma sanguessuga, por exemplo.Os rotíferos planctônicos nadam continuamente e são adaptados a se manter em uma posição favorável na coluna d’água A boca dos rotíferos é anterior e ventral geralmente rodeada por parte da corona. Ela se abre para um tubo bucal ciliado ou, nas espécies que se alimentam por suspensão, diretamente para a faringe. A faringe apresenta um conjunto de mandíbulas denominado mástax uma característica peculiar dos rotíferas. O mástax é utilizado para capturar e processar mecanicamente o alimento e como é típica de partes bucais, sua estrutura varia consideravelmente de acordo com táxon
Entre os rotíferos, os bdelóideos possuem o ciclo de vida mais simples. As fêmeas são diploides produzem apenas um tipo de ovo, enquanto os machos são ausentes e a reprodução é assexuada, por partenogênese,as fêmeas produzem ovos diploides por mitose e os ovos eclodem com mais fêmeas diploides. Já em rotíferos monogonontes, o ciclo de vida é mais complexo, pois a reprodução pode ser bissexual ou por partenogênese, e vários tipos de ovos podem ser produzidos.[11] No ciclo partenogenético ou amíctico, uma fêmea amíctica diploide produz um ovo de casca fina, que não pode ser fertilizado e consequentemente se desenvolve em uma fêmea idêntica a mãe, esse ovo é denominado ovo amíctico, de rápida eclosão de 12 a 24 horas.[12] Um segundo tipo de ovo descrito é o ovo monogononte, denominado ovo míctico, que também possui casca fina, mas provem da meiose e, portanto é haploide, se esses ovos não forem fertilizados, eles se desenvolverão em machos haploides.[13] Os ovos haploides, fertilizados por machos produzem cascas densas e resistentes, estes ovos são chamados de ovos dormentes. Estes ovos são diploides e eventualmente irão eclodir em fêmeas diploides. Os ovos dormentes tem um embrião em dia pausa, capaz de resistir a dessecação, temperaturas extremas e outras condições adversas e pode não eclodir por vários meses, ou anos.[7]. Os rotíferos exibem uma clivagem espiral modificada. A divisão nuclear é completada cedo no desenvolvimento e os indivíduos eclodem com o número de núcleos característico de cada espécie. O crescimento pós-embrionário é resultado do aumento do tamanho ou espaço das células, não adição de novas células. O desenvolvimento é determinado e direto sem estagio larval verdadeiro. .[14]
Ciclomorfose - Ruppert et al. (2005), tem uma visão peculiar na qual descreve a ciclomorfose dos rotíferos.De acordo com o autor, vários rotíferos pelágicos sofrem mudanças sazonais na forma ou nas proporções do corpo, um fenômeno conhecido como ciclomorfose. Durante uma estação do ano, indivíduos de algumas espécies possuem espinhos mais longos ou mais curtos que aqueles desenvolvidos por seus descendentes geneticamente idênticos durante outra estação como descreve Ruppert et al. (2005).
Em Brachionus sp, segundo Ruppert et al. (2005), os espinhos de gerações subsequentes podem ser induzidos por escassez de alimento, baixas temperaturas e cairomonios produzidos pelo rotífero predador Asplanchna sp. Espinhos maiores protegem o o Brachionus de ser predado. Mudanças morfológicas similares são observadas em cladoceros e em alguns dinoflagelados.
Criptobiose Os metazoários que habitam poças temporárias e solos úmidos se deparam com a ameaça da desidratação e outros extremos físicos, estes indivíduos desenvolvem adaptações reprodutivas, como no caso dos rotíferos, ovos resistentes, que permite que o embrião sobreviva em condições extremas, mesmos que os adultos morram.[15] Para uma criptobiose bem-sucedida, o animal deve dissecar lentamente, por vários dias, enquanto o animal seca, a forma do corpo muda para diminuir o volume e minimizar a área da superfície, os rotíferos diminuem cerca de 25% de seu tamanho normal[16] A criptobiose é verdadeiramente um estado de morte, o metabolismo é infinitamente baixo, na ausência de metabolismo e em desidratação quase total, os animais criptobioticos estão essencialmente mortos, mas capazes de reviver, por esse motivo os rotíferos sobrevivem em ambientes extremos.[17]
Rotiferos tem um cérebro pequeno, localizado logo acima da mastax, a partir do qual um certo número de nervos se estende por todo o corpo. O número de nervos varia entre espécies, embora o sistema nervoso normalmente tenha uma disposição simples. Perto do cérebro se encontra um órgão retrocerebral, consistindo de duas glândulas de ambos os lados de um saco medial. O saco drena para um duto que se divide em dois antes de abrir através de poros na parte superior da cabeça. Sua função não é clara.
Rotíferos normalmente possuem um ou dois pares de antenas curtas e até cinco olhos. Os olhos são simples na estrutura, às vezes com apenas um único células fotorreceptoras. Além disso, as cerdas da coroa é sensível ao toque, e também há um par de pequenos buracos revestidas pelos cílios sensoriais, na região da cabeça.[18]
Rev. John Harris primeiro descreveu os rotíferos em 1696 como "um animal como um grande verme que pode contrair-se em uma figura esférica e, em seguida, estender-se para fora de novo, no final de sua cauda apareceu com um fórceps, como que de uma tesourinha ". .[19] Em 1702, Anton van Leeuwenhoek deu uma descrição detalhada de Rotífero vulgaris e posteriormente descrito ringens Melicerta e outras espécies. Ele também foi o primeiro a publicar observações de certas espécies após a secagem . Outras formas foram descritas por outros observadores, mas não foi até a publicação do Christian Gottfried Ehrenberg Die Infusionsthierchen als vollkommene Organismen em 1838 que os rotíferos foram reconhecidos como sendo animais multicelulares.[20] Cerca de 2.200 espécies de rotíferos têm sido descritos. Sua taxonomia está atualmente em um estado de fluxo. O filo rotifera foi subdivididos em três classes: Seisonidea, Bdelloidea e Monogononta[21]
O grupo maior é a Monogononta, com cerca de 1500 espécies, seguida pela Bdelloidea, com cerca de 350 espécies.. Existem apenas dois gêneros conhecidos com três espécies de Seisonidea. O Acanthocephala, anteriormente considerada um filo separado, têm sido demonstrado ser rotíferos modificados. A relação exacta de outros membros do filo ainda não foi resolvido. Uma possibilidade é que o Acanthocephala estão mais próximos do Bdelloidea Monogononta e que ao Seisonidea;. Os nomes e as relações correspondentes são mostradas na cladogram presente nesse página.
O Rotifera, estritamente falando, estão confinados à Bdelloidea e o Monogonata. Rotifera, Acanthocephala e Seisonida compõem um clado chamado Syndermata.[22]
O estado de São Paulo possui 236 espécies de rotíferos das 457 encontradas no Brasil (aproximadamente metade). O conhecimento atual dos rotíferos necessita, porém, de intensa revisão na identificação de algumas espécies descritas, assim como expansão das áreas de coletas. A maioria dos trabalhos considera, em geral, pontos de amostragem na região limnética dos lagos e reservatórios (de onde estão descritas a maior parte das espécies ou os gêneros de rotíferos), negligenciando importantes microhábitats presentes em regiões litorâneas, em meio às macrófitas, locais rasos e outros (além dos rios que foram satisfatoriamente amostrados)(Projeto Biota Fapesp www.biota.org.br/iRead).
Rotíferos são considerados organismos oportunistas encontradas em alta densidade na maioria dos reservatórios superando os outros grupos do zooplâncton, várias espécies apresentam uma grande capacidade para se adaptar às mudanças ambientais.[23] São comumente registrados para ambientes com altos teores de nutrientes, tendo preferência por ambientes eutrofizados, principalmente reservatórios de água doce.
Encontra-se na biota aquática, variados indicadores biológicos, pois o meio vem sendo agredido constantemente pela ação humana, através de uso indiscriminado de fertilizante agrícola, dejetos industriais, escoamento de esgoto, lixo urbano, entre vários fatores.Entre os bioindicadores estudados do meio aquático, destacam-se os rotíferos.
Desta forma, investigações sobre a comunidade de rotíferos planctônicos, animais microscópicos, foram desenvolvidos tendo como objetivo identificar a presença destes seres indicadores da qualidade ambiental. sendo importante também por constituir o zooplâncton, fundamental para alimentação e sobrevivência de outros animais aquáticos. Amplamente utilizados como indicadores de poluição e das condições ambientais os rotíferos tornam-se animais de grande valor ecológico .Estes metazoários executam importante papel na restauração de nutrientes e na transferência de energia nas teias alimentares, estão diretamente envolvidos no crescimento das populações de bactérias e fungos abrangidos na remineralização de nutrientes.[24]
O principal uso desses animais na economia atual é o de alimento para larvas de peixes, pois os rotíferos têm ciclo de vida rápido e tem um alto teor energético, sendo mais eficientes na alimentação de larvas de peixes do que dietas artificiais (LUBZENS, ZMORA; BARR, 2000). Outro importante campo de pesquisa que envolve o grupo é o da ecotoxicologia em que são usados dados da dinâmica de população para avaliar a influência de determinados fatores abióticos sobre a população de rotíferos. As populações focais deste tipo de estudo são espécies de rotíferos e o gênero mais usado nesses ensaios é o Brachionus (Yúfera, 2001). Esse tipo de estudo que analisa as alterações na sua composição populacional natural decorrentes de variações ambientais, demonstram a possibilidade de uso de rotíferos como bioindicadores (Projeto Biota Fapesp www.biota.org.br/iRead).
Com base em certas similaridades morfológicas, os rotíferos e os acantocéfalos (os vermes parasitas que constituem o filo Acanthocephala) foram durante muito tempo considerados como parentes e estudos recentes das sequências do gene rRNA 18S corroboraram este parentesco. Estes dois grupos foram durante algum tempo classificados como pseudocelomados, juntamente com uma série de outros vermes, mas as últimas análises filogenéticas puseram em causa esta hipótese dos Pseudocoelomata serem um grupo natural. De fato, muitos animais pseudocelomados, como os priapulídeos e os nemátodes, parecem ter relações muito mais próximas dos artrópodes num grupo chamado Ecdysozoa, enquanto que outros animais com um pseudoceloma, como os rotíferos e os acantocéfalos, parecem estar mais próximos dos moluscos, anelídeos e braquiópodes, num grupo denominado Lophotrochozoa.
O filo Rotifera é dividido em três classes: Monogononta, Bdelloidea e Seisonidea, entre os quais o maior é o primeiro, com cerca de 1500 espécies, seguido pelos Bdelloidea, com cerca de 350 espécies. Conhecem-se apenas duas espécies de Seisonidea, que são normalmente consideradas como mais "primitivas".
|coautores= (ajuda) Os Rotifera ou rotíferos (do latim rota, roda + fera, do grego "phoros", aqueles que possuem) são um filo de animais aquáticos e microscópicos. O seu nome deriva do latim para “roda”, com referência à coroa de cílios que rodeiam a boca destes animais e que se movem rapidamente, para captar as partículas de alimento, parecendo uma roda a girar.
Por causa do seu pequeno tamanho e falta de peças duras, os rotíferos não fossilizam facilmente e o mais antigo que se conhece foi encontrado em âmbar da República Dominicana e data do período Eoceno, mas são comuns fósseis da espécie Habrotrocha angusticollis em turfa com 6000 anos (período Pleistoceno) em Ontário, no Canadá.
Conhecem-se cerca de 1700 espécies de rotíferos de vida livre, que vivem na maior parte das massas de água doce, incluindo pequenas poças de chuva, no solo húmido e também se encontram em musgos e líquenes que crescem em troncos de árvores e pedras, ou mesmo sobre fungos, crustáceos ou larvas aquáticas de insectos. Algumas espécies nadam livremente na água, mas outras são sésseis, agarrando-se a qualquer substrato, seja ele fixo ou flutuante.Os rotíferos são encontrados em todos os biomas terrestres, desde poças de água de chuva até grandes lagos de água fresca, também no meio de grãos de areia, e poucos em água salgada. São animais protostômios que são incluídos na meiofauna por serem multicelulares. A maioria das espécies é planctônica e podem contribuir com até 30% da biomassa total do plâncton. Em conseqüência disso, o conhecimento sobre a biologia dos rotíferos é baseado nas espécies planctônicas (RICCI; BALSAMO, 2000).
Eles estão em praticamente todos os lagos, lagoas e rios, em pedaços de musgos na floresta úmida, nas regiões tropicais e até mesmo em terrenos gelados do Ártico. Um lugar praticamente garantido para encontrar alguns é na beira de uma lagoa ou lago. Há rotíferos marinhos, mas a grande maioria ocorrem em água doce. Rotíferos possuem cerca de 0,1-0,5 mm de comprimento (embora o seu tamanho pode variar de 50 μm a mais de 2 milímetros). Alguns rotíferos são de natação livre e verdadeiramente planctônicas, outros se movem ao longo de um substrato, e alguns são sésseis, vivendo dentro de tubos ou superfícies gelatinosas que estão ligados a um substrato. Cerca de 25 espécies são coloniais (por exemplo, Sinantherina semibullata), sésseis ou planctônicas. Rotíferos são uma parte importante do zooplâncton de água doce, sendo um dos principais e com muitas espécies também contribuindo para a decomposição da matéria orgânica do solo. Em alguns estudos mais recentes, os rotíferos são classificados com acantocéfalos em um clado maior chamado Syndermata. Os membros da classe Bdelloidea encontram-se ocasionalmente em água salobra ou marinha. Estes rotíferos são capazes de sobreviver à dessecação, através do processo chamado criptobiose (ou anidrobiose), assim como os seus ovos – de facto, os embriões mais velhos têm mais possibilidade de vingar.
Rotiferele (Rotifera sau Rotatoria) (din latina rota = roată + ferre = a purta) sunt o încrengătură de nevertebrate acvatice microscopice, înrudite cu nematodele, nesegmentate, pseudocelomate, cu dimensiuni între 0,04–3 mm, cu corpul alungit, cilindric sau mai rar sferic, având la partea anterioară una sau două coroane de cili puternici, care se învârtesc ca o roată, de unde și numele, servind la antrenarea particulelor alimentare spre gură și la mișcare. Se cunosc circa 2 500 specii, majoritatea de apă dulce, puține marine, litorale și terestre (în mușchi, licheni). Unele dintre rotifere sunt forme libere, fie plutitoare sau înotătoare (planctonice), fie târându-se pe fundul apei (bentonice). Alte forme bentonice sunt sesile, adică fixate pe fundul apei prin extremitatea lor posterioară și, în cele mai dese cazuri, înconjurate de tuburi protectoare. Puține specii sunt parazite, unele la plante, altele la animale.
Corpul rotiferelor este împărțit în trei părți distincte: una anterioară, numită regiunea cefalică (cap), una mijlocie, numită trunchi și una posterioară, numită picior (coadă). Piciorul se termină de obicei cu o furcă, constituită din 2-4 degete, la extremitatea cărora se deschide câte o glandă cimentară (pedioasă). Cu ajutorul secreției produsă de aceste glande, animalul se poate fixa temporar pe suport. Extremitatea cefalică și piciorul poate fi retras în interiorul corpului prin telescoparea inelelor cuticulare, cu ajutorul unor mușchi speciali.
Organul caracteristic al rotiferelor este aparatul rotator situat în regiunea cefalică cu ajutorul căruia se deplasează activ în masa apei. Aparatul rotator este format din una sau două coroane concentrice de ciliatură: trocus (situat preoral) și cingul (situat postoral), între care se găsește un șanț. Corpul este acoperit la mai multe specii de o platoșă cuticulară numită lorica.
Rotiferele au un număr definit, specific, de celule. Astfel, Epiphanes senta are 959 celule.
Aparatul digestiv este format din orificiul bucal, faringe, esofag, stomac, intestin, cloacă, care se deschide la exterior prin orificiul anal. În faringe se află un aparat masticator, numit mastax, care constă din 7 piese numite trofi sau maxile: una impară, fulcru, 2 rami, 2 unci și 2 manubrii. Fulcrul și ramii formează incusul (nicovala), iar uncii și manubriile maleusul (ciocanul).
Aparatul respirator și circulator nu există. Aparatul excretor este reprezentat prin protonefridii și este format din 4-50 celule cu flamură vibratilă, două tuburi colectoare ce se unesc într-o vezică urinară care se deschide printr-un tub scurt în cloacă pe partea ventrală.
Organele de simț sunt reprezentate prin unul sau doi oceli, situați pe aparatul rotator și prin palpii ciliați. Sistemul nervos este format din ganglionul cerebroid, ganglionul mastaxului, ganglionul pedios. Ganglionul cerebroid este legat de ganglionul pedios prin două cordoane nervoase laterale.
Aparatul genital feminin la monogononte este format dintr-un un singur ovar sincițial și un vitelariu (germovitelariu) alcătuit din ovogonii degenerate, care se continuă cu un oviduct ce se deschide în cloacă, înaintea orificiului excretor; bdeloidele au germovitelariu pereche și un oviduct comun. Masculii nu se cunosc la bdeloide, la monogononte masculii sunt pigmei și au un singur testicul, un conduct spermatic, un gonopor și un cir sau penis. La seisonide masculii și femelele sunt aproape identici, gonadele sunt perechi la masculi și la femele; ovarele sunt lipsite de vitelariu. Fecundația se face prin impregnare hipodermică: penisul înțeapă corpul femelei într-un punct oarecare, injectând sperma în pseudocel.
Rotiferele au o durată de viață scurta, de 2-3 săptămâni. În perioada caldă a anului se dezvoltă mai multe generații succesive. Iarna sau în timpul secetei rezistă sub formă de ouă. Din astfel de ouă, numite ouă durabile, ies primăvara numai femele, care se înmulțesc toată vara partenogenetic, adică fără prezența masculilor. Când condițiile de mediu se schimbă, în special când temperatura apei ajunge maximă, ceea ce se întâmplă de obicei vara, tot partenogenetic apar și masculii care se împerechează imediat cu femelele generației care le-au dat naștere. Ouăle rezultate în urma fecundării rezistă peste iarnă, din ele se dezvoltă o nouă generație de femele în primăvara următoare.
Hrana rotiferelor constă din microorganisme vegetale sau animale, care sunt aduse la gură o dată cu curentul de apă produs de organul rotator. Rotiferele constituie o bună parte a microplanctonului și o hrană prețioasă pentru animale, în special pentru puietul mic de pește, ceea ce este foarte important pentru piscicultură. Prin modul lor de hrană, ele contribuie la curățirea apelor de bacteriile cu care se hrănesc.
După Marius Skolka, rotiferele se împart în două clase: Pararotatoria (Prorotatoria) și Eurotatoria.[1]
Clasificarea rotiferelor după Zhi-Qiang Zhang[2]
Încrengătura Rotifera (Cuvier, 1817) — 2 clase
Lista alfabetică a rotiferelor din România
Rotiferele (Rotifera sau Rotatoria) (din latina rota = roată + ferre = a purta) sunt o încrengătură de nevertebrate acvatice microscopice, înrudite cu nematodele, nesegmentate, pseudocelomate, cu dimensiuni între 0,04–3 mm, cu corpul alungit, cilindric sau mai rar sferic, având la partea anterioară una sau două coroane de cili puternici, care se învârtesc ca o roată, de unde și numele, servind la antrenarea particulelor alimentare spre gură și la mișcare. Se cunosc circa 2 500 specii, majoritatea de apă dulce, puține marine, litorale și terestre (în mușchi, licheni). Unele dintre rotifere sunt forme libere, fie plutitoare sau înotătoare (planctonice), fie târându-se pe fundul apei (bentonice). Alte forme bentonice sunt sesile, adică fixate pe fundul apei prin extremitatea lor posterioară și, în cele mai dese cazuri, înconjurate de tuburi protectoare. Puține specii sunt parazite, unele la plante, altele la animale.
Corpul rotiferelor este împărțit în trei părți distincte: una anterioară, numită regiunea cefalică (cap), una mijlocie, numită trunchi și una posterioară, numită picior (coadă). Piciorul se termină de obicei cu o furcă, constituită din 2-4 degete, la extremitatea cărora se deschide câte o glandă cimentară (pedioasă). Cu ajutorul secreției produsă de aceste glande, animalul se poate fixa temporar pe suport. Extremitatea cefalică și piciorul poate fi retras în interiorul corpului prin telescoparea inelelor cuticulare, cu ajutorul unor mușchi speciali.
Organul caracteristic al rotiferelor este aparatul rotator situat în regiunea cefalică cu ajutorul căruia se deplasează activ în masa apei. Aparatul rotator este format din una sau două coroane concentrice de ciliatură: trocus (situat preoral) și cingul (situat postoral), între care se găsește un șanț. Corpul este acoperit la mai multe specii de o platoșă cuticulară numită lorica.
Rotiferele au un număr definit, specific, de celule. Astfel, Epiphanes senta are 959 celule.
Aparatul digestiv este format din orificiul bucal, faringe, esofag, stomac, intestin, cloacă, care se deschide la exterior prin orificiul anal. În faringe se află un aparat masticator, numit mastax, care constă din 7 piese numite trofi sau maxile: una impară, fulcru, 2 rami, 2 unci și 2 manubrii. Fulcrul și ramii formează incusul (nicovala), iar uncii și manubriile maleusul (ciocanul).
Aparatul respirator și circulator nu există. Aparatul excretor este reprezentat prin protonefridii și este format din 4-50 celule cu flamură vibratilă, două tuburi colectoare ce se unesc într-o vezică urinară care se deschide printr-un tub scurt în cloacă pe partea ventrală.
Organele de simț sunt reprezentate prin unul sau doi oceli, situați pe aparatul rotator și prin palpii ciliați. Sistemul nervos este format din ganglionul cerebroid, ganglionul mastaxului, ganglionul pedios. Ganglionul cerebroid este legat de ganglionul pedios prin două cordoane nervoase laterale.
Aparatul genital feminin la monogononte este format dintr-un un singur ovar sincițial și un vitelariu (germovitelariu) alcătuit din ovogonii degenerate, care se continuă cu un oviduct ce se deschide în cloacă, înaintea orificiului excretor; bdeloidele au germovitelariu pereche și un oviduct comun. Masculii nu se cunosc la bdeloide, la monogononte masculii sunt pigmei și au un singur testicul, un conduct spermatic, un gonopor și un cir sau penis. La seisonide masculii și femelele sunt aproape identici, gonadele sunt perechi la masculi și la femele; ovarele sunt lipsite de vitelariu. Fecundația se face prin impregnare hipodermică: penisul înțeapă corpul femelei într-un punct oarecare, injectând sperma în pseudocel.
Rotiferele au o durată de viață scurta, de 2-3 săptămâni. În perioada caldă a anului se dezvoltă mai multe generații succesive. Iarna sau în timpul secetei rezistă sub formă de ouă. Din astfel de ouă, numite ouă durabile, ies primăvara numai femele, care se înmulțesc toată vara partenogenetic, adică fără prezența masculilor. Când condițiile de mediu se schimbă, în special când temperatura apei ajunge maximă, ceea ce se întâmplă de obicei vara, tot partenogenetic apar și masculii care se împerechează imediat cu femelele generației care le-au dat naștere. Ouăle rezultate în urma fecundării rezistă peste iarnă, din ele se dezvoltă o nouă generație de femele în primăvara următoare.
Hrana rotiferelor constă din microorganisme vegetale sau animale, care sunt aduse la gură o dată cu curentul de apă produs de organul rotator. Rotiferele constituie o bună parte a microplanctonului și o hrană prețioasă pentru animale, în special pentru puietul mic de pește, ceea ce este foarte important pentru piscicultură. Prin modul lor de hrană, ele contribuie la curățirea apelor de bacteriile cu care se hrănesc.
Vírniky (lat. Rotifera, Syndermata, Rotatoria) sú kmeň prvoústovcov.
Systém vírnikov je v každom ohľade sporný. Napríklad:
Podrobnosti o systéme pozri nižšie.
Sú to drobné, väčšinou len mikroskopom viditeľné živočíchy. Na povrchu tela je kutikula s redukovaným obrvením – zostali z neho len dva vence bŕv v prednej časti.
Telo sa skladá z hlavy, krčka (často býva redukovaný), trupu a nohy. Prisadnuté druhy majú na konci nohy cementózne žľazy, ktoré produkujú tuhnúci sekrét. Napriek mikroskopickým rozmerom sú orgánové sústavy dosť zložito stavané. Za ústami je trojhranný hltan so špecializovaným žuvacím aparátom mastax, tvoria ho pevné kutikulárne doštičky. Nervová sústava je pásová, zo zmyslov pozorujeme receptorické brvy a výrastky v prednej časti tela, niekedy primitívne ploché očká. Vylučovacími orgánmi sú protonefrídie, spájajú sa do močového mechúra, ktorý ústi do kloaky – je to spoločný vývod vylučovacej, tráviacej a rozmnožovacej sústavy. Mezenchým je syncytiálny a tvorí sa v ňom nevýrazný schizocoel.
Vírniky sú gonochoristy, samce sú menšie ako samičky, u mnohých druhov chýbajú. U mnohých druhov pozorujeme heterogóniu – striedanie niekoľkých partenogenetických generácií samíc s pohlavnou generáciou na jeseň, podobne ako u vošiek. Vývin je priamy. Je to kmeň charakteristický eutéliou – stálym (a pomerne nízkym) počtom buniek jednotlivých druhov. Žijú v moriach i v sladkých vodách, často aj vo vlhkom machu. Za východiskovú skupinu pre fylogenézu vírnikov sa považuje čeľaď Notonemmatidae – podobajú sa na maličké ploskule a ešte viac na brušnobrvky – ich telo je ešte nezreteľne diferencované na hlavu, trup a nohu. Lezú po dne pomocou obrveného políčka v prednej časti tela (kontinuita s Turbellaria), z ktorého vznikol obrvený aparát vyšších vírnikov.
V starých systémoch sa taxón, ktorý je predmetom tohto článku, zaraďoval (bez Acanthocephala):
V súčasných systémoch sa (vrátane alebo bez Acanthocephala) obyčajne zaraďuje ako kmeň Rotifera (Syndermata) do (prípadnej skupiny Gnathifera a tá do) nadkmeňa špirálovce (Spiralia). [5][6]
Systém podľa Grzimek et al. 1971: [3]
trieda Rotatoria:
Systém podľa Nogrady et al. 1993, Segers 2002 a pod.: [7]
kmeň Rotifera:
Systém podľa Markevich (1990):[7]
Rotifera:
Systém podľa Ruppert et al. 2004: [8]
Syndermata:
Systém podľa rôznych autorov: [5][9]
Syndermata:
Systém podľa Franc 2005 (v tejto wikipédii to je primárny systém): [1]
kmeň vírniky (Rotifera) - jediná trieda vírniky (Rotatoria):
Systém podľa Ruggiero et al. 2015 :[6]
kmeň Rotifera:
Systém podľa Tirjaková 2015: [5]
kmeň vírniky (Rotifera):
Systém podľa Sielaff 2016: [10]
a) jedna možnosť (Fig. 2):
Syndermata (Rotifera):
b) druhá možnosť (Fig. 4) [takmer rovnaký systém má Sørensen & Giribet (2006)]:
Syndermata (Rotifera):
Vírniky (lat. Rotifera, Syndermata, Rotatoria) sú kmeň prvoústovcov.
Systém vírnikov je v každom ohľade sporný. Napríklad:
v niektorých systémoch kmeň vírniky zahŕňa len jedinú triedu vírniky (lat. Rotatoria), t.j. bez háčikohlavcov; v iných systémoch zahŕňa triedu vírniky (Rotatoria) a triedu háčikohlavce (Acanthocephala), t.j. je rozšírený o taxón háčikohlavce; v iných systémoch je rozdelený na dve alebo viac inak pomenovaných a koncipovaných tried, pričom buď zahŕňa alebo nezahŕňa háčikohlavce; v niektorých systémoch sa rozlišuje medzi taxónom Syndermata a kmeňom Rotifera (kmeň Rotifera v takom prípade nezahŕňa buď Acanthocephala alebo ani Seisonida).Podrobnosti o systéme pozri nižšie.
Hjuldjur (Rotifera) är mikroskopiska vattenlevande flercelliga djur.
Namnet kommer av det så kallade hjulorganet som viftar ned föda i munnen och som används till förflyttning. Dessa små organismer som sällan blir större än en millimeter livnär sig på både döda och levande växter och djur. Trots att hjuldjuren är så små har de en matsmältningskanal. Även födan är i litet format - mikroskopiska växter, djurplankton och bakterier. Hjuldjur finns i många sötvattensmiljöer såsom sjöbottnar, likväl som i rinnande vattenmiljöer, men även i fuktig jord. Där finns de i de tunna vattenskikten som ligger runt jordpartiklarna. Hjuldjur är även vanliga i avloppsrör, vattenpölar och på svampar som växer nära döda träd. Så trots att hjuldjuren är vattenlevande, kan man finna dem nästan var som helst där det finns vatten. Vissa arter klarar även salt och bräckt vatten. Det finns en oerhörd mångfald och man får nästan garanterat med dem i alla sötvattenprover.
Hjuldjurets förökning är enkel och går till på så sätt att honorna bär på ägg som befruktas och sedan kläcks. I vissa fall kan det förekomma jungfrufödsel (partenogenes), när ägg som inte är befruktade kläcks.
Hos de flesta hjuldjur är antalet hanar nedsatt, och de kan till och med vara obefintliga. Vissa arter lägger två typer av ägg: en sort som utvecklas till normala honor, medan den andra utvecklas till en degenererad hane som inte kan föda sig själv och existerar endast för uppgiften att producera sperma.
Livslängden för honor är omkring en till två veckor.
Fiender är större djurplankton och andra vattendjur som livnär sig på mikroskopiska djur.
Det finns omkring 1800 arter, indelade i tre klasser.
De flesta arter är av fastsittande typ, en längre eller kortare period. De sitter fast med ett slags fot på bakkroppen. Andra flyter omkring som plankton.
Hjuldjur (Rotifera) är mikroskopiska vattenlevande flercelliga djur.
Namnet kommer av det så kallade hjulorganet som viftar ned föda i munnen och som används till förflyttning. Dessa små organismer som sällan blir större än en millimeter livnär sig på både döda och levande växter och djur. Trots att hjuldjuren är så små har de en matsmältningskanal. Även födan är i litet format - mikroskopiska växter, djurplankton och bakterier. Hjuldjur finns i många sötvattensmiljöer såsom sjöbottnar, likväl som i rinnande vattenmiljöer, men även i fuktig jord. Där finns de i de tunna vattenskikten som ligger runt jordpartiklarna. Hjuldjur är även vanliga i avloppsrör, vattenpölar och på svampar som växer nära döda träd. Så trots att hjuldjuren är vattenlevande, kan man finna dem nästan var som helst där det finns vatten. Vissa arter klarar även salt och bräckt vatten. Det finns en oerhörd mångfald och man får nästan garanterat med dem i alla sötvattenprover.
Hjuldjurets förökning är enkel och går till på så sätt att honorna bär på ägg som befruktas och sedan kläcks. I vissa fall kan det förekomma jungfrufödsel (partenogenes), när ägg som inte är befruktade kläcks.
Hos de flesta hjuldjur är antalet hanar nedsatt, och de kan till och med vara obefintliga. Vissa arter lägger två typer av ägg: en sort som utvecklas till normala honor, medan den andra utvecklas till en degenererad hane som inte kan föda sig själv och existerar endast för uppgiften att producera sperma.
Livslängden för honor är omkring en till två veckor.
Fiender är större djurplankton och andra vattendjur som livnär sig på mikroskopiska djur.
Det finns omkring 1800 arter, indelade i tre klasser.
De flesta arter är av fastsittande typ, en längre eller kortare period. De sitter fast med ett slags fot på bakkroppen. Andra flyter omkring som plankton.
Rotifera, çarklı hayvanlar ya da tekerlekli hayvanlar; solucan benzeri mikroskobik omurgasız hayvanlar şubesidir.
Tatlısularda bulunurlar. Vücut, gövde ve ayaktan oluşur. Sindirim sistemleri gelişmiştir. Başlarında halka şeklinde bir sil demeti bulunur. Bu siller hareketleri sırasında tekerlek görüntüsündedirler. Rotifera'da erkek bireyler çok küçülmüş ya da yokturlar. Yaklaşık 1.500-2.000 türü bilinmektedir.
На передньому кінці тіла розташований навколоротовий апарат. Він в простому варіанті складається з двох віночків війок (трохус (передній) і цингулум (задній)) і війкового поля між ними. Головний відділ може втягуватися за допомогою спеціальних м'язів-ретракторов. Тулуб містить велику частину внутрішніх органів. На його кінці, над основами ноги розташований отвір клоаки.
Нога — це м'язистий виріст тіла, що дозволяє коловерткам рухатися. На кінці ноги розташовується пара «пальців», в основі яких відкриваються цементуючі залози: таким чином, з їх допомогою тварина може прикріплятися до субстрату.
Покриви представлені синцитієм, на периферії якого розташовується термінальна мережа з актиноподібних філаментів. На поверхні розташований шар глікокаліксу, який може іноді досягати значної товщини. Синцитій підстилає базальна пластинка, під якою знаходиться первинна порожнина тіла.
Шкірно-мускульний мішок у коловерток відсутній, є окремі, переважно поперечно-смугасті, м'язові волокна.
Ротовий отвір розташований на черевній стороні між віночками війок навколоротового апарату. Він веде в ротову порожнину і потім в м'язисту глотку (мастакс) з характерним для коловерток жувальним апаратом, утвореним «зубами» з полісахаридів (у хижих форм вони можуть висуватися). У глотку відкриваються слинні залози. За глоткою йде стравохід, а за ним — ентодермальний шлунок, в передню частину якого відкриваються травні залози. Шлунок з'єднується з середньою кишкою, клітини якої мають довгі джгутики. Далі розташовується задня кишка з анальним отвором (фактично вона є клоакою, оскільки в неї впадають протоки статевої і видільної систем). Деякі коловертки (2 роди) можуть повторно втрачати задню кишку.
Видільна система представлена протонефридіями (від 4 до 50 циртоцитів). Їх протоки впадають в сечовий міхур, який відкривається в клоаку.
Присутній один виражений надглотковий ганглій, від якого йдуть нервові стовбури до різних органів, але також можуть бути нервові скупчення в районі ноги і статевої системи. Основними органами чуття є головні щупальця — конічні горбки з пучком війок на верхівці. У загальному їх три: пара вентральних і дорсальна (тім'яна).
У районі надглоткового ганглія у більшості є 1 або 2 фоторецептори інвертованого типу, будова яких достатньо примітивна (тільки у небагатьох є подібність склоподібного тіла).
Коловертки — роздільностатеві організми. Статева система самки представлена цілісним (клас Monogononta) або дволопатевим (клас Digononta) яєчником, що знаходиться в задній частині тіла, під кишкою. Яєчник підрозділений на частину, що власне продукує яйцеклітини і жовток. Від яєчника відходить короткий яйцепровід, що впадає в клоаку. Самці володіють єдиним сім'яником; від нього відходить сім'япровід, дистальна частина якого проходить в копулятивному органі, що вивертається.
У різних систематичних групах коловерток статевий диморфізм виражений по-різному. Так у представників класу Seisonidea самці і самки не дуже розрізняються на вигляд. Навпаки, у коловерток з класу Monogononta самці карликові, не живляться, і з внутрішніх органів у них представлена тільки гонада. У представників класу Digononta самці зовсім відсутні, а розмноження йде тільки шляхом партеногенезу.
Розвиток прямий, без метаморфозу. Дроблення дуже рано стає двобічно-симетричним. Після періоду ембріонального розвитку клітини перестають ділитися — з цим пов'язана постійність їх клітинного складу (також через це у коловерток відсутня здатність до регенерації).
Життєвий цикл (у Monogononta) є гетерогонією, тобто чергуванням партеногенетичного і статевого розмноження. У стабільних, сприятливих умовах в популяції присутні тільки самки, які розмножуються партеногенезом. За несприятливих умов з'являються гаплоїдні самці. В результаті запліднення утворюються яйця, що мають товсту оболонку. З цих яєць виходять партеногенетичні самки. Самки також виділяють спеціальний фермент. Коловертки можуть утворювати цисти, за допомогою яких можуть розповсюджуватися по земній кулі, перепливаючи океани.[2] Деякі класи коловерток (наприклад, бделоїдні — Bdelloidea) десятки мільйонів років обходяться без статевого розмноження. Дивовижно, але за відсутності цього важливого, як традиційно вважається, фактору мінливості ці класи збагачуються десятками нових видів. Припускається — через здатність використовувати гени інших малих істот.
Luân trùng hay Trùng bánh xe là những động vật khoang giả, có kích thước hiển vi. Hầu hết luân trùng có chiều dài từ 0,1 – 0,5 mm, sống phổ biến trong các khu vực nước ngọt, bên cạnh đó có một số loài sống ở nước mặn.
Tên gọi "trùng bánh xe" bắt nguồn từ tên Latin Rotifera có nghĩa là bánh xe. Luân trùng có cấu tạo từ một vài búi lông mao xung quanh miệng có chức năng vận động giống như một bánh xe đang quay. Chúng tạo ra một dòng nước cuốn thức ăn vào trong miệng, ở đó thức ăn được nghiền bởi một hầu chuyên hoá (còn được gọi là mastax) có chứa những chiếc hàm răng nhỏ xíu. Hầu hết luân trùng sống tự do có các cặp chân hậu để gắn chúng vào giá thể trong khi ăn.
Hình dạng của luân trùng rất đa dạng. Lớp biểu bì của chúng phát triển tốt, lớp này có thể dày và cứng làm chúng có dạng như một cái hộp hoặc mềm dẻo trông như những con giun. Những dạng luân trùng trên được gọi là luân trùng có mai (vỏ cứng) hoặc luân trùng không mai. Nhiều loài luân trùng có khả năng bơi và một số dạng luân trùng không mai di chuyển theo cách giun bò trên các cơ chất. Các loại luân trùng khác không có cuống, chúng sống bên trong các vỏ bằng gelatin. Có khoảng 25 loài có dạng sống tập đoàn, các loài khác sống trôi nổi (sinh vật phù du). Cũng như nhiều động vật có kích thước hiển vi khác, luân trùng trưởng thành có một số lượng tế bào cố định đặc trưng cho loài.
Hầu hết ở các loài luân trùng thì giống đực thường rất ít, thậm chí có thể biến mất trong trường hợp sinh sản đơn tính. Ở một số loài, quá trình sinh sản đơn tính tạo ra hai loại trứng; một loại trứng phát triển thành giống cái bình thường trong khi loại còn lại phát triển thành dạng giống đực thoái hoá, chúng thậm chí không dinh dưỡng và chỉ tồn tại để sinh ra tinh trùng. Trong các loài này, trứng được thụ tinh tạo thành dạng hợp tử chống chịu khô hạn có khả năng sống sót trong điều kiện khô hạn của các ao hồ, chúng chỉ phát triển thành thế hệ giống cái khi các điều kiện sống thuận lợi trở lại. Thời gian sống của con cái trong khoảng 1 – 2 tuần. Luân trùng là một trong nhiều dạng động, thực vật chịu khô hạn. Chúng có thể sống sót sau một thời gian dài khô hạn và các điều kiện bất lợi khác. Khi điều kiện môi trường trở nên bất lợi, luân trùng kết bào xác dạng trứng (bao nang), khi gặp môi trường nước chúng thoát ra trở thành dạng sinh vật bơi tự do. Ở dạng bào xác, chúng có thể sống thiếu nước trong một thời gian dài, có thể là sống tiềm sinh vô hạn. Sự kích thích do nước thường diễn ra rất nhanh, thường không đầy 2 giờ đồng hồ. Quá trình sinh hoá này được hoàn thành đồng thời với khả năng tạo ra đường trehalose (đường trihaloza - một dạng đường kiểu 1-alpha) của luân trùng để tạo ra trạng thái dạng gel giúp cho các bào quan tránh được những tác động bất lợi do điều kiện mất nước này. Quá trình này cũng thấy ở nhiều sinh vật khác như ngành động vật bò chậm (Tardigrada), loài giáp xác Artemia salinis.
Có khoảng 2.000 loài luân trùng, được chia thành 3 lớp. Động vật đầu móc ký sinh (Acanthocephala) cũng có thể coi là thuộc nhóm luân trùng. Những ngành này thuộc nhóm được gọi là động vật dạng dẹt (Platyzoa). Những loài luân trùng thuộc lớp Bdelloidea là nhóm sinh vật đa bào có số lượng cá thể đông đảo thứ hai trên Trái Đất sau nhóm đông nhất là ngành giun tròn (Nematoda).
Luân trùng hay Trùng bánh xe là những động vật khoang giả, có kích thước hiển vi. Hầu hết luân trùng có chiều dài từ 0,1 – 0,5 mm, sống phổ biến trong các khu vực nước ngọt, bên cạnh đó có một số loài sống ở nước mặn.
Tên gọi "trùng bánh xe" bắt nguồn từ tên Latin Rotifera có nghĩa là bánh xe. Luân trùng có cấu tạo từ một vài búi lông mao xung quanh miệng có chức năng vận động giống như một bánh xe đang quay. Chúng tạo ra một dòng nước cuốn thức ăn vào trong miệng, ở đó thức ăn được nghiền bởi một hầu chuyên hoá (còn được gọi là mastax) có chứa những chiếc hàm răng nhỏ xíu. Hầu hết luân trùng sống tự do có các cặp chân hậu để gắn chúng vào giá thể trong khi ăn.
Hình dạng của luân trùng rất đa dạng. Lớp biểu bì của chúng phát triển tốt, lớp này có thể dày và cứng làm chúng có dạng như một cái hộp hoặc mềm dẻo trông như những con giun. Những dạng luân trùng trên được gọi là luân trùng có mai (vỏ cứng) hoặc luân trùng không mai. Nhiều loài luân trùng có khả năng bơi và một số dạng luân trùng không mai di chuyển theo cách giun bò trên các cơ chất. Các loại luân trùng khác không có cuống, chúng sống bên trong các vỏ bằng gelatin. Có khoảng 25 loài có dạng sống tập đoàn, các loài khác sống trôi nổi (sinh vật phù du). Cũng như nhiều động vật có kích thước hiển vi khác, luân trùng trưởng thành có một số lượng tế bào cố định đặc trưng cho loài.
Hầu hết ở các loài luân trùng thì giống đực thường rất ít, thậm chí có thể biến mất trong trường hợp sinh sản đơn tính. Ở một số loài, quá trình sinh sản đơn tính tạo ra hai loại trứng; một loại trứng phát triển thành giống cái bình thường trong khi loại còn lại phát triển thành dạng giống đực thoái hoá, chúng thậm chí không dinh dưỡng và chỉ tồn tại để sinh ra tinh trùng. Trong các loài này, trứng được thụ tinh tạo thành dạng hợp tử chống chịu khô hạn có khả năng sống sót trong điều kiện khô hạn của các ao hồ, chúng chỉ phát triển thành thế hệ giống cái khi các điều kiện sống thuận lợi trở lại. Thời gian sống của con cái trong khoảng 1 – 2 tuần. Luân trùng là một trong nhiều dạng động, thực vật chịu khô hạn. Chúng có thể sống sót sau một thời gian dài khô hạn và các điều kiện bất lợi khác. Khi điều kiện môi trường trở nên bất lợi, luân trùng kết bào xác dạng trứng (bao nang), khi gặp môi trường nước chúng thoát ra trở thành dạng sinh vật bơi tự do. Ở dạng bào xác, chúng có thể sống thiếu nước trong một thời gian dài, có thể là sống tiềm sinh vô hạn. Sự kích thích do nước thường diễn ra rất nhanh, thường không đầy 2 giờ đồng hồ. Quá trình sinh hoá này được hoàn thành đồng thời với khả năng tạo ra đường trehalose (đường trihaloza - một dạng đường kiểu 1-alpha) của luân trùng để tạo ra trạng thái dạng gel giúp cho các bào quan tránh được những tác động bất lợi do điều kiện mất nước này. Quá trình này cũng thấy ở nhiều sinh vật khác như ngành động vật bò chậm (Tardigrada), loài giáp xác Artemia salinis.
Коловратки (лат. Rotifera) — тип многоклеточных животных, ранее относимых к группе первичнополостных червей. Основным характерным признаком является наличие так называемого коловращательного аппарата — ресничного образования на переднем конце тела, который используется для питания и движения.
Известно около 1500 видов коловраток, в России — около 600 видов[1]. В основном это пресноводные обитатели, но также они водятся в море и влажных почвах. Большинство коловраток свободноподвижные, но есть и прикреплённые формы. Подавляющее большинство видов свободноживущие, но встречаются также паразиты и комменсалы.
По размеру коловратки не превышают 2 мм (в среднем гораздо меньше). Представитель этого типа Ascomorpha minima — самое мелкое многоклеточное животное, размер его составляет около 40 микрон. В связи с малыми размерами ткани часто приобретают синцитиальное строение. Для типа в целом характерна эутелия — постоянство клеточного состава, то есть постоянное число клеток у всех особей одного вида. Некоторые коловратки способны впадать в криптобиоз.
На переднем конце тела расположен коловращательный аппарат. Он в простейшем варианте состоит из двух венчиков ресничек (трохус (передний) и цингулюм (задний)) и ресничного поля между ними. Головной отдел может втягиваться с помощью специальных мышц-ретракторов.
Туловище содержит большую часть внутренних органов. На его конце, над основанием ноги расположено отверстие клоаки.
Нога — это мускулистый вырост тела, позволяющий коловраткам ползать. На конце ноги располагается пара «пальцев», в основании которых открываются цементные железы: таким образом, с их помощью животное может прикрепляться к субстрату.
Покровы представлены синцитием, на периферии которого располагается терминальная сеть из актиноподобных филаментов. На поверхности расположен слой гликокаликса, который может иногда достигать значительной толщины. Синцитий подстилает базальная пластинка, под ней находится первичная полость тела.
Кожно-мускульный мешок у коловраток отсутствует, имеются отдельные, большей частью поперечнополосатые мышечные волокна.
Ротовое отверстие расположено на брюшной стороне между венчиками ресничек коловращательного аппарата. Оно ведёт в ротовую полость и затем в мускулистую глотку (мастакс) с характерным для коловраток жевательным аппаратом, образованным «зубами» из полисахаридов (у хищных форм они могут выдвигаться). В глотку открываются слюнные железы. За глоткой идёт пищевод, а за ним — энтодермальный желудок, в переднюю часть которого открываются пищеварительные железы. Желудок соединяется со средней кишкой, клетки которой несут длинные реснички. Дальше располагается задняя кишка с анальным отверстием (фактически она является клоакой, поскольку в неё впадают протоки половой и выделительной системы). Некоторые коловратки (2 рода) могут вторично терять заднюю кишку.
Выделительная система представлена протонефридиями (от 4 до 50 циртоцитов); их протоки впадают в мочевой пузырёк, который открывается в клоаку.
Присутствует один выраженный надглоточный ганглий, от которого идут нервные стволы к разным органам; но также могут быть нервные скопления в районе ноги и половой системы. Основными органами чувств являются головные щупальца — конические бугорки с пучком ресничек на вершине. В общем случае их три: пара вентральных и непарное дорсальное (теменное).
В районе надглоточного ганглия у большинства имеются 1 или 2 фоторецептора инвертированного типа, строение которых достаточно примитивно (только у немногих имеется подобие стекловидного тела).
Коловратки раздельнополы. Половая система самки представлена цельным (подкласс Monogononta) или двулопастным (подкласс Bdelloidea=Digononta) яичником, лежащим в задней части тела, под кишкой. Яичник подразделён на часть собственно продуцирующую яйцеклетки и желточник. От яичника отходит короткий яйцевод, впадающий в клоаку. Самцы обладают единственным семенником; от него отходит семяпровод, дистальная часть которого проходит в выворачивающемся копулятивном органе.
В разных систематических группах коловраток половой диморфизм выражен по-разному. Так у представителей класса Pararotatoria самцы и самки не слишком различаются по внешнему виду. Напротив, у коловраток из подкласса Monogononta самцы карликовые, не питающиеся, и из внутренних органов у них представлена только гонада. У представителей подкласса Bdelloidea самцы вовсе отсутствуют, а размножение идёт только путём партеногенеза.
Развитие прямое, без метаморфоза. Дробление очень рано становится билатерально-симметричным. После периода эмбрионального развития клетки перестают делиться — с этим связано постоянство клеточного состава коловраток (также из-за этого у коловраток отсутствует способность к регенерации).
(на примере класса Monogononta)
Жизненный цикл представляет собой гетерогонию, то есть чередование партеногенетического и полового размножения.
В стабильных, благоприятных условиях в популяции присутствуют только самки, которые размножаются партеногенезом, то есть производя на свет таких же самок. В неблагоприятных же условиях (например, угроза пересыхания водоёма, приближение морозов) появляются гаплоидные самцы, оплодотворяющие самок, в результате чего образуются «зимние» или покоящиеся яйца с толстой защитной оболочкой. Из этих яиц выходят впоследствии (при наступлении благоприятных условий) партеногенетические самки, и цикл повторяется.
Некоторые классы коловраток (например, бделлоидные — Bdelloidea) десятки миллионов лет обходятся без полового размножения. Удивительно, но в отсутствие этого важнейшего, как традиционно считается, фактора изменчивости, эти классы обогащаются десятками новых видов. Предположительно — из-за способности использования генов других мелких существ (горизонтальный перенос генов).
В типе выделяют следующие таксоны до отряда включительно[2]:
Коловратки (лат. Rotifera) — тип многоклеточных животных, ранее относимых к группе первичнополостных червей. Основным характерным признаком является наличие так называемого коловращательного аппарата — ресничного образования на переднем конце тела, который используется для питания и движения.
Известно около 1500 видов коловраток, в России — около 600 видов. В основном это пресноводные обитатели, но также они водятся в море и влажных почвах. Большинство коловраток свободноподвижные, но есть и прикреплённые формы. Подавляющее большинство видов свободноживущие, но встречаются также паразиты и комменсалы.
По размеру коловратки не превышают 2 мм (в среднем гораздо меньше). Представитель этого типа Ascomorpha minima — самое мелкое многоклеточное животное, размер его составляет около 40 микрон. В связи с малыми размерами ткани часто приобретают синцитиальное строение. Для типа в целом характерна эутелия — постоянство клеточного состава, то есть постоянное число клеток у всех особей одного вида. Некоторые коловратки способны впадать в криптобиоз.
見内文
轮形动物门(學名:Rotifera),又稱輪蟲動物門,是动物界的一个门。是主要生活在淡水中的小型动物,约有1800种左右。轮形动物在假体腔动物中是相当繁盛的一类。
身体短圆,有明亮的壳,两侧对称,身体的后端多数有尾状部;前端有一纤毛盘,具有运动功能;纤毛摆动时状如旋转的轮盘,所以得名;咽内具有咀嚼器,无真体腔。
輪形動物各子類群之間的關係尚未明確[1]。 傳統上按照生殖器特徵分爲兩大類群三個綱:
有的分類法中將搖輪蟲目(Seisonidea)划到雙巢綱(Digononta)之下。
轮形动物门(學名:Rotifera),又稱輪蟲動物門,是动物界的一个门。是主要生活在淡水中的小型动物,约有1800种左右。轮形动物在假体腔动物中是相当繁盛的一类。
身体短圆,有明亮的壳,两侧对称,身体的后端多数有尾状部;前端有一纤毛盘,具有运动功能;纤毛摆动时状如旋转的轮盘,所以得名;咽内具有咀嚼器,无真体腔。

輪形動物(りんけいどうぶつ)は、いわゆるワムシ類と総称される動物の分類群である。
輪形動物門 (Rotifera) は、ワムシと呼ばれる水中の微小動物からなる動物群である。主として淡水に生息し、若干の海産種や陸生種がある。多くは1 mm に満たず、たいていは100-500 μm 程度の大きさである。浮遊生活か、藻類や沈殿物の表面を匍匐して暮らしている。一部に固着性の種がある。世界で約3,000種が知られる。
単為生殖をする種が多く、雄が常時出現する例は少ない。雄が全く見られない群もある。なお、雄は雌よりはるかに小さく、形態も単純で消化管等も持たない。以下の構造等の記述は主として雌に関するものである。
壷型の胴体と、後方に伸びる尾部を持ち、頭にある繊毛を使って運動する。全体としては左右相称で、腹背の区別はあるが、さほどはっきりしない例もある。
体の先端部は幅広く、ここには繊毛が円をなして配置し、繊毛冠(Corona)を形成する。この繊毛は摂食にも運動にも使われる。この部分は形態的にははっきりしない場合もあるが、頭部と言われる。眼点や特殊な感覚器を備える例もある。
頭部に続く部分は胴部で、円筒形から壷型、内臓の大部分がここに収まる。体表はキチン質の表皮に覆われる。がっしりとした被甲に覆われる例も多い。
それに続く尾部は、いくつかの節に分かれて、よく伸縮する。先端に二本の指と爪があり、また粘液腺などを持って体を支えるのに使われる。匍匐性の種では胴と同じくらいの幅と長さを持ち、節があるものもあるが、体節とは認められていない。また、この付近に卵をぶら下げて活動するものがよくある。
多くのものは付属肢を持たないが、ミジンコワムシは二対の付属肢があり、それを使って泳ぐ。また、可動の棘を持つものもある。ミツウデワムシは胴部前方に一対、後方に一本の棘があり、この前方の一対を大きく動かして撥ねるように泳ぐ。
消化系は直線的。繊毛冠の中央に口が開き、胴部の前端付近に咽頭部がある。この部分は厚い筋肉に覆われ、石灰質の咀嚼板が組合わさって咀嚼器を構成している。 それに続いて胃と腸があり、肛門は胴部の後端にある。フクロワムシは腸と肛門を欠く。
胴部の内部を広く占める体腔内は、消化系の表面に上皮層を欠くので偽体腔である。また、縦に走る筋肉がよく発達し、これによって体を伸び縮みさせ、よく運動する。
神経系としては咀嚼嚢の背面に脳神経節があり、ここから全身に末梢神経が走る。 排出系は原腎管を左右一対持ち、その末端は肛門につながる膀胱に開く。生殖巣は消化器の腹側にあり、やはり肛門に口を開く。なお、雄ではこの位置に陰茎がある。
基本的には水中動物であり、陸で見られるものも、特に湿った状態の時に出現する。繊毛を動かしてデトリタスなどを集めて食べているものが多いが、植物の汁を吸うもの、捕食性で原生動物や他のワムシ類などを捕らえるものも知られる。寄生性のものも知られている。
多くは自由生活で、浮遊性のものもあれば、基質上をはい回ることの多いものもある。繊毛を動かして泳ぐか、尾部で基質表面に付着し、尾を動かして運動する。ヒルガタワムシは頭部と尾部を使い、ヒルやシャクトリムシのように這う。ヒルガタワムシは乾燥にさらされると脱水して乾眠とよばれる無代謝状態になる。
固着性の種もあり、それらは基質表面に棲管を作り、そこに体をいれ、伸び出して管の口から繊毛冠を広げる。その仲間で変わっているのはテマリワムシで、多数個体が互いに尾の先端でくっつき合い、それが寒天質に包まれてくす玉のような群体となり、水中を回転しながらただよう。
多くの種が単為生殖をする。それらは条件のいい間は夏卵と言われる殻の薄い卵を産み、この卵はすぐに孵化して雌となり、これを繰り返す。条件が悪化するなどの場合には減数分裂が行われて雄が生まれ、受精によって生じた卵は休眠卵となる。休眠卵は乾燥にも耐え、条件がよくなれば孵化する。なお、ヒルガタワムシ類では雄は全く知られていない。他方、ウミヒルガタワムシでは雄が常時存在することが知られる。
なお、単為生殖を繰り返す期間に、殻の角が伸びるなど表現型が世代を繰り返す間に変化する例があり、周期的体型輪廻(英語版)と言われる。
特に害をなす局面はない。
野外ではさまざまな水域で生息密度も高く、小魚等の重要な餌になっている。また、デトリタス食のものは水の浄化にも効果が高いと言われる。特にシオミズツボワムシは人工培養も行われており、アルテミアよりも小さな生き餌として、養魚場等では重宝される。
偽体腔を持つとされ、かつて袋型動物とされたこともある。その中では小型であること、繊毛で運動する点で腹毛動物に似ている。原腎管を持つ点では扁形動物と共通点がある。
その体制をトロコフォアと対比させる説が唱えられたこともある。
なお、近年の系統研究から鉤頭虫がこの門に含まれるべきとの提案もなされている。
すべてを輪形動物綱にまとめ、伝統的には生殖器によって三群に分けるが、あまり安定した体系はないようである。以下に旧来の分類体系を示す。
輪虫綱 Rotatoria
輪形動物(りんけいどうぶつ)は、いわゆるワムシ類と総称される動物の分類群である。
윤형동물(輪形動物, Rotifer) 또는 윤충류(輪蟲類)는 동물분류학상의 한 문(門)이며, 편충동물상문에 속하는 무척추동물이다.
대부분 연못이나 호수에서 자유로운 생활을 하거나 다른 동물에 붙어 산다. 또 부식성 토양이나 이끼 또는 바닷물 속에서도 산다. 몸의 앞끝에 달린 섬모 꼬리를 움직여 섬모를 수레바퀴처럼 운동시키므로 윤충류라는 이름이 붙었다. 윤충류는 여러 동물의 먹이가 되며 생태계에서 중요한 위치를 차지한다. 전 세계에 약 1,500종 정도가 알려져 있다. 암컷은 껍질이 얇은 2배성의 여름 알을 낳고 이 알은 수정되지 않은 채 그대로 발생하여 암컷의 개체가 된다. 이때 알은 물 속이나 수생 식물 위에 붙어 있거나 부화할 때까지 어미의 몸 표면에 붙어 있다. 알은 곧 난할이 진행되어 1-2일 만에 부화하며, 수일 후에는 번식 능력을 갖는 개체로 성숙한다. 그러나 난태생의 종류는 부화할 때까지 알이 자궁 안에 머무르므로 몸 안에서 한 개 내지 여러 개로 발생한 유생을 찾아볼 수 있다. 한편, 단위생식이 오래 계속되거나 주변 환경이 극단적으로 춥거나 덥고, 또 개체수의 밀도가 높아 환경이 나빠지면, 암컷은 장차 수컷이 될 작은 알과 암컷이 될 큰 알을 각각 낳는다. 이리하여 부화한 수컷과 암컷이 교미를 하면, 여름알보다 저항력이 강한 두꺼운 알껍질에 싸여진 수정란, 즉 겨울알(내구알)을 낳는다. 이 알의 껍질에는 가시나 혹 모양의 돌기 등 여러 가지 부속물이 있다. 대부분의 겨울알은 어미가 죽어도, 또한 내용물이 없어진 후에도 계속하여 어미의 몸 안에서 보호된다.
















.JPG)








_-_40X_view.jpg)





















