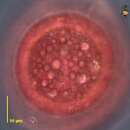
Els Chromatiaceae són la principal família dels bacteris Chromatiales. Viuen generalment en condicions anòxiques sota la llum, però algunes espècies viuen en la foscor sota condicions quimiotròfiques. Es distingeixen pel fet de produir glòbuls de sofre dins les seves cèl·lules. Aquestes són un intermedi en l'oxidació de sulfit, que finalment es converteix en sulfat, i pot servir com a reserva. Els membres d'aquesta família es poden trobar tant en l'aigua dolça com en la salada i són especialment abundant en aigües estancades.
Els Chromatiaceae són la causa que els llacs pobres en oxigen es tornin del color de "sang vermella" com a San Angelo, Texas, USA. http://www.foxnews.com/scitech/2011/08/02/end-times-texas-lake-turns-blood-red
Els Chromatiaceae són la principal família dels bacteris Chromatiales. Viuen generalment en condicions anòxiques sota la llum, però algunes espècies viuen en la foscor sota condicions quimiotròfiques. Es distingeixen pel fet de produir glòbuls de sofre dins les seves cèl·lules. Aquestes són un intermedi en l'oxidació de sulfit, que finalment es converteix en sulfat, i pot servir com a reserva. Els membres d'aquesta família es poden trobar tant en l'aigua dolça com en la salada i són especialment abundant en aigües estancades.
Els Chromatiaceae són la causa que els llacs pobres en oxigen es tornin del color de "sang vermella" com a San Angelo, Texas, USA. http://www.foxnews.com/scitech/2011/08/02/end-times-texas-lake-turns-blood-red
Die Chromatiaceae sind eine Familie von Bakterien innerhalb der Proteobacteria. Wie auch die Ectothiorhodospiraceae gehören sie zu der Ordnung Chromatiales, zusammen bilden sie die physiologische Gruppe der Schwefelpurpurbakterien. Sie betreiben eine anoxische Photosynthese mit der Oxidation von Schwefelwasserstoff zu Schwefel oder Sulfat. Dabei lagern sie den elementaren Schwefel, der als Endprodukt oder als Zwischenprodukt der Schwefelwasserstoffoxidation entsteht, in Form von Kügelchen oder Körnchen innerhalb der Zelle ab. Die meist anaeroben Mitglieder dieser Familie findet man hauptsächlich in Gewässern, sowohl in Süß- als auch in Salzwasser. Früher wurden viele Arten dieser Familie zu den ehemaligen Thiorhodaceae gestellt.
Die Zellen der meisten Arten sind unbeweglich und kugelförmig, auch Stäbchen (z. B. Thiobaca) und Spirillen (z. B. Thiorhodovibrio) kommen vor. Einige Arten bilden Gasvesikel, z. B. Lamprocystis roseopersicina, L. roseopersicina, Thiocapsa rosea und Thiolamprovum pedioforme. Die Gasvesikel dienen dazu die optimale Wasserschichten zu erreichen, Geißeln sind in diesem Fall nicht nötig. Die meisten begeißelten Arten besitzen somit auch keine Gasvesikel, eine der Ausnahmen ist Lamprocystis roseopersicina. Zu den durch Geißeln beweglichen Formen zählen u. a. Chromatium, Allochromatium, Thermochromatium und Thiocystis.
Die Schwefelpurpurbakterien verwenden im Allgemeinen Sulfid-Ionen (S2−), bzw. Schwefelwasserstoff (H2S) als Elektronendonor für die Reduktion von CO2. Sie unterscheiden sich damit von Phototrophen, die Wasser (H2O) als Elektronendonor nutzen, wie beispielsweise Cyanobakterien und Pflanzen, und die deshalb als Oxidationsprodukt des Wassers elementaren Sauerstoff (O2) bilden (oxygene Photosynthese). Bei der Photosynthese der Schwefelpurpurbakterien wird dagegen kein Sauerstoff freigesetzt, ihre Photosynthese ist somit anoxygen.
Die Chromatiaceae oxidieren Sulfide, bzw. Schwefelwasserstoff zu elementaren Schwefel und lagern den Schwefel in Form von Kügelchen innerhalb der Zelle (intrazellulär) ab. Der Schwefel kann dann weiter zu Sulfat oxidiert werden. Dieser Verlauf entspricht der Schwefeloxidation der Ectothiorhodospiraceae, nur dass diese den Schwefel extrazellulär ablagern. Eine Art der Ectothiorhodospiraceae, Thiorhodospira sibirica lagert den Schwefel allerdings nicht nur extrazellulär, sondern auch im periplasmatischen Raum der Zelle ab.[4]
Bei autotrophem Wachstum und wenn als einzige Kohlenstoffquelle Kohlenstoffdioxid (CO2) genutzt wird, erfolgt der Aufbau von Zellmaterial unter Assimilation von CO2 mit Hilfe des Calvinzyklus. Chromatiaceae können in der Regel auch einfache organische Verbindungen verwenden, CO2 ist also nicht die einzige Kohlenstoffquelle, wie es bei den Pflanzen der Fall ist. Acetat und Pyruvat sind die am häufigsten genutzten organischen Kohlenstoffquellen. Polysaccharide, Poly-β-hydroxybutyrat und Polyphosphate werden oft als Energie- bzw. Phosphatreserven gebildet und in den Zellen eingelagert. Der in der Zelle abgelagerte elementare Schwefel fungiert als Elektronendonor- und Energiereserve, bei Abwesenheit von Schwefelwasserstoff wird der gespeicherte Schwefel weiter zu Sulfat oxidiert.
Chlorophyll ist meist das Bacteriochlorophyll α. Einige Arten besitzen auch Bacteriochlorophyll β, Beispiele hierfür sind: Thiococcus pfennigii, Thioalkalicoccus sibiricus und Thioflavicoccus mobilis. Die Carotinoiden, die in dieser Familie vorkommen, zählen zu den Gruppen der Spirilloxanthine (Thermochromatium, Thiocapsa und einige Arten von Allochromatium), Rhodopinale (Allochromatium) und Okenone (z. B. Chromatium). Auch Tetrahydrospirilloxanthin kommt vor (Thiococcus pfennigii).
Zusammen mit den schwefelfreien Purpurbakterien, den sogenannten grünen Schwefelbakterien und den grünen Nichtschwefelbakterien (Chloroflexi) gehören die Schwefelpurpurbakterien zu den anoxygen phototrophen Bakterien. Die bekannten phototrophen Cyanobakterien zeichnen sich dagegen durch die Sauerstoffbildung aus: Da Wasser als Elektronendonator dient wird Sauerstoff freigesetzt. Sie sind somit oxygen phototroph.
Den Stoffwechsel betreffend kann man zwischen zwei Linien innerhalb der Chromatiaceae unterscheiden, den flexiblen und den spezialisierten (also unflexiblen) Arten. Physiologisch eher unflexible Arten sind beispielsweise Chromatium okenii, Chromatium weissei, Allochromatium warmingii, Isochromatium buderi, Thiospirillum jenense und Thiococcus pfennigii. Ohne verfügbares Sulfid erfolgt kein Wachstum, Sulfid ist der einzige nutzbare Elektronendonor, Wasserstoff kann nicht als Elektronendonor verwendet werden. Trotz der Fähigkeit auch Acetat und Pyruvat zu nutzen, bleiben sie abhängig von Kohlenstoffdioxid.
Zu den physiologisch vielseitigen Arten zählen unter anderem Thiocystis violacea, Allochromatium vinosum, Thiocapsa roseopersicina und Lamprobacter modestohalophilus. Einige von ihnen sind in der Lage, ohne reduzierte Schwefelverbindungen zu wachsen. Allochromatium vinosum kann zum Beispiel Wasserstoff als Elektronendonor verwenden. Einige können organische Verbindungen als Elektronendonoren nutzen. Auch Thiosulfat und Eisen(II)-Ionen können als Elektronendonoren fungieren.[5]
Die Mehrheit der Chromatiaceae sind auch Stickstofffixierer: Sie reduzieren elementaren Stickstoff (N2) zu Ammoniak und zählen somit zu den diazotrophen Bakterien.
Folgende Gattungen zählen zu der Familie Chromatiaceae[6]:
Die Chromatiaceae sind eine Familie von Bakterien innerhalb der Proteobacteria. Wie auch die Ectothiorhodospiraceae gehören sie zu der Ordnung Chromatiales, zusammen bilden sie die physiologische Gruppe der Schwefelpurpurbakterien. Sie betreiben eine anoxische Photosynthese mit der Oxidation von Schwefelwasserstoff zu Schwefel oder Sulfat. Dabei lagern sie den elementaren Schwefel, der als Endprodukt oder als Zwischenprodukt der Schwefelwasserstoffoxidation entsteht, in Form von Kügelchen oder Körnchen innerhalb der Zelle ab. Die meist anaeroben Mitglieder dieser Familie findet man hauptsächlich in Gewässern, sowohl in Süß- als auch in Salzwasser. Früher wurden viele Arten dieser Familie zu den ehemaligen Thiorhodaceae gestellt.
Ang Chromatiaceae [(Bigkas: Chro.ma.ti.a'ce.ae)(Medieval Latin: Chromatium, uri ng Genus ng pamilya, -aceae, tinutukoy ang pamilya)] ay isang indibidwal na selula na bilog, hugis ovoid, rod, vibrio o spiral shaped, gumagalaw o hindi gumagalaw, mayroon o walang Gas Vacuoles.
Natuklasan ni Bavendamn noong 1924, 125 at inilabas nina Pfenning at Truper noong 1971, 16.
Gumagalaw ito sa pamamagitan ng Polar Flagella sa mataas na uri, ang dulo nito ay nakikita sa ilalim ng isang Light Microscope. Nakikita ang napakabagal na grupo ng selula at maliksing galaw ng isang indibidwal na selula sa isang naglalaman ng Gas Vacuoles at inilarawan ito ni Winogradsky noong 1888 para sa pag-angat ng Amoebobacter bilang artipak sa isang obserbasyong Mikroskopik at walang koneksiyon ito kasama ang galaw ng maliksing bakterya. Naglalamn ng Bacteriochlorophylls a o b ang bawat selula, grupo ng carotenoids 1,3 ta 4 o tetrahydrospirilloxanthin
![]() Ang lathalaing ito na tungkol sa Agham ay isang usbong. Makatutulong ka sa Wikipedia sa nito.
Ang lathalaing ito na tungkol sa Agham ay isang usbong. Makatutulong ka sa Wikipedia sa nito.
The Chromatiaceae are one of the two families of purple sulfur bacteria, together with the Ectothiorhodospiraceae. They belong to the order Chromatiales of the class Gammaproteobacteria, which is composed by unicellular Gram-negative organisms.[2][3] Most of the species are photolithoautotrophs and conduct an anoxygenic photosynthesis, but there are also representatives capable of growing under dark and/or microaerobic conditions as either chemolithoautotrophs or chemoorganoheterotrophs.[3][4]
Both Ectothiorhodospiraceae and Chromatiaceae bacteria produce elemental sulfur globules, the difference is that in the second case they are stored inside the cells rather than outside. Sulfur is an intermediate in the oxidization of sulfide, which is ultimately converted into sulfate, and may serve as a reserve.[2]
Although purple sulfur bacteria have been known for some time, the difficulty in cultivating these microorganisms in the laboratory has made that few scientific depositions are available to date, and even less are those that provide comparative studies between the two families of the order: Chromatiaceae and Ectothiorhodospiraceae. This is evidenced by the fact that most of the publications at disposal of the scientific community are the result of the work of a relatively small number of scientists, first of all Norbert Pfennig, Johannes Imhoff and Jörg Overmann.[5]
The taxonomy of these two families was originally drawn up entirely on characteristics easy to observe, such as the storage of elemental sulfur inside or outside the cells, the presence of gas vesicles and the internal membrane systems.[3] This led to a first subdivision based on phenotypic properties without taking into account the phylogenetic relationships between the two families.[6][7][8]
The first taxonomic system (1907) was based on the Molisch's pigment and the storage of elemental sulfur. According to these traits, Molisch made a first distinction of the purple sulfur bacteria into two groups: the Thiorhodaceae, including all the members that store globules of elemental sulfur inside their cells, and the Athiorhodaceae, without this feature.[9] These two groups appeared as families of the order Pseudomonadales in the Bergey's Manual of Determinative Bacteriology, 7th ed. In the next edition, they were split in the Rhodospirillaceae (Pfennig and Triiper, 1971) and the Chromatiaceae (Bavendamm, 1924), respectively.[2][10]
Then, the term Chromatiaceae was used for the first time by Bavendamm in 1924, in particular as referred to all those purple bacteria which follow the pathway of anoxygenic photosynthesis, that is to say without using water as electron donor (and consequently without oxygen production), but rather oxidizing sulfide and storing the resulting sulfur either inside or outside the cells. As a consequence of this definition, both the bacteria that accumulate sulfur inside (current Chromatiaceae) and outside the cells (current Ectothiorhodospiraceae) were initially grouped together.[5][7][11]
With the development of chemotaxonomy and DNA sequencing, thanks to analyses on 16S rRNA and pufLM genes as well as on some peculiar phenotypic features (such as lipid and fatty acid composition, quinone structure and amino acid sequence of cytochrome c551), it was possible to demonstrate the phylogenetic distance between the two families. This emerging data led Imhoff (1984) to redefine them: he assigned the genus Ectothiorhodospira (Pelsh, 1936) to a new family called Ectothiorhodospiraceae, and the genus Chromatium (Perty, 1852) to the family Chromatiaceae.[3][5][9][12][13][14]
Referring to the latest definition, Chromatiaceae family includes only those purple sulfur bacteria that perform anoxygenic photosynthesis and store elemental sulfur inside their cells. This is in line with the first Molisch definition of the Thiorhodaceae group and reflects the distance, but phylogenetic correlation between the two families.[11] Therefore, the current taxonomic system for Chromatiaceae corresponds primarily to the phylogenetic knowledge,[5] but it also takes into account phenotypic characteristics and ecological features, which allow to broadly distinguish the members of this family to the Ectothiorhodospiraceae ones. For example, the pH and the response to salinity play an important role in terms of ecological distribution: the species belonging to the family Ectothiorodospiraceae prefer basic pH and habitats with high salt concentration, while the optimal conditions for the Chromatiaceae species include a pH close to neutral and a higher range of salinity, enabling them to occupy more varied habitats, from fresh to brackish and marine water.[11]
The Chromatiaceae bacteria have been divided into four major phylogenetic branches set on the basis of all the aforementioned information:[3][15]

There is a great variety among Chromatiaceae species. From a physiological and morphological point of view, individual members can be distinguished from each other on the basis of cell size and shape (sphere, rod, vibrio, spirillum), motility (polar flagella-dependent motility or nonmotile members), presence of gas vesicles and ability to form cellular aggregates.[3]
Unlike Ectothiorhodospiraceae and Chlorobiaceae bacteria, which store elemental sulfur outside their cells, the intracellularly formed elemental sulfur by Chromatiaceae bacteria gives them great advantages. The chemical element, as it is inaccessible to other bacteria, could serve as a useful personal reserve:[3][16]
Regarding their photosynthetic pigment composition, in the Chromatiaceae can be found bacteriochlorophyll a or b and different types of carotenoids, according to the species. They confer to these bacteria their characteristic pigmentation, and if the amount of cells in the medium is high enough, the coloring may also be visible to the naked eye. Most of them have bacteriochlorophyll a, with a maximum absorption wavelength of 800-900 nm, and a number of species uniquely biosynthesize the red-colored aromatic carotenoid okenone. Okenone has an absorption peak around 500 nm and is involved in both photosynthesis and photoprotection. Bacteriochlorophylls and carotenoids are part of the light-harvesting complex; together with the reaction center, this one makes up the photosynthetic apparatus of Chromatiaceae, which is localized within an intracytoplasmic membrane system arranged as vesicles or tubules, depending on the species. Those able to synthesize okenone are advantaged in deeper water layers, since water absorbs the longer wavelengths in the upper 10 meters thus limiting the use of bacteriochlorophylls for light harvesting.[3][5]
Out of a metabolic point of view, Chromatiaceae bacteria are divided into two main groups: specialized and versatile species. The specialized species are obligate anaerobic photolithoautotrophs, they use reduced sulfur compounds as electron donors and photoassimilate exclusively acetate and pyruvate (or propionate). This group does not include chemotrophs. The versatile species photoassimilate a wider range of organic substrates and most of them do not require the presence of reduced sulfur compounds for their growth, since they are also capable of alternative metabolic strategies.[3][17]
Various metabolic activities are observed in Chromatiaceae:
Chromatiaceae bacteria carry out anoxygenic photosynthesis, during which they oxidize reduced sulfur compounds to sulfate (SO42-). All the species are able to use elemental sulfur (S0) and hydrogen sulfide (H2S) as electron donors. As early as 1931, the stoichiometric link between photosynthetic CO2 fixation and sulfide oxidation was demonstrated: the overall reaction contains the reduction of 2 carbon dioxide molecules to 2 carbohydrates molecules coupled with the oxidation of 1 sulfide molecule to 1 sulfate molecule.[3][18]
2 CO 2 + H 2 S + 2 H 2 O ⟶ 2 CH 2 O + H 2 SO 4 {displaystyle {ce {2CO2 + H2S + 2H2O -> 2CH2O + H2SO4}}} 
Many species utilize thiosulfate (S2O32-) as electron donor, whereas only few of them oxidize sulfite (SO32-) and tetrathionate (S4O62-). In the following example reaction, thiosulfate appears as sodium thiosulfate (Na2S2O3) and is oxidized to sulfuric acid (H2SO4) and sodium sulfate (Na2SO4).[3][19]
2 CO 2 + Na 2 S 2 O 3 + 3 H 2 O ⟶ 2 CH 2 O + H 2 SO 4 + Na 2 SO 4 {displaystyle {ce {2CO2 + Na2S2O3 + 3H2O -> 2CH2O + H2SO4 + Na2SO4}}} 
The final goal of those enzymatic reactions is the oxidation to sulfate. Meanwhile, metabolic intermediates can be formed as sulfur and sulfite. Sulfur is stored as granules inside the cells and can be used as an electron reserve in dark growth conditions during respiration and fermentation. Conversely, sulfite does not accumulate intracellularly since it is readily oxidized to sulfate.[3][20][21]
Most of the species can perform biological nitrogen fixation, during which atmospheric nitrogen or dinitrogen (N2) is converted into ammonia by a nitrogenase.
N 2 + 16 ATP + 16 H 2 O + 8 e − + 8 H + ⟶ 2 NH 3 + H 2 + 16 ADP + 16 Pi {displaystyle {ce {N2 + 16ATP + 16H2O + 8e- + 8H+ -> 2NH3 + H2 + 16ADP + 16Pi}}} 
The favorite nitrogen source of all Chromatiaceae is ammonia; as in many other bacteria, its assimilation is catalyzed by the enzymes GS (glutamine synthetase) and GOGAT (glutamine oxoglutarate aminotransferase), which incorporate the inorganic ammonia into amino acids. Regarding the organic nitrogen sources used by these bacteria, glutamate and aspartate are among the most common.[3][17][22]
Hydrogen (H2) is a major electron donor for photoautotrophic Chromatiaceae. It can be obtained in different ways, according to the growth conditions:[3][23][24][25]
The majority of Chromatiaceae species utilize CO2 as the sole carbon source for autotrophic growth. They employ the Calvin cycle with the key enzymes ribulose bisphosphate carboxylase (RubisCO) and phosphoribulokinase. Versatile species are able to metabolize a larger number of organic compounds than specialized species, which instead mainly assimilate acetate and pyruvate.[3][26]
As mentioned above, although Chromatiaceae largely practice anoxygenic photosynthesis, there are also versatile species able to switch their metabolism in the presence of oxygen.[27]
As in the case of the other purple sulfur bacteria, Chromatiaceae are mainly found in all those anaerobic habitats where the presence of light and the geochemical or biological production of sulfide can support their growth. These types of habitats occur especially in the anoxic sediments of both fresh and sea water, salt marshes, saline and soda lakes, sulfur springs and in stratified environments such as microbial mats and meromictic lakes.
The natural distribution of Chromatiaceae strongly depends on the selective environmental factors described above (anoxic conditions, availability of light and sulfide) and follows daily and seasonal fluctuations, especially in response to the changes in sunlight intensity and temperature. Sulfide concentration tends to fall during the day and rise during the night, this is due to the fact that oxygen production by algae and Cyanobacteria and sulfide oxidation by phototrophic bacteria are diurnal activities; conversely, sulfide production by sulfate reducers and oxygen utilization by respiratory bacteria occur overnight since they are light-independent reactions.[3]
Light absorption changes also according to the type of habitat: the photic zone in sediments is less extensive compared to that in the water, in fact, while in the latter the penetration depth of light is many meters, sediment particles strongly reduce the amount of light entering, and this is the reason why phototrophic bacteria in sediments often form thinner layers within the uppermost millimeters.[3][28] Sulfate present in the sediments is metabolised to sulfide by sulfate-reducing bacteria and the sulfide thus produced diffuses upwards into the water column. Then, sulfide forms a vertical gradient with an opposite trend to that of the light, which on the contrary decreases going downwards.[4] This is the reason why these bacteria can find their optimal growth conditions in small areas of overlap between the countercurrent gradients of the above-mentioned factors.[29][30]
The bacteria inhabiting meromictic lakes generally grow at a quite fixed depth and in a relatively stable way over time, as in these lakes there is a permanent stratification due to the higher salinity of the bottom water layers; in particular, photosynthetic purple sulfur bacteria are located at the level of the chemocline, where they can benefit from both the sunlight from above and the sulfide produced by the underlying anaerobic bacteria. Since the chemocline is usually relatively deep (from a few centimeters up to several meters, depending on the lake), the blooms of these bacteria tend not to be visible at the water surface. Regarding the holomictic lakes, here occurs a seasonal, thermal stratification maintained by temperature differences (especially during summer), which also provides a stable enough layering for purple sulfur bacteria growth; in this case, they cluster at the level of the anoxic and sulfidic hypolimnion, that is to say the dense, bottom water layer of the lake.[3][29][31]

Ectothiorhodospiraceae and Chlorobiaceae are the only other families of phototrophic bacteria which thrive under similar environmental conditions:[28] despite their dissimilarities from a genetic and evolutionary point of view, both purple and green sulfur bacteria depend on reduced inorganic sulfur compounds for their growth and it allows them to play similar ecological roles.[32] The metabolic flexibility of the versatile species belonging to the Chromatiaceae family provides a better adaptation to the environmental changes, such as those occurring in the intermittently oxygenated areas; anyway, the growth they obtain through photosynthesis is greater than that obtained through any other metabolic strategy, regardless of the fact that using forms of metabolism different from photosynthesis means having to compete with those nonphototrophic bacteria and facultative chemotrophic purple nonsulfur bacteria which share the same habitat.[4][16]
Generally, in the multi-layering of a microbial mat, the green sulfur bacteria are placed immediately underneath layers of phototrophic purple sulfur bacteria (and of algae and Cyanobacteria); this coexistence is strategic for several reasons, just to name a few:[4][30][33]
The diagenetic end product of okenone, okenane, is considered a valid biomarker. When bacteria die, they sink to the seafloor together with some of their pigment molecules, including okenone. Under conditions of preservation, the environment is often anoxic and reducing, leading to the chemical loss of functional groups and the partial saturation of okenone to okenane, which in this form can evade microbial and chemical degradation and therefore is buried in marine sediments. This molecule allows to carry out paleoenvironmental reconstructions, since its discovery in marine sediments implies the presence of purple sulfur bacteria during the time of burial and thus a past euxinic environment (that is to say, anoxic and sulfidic waters).[34][35]
An anaerobic lagoon is a shallow, man-made, covered basin designed to hold and pretreat industrial, urban or other kinds of wastewaters. Here Chromatiaceae (and purple sulfur bacteria in general) are the predominant phototrophic bacteria, since they play a key role in metabolizing and thus lowering the concentration of undesirable compounds present in the influent wastewater stream. This is an example of in situ bioremediation, during which the bacteria inhabiting the lagoon exploit these compounds to support their photoheterotrophic growth. For example, they can use the toxic and bad-smelling hydrogen sulfide as the electron source and the greenhouse gas methane as the carbon source for anoxygenic photosynthesis; in this way, the contaminants are removed from the lagoon thus reducing odour and toxicity as well as atmospheric pollution, respectively.[36][37]
{{cite book}}: CS1 maint: others (link) The Chromatiaceae are one of the two families of purple sulfur bacteria, together with the Ectothiorhodospiraceae. They belong to the order Chromatiales of the class Gammaproteobacteria, which is composed by unicellular Gram-negative organisms. Most of the species are photolithoautotrophs and conduct an anoxygenic photosynthesis, but there are also representatives capable of growing under dark and/or microaerobic conditions as either chemolithoautotrophs or chemoorganoheterotrophs.
Both Ectothiorhodospiraceae and Chromatiaceae bacteria produce elemental sulfur globules, the difference is that in the second case they are stored inside the cells rather than outside. Sulfur is an intermediate in the oxidization of sulfide, which is ultimately converted into sulfate, and may serve as a reserve.
Las Chromatiaceae son la familia principal de las sulfobacterias. Se distinguen por producir glóbulos sulfurosos dentro de sus células. Son un intermediario en la oxidización del sulfito, que será últimamente convertido en sulfato, y puede servir de reserva. Sus miembros se encuentran tanto en agua dulce como salina, y muy especialmente comunes en agua estancada en estanques.
I Chromatiaceae sono dei batteri appartenenti alla famiglia del purple sulfur bacteria. Si distinguono per la produzione di globuli di zolfo fuori dal proprio organismo. Sono intermediari nella conversione di solfiti in solfuro che ne diventa una riserva. Organismi di questa famiglia sono presenti sia in acque dolci che in acque salate.
Chromatiaceae é uma família de bactérias gram-negativas, não formadoras de esporos do filo Proteobacteria. Constitui um dos principais grupos de bactérias púrpuras sulfurosas.
Chromatiaceae é uma família de bactérias gram-negativas, não formadoras de esporos do filo Proteobacteria. Constitui um dos principais grupos de bactérias púrpuras sulfurosas.
Chromatiaceae là một họ trong bộ vi khuẩn lưu huỳnh tía (Chromatiales). Chúng có khả năng tạo ra lưu huỳnh ở dạng hạt bên trong các tế bào của chúng. Lưu huỳnh này được sinh ra từ quá trình ôxy hóa chất trung gian sulfua, cuối cùng tạo ra sulfat. Các loài trong họ này được tìm thấy cả trong môi trường nước ngọt và nước mặn[1], và đặc biệt phổ biến trong các hồ nước tù đọng.
Các khi khuẩn lưu huỳnh tía thường sử dụng các ion sulfua (S2-), hoặc hydrogen sulfua (H2S) để làm chất ôxy hóa (cho điện tử) để khử CO2. Do đó, chúng khác với các phototrophs, sử dụng nước (H2O) là chất khử, như cyanobacteria và thực vật, và tạo ra sản phẩm ôxi hóa từ nước là nguyên tố oxy (O2) (quang hợp). Tuy nhiên, quá trình quang hợp của vi khuẩn lưu huỳnh tía không tạo ra ôxy.
Chromatiaceae xôxi hóa các sulfua, hoặc hydrogen sulfua để tạo ra lưu huỳnh nguyên tố và lượng lưu huỳnh này được chứa ở dạng hạt bên trong tế bào của chúng. Lưu huỳnh sau đó có thể bị ôxi hóa tiếp để tạo ra sulfat. Quá trình này giống với sự ôxi hóa lưu huỳnh của Ectothiorhodospiraceae, ngoại trừ một điểm đó là lưu huỳnh tích tụ bên ngoài tế bào. Loài đặc trưng của Ectothiorhodospiraceae, Thiorhodospira sibirica tạo ra nhiều lưu huỳnh xếp lên nhau, tuy nhiên, lưu huỳnh không chỉ tích tụ bên ngoài tế bào mà nó còn có thể tích tục vào khoảng trống giữa màng ngoài và màng trong của tế bào.[2]
Đối với quá trình phát triển tự dưỡng, nếu nó chỉ dùng nguồn carbon dioxide, thì quá trình hình thành các vật chất của tế bào cần sự đồng hóa CO2 với sự hỗ trợ của chu trình Calvin. Ngoài CO2, Chromatiaceae cũng có thể sử dụng các hợp chất hữu cơ đơn giản như vỏ thực vật như lacetate và pyruvate. Polysaccharide, poly-β-hydroxybutyrate và polyphosphate thường được tạo ra ở dạng năng lượng hoặc photphat dự trữ và chứa trong các tế bào. Lượng lưu huỳnh nguyên tố chứ trong tế bào đóng vai trò là nguồn cung cấp electron và năng lượng dự trữ để sau đó bị ôxi hóa tạo ra sulfat.
Chromatiaceae là một họ trong bộ vi khuẩn lưu huỳnh tía (Chromatiales). Chúng có khả năng tạo ra lưu huỳnh ở dạng hạt bên trong các tế bào của chúng. Lưu huỳnh này được sinh ra từ quá trình ôxy hóa chất trung gian sulfua, cuối cùng tạo ra sulfat. Các loài trong họ này được tìm thấy cả trong môi trường nước ngọt và nước mặn, và đặc biệt phổ biến trong các hồ nước tù đọng.
