The brachiopods are a very important group for paleontologists as they have a rich 600 million year old fossil history. With more than 12,000 fossil species described they were possibly the most abundant animals of the Paleozoic era, and important ancient reef builders. Several historical extinction events, most notably the Permian-Triassic extinction, dramatically reduced their numbers and diversity and today only about 335 living species exist.
There are two highly divergent extant classes of brachiopods. The Inarticulata (with about 45 species) appeared first in the fossil record at the beginning of the Cambrian; these were followed by the Articulata (290+ species). The extant genus Lingula shows minimal change from Ordovician fossils and are considered "living fossils".
(Brusca and Brusca 2003; UCMP Berkeley).
Since modern brachiopods inhabit mostly deep or cold marine environments, where they can live by expending very little energy, they are not commonly encountered although they are not rare in modern oceans (Brusca and Brusca 2003; UCMP Berkeley).
Brachiopod larvae are termed “lobate larvae”, but are considerably different between the Articulata and Inarticulata in terms morphology and timing of settlement. Articulate larvae have no mouth so settle after a planktonic period of several days, where they undergo reorganization from a three-part lobed larva to a sessile adult. The morphology of the Inarticulata larvae is similar to adults. They have a far longer planktonic period, at the end of which they gradually uncurl a pedicule and start secreting calcium carbonate or calcium phosphate (Lingulata) to form a shell, which, because it is heavy causes them to settle without such a dramatic metamorphosis as in the Articulata (Brusca and Brusca 2003).
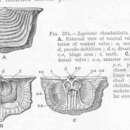
These exclusively benthic marine organisms superficially resemble bivalve molluscs in having two hinged shells (valves), yet they have vastly different body plans. While bivalves have two equally sized valves that are oriented laterally (left and right) with respect to the animal’s body, brachiopod shells are oriented up and down as a dorsal valve and (usually) larger ventral valve. Clearly, shell production has converged from separate origins between these two phyla. Another anatomical peculiarity that obviously distinguishes the brachiopods from the bivalves is the presence of a feeding structure called a lophophore. Only two other phyla have lophophores, the ectoprocts (bryozoans) and phoronids. (These three phyla have long been considered a monophyletic group, but recent evidence suggests the “Lophophorates” is paraphyletic; Halanych, 2004.) In brachiopods the lophophore is made up of two arms of ciliated tentacles held out in a rigid ring circling the mouth. Depending on the species, the lophophore may be a simple circle or highly coiled structure, and in most species it has an internal skeletal support (the Brachidium). The lophophore sits inside the water-filled mantle cavity, taking up about two thirds of the space inside the valves and the brachiopod body takes up the posterior third.
Most brachiopods attach themselves to substrates on the ocean floor or bury into soft substrates using a muscular stalk (called a pedicle, but this structure is not homologous between the two brachiopod classes; Kozloff 1990) although some do not attach to anything and remain free-living. To feed, they crack their shells open using didactor muscles (class Articulata, which have valves that articulate with tooth and groove hinges) or hydrostatic pressure (class Inarticulata, which has unhinged valves and no didactor muscles). The cilia on the lophophore tentacles create a feeding current very specific to the shape of the lophophore, which moves particulates along the tentacles. Food particles are then transported to the mouth via branchial food grooves. In general, lophophorates have a blind, U-shaped gut. The Inarticulata has retained an anus and expel solid waste outside of the mantle cavity, however, the larger order Articulata has lost this feature, so secretes fecal pellets back to the mantle cavity though the mouth. Brachiopods can also clear the mantle cavity and lophophore of lodged non-food contaminants and waste products by contracting the adductor muscles sharply to snap the valves closed and create out-flowing currents.
Brachiopods have a contractile heart, but their open circulatory system appears to function primarily in distributing nutrients. Oxygen diffuses mainly across the lophophore and mantle, and is dispersed in the coelomic fluid. Metabolic wastes are discharged from the body into the mantle cavity though metanephridioducts, which are also used to spawn gametes. Brachiopods have a rudimentary nervous system, and a range of sensory organs dependent on where and how they live.
(Brusca and Brusca 2003; UMCP Berkeley)
Much literature discusses brachiopods as generally avoided as food by fish, sea stars and crabs, and only rare boring predation possibly because there is not much edible tissue in a brachiopod (Thayer, 1985). But other literature disputes this (Harper & Wharton 2000), indicating that predation pressure may indeed have evolutionary significance on brachiopods.
Brachiopods have separate sexes. In most cases fertilization is external in brachiopods, except in a few genera of Articulata where eggs fertilized by sperm in the mangle cavity are brooded in the metanephridia, then released as swimming larvae.
(Brusca and Brusca 2003; Kozloff 1990)
While fossil species got up to 20 cm across, modern brachiopods range from 1 mm to 10 cm.
The phylogenetic position of the brachiopods has been controversial. Many traditional classifications have considered brachiopods (and other lophophorates) to be basal deuterostomes, based on several classically deuderostomic characters: initial cell division of the egg (cleavage) is radial (the cells are arranged in rows, as opposed to spiral cleavage); enterocoelic development leads to a three part body plan; and the mouth is not derived from the blastopore. However, some brachiopods and other lophophorates show exceptions to these characters and other analyses have allied the lophophorates with the protostomes. Molecular phylogenetic work with many independent genes provides consistently strong support for the formal reclassification of the brachiopods as the sister group to pheronids in a group called the “Lophotrochozoa,” essentially derived protostomes (along with molluscs and annelids), and interprets the classical deuderostome characters as more labile and less informative than previously considered. Molecular reconstructions have not allowed much understanding of the relationships within the Lophotrochozoa, but there is much support for a sister relationship between the brachiopods and the phoronids.
(Halanych et al. 1995)
Like other lophophorates, brachiopods are solitary, sessile filter feeders.