Phaeocystis ist eine Gattung von Algen, die zur Klasse der Prymnesiophyten (Prymnesiophyceae) und damit zum großen Phylum (Abteilung) der Haptophyten (Haptophyta) gehört.[1] Phaeocystis-Arten sind im marinen Phytoplankton weit verbreitet und können in einem weiten Temperaturbereich („eurythermal“) und Salzgehaltsbereich („euryhalin“) vorkommen.[2] Mitglieder dieser Gattung leben sowohl im offenen Ozean als auch im Meereis.[3]
Phaeocystis-Arten haben einen vielgestaltigen (polymorphen) Lebenszyklus, der von freilebenden Zellen bis zu großen Kolonien reicht.[2] Auf diese Weise sind einige Arten von Phaecocystis in der Lage, schädliche Algenblüten (en. harmful algal blooms, HABs) zu bilden.
Gattung Phaeocystis[5][6][7][8]
Anmerkungen:
Ein Genomvergleich hat gezeigt, dass die RuBisCO-Spacer-Region (im Plastom, d. h. der Plastiden-DNA, zwischen zwei Untereinheiten des Enzyms Ribulose-1,5-bisphosphat-Carboxylase) unter eng verwandten kolonialen Phaeocystis-Arten hoch konserviert ist. Sie ist in P. antarctica, P. pouchetii und zwei warm-temperierten Stämmen von P. globosa identisch, es gibt lediglich eine einzige Basensubstitution in zwei kalt-temperierten Stämmen von P. globosa.[13]
Phaeocystis kann entweder in der Form freilebender Zellen oder als Kolonie existieren. Ein Sonderfall ist die endosymbiotische Lebensweise.
Ein haploid-diploider Lebenszyklus wurde bisher nur bei P. globosa beobachtet, wird aber auch bei anderen Arten wie P. pouchetii und P. antarctica vermutet. In diesem Zyklus ist die sexuelle Fortpflanzung bei der Bildung und dem Absterben der Kolonieblüte dominant. Abgesehen davon wurden zwei unterschiedliche Arten der vegetativen Fortpflanzung beobachtet.[14]
Freilebende Formen von Phaeocystis sind weltweit verbreitet und kommen in einer Vielzahl von marinen Lebensräumen vor, darunter Küstenmeere, offene Ozeane, Polarmeere und im Meereis.[15]
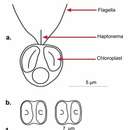
Freilebende Zellen können je nach Art eine Vielzahl von Morphologien aufweisen. Alle Arten können als geschuppte Flagellaten (Geißeltierchen) existieren; bei P. scrobiculata und P. cordata ist dies sogar die einzige beobachtete Form.
Die Bildung schwimmender Kolonien ist eine besondere und kennzeichnende Eigenschaft der Gattung Phaeocystis. In einer solchen Kolonie sind Hunderte von Zellen in eine Matrix aus Polysaccharid-Gel eingebettet, die bei Algenblüten massiv an Größe zunehmen kann.[3]
Bei drei Arten wurden Kolonienbildung beobachtet (P. globosa, P. pouchetii und P. antarctica), diese können auch als ohne Schuppen und Filamente existieren.[14] Bei Kolonien von Phaeocystis kann eine Koloniehaut (en. colony skin) Schutz vor kleineren Zooplanktonfressern und Viren bieten.[16]
Die Allgegenwart und die Bildung großer Algenblüten machen die Gattung Phaeocystis zu einem wichtigen Bestandteil des Kohlenstoffkreislaufs im Ozean.[17][18] Außerdem produziert Phaeocystis Dimethylsulfid (DMS), einen Schlüsselakteur im Schwefelkreislauf.[19][20]
Die Bildung von Algenblüten in nährstoffreichen Gebieten wird durch die drei koloniebildenden Arten (P. globosa, P. pouchetii und P. antarctica) verursacht.[21] Diese können natürliche Ursache haben (z. B. im Rossmeer, in der Grönlandsee oder in der Barentssee)[22] oder durch anthropogene (menschenverursachte) Einträge hervorgerufen werden (z. B. in der Southern Bight in der südlichen Nordsee oder im Persischen Golf).
Im Allgemeinen blüht P. globosa in gemäßigten und tropischen Gewässern, während P. pouchetii und P. antarctica besser an die kalten Temperaturen angepasst sind, die in arktischen bzw. antarktischen Gewässern herrschen. P. pouchetii toleriert jedoch auch wärmere Temperaturen[23] und wurde tatsächlich schon in gemäßigten Gewässern gesehen.[24]
Die größten Phaeocystis-Blüten bilden sich in den Polarmeeren: P. pouchetii im Norden und P. antarctica im Süden.[1] Diese intensive Phaeocystis-Produktivität hält im Allgemeinen etwa drei Monate lang an und überspannt den größten Teil des Sommers auf der Südhalbkugel. Phaeocystis-reiche Ökosysteme stehen in der Regel mit kommerziell wichtigen Beständen von Krebstieren, Mollusken, Fischen und auch Säugetieren in Verbindung. Phaeocystis kann negative Auswirkungen auf höhere trophische Ebenen im marinen Ökosystem und damit auch auf menschliche Aktivitäten (z. B. Fischzucht und Küstentourismus) haben, indem sie während des Abklingens einer Blüte geruchsintensive Schäume an den Stränden bildet.[25]
Phaeocystis-Arten treten als Endosymbionten von Acantharien (Acantharea, informelle Gruppe der Strahlentierchen alias Radiolarien, wohl zu unterscheiden von der Pilzgattung Acantharia) auf.[26][11] Ob diese Symbiose einen echten Mutualismus darstellt, bei dem beide Partner profitieren, ist umstritten.[27] Aus verschiedenen Ozeanbecken gesammelte Acantharien beherbergen unterschiedliche Arten von Phaeocystis als dominante Symbionten:[26]
Zusätzlich zu den beschriebenen Phaeocystis-Arten machen Sequenzen aus einer mit „Phaeo2“ bezeichneten molekularen Klade oft einen Großteil der aus Acanthariern in warmen Wasserregionen gewonnenen symbiotischen Sequenzen aus (neben denen der sog. Klade „Phaeo1“[26][11][28], wohl zu unterscheiden von den gleichnamigen Kladen der Spezies Phaeodactylum tricornutum)[29][30]
Bei symbiotischen Phaeocystis-Arten wird ein extremer Zellumbau beobachtet, einschließlich einer drastischen Zunahme der Chloroplastenzahl und einer vergrößerten zentralen Vakuole.[26][11] Diese phänotypische Veränderung wird wahrscheinlich vom Wirt induziert, um die photosynthetische Leistung der Symbionten zu erhöhen. Die Endosymbiose wird jedoch zur Sackgasse für Phaeocystis, wenn sie die symbiotischen Zellen unfähig für weitere Zellteilungen macht.[27] Die Symbiose ist ökologisch relevant, weil sie Brennpunkte der Primärproduktion in nährstoffarmen Regionen schafft.[31] Es bleibt aber noch zu klären, inwieweit die Symbiose die Evolution von Phaeocystis beeinflusst hat.
Von den Arten P. globosa und P. pouchetii sind eine Reihe von Virusspezies mit Kandidatenstatus bekannt.[32] Diese wurden zunächst vorgeschlagen als Mitglieder der Virusgattung Prymnesiovirus, Familie Phycodnaviridae im Phylum Nucleocytoviricota (NCLDV). Nach neueren Untersuchungen sind einige der Kandidaten jedoch mit Gruppe der sog. „OLPG“ eher der Familie Mimiviridae (oder einer Schwesterfamilie Mesomimiviridae derselben) zuzuordnen.
Anmerkung: In der Prymnesiovirus-Schwestergattung Phaeovirus sind (trotz der Namensähnlichkeit) keine Phaeocystis-infizierende oder -assoziierte Viren bekannt.
Man vermutet, dass die Algenblühen verursachende Mikroorganismen durch sie parasitierende Viren kontrolliert werden und diese sogar in der Lage sind, eine Blüte zusammenbrechen zu lassen.
Die Gattung Phaeocystis ist ein wichtiger Produzent von 3-Dimethylsulphoniopropionat (DMSP). DMSP wird von weiteren marinen Mikroben in flüchtige Schwefelverbindungen zersetzt, insbesondere Methanthiol (CH3SH) und Dimethylsulfid (DMS, (CH3)2S).[1] Ein Großteil des DMS im Meerwasser wird zu Dimethylsulfoxid (DMSO, (CH3)2S=O) oxidiert.[45] Biogenes DMS trägt jährlich etwa 1.5×1010 kg Schwefel zur Atmosphäre bei und spielt daher eine wichtige Rolle im globalen Schwefelkreislauf, der die Wolkenbildung und möglicherweise die Klimaregulierung beeinflussen kann.[1]
Phaeocystis ist eine Gattung von Algen, die zur Klasse der Prymnesiophyten (Prymnesiophyceae) und damit zum großen Phylum (Abteilung) der Haptophyten (Haptophyta) gehört. Phaeocystis-Arten sind im marinen Phytoplankton weit verbreitet und können in einem weiten Temperaturbereich („eurythermal“) und Salzgehaltsbereich („euryhalin“) vorkommen. Mitglieder dieser Gattung leben sowohl im offenen Ozean als auch im Meereis.
Phaeocystis-Arten haben einen vielgestaltigen (polymorphen) Lebenszyklus, der von freilebenden Zellen bis zu großen Kolonien reicht. Auf diese Weise sind einige Arten von Phaecocystis in der Lage, schädliche Algenblüten (en. harmful algal blooms, HABs) zu bilden.
Phaeocystis is a genus of algae belonging to the Prymnesiophyte class and to the larger division of Haptophyta.[1] It is a widespread marine phytoplankton and can function at a wide range of temperatures (eurythermal) and salinities (euryhaline).[2] Members of this genus live in the open ocean, as well as in sea ice.[3] It has a polymorphic life cycle, ranging from free-living cells to large colonies.[2]
The ability to form a floating colony is one of the unique attributes of Phaeocystis – hundreds of cells are embedded in a polysaccharide gel matrix, which can increase massively in size during blooms.[3] The largest Phaeocystis blooms form in the polar seas: P. pouchetii in the north and P. antarctica in the south.[1] This intense Phaeocystis productivity generally persists for about a three-month period, spanning most of the summer in the Southern Hemisphere. Phaeocystis-abundant ecosystems are generally associated with commercially important stocks of crustaceans, molluscs, fish and mammals. Phaeocystis may have negative effects on higher trophic levels in the marine ecosystem, and consequent impacts on human activities (such as fish farming and coastal tourism), by forming odorous foams on beaches during the wane of a bloom.[4]
The ability to form large blooms and its ubiquity make Phaeocystis an important contributor to the ocean carbon cycle.[5][6] In addition, Phaeocystis produces dimethyl sulfide (DMS), a key player in the sulfur cycle.[7][8]


Free-living forms of Phaeocystis are globally distributed and occur in a variety of marine habitats, including coastal oceans, open oceans, polar seas and sea ice.[10] Seven species are currently assigned to the genus: P. antarctica, P. jahnii, P. globosa, P. pouchetti, P. scrobiculata (not in culture), P. cordata, and P. rex.[11] Three species (P. globosa, P. pouchetii, and P. antarctica) are associated with bloom formation in nutrient-rich areas,[12] which can occur either naturally (e.g. in the Ross Sea, Greenland Sea or the Barents Sea) or due to anthropogenic inputs (e.g. in the Southern Bight of the North Sea or the Persian Gulf). Generally, P. globosa blooms in temperate and tropical waters, whereas P. pouchetii and P. antarctica are better adjusted to the cold temperatures prevailing in Arctic and Antarctic waters, respectively. However, P. pouchetii also tolerates warmer temperatures[13] and has been seen in temperate waters.[14]
Genome comparison has shown that the RUBISCO spacer region (located in the plastid DNA, between two subunits of the enzyme 1,5 -bisphosphate carboxylase) is highly conserved among closely related colonial Phaeocystis species and identical in P. antarctica, P. pouchetii and two warm-temperate strains of P. globosa, with a single base substitution in two cold-temperate strains of P. globosa.[15]
Phaeocystis can exist as either free-living cells or colonies. Free-living cells can show a variety of morphologies, depending on the species. All species can exist as scaled flagellates, and this is the only form that has been observed for P. scrobiculata and P. cordata. Three species have been observed as colonies (P. globosa, P. pouchetii and P. antarctica) and these can also exist as a flagellate devoid of scales and filaments.[16] In colonies of Phaeocystis, the colony skin may provide protection against smaller zooplankton grazers and viruses.[17]
While suspected in other species (P. pouchetii and P. antarctica), a haploid-diploid life cycle has only been observed in P. globosa. In this cycle, sexual reproduction is dominant in colony bloom formation/termination, and two types of vegetative reproduction exist.[16]
The genus Phaeocystis is a major producer of 3-dimethylsulphoniopropionate (DMSP), the precursor of dimethyl sulfide (DMS). Biogenic DMS contributes approximately 1.5×1013 g sulfur to the atmosphere annually and plays a major part in the global sulfur cycle, which can affect cloud formation and, potentially, climate regulation.[1]
Phaeocystis species are endosymbionts to acantharian radiolarians.[18][19] Acantharians collected in different ocean basins host different species of Phaeocystis has their dominant symbionts: P. antarctica is found as the primary symbiont to acantharians in the Southern Ocean and P. cordata and P. jahnii are among the dominant symbionts found in acantharians collected in warm oligotrophic regions of the Indian and Pacific oceans.[18] In addition to the described Phaeocystis species, sequences belonging to the molecular clade Phaeo02 often make up a majority of symbiotic sequences recovered from acantharians in warm-water regions.[18][19] Whether or not this symbiosis represents a true mutualism with both partners benefiting, is debated. [20] Extreme cellular remodeling is observed in symbiotic Phaeocystis, including a drastic increase in chloroplast number and an enlarged central vacuole.[18] [19] This phenotypic change is probably induced by the host to increase photosynthetic output by symbionts, but if it renders symbiotic cells incapable of future cell-division, the symbiosis is a dead end for Phaeocystis.[20] The symbiosis is ecologically relevant because it creates primary production hot spots in low-nutrient regions,[21] but it remains to be determined how the symbiosis has affected Phaeocystis evolution.
Phaeocystis is a genus of algae belonging to the Prymnesiophyte class and to the larger division of Haptophyta. It is a widespread marine phytoplankton and can function at a wide range of temperatures (eurythermal) and salinities (euryhaline). Members of this genus live in the open ocean, as well as in sea ice. It has a polymorphic life cycle, ranging from free-living cells to large colonies.
The ability to form a floating colony is one of the unique attributes of Phaeocystis – hundreds of cells are embedded in a polysaccharide gel matrix, which can increase massively in size during blooms. The largest Phaeocystis blooms form in the polar seas: P. pouchetii in the north and P. antarctica in the south. This intense Phaeocystis productivity generally persists for about a three-month period, spanning most of the summer in the Southern Hemisphere. Phaeocystis-abundant ecosystems are generally associated with commercially important stocks of crustaceans, molluscs, fish and mammals. Phaeocystis may have negative effects on higher trophic levels in the marine ecosystem, and consequent impacts on human activities (such as fish farming and coastal tourism), by forming odorous foams on beaches during the wane of a bloom.
The ability to form large blooms and its ubiquity make Phaeocystis an important contributor to the ocean carbon cycle. In addition, Phaeocystis produces dimethyl sulfide (DMS), a key player in the sulfur cycle.
Phaeocystales es un orden de protistas del subfilo Haptophyta[1][2] que comprende únicamente al género Phaeocystis.[3] Son algas unicelulares que presentan una alternancia de generaciones entre etapas móviles e inmóviles. La etapa móvil tiene un tamaño de 3-10 µm, presenta dos flagelos y un apéndice corto denominado haptonema. La etapa inmóvil es colonial, está embebida en una matriz gelatinosa y puede alcanzar un tamaño de varios mm y ser visible a simple vista. La superficie celular está cubierta por placas orgánicas de dos tamaños diferentes y presentan de uno a cuatro cloroplastos.[4] Phaeocystis produce regularmente floraciones en las que el material gelatinoso a menudo se acumula en las playas en forma de espuma.[5]
Phaeocystales es un orden de protistas del subfilo Haptophyta que comprende únicamente al género Phaeocystis. Son algas unicelulares que presentan una alternancia de generaciones entre etapas móviles e inmóviles. La etapa móvil tiene un tamaño de 3-10 µm, presenta dos flagelos y un apéndice corto denominado haptonema. La etapa inmóvil es colonial, está embebida en una matriz gelatinosa y puede alcanzar un tamaño de varios mm y ser visible a simple vista. La superficie celular está cubierta por placas orgánicas de dos tamaños diferentes y presentan de uno a cuatro cloroplastos. Phaeocystis produce regularmente floraciones en las que el material gelatinoso a menudo se acumula en las playas en forma de espuma.
Phaeocystis on yhteyttävistä ja yksisoluisista tarttumalevistä muodostuva suku. Sukuun kuuluvia leviä esiintyy maailman kaikilla merialueilla, myös merijäällä. Ne muodostavat kelluvia lauttoja, joiden koko voi kasvaa räjähdysmäisesti kukinnan aikana. Lautat muodostuvat sadoista soluista, jotka elävät geelimäisistä polysakkarideista muodostuvalla kasvualustalla.[2]
Pohjanmeren eteläosissa esiintyy joinakin kesinä massiivisia Phaeocystis-kukintoja, joiden hajotessa rannoille ajautuu näkö- ja hajuhaittoja aiheuttavaa paksua vaahtoa. Levien hajoamistuotteet saattavat edistää myös happosateiden syntymistä.[1]
Sukuun luetaan seuraavat lajit:[3]
Phaeocystis on yhteyttävistä ja yksisoluisista tarttumalevistä muodostuva suku. Sukuun kuuluvia leviä esiintyy maailman kaikilla merialueilla, myös merijäällä. Ne muodostavat kelluvia lauttoja, joiden koko voi kasvaa räjähdysmäisesti kukinnan aikana. Lautat muodostuvat sadoista soluista, jotka elävät geelimäisistä polysakkarideista muodostuvalla kasvualustalla.
Pohjanmeren eteläosissa esiintyy joinakin kesinä massiivisia Phaeocystis-kukintoja, joiden hajotessa rannoille ajautuu näkö- ja hajuhaittoja aiheuttavaa paksua vaahtoa. Levien hajoamistuotteet saattavat edistää myös happosateiden syntymistä.
Phaeocystis est un genre d'algues nanoplanctoniques marines de la classe des Prymnésiophycées.
Ces algues unicellulaires photosynthétiques sont des eucaryotes présents dans les océans ouverts du monde, également trouvés sous et dans la banquise (Phaeocystis antarctica[1]).
Quand elles passent d'un mode de vie solitaire à une vie coloniale, elles peuvent être à l'origine de blooms planctoniques très importants qui se manifestent notamment en mer du Nord[2], en Manche/mer du Nord, particulièrement dans le pas de Calais et en Belgique[3], aux Pays-Bas [4],[5], mais également périodiquement jusqu'en zone paléarctique[1] ou en Australie comme à Mooloolaba le lundi 28 janvier 2013[6].
Ces blooms se traduisent par l'apparition de colonies flottantes formant un épais mucilage[7] dans l'eau [8]. Ce mucilage est constitué de micro-structures faites de centaines de cellules noyées dans une matrice de gel de polysaccharide[1]. Il peut gêner la pêche et forme parfois sur les plages et les laisses de mer une épaisse couche d'écume (de couleur blanc-gris à brunâtre ou blanc jaunâtre, de quelques millimètres à quelques dizaines de cm, voire à un mètre de hauteur)[9].
Les Phaeocystis jouent un rôle majeur dans les cycles mondiaux du carbone et de soufre et dans les cycles biogéochimiques planétaires en séquestrant efficacement de grandes quantités de CO2 sur de grandes zones géographiques[1]. Lors des proliférations elles sont aussi un gros émetteur de 3-diméthylsulphoniopropionate (DMSP), le précurseur du sulfure de diméthyle (DMS), émis biogéniquement dans l'atmosphère à raison d'environ 1,5 × 1013 grammes de soufre par an, ce qui lui donne un rôle majeur dans le cycle global du soufre, lequel permet la nucléation des gouttes d'eau et la formation d'une grande partie des nuages qui contribuent à la régulation climatique et aux pluies. Le cycle de vie et le régime de croissance des algues de ce genre sont encore mal compris[1].
Selon l'étude faite par l'IFREMER en 2004, bien que les pêcheurs s'y soient habitués et le considèrent comme d'origine naturelle, il s'agit d'un phénomène plus important qu'autrefois, mais qui a déjà été observé en Angleterre en 1923 sur l'estuaire de la Tamise. Il ne semble pas y avoir eu d'observations antérieures relatées par les naturalistes ou chroniqueurs des époques précédentes.
Les pêcheurs de la Manche occidentale française nomment ces blooms « vert de mai », « crasse » (synonyme d'écume) ou parlent du « gluant » ou du « limon ».
Côté anglais les pêcheurs parlent de tobacco juice ou de baccy juice ou de fisherman's signs ou encore de foul water ou de stinking water.
Ce genre regroupe des espèces d'abord décrites par le biologiste Kornmann en 1955, caractérisées par un cycle de vie polymorphique et complexe.
Il est observé dans le milieu naturel sous diverses formes[10] dont :
Le public connaît surtout ces espèces par l'écume de couleur blanc-crème couvrant l'eau ou déposée sur les plages et rochers du littoral par la marée descendante. Cette écume peut devenir nauséabonde quand elle est épaisse (elle a pu atteindre voire dépasser 2 mètres là où le vent et le courant l'accumule). Elle est formée à partir du mucus algal lors des blooms planctoniques qui sont modifiés par une mer très agitée. Ces blooms sont de plus en plus communs de mars à juin sur les littoraux eutrophisés (notamment en Manche-Est/mer du Nord). Dans ces cas, l'espèce en cause est généralement Phaeocystis pouchetii.
C'est le seul phytoplancton marin connu pour être capable de brutalement devenir l'espèce dominante de tout un écosystème[13],[14].
Ces algues sont bien connues pour leur efflorescence algale spectaculaire (dite aussi le "bloom") à la surface de la mer au printemps, favorisée par un excès de nitrates et/ou phosphates dans l'eau[15].
Elle peut être responsable de changements brutaux et importants dans la structure[16] et le fonctionnement des réseaux trophiques (planctonique et benthique), avec des conséquences en matière de biogéochimie[17].
En Manche orientale, où cette espèce a des caractéristiques invasives, elle est – par ses pullulations printanières – susceptible de poser des problèmes écologiques mais aussi économiques (pour la pêche et la conchyliculture), ce qui a justifié le déclenchement par l'IFREMER et l'ULCO d'une étude spécifique de 2002 à 2006, dans le cadre du PNEC (Programme national d’environnement côtier) intitulé « Déterminisme du bloom de Phaeocystis et ses conséquences sur l’écosystème Manche orientale-Sud Mer du Nord », de la frontière franco-belge au littoral du Pays de Caux.
Phaeocystis est un genre eurytherme et ubiquiste qui a colonisé une grande partie de la planète[18]. Diverses espèces de ce genre sont fréquemment observées (avec des blooms printaniers) sur les côtes de la mer du Nord[19] (Belgique[20], Pays-Bas[21],[22], Allemagne[23]).
Divers auteurs[24] en ont trouvé jusqu'aux mers polaires, en Arctique (mer de Barents, mer du Groenland, mer de Béring) comme au sud dans l'Antarctique(mer de Ross, mer de Weddell). On l'a également trouvé en plein Atlantique et dans le Pacifique, et sur les côtes de Floride et d'Australie, comme dans le golfe Arabo-Persique.
Ce sont des espèces qui sont aussi susceptibles d'être transportées à grande distance dans les ballasts de navires marchands.
En zone froide, les espèces dominantes sont :
En zone tempérée, les espèces dominantes seraient[25] :
Des abondances importantes peuvent être mesurées lors des blooms (plus de 1 000 cellules/litre d'eau de mer, avec un record de plus de 37 106 cellules/litre en baie de Somme début avril, sur le littoral picard et du Nord-Pas-de-Calais [26]. Ifremer [27] a par exemple détecté des Phaeocystis abondants sur le littoral boulonnais et picard en mars et mai 2003, et du mois de mars à celui de juin 2003 dans le Dunkerquois.
Les pullulations printanières semblent multiples, mais surtout dépendre de la quantité et proportion de nutriments :
Le bloom prend généralement fin aussi « brutalement » qu'il est apparu. Ce mécanisme semble avoir plusieurs explications :
Les fortes pullulations gênent la pêche, surtout au filet fixe. Le fileyage est plus gêné que le chalutage, notamment à cause du colmatage des filets fixes et des filets à mailles fines (pour la pêche à la crevette en particulier) ; les filets sont colmatés ou alourdis par les mucilages algaux. Les crépines de prise d'eau de refroidissement des moteurs tendent à se boucher. Les pêcheurs signalent que le phénomène est de plus en plus précoce et long, et que le poisson pêché dans ces filets est anormalement gluant et malodorant et doit être lavé et relavé par les pêcheurs[38], etc.
La question des impacts toxicologiques ou écotoxicologiques de ces blooms reste discutée et étudiée, comme en France dans le cadre du PNEC (programme national environnement côtier).
IFREMER cite dans son rapport[39]
Les colonies à l'origine de blooms produisent de grandes quantités de gaz libérés dans l'eau et l'air, dont le diméthylsulfide (DMS) qui est un acidifiant et pourrait jouer un rôle climatique (nucléation de l'eau de pluie, augmentation de la nébulosité et donc de l'albédo nuageux)[43]. En augmentant les pluies, ce type de phénomène peut se traduire par une augmentation du lessivage des terres et donc d'une augmentation de l'eutrophisation des eaux littorales.
Selon World Register of Marine Species (22 nov. 2011)[44] :
Selon ITIS (22 nov. 2011)[45] :
Un projet en cours vise le séquençage génétique de Phaeocystis antarctica[1].
Phaeocystis est un genre d'algues nanoplanctoniques marines de la classe des Prymnésiophycées.


.jpg)






{kind=link}
{kind=link}
{kind=link}