Poxviridae són una família de virus que poden infectar tant a animals vertebrats com a invertebrats. El prototip de la família poxviridae és el virus vaccinia, el qual s’ha utilitzat amb èxit per erradicar la verola o pigota.
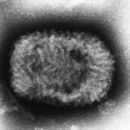
Les partícules virals dels Poxviridae (virions) estan envoltades per substàncies que també són infeccioses. La forma és variable segons les espècies però normalment semblen un maó o un maó arrodonit. El virió és excepcionalment gros i fa uns 200 nm de diàmetre i 300 nm de llargada i porta el seu genoma en doble capa linear d’ADN.[1] Comparat amb el, Rhinovirus aquest només té una mida del 10% d’un virió de Poxviridae .[2]
La replicació d'un poxvirus passa per diferents estadis. El primer que fa aquest virus és unir-se a un receptor en la superfície de la cèl·lula; es creu que el receptor són els Glucosaminoglucans (GAGs). Dins la cèl·lula es replica ràpidament en unes 12 hores.
El nom de la família, Poxviridae, és una herència del virus de la verola o pigota (pox) de la pell. Hi ha quatre gèneres de poxvirus que poden infectar els humans: orthopox, parapox, yatapox i molluscipox.
Inclou els següents gèneres:
Les següents famílies no estan actualment assignades a un gènere:
Poxviridae són una família de virus que poden infectar tant a animals vertebrats com a invertebrats. El prototip de la família poxviridae és el virus vaccinia, el qual s’ha utilitzat amb èxit per erradicar la verola o pigota.
Poxviry (Poxviridae) je čeleď velkých DNA virů o velikosti 230 × 300 nm, jejichž genom se skládá z dvou lineárních řetězců dvouvláknové DNA. Mimo to každý virion obsahuje také enzymatickou výbavu pro syntézu RNA (transkripci) i syntézu DNA.
Mezi poxviry patří virus pravých neštovic (variola virus), který však se již v přírodě pravděpodobně nevyskytuje, dále však také molluscum contagiosum virus (MCV) a některé viry, které normálně člověka nenapadají, ale způsobují choroby zvířat, tzv. zoonózy. Z nemocí ptáků jsou to např. neštovice ptáků. Jako očkování proti pravým neštovicím byl použit rovněž poxvirus, a to virus Vaccinia (jeden z původců kravských neštovic).
Poxviry (Poxviridae) je čeleď velkých DNA virů o velikosti 230 × 300 nm, jejichž genom se skládá z dvou lineárních řetězců dvouvláknové DNA. Mimo to každý virion obsahuje také enzymatickou výbavu pro syntézu RNA (transkripci) i syntézu DNA.
Mezi poxviry patří virus pravých neštovic (variola virus), který však se již v přírodě pravděpodobně nevyskytuje, dále však také molluscum contagiosum virus (MCV) a některé viry, které normálně člověka nenapadají, ale způsobují choroby zvířat, tzv. zoonózy. Z nemocí ptáků jsou to např. neštovice ptáků. Jako očkování proti pravým neštovicím byl použit rovněž poxvirus, a to virus Vaccinia (jeden z původců kravských neštovic).
Die Poxviridae (Pockenviren) sind eine Familie von Viren, die dem Phylum der Nucleocytoviricota (veraltet Nucleocytoplasmic large DNA viruses, NCLDV; frühere Vorschläge hatten auf „Nucleocytoplasmaviricota“ bzw. – im Rang einer Ordnung – „Megavirales“ gelautet) zugerechnet wird.[1] Zu dieser Familie gehören die Unterfamilien Chordopoxvirinae und Entemopoxvirinae.
Vertreter dieser Virenfamilie gehören zu den größten bekannten Viren. Sie können bei ihrer rechteckigen bis ovalen Form eine Größe von 200 bis 400 nm erreichen und sind daher auch in einem sehr guten Lichtmikroskop zu erkennen. Bei allen Mitgliedern dieser Familie handelt es sich um behüllte, doppelsträngige DNA-Viren (dsDNA), denn sie besitzen neben dem Kapsid noch eine weitere Hülle mit einem äußeren und inneren Teil. Dieses Kapsid beinhaltet zusammen mit assoziierten Proteinen den DNA-Doppelstrang in einer S-förmigen Faltung. Die Struktur dieser Viren ist relativ komplex, da sie zusätzlich zu einem bikonkaven Kern zwei Lateralkörper enthalten. Alle zur Familie der Poxviridae gehörenden Viren verfügen über viruskodierte Enzyme, die zur mRNA-Synthese in der Wirtszelle benötigt werden. Weiterhin besitzen sie ein lineares doppelsträngiges DNA-Molekül (DNA-Viren) einer Größe von etwa 130.000 bis 375.000 Basenpaaren. Im Zytoplasma ihres jeweiligen Wirtes können sie sich außerdem leicht vermehren, da sie viele Steuerproteine mitbringen beziehungsweise selbst produzieren.
Die Viren der Unterfamilie Chordopoxvirinae infizieren meist Säugetiere und Vögel, die Entemopoxvirinae aber auch Insekten und verursachen verschiedene Erkrankungen.
In der Geschichte der Wissenschaft besitzen sie durch die Pockenepidemien und die Versuche zur Herstellung von Impfstoffen eine lange Vergangenheit.
Die innere Systematik der Poxviridae ist nach International Committee on Taxonomy of Viruses (ICTV), Stand Master Species List #36, Stand 18. Juni 2021, wie folgt:[2]
Die ICTV Master Species List wurde um einige der wichtigsten Viren (Unterarten/Isolate) zu den jeweiligen Spezies ergänzt. Typusspezies sind fett dargestellt. Die zwar den neuen Vorgaben des ICTV entsprechenden, aber (noch) nicht offiziellen binären Namen (Stand April 2022) finden sich beispielsweise bei Anton Mayr (2007).[17]
Das folgende Kladogramm der inneren Systematik der Poxviridae folgt Clara Rolland et al. (2019):[18]
PoxviridaeEntomopoxvirinae: Alphaentomopoxvirus
Schulz et al. (2018), Fig. 2[19] schlugen eine Systematik der NCLDV (jetzt Nucleocytoviricota) vor, in der die Poxviridae eine basale Gruppe darstellen und die (erweiterten) Asfarviridae im Zweig der Marseilleviridae verortet werden. Das hätte eine gemeinsame Wurzel aller Riesenviren unter den NCLDV bedeutet.
Koonin et al. (2015 und 2019) sowie Bäckström et al. (2019) schlugen hingegen eine Systematik der NCLDV vor, in der die (erweiterte) Familie der Asfarviridae eine Schwestergruppe der Poxviridae bildet. Zusammen bilden sie dann neben einem einen 3. Zweig (englisch branch) der NCLDV, neben einem 1. Zweig mit Mimiviridae und Phycodnaviridae, und einem 2. Zweig mit den Pithoviridae, Marseilleviridae und Iridoviridae.[20][21] Das ICTV ist diesen Vorschlägen mit seiner Master Species List (MSL) #35 im März 2020 gefolgt.
Die Poxviridae (Pockenviren) sind eine Familie von Viren, die dem Phylum der Nucleocytoviricota (veraltet Nucleocytoplasmic large DNA viruses, NCLDV; frühere Vorschläge hatten auf „Nucleocytoplasmaviricota“ bzw. – im Rang einer Ordnung – „Megavirales“ gelautet) zugerechnet wird. Zu dieser Familie gehören die Unterfamilien Chordopoxvirinae und Entemopoxvirinae.
Les pocvirûsses, c' est des virûsses ki dnèt des maladeyes avou des ploketes, ås djins et ås biesses.
C' est des virûsses å SDN (a l' ADN).
noerès plokes
prindåves bocrês ås bedots, passés a ene djin
vaccene
plokes ås vatches passêyes a ene djin
noerès plokes ås bedots
Les pocvirûsses, c' est des virûsses ki dnèt des maladeyes avou des ploketes, ås djins et ås biesses.

Orf-Wiirus
Süstemaatik Hoodkategorii: Wiiren Famile: Poxviridae Taksonomii Genoom: dsDNA linear Baltimore-klas: Skööl 1 Sümetrii: kompleks Wiirusskan: diar Wedenskapelk nööm Poxviridae (ingelsk) FerwisangenPoxviridae san en famile faan wiiren. Jo liase pooken ütj.
Poxviridae is a family of double-stranded DNA viruses. Vertebrates and arthropods serve as natural hosts. There are currently 83 species in this family, divided among 22 genera, which are divided into two subfamilies. Diseases associated with this family include smallpox.[1][2]
Four genera of poxviruses may infect humans: Orthopoxvirus, Parapoxvirus, Yatapoxvirus, Molluscipoxvirus. Orthopoxvirus: smallpox virus (variola), vaccinia virus, cowpox virus, monkeypox virus; Parapoxvirus: orf virus, pseudocowpox, bovine papular stomatitis virus; Yatapoxvirus: tanapox virus, yaba monkey tumor virus; Molluscipoxvirus: molluscum contagiosum virus (MCV).[3] The most common are vaccinia (seen on the Indian subcontinent) and molluscum contagiosum, but monkeypox infections are rising (seen in west and central African rainforest countries). The similarly named disease chickenpox is not a true poxvirus and is caused by the herpesvirus varicella zoster.
The name of the family, Poxviridae, is a legacy of the original grouping of viruses associated with diseases that produced poxes in the skin. Modern viral classification is based on phenotypic characteristics; morphology, nucleic acid type, mode of replication, host organisms, and the type of disease they cause. The smallpox virus remains the most notable member of the family.
Diseases caused by pox viruses, especially smallpox, have been known about for centuries. One of the earliest suspected cases is that of Egyptian pharaoh Ramses V who is thought to have died from smallpox circa 1150 years BCE.[4][5] Smallpox was thought to have been transferred to Europe around the early 8th century and then to the Americas in the early 16th century, resulting in the deaths of 3.2 million Aztecs within two years of introduction. This death toll can be attributed to the indigenous population's complete lack of exposure to the virus over millennia.
A century after Edward Jenner showed that the less potent cowpox could be used to effectively vaccinate against the more deadly smallpox, a worldwide effort to vaccinate everyone against smallpox began with the ultimate goal to rid the world of the plague-like epidemic. The last case of endemic smallpox occurred in Somalia in 1977. Extensive searches over two years detected no further cases, and in 1979 the World Health Organization (WHO) declared the disease officially eradicated.
In 1986, all virus samples were destroyed or transferred to two approved WHO reference labs: at the headquarters of the federal Centers for Disease Control and Prevention (the C.D.C.) in Atlanta, Georgia (the United States) and at the Institute of Virus Preparations in Moscow.[6] After September 11, 2001 the American and UK governments have had increased concern over the use of smallpox, or a smallpox-like disease, in bioterrorism. However, several poxviruses including vaccinia virus, myxoma virus, tanapox virus and raccoon pox virus are currently being investigated for their therapeutic potential in various human cancers in preclinical and clinical studies.[7][8][9]
Poxviridae viral particles (virions) are generally enveloped (external enveloped virion), though the intracellular mature virion form of the virus, which contains different envelope, is also infectious. They vary in their shape depending upon the species but are generally shaped like a brick or as an oval form similar to a rounded brick because they are wrapped by the endoplasmic reticulum. The virion is exceptionally large, its size is around 200 nm in diameter and 300 nm in length and carries its genome in a single, linear, double-stranded segment of DNA.[10] By comparison, rhinoviruses are 1/10 as large as a typical Poxviridae virion.[11]
Phylogenetic analysis of 26 different chordopoxvirus genomes has shown that the central region of the genome is conserved and contains ~90 genes.[12] The termini in contrast are not conserved between species. Of this group Avipoxvirus is the most divergent. The next most divergent is Molluscipoxvirus. Capripoxvirus, Leporipoxvirus, Suipoxvirus and Yatapoxvirus genera cluster together: Capripoxvirus and Suipoxvirus share a common ancestor and are distinct from the genus Orthopoxvirus. Within the Othopoxvirus genus Cowpox virus strain Brighton Red, Ectromelia virus and Monkeypox virus do not group closely with any other member. Variola virus and Camelpox virus form a subgroup. Vaccinia virus is most closely related to CPV-GRI-90.
The GC-content of family member genomes differ considerably.[13] Avipoxvirus, capripoxvirus, cervidpoxvirus, orthopoxvirus, suipoxvirus, yatapoxvirus and one Entomopox genus (Betaentomopoxvirus) along with several other unclassified Entomopoxviruses have a low G+C content while others - Molluscipoxvirus, Orthopoxvirus, Parapoxvirus and some unclassified Chordopoxvirus - have a relatively high G+C content. The reasons for these differences are not known.
Replication of the poxvirus involves several stages.[14] The virus first binds to a receptor on the host cell surface; the receptors for the poxvirus are thought to be glycosaminoglycans. After binding to the receptor, the virus enters the cell where it uncoats. Uncoating of the virus is a two step process. Firstly the outer membrane is removed as the particle enters the cell; secondly the virus particle (without the outer membrane) fuses with the cellular membrane to release the core into the cytoplasm. The pox viral genes are expressed in two phases. The early genes encode the non-structural protein, including proteins necessary for replication of the viral genome, and are expressed before the genome is replicated. The late genes are expressed after the genome has been replicated and encode the structural proteins to make the virus particle. The assembly of the virus particle occurs in five stages of maturation that lead to the final exocytosis of the new enveloped virion. After the genome has been replicated, the immature virion assembles the A5 protein to create the intracellular mature virion. The protein aligns and the brick-shaped envelope of the intracellular enveloped virion. These particles are then fused to the cell plasma to form the cell-associated enveloped virion, which encounters the microtubules and prepares to exit the cell as an extracellular enveloped virion. The assembly of the virus particle occurs in the cytoplasm of the cell and is a complex process that is currently being researched to understand each stage in more depth. Considering the fact that this virus is large and complex, replication is relatively quick taking approximately 12 hours until the host cell dies by the release of viruses.
The replication of poxvirus is unusual for a virus with double-stranded DNA genome because it occurs in the cytoplasm,[15] although this is typical of other large DNA viruses.[16] Poxvirus encodes its own machinery for genome transcription, a DNA dependent RNA polymerase,[17] which makes replication in the cytoplasm possible. Most double-stranded DNA viruses require the host cell's DNA-dependent RNA polymerase to perform transcription. These host polymerases are found in the nucleus, and therefore most double-stranded DNA viruses carry out a part of their infection cycle within the host cell's nucleus.
The ancestor of the poxviruses is not known but structural studies suggest it may have been an adenovirus or a species related to both the poxviruses and the adenoviruses.[18]
Based on the genome organisation and DNA replication mechanism a phylogenetic relationships may exist between the rudiviruses (Rudiviridae) and the large eukaryal DNA viruses: the African swine fever virus (Asfarviridae), Chlorella viruses (Phycodnaviridae) and poxviruses (Poxviridae).[19]
The mutation rate in poxvirus genomes has been estimated to be 0.9–1.2 x 10−6 substitutions per site per year.[20] A second estimate puts this rate at 0.5–7 × 10−6 nucleotide substitutions per site per year.[21] A third estimate places the rate at 4–6 × 10−6.[22]
The last common ancestor of the extant poxviruses that infect vertebrates existed 0.5 million years ago. The genus Avipoxvirus diverged from the ancestor 249 ± 69 thousand years ago. The ancestor of the genus Orthopoxvirus was next to diverge from the other clades at 0.3 million years ago. A second estimate of this divergence time places this event at 166,000 ± 43,000 years ago.[21] The division of the Orthopoxvirus into the extant genera occurred ~14,000 years ago. The genus Leporipoxvirus diverged ~137,000 ± 35,000 years ago. This was followed by the ancestor of the genus Yatapoxvirus. The last common ancestor of the Capripoxvirus and Suipoxvirus diverged 111,000 ± 29,000 years ago.
An isolate from a fish – salmon gill poxvirus – appears to be the earliest branch in the Chordopoxvirinae.[23] A new systematic has been proposed recently after findings of a new squirrel poxvirus in Berlin, Germany.[24]
The date of the appearance of smallpox is not settled. It most likely evolved from a rodent virus between 68,000 and 16,000 years ago.[25][26] The wide range of dates is due to the different records used to calibrate the molecular clock. One clade was the variola major strains (the more clinically severe form of smallpox) which spread from Asia between 400 and 1,600 years ago. A second clade included both alastrim minor (a phenotypically mild smallpox) described from the American continents and isolates from West Africa which diverged from an ancestral strain between 1,400 and 6,300 years before present. This clade further diverged into two subclades at least 800 years ago.
A second estimate has placed the separation of variola from Taterapox at 3000–4000 years ago.[22] This is consistent with archaeological and historical evidence regarding the appearance of smallpox as a human disease which suggests a relatively recent origin. However, if the mutation rate is assumed to be similar to that of the herpesviruses the divergence date between variola from Taterapox has been estimated to be 50,000 years ago.[22] While this is consistent with the other published estimates it suggests that the archaeological and historical evidence is very incomplete. Better estimates of mutation rates in these viruses are needed.
The species in the subfamily Chordopoxvirinae infect vertebrates and those in the subfamily Entomopoxvirinae infect insects. There are ten recognised genera in the Chordopoxvirinae and three in the Entomopoxvirinae.
The following subfamilies and genera are recognized (-virinae denotes subfamily and -virus denotes genus):[2]
Both subfamilies also contain a number of unclassified species for which new genera may be created in the future.
The prototypical poxvirus is vaccinia virus, known for its role in the eradication of smallpox. The vaccinia virus is an effective tool for foreign protein expression, as it elicits a strong host immune-response. The vaccinia virus enters cells primarily by cell fusion, although currently the receptor responsible is unknown.
Vaccinia contains three classes of genes: early, intermediate and late. These genes are transcribed by viral RNA polymerase and associated transcription factors. Vaccinia replicates its genome in the cytoplasm of infected cells, and after late-stage gene expression undergoes virion morphogenesis, which produces intracellular mature virions contained within an envelope membrane. The origin of the envelope membrane is still unknown. The intracellular mature virions are then transported to the Golgi apparatus where it is wrapped with an additional two membranes, becoming the intracellular enveloped virus. This is transported along cytoskeletal microtubules to reach the cell periphery, where it fuses with the plasma membrane to become the cell-associated enveloped virus. This triggers actin tails on cell surfaces or is released as external enveloped virion.
{{cite web}}: CS1 maint: multiple names: authors list (link) Poxviridae is a family of double-stranded DNA viruses. Vertebrates and arthropods serve as natural hosts. There are currently 83 species in this family, divided among 22 genera, which are divided into two subfamilies. Diseases associated with this family include smallpox.
Four genera of poxviruses may infect humans: Orthopoxvirus, Parapoxvirus, Yatapoxvirus, Molluscipoxvirus. Orthopoxvirus: smallpox virus (variola), vaccinia virus, cowpox virus, monkeypox virus; Parapoxvirus: orf virus, pseudocowpox, bovine papular stomatitis virus; Yatapoxvirus: tanapox virus, yaba monkey tumor virus; Molluscipoxvirus: molluscum contagiosum virus (MCV). The most common are vaccinia (seen on the Indian subcontinent) and molluscum contagiosum, but monkeypox infections are rising (seen in west and central African rainforest countries). The similarly named disease chickenpox is not a true poxvirus and is caused by the herpesvirus varicella zoster.
Poxviridae es una familia de virus de ADN relacionados entre sí llamados poxvirus, infectivos para animales. Tienen un genoma ADN bicatenario y por lo tanto pertenece al Grupo I de la Clasificación de Baltimore. El nombre de la familia proviene de un agrupamiento inicial de enfermedades asociados a la piel denominados en conjunto pox en otros idiomas. La clasificación viral más moderna está basada en las características moleculares y la forma de los virus miembros. El más notable de ellos es la viruela y desde su erradicación mundial, solo el virus del molusco contagioso infecta específicamente a los humanos, más que cualquier otro poxvirus.[1]
Son virus gigantes, la cápside está estructuralmente definida por una simetría compleja, de un tamaño de 300-400 nm x 250-290 nm. Todos tienen silueta ovalada característica en forma de ladrillo, con una membrana externa que muestra crestas de función desconocida y corpúsculos laterales que encajan en las concavidades.[2] La mayoría poseen una envoltura viral.
Se caracterizan por poseer un genoma con ADN de cadena doble lineal como ácido nucleico; de 130 - 375 kpb y ensamblar los viriones maduros en el citoplasma como compartimento celular.[3] El ADN así ensamblado posee asas terminales y un bajo contenido GC (30-40%), excepto el Parapoxvirus (en la foto), que tiene alrededor del 60%. El mecanismo de replicación viral se ensambla en el citoplasma.
Los poxvirus están constituidos por un 90% proteínas, 3% ADN y 5% lípidos. Contienen más de 100 polipéptidos, en el núcleo contiene sus propias enzimas incluyendo un sistema de transcripción genética.[4] Las proteínas codificadas por el virus ayudan a evadir el sistema inmune de defensa del huésped.
Poxviridae es una familia de virus de ADN relacionados entre sí llamados poxvirus, infectivos para animales. Tienen un genoma ADN bicatenario y por lo tanto pertenece al Grupo I de la Clasificación de Baltimore. El nombre de la familia proviene de un agrupamiento inicial de enfermedades asociados a la piel denominados en conjunto pox en otros idiomas. La clasificación viral más moderna está basada en las características moleculares y la forma de los virus miembros. El más notable de ellos es la viruela y desde su erradicación mundial, solo el virus del molusco contagioso infecta específicamente a los humanos, más que cualquier otro poxvirus.
Poksviirused ehk rõugeviirused on kaksikahelalised DNA-viirused, mille replikatsioon toimub tsütoplasmas, mistõttu nad saavad oma genoomi iseseisvalt transkribeerida ja replitseerida.
Poksviirused moodustavad sugukonna Poxviridae.
Poxviridés
Les Poxviridés (Poxviridae) est une famille qui appartient au groupe I des virus à ADN à double brin et comprend 2 sous-familles : les Chordopoxvirinae chez les vertébrés et les Entomopoxvirinae chez les insectes.
« Pox » est le pluriel de « pock » qui en anglais signifie « pustule ».
Ce sont de grands virus de forme généralement ovale de 200 nm de diamètre et 300 nm de long. Ils contiennent un core central en forme d'haltère ou nucléoïde qui contient le génome et qui est flanqué de deux corps latéraux.
Les poxvirus possèdent un génome d'ADN linéaire double-brin variant de 134 kbp (134 000 paires de bases) dans les parapoxvirus à plus de 300 kbp dans certains avipoxvirus. En lui-même, l'ADN d'un poxvirus n'est pas infectieux car l'ARN polymérase virale et d'autres enzymes et facteurs viraux sont nécessaires pour exprimer le génome viral dans le cytoplasme. Tous les poxvirus possèdent des répétitions terminales inversées (en anglais, ITR pour inverted terminal repetitions) qui sont des séquences identiques mais d'orientation opposée aux deux extrémités du génome.
La réplication des poxvirus comporte plusieurs étapes. Le virus se lie à un récepteur sur la surface de la cellule hôte, on pense que les récepteurs pour les poxvirus sont les glycosaminoglycanes (GAG). Après liaison au récepteur, le virus pénètre dans la cellule où il est déshabillé. La décapsidation du virus est un processus en deux étapes. Tout d'abord la membrane externe est enlevée alors que la particule pénètre dans la cellule, ensuite la particule virale, sans la membrane externe, se désagrège jusqu'à libérer le core dans le cytoplasme. Les gènes viraux sont exprimés en deux phases. Les gènes précoces sont d'abord exprimés. Ces gènes codent des protéines non structurelles, comme les protéines nécessaires à la réplication du génome viral et sont exprimés avant que le génome ne soit répliqué. Les gènes tardifs sont exprimés après que le génome a été répliqué et codent les protéines structurelles permettant de rendre la particule infectieuse. L'assemblage de la particule virale se produit dans le cytoplasme de la cellule et est un processus complexe qui est mal compris et actuellement à l'étude.
La réplication des poxvirus est inhabituelle pour des virus à ADN double brin (ADNdb) parce qu'elle se produit dans le cytoplasme. Les poxvirus codent leur propre machine pour la transcription du génome, une ARN polymérase dépendante de l'ADN, ce qui rend la réplication dans le cytoplasme possible. La plupart des virus à ADNdb nécessitent des protéines de la cellule hôte pour effectuer la transcription. Ces protéines de l'hôte se trouvent dans le noyau et donc la plupart des virus ADNdb effectuent une partie de leur cycle d'infection au sein du noyau de la cellule hôte.
Poxviridés
Les Poxviridés (Poxviridae) est une famille qui appartient au groupe I des virus à ADN à double brin et comprend 2 sous-familles : les Chordopoxvirinae chez les vertébrés et les Entomopoxvirinae chez les insectes.
« Pox » est le pluriel de « pock » qui en anglais signifie « pustule ».
Ce sont de grands virus de forme généralement ovale de 200 nm de diamètre et 300 nm de long. Ils contiennent un core central en forme d'haltère ou nucléoïde qui contient le génome et qui est flanqué de deux corps latéraux.
Poxviridae (poxvíridos), é unha familia de virus que infectan a vertebrados e invertebrados, cuxos membros se coñecen comunmente como poxvirus. A raíz pox do seu nome procede do inglés, idioma no que inicialmente se aplicou á doenza máis famosa que producen, a varíola, que despois pasou a chamarse smallpox.
Hai catro xéneros de poxvirus que poden infectar a humanos, que son:
Anteriormente o virus da varíola causaba grandes epidemias, pero foi erradicado pola vacinación. Actualmente, os máis comúns son o virus da vaccinia, que se pode atopar no subcontinente indio, e o virus do molusco contaxioso, aínda que están aumentando os casos de infeccións por virus da varíola do mono en zonas tropicais de África central.
As partículas virais (virións) dos Poxviridae teñen xeralmente envoltura (virión con envoltura externa ou EEV), aínda que a forma virión maduro intracelular (IMV) deste virus, contén envolturas diferentes, e é tamén infeccioso. Varían na súa forma dependendo da especie, pero xeralmente teñen forma de ladrillo ou forma oval similar a un ladrillo redondeado porque están envoltos polo retículo endoplasmático da célula. O virión é excepcionalmente grande, o seu tamaño é de arredor de 200 nm de diámetro e 300 nm de lonxitude e leva o seu xenoma nun segmento de ADN de dobre cadea linear e único.[3] En comparación, a lonxitude dun Rhinovirus é só a décima parte da dun virión típico de Poxviridae.[4]
A replicación dos poxvirus implica varios estadios. A primeira cousa que fai o virus é unirse a un receptor da superficie da célula hóspede; crese que os receptores para os pxvirus son glicosaminoglicanos (GAGs). Despois de unirse ao receptor, o virus entra na célula, onde perde a envoltura. Esta perda da envoltura é un proceso en dúas etapas. Primeiramente, elimínase a membrana externa a medida que a partícula entra na célula; en segundo lugar, a partícula vírica (sen a membrana externa) fusiónase coa membrana celular para liberar a súa parte central (core) no citoplasma. Os xenes virais pox exprésanse en dúas fases. Os xenes temperáns codifican as proteínas non estruturais, incluíndo as proteínas necesarias para a replicación do xenoma viral, e exprésanse antes de que se replique o xenoma. Os xenes tardíos exprésanse despois da replicación do xenoma e codifican as proteínas estruturais necesarias para construír a partícula do virus. A ensamblaxe da partícula do virus ocorre en cinco etapas de maduración que acaban coa exocitose final dos virións novos envoltos. O número de membranas que envolven os virións foi discutido, pero crese que son dunha a tres segundo a etapa do ciclo vital.
En virus como o da vaccinia distínguense varias formas do virión, que son:[5]
A replicación dos poxvirus é pouco usual para un virus de ADN bicatenario (que xeralmente son pequenos) porque ocorre no citoplasma,[6] aínda que isto é típico noutros virus de ADN gran tamaño.[7] Os poxvirus codifican a súa propia maquinaria para a transcrición do xenoma, unha ARN polimerase ADN dependente propia,[8] o que fai posible a replicación no citoplasma. A maioría dos virus de ADN bicatenario necesitan usar a ARN polimerase ADN dependente da célula hóspede para realizar a transcrición. Este encima do hóspede encóntrase no núcleo celular, e, por tanto, a maioría dos virus de ADN bicatenario levan a cabo unha parte do seu ciclo de infección no núcleo da célula.
O antepasado dos poxvirus non se coñece pero os estudos estruturais suxiren que este puido ser un adenovirus ou unha especie relacionada tanto cos poxvirus coma cos adenovirus.[9]
Baseándose na organización do xenoma e no mecanismo da replicación do ADN parece que pode haber relacións filoxenéticas entre os rudivirus (Rudiviridae) e os virus de ADN eucariotas grandes: o virus da febre suína africana (Asfarviridae), virus de Chlorella (Phycodnaviridae) e os poxvirus (Poxviridae).[10]
A taxa de mutación neses xenomas estímase que está entre 0,9 e 1,2 x 10−6 substitucións por sitio por ano.[11] Outra estimación considera que está entre 0,5 e 7 × 10−6 substitucións de nucleótidos por sitio e por ano.[12] Unha terceira estimación sitúa a cifra en 4-6 × 10−6.[13]
O último antepasado común dos poxviruses existentes que infectan a vertebrados existiu hai 500 mil anos. O xénero Avipoxvirus diverxiu do seu antepasado hai 249 ± 69 miles de anos. O devanceiro do xénero Orthopoxvirus foi o seguinte en diverxir dos outros clados hai 300 mil anos. Unha segunda estimación do tempo desta diverxencia sitúa este evento hai 166.000 ± 43.000 anos.[12] A división dos Orthopox ocorreu hai ~14.000 anos. O xénero Leporipoxvirus diverxiu hai ~137.000 ± 35.000 anos. Isto foi seguido polo antepasado do xénero Yatapoxvirus. O último antepasado común do Capripoxvirus e Suipoxvirus diverxiu hai 111.000 ± 29.000 anos.
A data en que apareceu a varíola non foi establecida. Probablemente evolucionou a partir dun virus de roedores hai entre 68.000 e 16.000 anos.[14][15] O amplo rango das datacións débese aos diferentes rexistros utilizados para calibrar o reloxo molecular. Un clado foi o das cepas principais da varíola (as clinicamente máis graves) que se espallou por Asia entre hai 400 e 1.600 anos. Un segundo clado incluía o alastrim (unha varíola fenotipicamente leve) descrita nos continentes americanos e illada en África occidental, que diverxiu a partir dunha capa ancestral hai entre 1.400 e 6.300 anos. Este clado despois diverxiu en dous subclados hai polo menos 800 anos.
Unha segunda estimación situou a separación da varíola do Taterapox virus (de roedores) hai 3.000–4.000 anos.[13] Isto é consistente coas probas arqueolóxicas e históricas en relación coa aparición da varíola como doenza humana, o que suxire unha orixe relativamente recente. Porén, se a taxa de mutación se asume que é similar á dos herpesvirus, a datación da diverxencia entre a varíola e o Taterapox virus estimouse que se produciu hai 50.000 anos.[13] Aínda que isto está en concordancia con outras estimacións publicadas, tamén indica que as probas arqueolóxicas e históricas son moi incompletas. Necesítanse mellores estimacións das taxas de mutación nestes virus.
As especies da subfamilia Chordopoxvirinae infectan vertebrados e as da subfamilia Entomopoxvirinae infectan insectos. Hai 10 xéneros recoñecidos entre os Chordopoxvirinae e 3 entre os Entomopoxvirinae. Ambas as subfamilias conteñen ademais varias especies non clasificadas en xéneros para as cales seguramente se crearán novos xéneros no futuro. O virus Cotia é un virus inusual que pode pertencer a un novo xénero.[16]
O contido GC dos xenomas destes virus difire considerablemente.[17] Os xéneros Avipoxvirus, Capripoxvirus, Cervidpoxvirus, Orthopoxvirus, Suipoxvirus, Yatapoxvirus e un xénero de entomopoxvirus, o Betaentomopoxvirus, xunto con varios outros entomopoxvirus non clasificados, teñen un contido GC baixo, mentres que outros, como Molluscipoxvirus, Orthopoxvirus, Parapoxvirus e algúns cordopoxvirus non clasificados, teñen un contido GC relativamente alto. Non se coñecen as razóns destas diferenzas.
A análise filoxenética de 26 xenomas de cordopoxvirus indicou que a rexión central do xenoma está conservda e contén ~90 xenes.[18] As rexións terminais, ao contrario, non están conservadas entre especies. Deste grupo Avipoxvirus é o máis diverxente. O seguinte máis diverxente é Molluscipoxvirus. Os xéneros Capripoxvirus, Leporipoxvirus, Suipoxvirus e Yatapoxvirus poden agruparse: Capripoxvirus e Suipoxvirus comparten un devanceiro común e son diferentes do xénero Orthopoxvirus. Dentro do xénero Othopoxvirus o virus da varíola bovina cepa Brighton Red, o virus Ectromelia e o virus da varíola do mono non están agrupados estreitamente con ningún outro membro. O virus da varíola humana e o virus da varíola do camelo forman un subgrupo. O virus vaccinia é o que está máis relacionado con CPV-GRI-90.
As doenzas causadas polos poxvirus, especialmente a varíola, coñécense desde hai séculos. Un dos primeiros casos sospeitosos é o do faraón exipcio Ramsés V, que se cre que morreu de varíola aproximadamente no ano 1150 a. C..[19][20] A varíola pénsase que foi transferida a Europa arredor de finais do século VIII e despois a América a inicios do século XVI. Acéptase que a gran mortaldade causada entre os aztecas, que contribuíu grandemente á súa derrota, foi causada por unha epidemia de varíola e que en dous anos morreron uns 3,2 millóns de indíxenas mexicanos. Esta elevada cifra pode explicarse pola completa falta de inmunización contra ese virus da poboación americana. Un século despois Edward Jenner atopou que o virus máis leve da varíola bovina podía usarse para vacinar efectivamente contra a máis grave e mortal varíola humana, e no século XX fíxose un esforzo mundial para estender a vacinación contra a varíola. O último caso de varíola epidémica ocorreu en Somalia en 1977. Despois dunha ampla investigación durante dous anos sen que se detectasen novos casos, en 1979 a Organización Mundial da Saúde (OMS) declarou que a doenza estaba oficialmente erradicada. En 1986, todas as mostras do virus foron destruídas ou transferidas a dous laboratorios de referencia aprobados pola OMS: na sede central dos Centros para a Prevención e Control de Enfermidades (CDC) federais de EUA en Atlanta, Xeorxia e no Instituto de Preparacións de Virus de Moscova, Rusia.[21] A partir dos atentados do 11 de setembro de 2001 en EUA, os gobernos de EUA e Reino Unido incrementaron a súa preocupación sobre o uso de virus causantes varíola ou de doenzas similares no bioterrorismo.
Poxviridae (poxvíridos), é unha familia de virus que infectan a vertebrados e invertebrados, cuxos membros se coñecen comunmente como poxvirus. A raíz pox do seu nome procede do inglés, idioma no que inicialmente se aplicou á doenza máis famosa que producen, a varíola, que despois pasou a chamarse smallpox.
Hai catro xéneros de poxvirus que poden infectar a humanos, que son:
Orthopoxvirus: virus da varíola, virus vaccinia, virus da varíola bovina, virus da varíola do mono; tamén é deste grupo o virus da varíola do camelo, que ás veces contaxia as mans dos humanos que manipulan eses animais. Parapoxvirus: virus orf , virus da pseudovaríola bovina, virus da estomatite papular bovina. Yatapoxvirus: virus da varíola do río Tana, virus do tumor do mono de Yaba. Molluscipoxvirus: virus do molusco contaxioso (MCV).Anteriormente o virus da varíola causaba grandes epidemias, pero foi erradicado pola vacinación. Actualmente, os máis comúns son o virus da vaccinia, que se pode atopar no subcontinente indio, e o virus do molusco contaxioso, aínda que están aumentando os casos de infeccións por virus da varíola do mono en zonas tropicais de África central.
Poxviridae adalah keluarga virus DNA beramplop yang termasuk golongan kelas I dalam klasifikasi Baltimore.[1] Poxviridae memiliki DNA untai ganda dengan morfologi yang besar dan komplek, serta tidak bersegmen.[1]
Anggota dari Poxviridae diantaranya adalah Orthopoxvirus, Parapoxvirus, Leporipoxvirus, Avipoxvirus, Molluscipoxvirus, Yatapoxvirus, Capripoxvirus, dan Suipoxvirus.[2] Smallpox yang menyebabkan cacar pada manusia termasuk dalam genus Orthopoxvirus. Kini smallpox telah dimusnahkan, dan imunisasi untuk pencegahan smallpox sudah tidak diberikan lagi.[2]
Virus lain yang termasuk dalam Orthopox diantaranya bufallopox, human monkeypox, dan cowpox. Ketiga virus ini memiliki inang hewan, tetapi dapat menginfeksi manusia dan menyebabkan lesi pada kulit.[2]

Poxviridae adalah keluarga virus DNA beramplop yang termasuk golongan kelas I dalam klasifikasi Baltimore. Poxviridae memiliki DNA untai ganda dengan morfologi yang besar dan komplek, serta tidak bersegmen.
Anggota dari Poxviridae diantaranya adalah Orthopoxvirus, Parapoxvirus, Leporipoxvirus, Avipoxvirus, Molluscipoxvirus, Yatapoxvirus, Capripoxvirus, dan Suipoxvirus. Smallpox yang menyebabkan cacar pada manusia termasuk dalam genus Orthopoxvirus. Kini smallpox telah dimusnahkan, dan imunisasi untuk pencegahan smallpox sudah tidak diberikan lagi.
Virus lain yang termasuk dalam Orthopox diantaranya bufallopox, human monkeypox, dan cowpox. Ketiga virus ini memiliki inang hewan, tetapi dapat menginfeksi manusia dan menyebabkan lesi pada kulit.
Poxviridae è una famiglia del phylum Nucleocytoviricota di virus a DNA a doppio filamento rivestiti da una doppia membrana virale.
La famiglia dei Poxviridae comprende due sottofamiglie:
Quattro generi di poxvirus possono infettare gli esseri umani: Orthopoxvirus, Parapoxvirus, Yatapoxvirus e Molluscipoxvirus[2]. I più comuni sono il virus del vaccino e del "Mollusco contagioso", ma infezioni per l'uomo possono provenire anche da alcuni Poxvirus degli animali come, per esempio, dal virus del vaiolo delle scimmie, responsabile di casi che si sono verificati in alcune remote località dell'Africa[3]. Apparteneva a questa famiglia anche il virus del vaiolo e dell'alastrim, malattie ormai eradicate[4].
Altri generi sono: Suipoxvirus, Avipoxvirus, Capripoxvirus, Leporipoxvirus
Le particelle virali dei Poxvirus sono normalmente dotate di un envelope formato da strati multipli di membrane modificate; in effetti, questo fa sì che spesso anche le forme immature intracellulari dei Poxvirus contengano envelope differenti rispetto al virus adulto e possono essere esse stesse infettive. Variano per forma e dimensioni a seconda delle specie e della maturazione: al microscopio elettronico appaiono a forma di manubrio all'esterno della cellula oppure di forma compatta, tondeggianti o simili a mattoni, all'interno della cellula. Il capside misura circa 200 nanometri di diametro e fino a 300 nanometri in lunghezza, mentre il genoma in esso contenuto è nella forma di una singola molecola lineare di DNA a doppio filamento.[5] In confronto, un Rhinovirus è grande solo un decimo di un tipico virione di Poxviridae.[6]
La replicazione dei Poxvirus richiede diversi stadi. Per prima cosa il virus si lega ad un recettore sulla superficie della cellula bersaglio; tale molecola è rappresentata con ogni probabilità da Glicosamminoglicani (GAGs). Dopo il contatto con il recettore, il virus entra nella cellula ove si libera del capside. La liberazione del genoma virale è un processo consistente di due fasi. Dapprima, al momento dell'entrata nella cellula ospite, viene rimossa la membrana virale esterna; successivamente la particella virale (rimasta senza membrana esterna) viene aperta per rilasciare il core nel citoplasma della cellula. Si compone di due fasi anche l'espressione genica.
I geni precoci codificano per le proteine non-strutturali, compresi gli enzimi necessari alla replicazione del genoma virale; i geni tardivi vengono espressi dopo la replicazione del patrimonio genetico e codificano le proteine strutturali del capside. L'assemblaggio della particella virale viene espletato nel citoscheletro della cellula: è un processo complesso ancora oggi poco compreso. Nonostante questi virus siano di grandi dimensioni e complessi, la loro replicazione risulta relativamente rapida, in quanto si svolge in sole 12 ore. La modalità di replicazione di questi virus sono insolite per un virus a DNA a doppio filamento sia perché il genoma virale codifica completamente il proprio sistema di riproduzione sia perché tale processo si verifica nel citoplasma, e non nel nucleo cellulare, dove sono presenti le DNA-polimerasi. La maggior parte degli altri virus a DNA sfrutta infatti l'apparato enzimatico dell'ospite per espletare le funzioni di replicazione del genoma.
Malattie causate da Poxvirus, specialmente il vaiolo, sono conosciute da secoli. Una delle prime prove documentate di questo è rappresentata dalla mummia del faraone Ramses V che morì di vaiolo attorno al 1000 a.C. Questa malattia fu probabilmente portata in Europa nel III secolo e quindi esportata in America dai Conquistadores ai primi del XVI secolo. È oramai riconosciuto che una pandemia di vaiolo fu la principale causa della rapida sconfitta degli Aztechi e che circa 3,2 milioni di essi perirono in soli due anni. Questo tasso di mortalità ed infettività fu dovuto all'assenza, nelle popolazioni americane, di anticorpi contro il virus di cui invece erano dotati gli europei.
Oltre due secoli dopo, Edward Jenner dimostrò che l'inoculazione del meno pericoloso vaiolo bovino poteva essere utilizzato per prevenire la forma più aggressiva di vaiolo. Un impegno mondiale per la vaccinazione di tutta la popolazione fu intrapreso con lo scopo di eradicare completamente la malattia dalla popolazione umana. L'Organizzazione Mondiale della Sanità dichiarò il virus ufficialmente sconfitto nel 1979, con campioni vivi in possesso ai laboratori delle due superpotenze del periodo, USA e Unione Sovietica. Dopo l'11 settembre 2001 i governi hanno incrementato la loro preoccupazione riguardo ad un possibile, futuro utilizzo del vaiolo o di altre malattie da Poxvirus in atti di bioterrorismo. Pertanto, sebbene dal 1979 nessun paese abbia vaccinato di routine i propri cittadini contro il vaiolo, dal 2001 gli USA hanno iniziato a vaccinare alcuni operatori della sanità e membri selezionati delle forze armate[7]
Debellato il virus del vaiolo con la vaccinazione, stanno comparendo altre patologie legate ai poxvirus, come per esempio il vaiolo delle scimmie. Statistiche recenti hanno mostrato un aumento di molte volte nel tasso di infezione di quest'ultima malattia, tenuta fino a poco tempo fa sotto controllo dalla vaccinazione obbligatoria contro il vaiolo umano[3].
Poxviridae è una famiglia del phylum Nucleocytoviricota di virus a DNA a doppio filamento rivestiti da una doppia membrana virale.
Pokswirusy (Poxviridae, z ang. pokkes - kieszonka) – rodzina wirusów wykazujących następujące cechy wspólne:
W obrębie rodziny Poxviridae wyróżnia się 8 rodzajów, ale tylko 2 gatunki są specyficzne w stosunku do człowieka. Inne (choć nie wszystkie) mogą także okazyjnie być chorobotwórcze dla ludzi, ale nie są to choroby szczególnie niebezpieczne. Podział systematyczny niniejszej rodziny przedstawia się następująco:
Wbrew zbieżności nazw z powyższymi chorobami, ospa wietrzna nie jest wywoływana przez pokswirusy, ale przez należącego do herpeswirusów wirusa ospy wietrznej-półpaśca.
Pokswirusy (Poxviridae, z ang. pokkes - kieszonka) – rodzina wirusów wykazujących następujące cechy wspólne:
Symetria: złożona, kształt zależy od rodzaju, są jednymi z najbardziej skomplikowanych pod względem budowy wirusów Osłonka lipidowa: czasami występuje, ale nie zawsze Kwas nukleinowy: liniowy dsDNA o kowalencyjnie związanych końcach. Na końcach tych znajdują się ITR (inverted terminal repetitions - odwrócone sekwencje powtórzone). Replikacja: zachodzi w cytoplazmie Wielkość: 270 x 300 nm (w przypadku wirusów owalnych), dzięki temu mogą nawet być dostrzeżone w mikroskopie świetlnym Cechy dodatkowe: ponieważ nie wnikają do jądra, pokswirusy tworzą ciała wtrętowe w cytoplazmie. wiriony zawierają kilkanaście właściwości enzymatycznych. Między innymi własną DNA-zależną polimerazę RNA, oraz ligazę DNA.Poxviridae ou poxvírus[1] é uma família que têm como principal característica a capacidade de infectar tanto os animais vertebrados como também os invertebrados. Partículas viróticas de poxvirus são geralmente envelopadas e variam em forma, dependendo da espécie, mas geralmente têm forma de tijolo ou uma forma mais oval. O virião[2] (o vírus que ainda não parasitou uma célula) tem um tamanho de aproximadamente 200 nm de diâmetro e 300 nm de comprimento e leva o seu genoma num simples e linear segmento de DNA (1). Em comparação, o Rhinovirus possui 1/10 do tamanho dum virião Poxviridae típico(2). A Microscopia electrónica dos gêneros Orthopoxvirus e Parapoxvirus, incluindo o vírus da varíola, foi coligida pelo International Committee on Taxonomy of Viruses no site Poxviridae picture gallery.
O protótipo da família poxvírus é o vírus vaccinia, o qual tem sido usado com sucesso como vacina para erradicar o vírus da varíola. O vírus vaccinia também tem sido utilizado como uma ferramenta eficaz capaz de provocar uma forte resposta imunitária por parte de um hospedeiro. O vírus vaccinia penetra nas células principalmente por fusão celular, apesar de actualmente ainda não se conhecer qual o receptor responsável pela ligação do vírus à célula.
O nome da família Poxviridae é uma herança do agrupamento original dos vírus associados com doenças que produzem vesículas (pox em inglês). A moderna classificação viral é baseada na forma e características moleculares dos vírus, sendo que o vírus da varíola permanece como o mais notável desta família.
O outro poxvirus conhecido por especificamente infectar humanos é o responsável pelo molluscum contagiosum virus (MCV).(3)
Os seguintes géneros são atualmente incluídos aqui:
Poxviridae ou poxvírus é uma família que têm como principal característica a capacidade de infectar tanto os animais vertebrados como também os invertebrados. Partículas viróticas de poxvirus são geralmente envelopadas e variam em forma, dependendo da espécie, mas geralmente têm forma de tijolo ou uma forma mais oval. O virião (o vírus que ainda não parasitou uma célula) tem um tamanho de aproximadamente 200 nm de diâmetro e 300 nm de comprimento e leva o seu genoma num simples e linear segmento de DNA . Em comparação, o Rhinovirus possui 1/10 do tamanho dum virião Poxviridae típico. A Microscopia electrónica dos gêneros Orthopoxvirus e Parapoxvirus, incluindo o vírus da varíola, foi coligida pelo International Committee on Taxonomy of Viruses no site Poxviridae picture gallery.
O protótipo da família poxvírus é o vírus vaccinia, o qual tem sido usado com sucesso como vacina para erradicar o vírus da varíola. O vírus vaccinia também tem sido utilizado como uma ferramenta eficaz capaz de provocar uma forte resposta imunitária por parte de um hospedeiro. O vírus vaccinia penetra nas células principalmente por fusão celular, apesar de actualmente ainda não se conhecer qual o receptor responsável pela ligação do vírus à célula.
O nome da família Poxviridae é uma herança do agrupamento original dos vírus associados com doenças que produzem vesículas (pox em inglês). A moderna classificação viral é baseada na forma e características moleculares dos vírus, sendo que o vírus da varíola permanece como o mais notável desta família.
O outro poxvirus conhecido por especificamente infectar humanos é o responsável pelo molluscum contagiosum virus (MCV).
Os seguintes géneros são atualmente incluídos aqui:
Subfamília Chordopoxvirinae Gênero Orthopoxvirus; espécies: Vaccinia virus; diseases: cowpox, vaccinia, smallpox Gênero Parapoxvirus; espécies: Orf virus Gênero Avipoxvirus; espécies: Fowlpox virus Gênero Capripoxvirus; espécies: Sheeppox virus Gênero Leporipoxvirus; espécies: Myxoma virus Gênero Suipoxvirus; espécies: Swinepox virus Gênero Molluscipoxvirus; espécies: Molluscum contagiosum virus Gênero Yatapoxvirus; espécies: Yaba monkey tumor virus Subfamília Entomopoxvirinae Gênero Entomopoxvirus A; espécies: Melolontha melolontha entomopoxvirus Gênero Entomopoxvirus B; espécies: Amsacta moorei entomopoxvirus Gênero Entomopoxvirus C; espécies: Chironomus luridus entomopoxvirusPoxviridae, aparține clasei 1 conform clasificării Baltimore, genomul viral fiind alcătuit din ADn dublu catenar.Infecțiile determinate de reprezentanții acestei clase infectează atât vertebratele cât și nevertebratele.
Este format din ADN dublucatenar, format din 130000-375000 nucleotide.Genomul are o secvență TR (terminal repeat),și ITR (inverted terminal repeat), situate la capetele lanțului nucleotidic.Aceste secvențe conțin [1]:
Guanina șicitozina au un procent de circa 35-64%, sau 20%(Entopomoxvirus).
[2] Virionul are un diametru de circa 140-260 nm, ADN monocatenar, putând fi găsit sub 4 forme
Mai există și cea de a 5-a formă numită A-type sau ATI întâlnită la unele pox virusuri. „Prototipul” pox virusurilor este Vaccinia virus(VV,VACV) , utilizat pentru vaccinarea împotriva variolei.
Replicarea este un proces ce se desfășoară în căteva stadii:
Poxviridae, aparține clasei 1 conform clasificării Baltimore, genomul viral fiind alcătuit din ADn dublu catenar.Infecțiile determinate de reprezentanții acestei clase infectează atât vertebratele cât și nevertebratele.
Poddružina Chordopoxvirinae
Avipoxvirus
Capripoxvirus
Cervidpoxvirus
Crocodylipoxvirus
Leporipoxvirus
Molluscipoxvirus
Orthopoxvirus
Parapoxvirus
Suipoxvirus
Yatapoxvirus
nedodeljeni
Poddružina Entomopoxvirinae
Alphaentomopoxvirus
Betaentomopoxvirus
Gammaentomopoxvirus
nedodeljeni
Poksvirusi ali Poxviridae so družina velikih kompleksnih virusov z dvoverižno DNK in večslojnimi ovojnicami, ki imajo afiniteto do kože in so patogeni za človeka in živali (za človeka so pomembni rodovi Orthopoxvirus, Parapoxvirus, Molluscipoxvirus in Yatapoxvirus).[1][2] So opekaste ali jajčaste oblike. Vsi predstavniki imajo skupen nukleoproteinski antigen; razmnožujejo se v citoplazmi gostiteljske celice. So povzročitelji koz pri človeku (virus črnih koz) in živalih (govedo, koze, prašiči, perutnina, kunci, zajci). Nekateri predstavniki so patogeni za človeka in živali (virus govejih koz). Med poksviruse spada tudi virus vakcinije, ki so ga uporabljali za cepljenje proti črnim kozam.[3]
Pri človeku lahko povzročijo bolezen štirje rodovi: ortopoksvirusi, parapoksvirusi, jatapoksvirusi in moluskipoksvirusi:
ortopoksvirusi: virus črnih koz (virus variole), virus vakcinije, virus kravjih koz, virus opičjih koz;
parapoksvirusi: virus orfa, virus psevdokravjih koz, virus govejega papularnega stomatitisa;
jatapoksvirusi: tanapoksvirus, tumorski virus opic jaba;
moluskipoksvirusi: virus mehkužk (virus molluscum contagiosum).[4]
Poksvirusni virioni imajo praviloma zunanjo virusno ovojnico, vendar so kužni tudi znotrajcelični zreli virioni, ki vsebujejo drugačno obliko ovojnice. Vrste se med seboj razlikujejo po obliki, vendar so nasplošno opekaste ali jajčaste oblike. Poksivirusi so nenavadno veliki in merijo okoli 200 nm v premer in 300 nm v dolžino. Virusna dednina je v obliki enojnega, linearnega dvoverižnega odseka DNK.[5] Rinovirusi, ki povzročajo prehlad, so na primer desetkrat manjši od tipičnega poksvirusa.[6]
Podvojevanje poksvirusov se začne v citoplazmi, kar sicer ni značilno za viruse z dvoverižno DNK (dsDNA),[7] je pa značilno tudi za druge velike viruse.[8] Poksvirusi nosijo sami zapis za ustroj, potreben za prepis genoma, in sicer zapis za encim od DNK odvisno RNK-polimerazo.[9] Le-ta omogoči prepisovanje virusne DNK v gostiteljevi citoplazmi. Sicer večina drugih dsDNA-virusov potrebuje gostiteljevo od DNK odvisno RNK-polimerazo, ta pa se nahaja v celičnem jedru gostiteljeve celice. Zato se cikel podvojevanja drugih dsDNA-virusov dogodi v samem jedru.
Poksvirusi ali Poxviridae so družina velikih kompleksnih virusov z dvoverižno DNK in večslojnimi ovojnicami, ki imajo afiniteto do kože in so patogeni za človeka in živali (za človeka so pomembni rodovi Orthopoxvirus, Parapoxvirus, Molluscipoxvirus in Yatapoxvirus). So opekaste ali jajčaste oblike. Vsi predstavniki imajo skupen nukleoproteinski antigen; razmnožujejo se v citoplazmi gostiteljske celice. So povzročitelji koz pri človeku (virus črnih koz) in živalih (govedo, koze, prašiči, perutnina, kunci, zajci). Nekateri predstavniki so patogeni za človeka in živali (virus govejih koz). Med poksviruse spada tudi virus vakcinije, ki so ga uporabljali za cepljenje proti črnim kozam.
Pri človeku lahko povzročijo bolezen štirje rodovi: ortopoksvirusi, parapoksvirusi, jatapoksvirusi in moluskipoksvirusi:
ortopoksvirusi: virus črnih koz (virus variole), virus vakcinije, virus kravjih koz, virus opičjih koz;
parapoksvirusi: virus orfa, virus psevdokravjih koz, virus govejega papularnega stomatitisa;
jatapoksvirusi: tanapoksvirus, tumorski virus opic jaba;
moluskipoksvirusi: virus mehkužk (virus molluscum contagiosum).
Poxvirus är de största och mest komplexa virusarterna. Generna är uppbyggda av linjärt dubbelsträngat DNA och är försedda med ett hölje. Till skillnad från andra virus med DNA replikerar viruset i cellens cytoplasma vilket innebär att viruset behöver föra med sig olika enzymer t.ex. RNA-polymeras för att möjliggöra transkription. Inom virusfamiljen återfinns variolaviruset som orsakar smittkoppor.
Poxvirus är de största och mest komplexa virusarterna. Generna är uppbyggda av linjärt dubbelsträngat DNA och är försedda med ett hölje. Till skillnad från andra virus med DNA replikerar viruset i cellens cytoplasma vilket innebär att viruset behöver föra med sig olika enzymer t.ex. RNA-polymeras för att möjliggöra transkription. Inom virusfamiljen återfinns variolaviruset som orsakar smittkoppor.
Віріон має цеглиноподібну форму з округлими краями розміром 300-450 * 170-260 нм. Це найбільший вірус тварин. Він має складну будову та складається із ядра, що має форму двоввігнутої лінзи. По обидва боки від якої знаходяться овальні утвори що називаються боковими тільцями. Ядро містить ДНК, упаковану в білковий футляр у вигляді циліндричних чи ниткоподібних структур і оточені внутрішньою та зовнішньою мембранами. Ядро та білкові тіла оточені зовнішньою оболонкою з характерними кулеподібними виступами.
Вірус віспи є стійким до впливу зовнішнього середовища. Він стійкий до ефіру, більшості дезінфікуючих засобів, до температури та може зберігатися в сухих ексудатах на протязі багатьох місяців при кімнатній температурі, а в разчинах везикул на холоді – на протязі декількох років. В 50% розчині гліцерину вірус зберігає інфекційну активність при 4°С протягом декількох років.
Геном вірусу віспи являє собою лінійну молекулу ДНК з молекулярно массою більш ніж 130*106, значно більшою ніж у інших вірусів тварин. ДНК містить близько 240 000 пар нуклеотидів при загальній довжині 82 мкм. На обох кінцях молекули знаходяться ковалентні зшивки, тому ниткам ДНК властива незвичайна міцність і не розділяються повністю при лужній денатурації.
Розмір геному 130-375kb.
Стратегія реплікації: Цитоплазматична
Віріони містять у своєму складі дволанцюгову ДНК, що становить 5 — 7,5 % від маси вібріона, більш ніж 30 структурних білків, включаючи ферменти, ліпіди (4 %), вуглеводи (3 %). Ліпіди входять до складу зовнішньої оболонки вібріонів.
В складі вібріонів містяться більше 30 структурних білків з молекулярною масою 11*103 — 150*103. З них 17 виявлені в серцевині, в тому числі два білка, що становлять 50 % від сумарного білка серцевини. П'ять білків локалізовано на поверхні вібріона, вісім — в більш глибоких шарах зовнішньої оболонки. Поверхневі білки є глікопротеїнами і скоріш за все з ними пов'язаний імунітет до віспи.
В вібріонах віспи виявлено 12-13 ферментів. Серед них знаходиться ДНК-залежна РНК-полімераза та інші ферменти зв'язані з транскрипцією ДНК, модифікацією мРНК, кінази та протеази.
Віруси віспи та вісповакцини подібні в антигенному відношенні і відрізняються один від одного не більш ніж одним антигеном. Всі антигени можна розділити на структурні, розчинні та гемаглютиніни. До структурних білків антигенів належать NP-антиген, загальний всього сімейства. Цей білок зв'язаний з білком нуклеокапсида. Серед розчинних антигенів є термолабільний та термостабільний. Гемаглютинін є ліпопротеїновим комплексом, що містить три глікопротеїни.
Вірус проникає в клітини слизової оболонки верхніх дихальних шляхів і після розмноження по лімфатичній системі переноситься в регіональні лімфатичні вузли. В них відбувається подальше розмноження вірусу і наступає первинна вірусемія яка приводить до інвазії ретикулоендотеліальної системи. Починається інтенсивне розмноження вірусу, що призводить до вторинної вірусемії. Після цього вірус накопичується в епідермісі і викликає ураження шкіри. З появою висипу хворий стає заразним. Захворювання розвивається після інкубаційного періоду 8-18 днів і починається раптовою лихорадкою, головним болем, болем в м'язах, прострацією та появою характерного висипу. Висип проходить стадії макули, папули, везикули, пустули і завершується утворенням рубця – все це за 3 тижні.
Натуральна віспа передається повітряно-крапельним шляхом, зараження наступає при контакті з хворим. Зараження може також відбутися через предмети побуту та одяг хворого тому, що вірусу характерна стійкість до зовнішнього середовища. Хворий заразний протягом всього періоду хвороби, та найбільш всього в період перших 8-10 днів.
Епідемії виникають в багатьох країнах але головними є Південно-Західна Азія (Індія, Пакистан, Бангладеш). В СРСР віспа була ліквідована в 1936 р., проте і після цього вона не одноразово заносилася з сусідніх країн (Іран, Афганістан).
Господар: як хребетні (в тому числі людина), так і безхребетні. Легко передається від тварини до людини (зоонози).
Географічне розповсюдження: по всьому світі.
Топізм: епітеліальні тканини (особливо респіраторної системи)
Для лікування крім патогенетичної та симптоматичної терапії, використовували хіміотерапевтичний препарат із группи β-тіосемикарбазонів – метисазон. Його ж застосували для профілактики.
Однак основним засобом за допомогою якого була досягнута перемога над віспою була віспяна вакцина запропонована в 1796 році. Звичайно в наш час препарати віспяної вакцини значно вдосконалені як і методи щеплення. Для масових щеплень застосовують безголкові ін’єкції, а для індивідуальних – біфуркаціонна голка, що дозволяє точно дозувати вакцину. Перемога над віспою була досягнена також в результаті правильної організації виявлення та подавлення іі появ.
Poxviridae
Подсемейства Группа по БалтиморуI: дцДНК-вирусы
Поксви́русы[2] или вирусы оспы[3] (лат. Poxviridae, от англ. рох — оспа + вирусы) — семейство вирусов животных, объединяющее самые крупные ДНК-содержащие вирусы, вирионы которых имеют кирпичеобразную или овоидную форму; включает виды, патогенные для человека, например, возбудителей оспы, пустулезного дерматита.
Геном, представленный двунитчатой линейной гантелеобразной формы ДНК, покрыт двухслойным капсидом, между слоями которого находятся боковые тела. Поверх нуклеокапсида расположена двухслойная липопротеидная оболочка с воронкообразными фибрами.
Поксвирусы обладают наиболее сложным репродуктивным циклом, при этом синтезируется более 100 различных белков, входящих в состав вирионов (большинство образует наружную оболочку). Репродукция поксвирусов характеризуется следующими особенностями.
В репродуктивном цикле выделяют три стадии[4]:
По данным Международного комитета по таксономии вирусов (ICTV), на март 2017 года в семейство включают 2 подсемейства с 11 и 3 родами соответственно[5]:
Поксви́русы или вирусы оспы (лат. Poxviridae, от англ. рох — оспа + вирусы) — семейство вирусов животных, объединяющее самые крупные ДНК-содержащие вирусы, вирионы которых имеют кирпичеобразную или овоидную форму; включает виды, патогенные для человека, например, возбудителей оспы, пустулезного дерматита.
Геном, представленный двунитчатой линейной гантелеобразной формы ДНК, покрыт двухслойным капсидом, между слоями которого находятся боковые тела. Поверх нуклеокапсида расположена двухслойная липопротеидная оболочка с воронкообразными фибрами.
亞科 痘病毒脊索亞科(Chordopoxvirinae)
正痘病毒屬(Orthopoxvirus)
副痘病毒屬(Parapoxvirus)
禽痘病毒屬(Avipoxvirus)
山羊痘病毒屬(Capripoxvirus)
野兔痘病毒屬(Leporipoxvirus)
豬痘病毒屬(Suipoxvirus)
軟疣痘病毒屬(Molluscipoxvirus)
亞塔痘病毒屬(Yatapoxvirus)
亞科 痘病毒昆蟲亞科(Entomopoxvirinae)
A昆蟲痘病毒屬(Entomopoxvirus A)
B昆蟲痘病毒屬(Entomopoxvirus B)
C昆蟲痘病毒屬(Entomopoxvirus C)
痘病毒(Poxviridae,pox=發疹的病)為哺乳動物的最大及最複雜的病毒(170×950nm)。包含兩個亞科痘病毒脊索亞科 (Chordopoxvirinae)、痘病毒昆蟲亞科 (Entomopoxvirinae),這兩個亞科各有8個和4個屬。不同的動物有不同品系的病原病毒。遺傳物質為雙股DNA,長度約130-375knt; 病毒直徑約140-260 nm,最大的病毒就是屬於痘病毒科。通常可區分為8屬(genera),分別為正痘病毒屬 (Orthopoxvirus)、駱駝痘病毒屬 (Capripoxvirus)、豬痘病毒屬 (suipoxvirus)、兔痘病毒屬 (Leporipoxvirus)、軟體動物痘病毒屬 (Molluscipoxvirus) 、雅塔痘病毒屬(Yatapoxvirus)、禽痘病毒屬 (Avipoxvirus)、副痘病毒屬 (Parapoxvirus),其中兩屬(Orthopoxvirus及Parapoxvirus)為人類的重要病原。同一屬病毒的血清反應常有廣泛的交叉反應,但各屬間僅有一種共同抗原,此抗原一般認為是一種核蛋白複合體。天花的病原Variola病毒為Orthopoxvirus屬,此病毒曾經摧殘全人類,目前已被撲滅,然最近常被認為為一種有可能之再新興之疾病。其他的痘病毒如駱駝痘(camel pox)及ectromelia造成低等動物的皮膚發疹。天花(Variola)及傳染性軟疣(molluscum contagiosum)病毒只對人類有病原性。
組織培養或水痘液的病毒顆粒有C及M兩型 (Cubed, Mulberry)。C型為磚塊型有一寬界明顯的外膜,中心電子密度較大。Mulberry(桑椹)型簡稱M型,外表有紡錘絲狀排列。M型顆粒之中心常凹陷,代表兩個圓形側體(lateral body),核仁及雙鏈DNA基因,其功用目前尚不十分明瞭。所有的痘病毒都於細胞質內增生,造成嗜酸性質內包涵體。痘病毒通常感染上皮細胞造成丘疹水及膿。
其他的病如綿羊痘(sheep pox)及羊接觸傳染性化膿性口炎(orf)及牛痘(cow pox)為其它引起動物發疹的病毒,也會傳染給人類,除了parapox屬外,所有的痘病毒都是170~250×300~450nm之磚形顆粒,表面有紡錘絲狀的結構。parapox(orf等)較呈卵形,表面絲狀排列為規則之螺旋型,平均僅為150×200nm所有痘病毒的內部結構大致上都是一致的。
根据最新的研究痘病毒科可能属于核质巨DNA病毒。
痘病毒(Poxviridae,pox=發疹的病)為哺乳動物的最大及最複雜的病毒(170×950nm)。包含兩個亞科痘病毒脊索亞科 (Chordopoxvirinae)、痘病毒昆蟲亞科 (Entomopoxvirinae),這兩個亞科各有8個和4個屬。不同的動物有不同品系的病原病毒。遺傳物質為雙股DNA,長度約130-375knt; 病毒直徑約140-260 nm,最大的病毒就是屬於痘病毒科。通常可區分為8屬(genera),分別為正痘病毒屬 (Orthopoxvirus)、駱駝痘病毒屬 (Capripoxvirus)、豬痘病毒屬 (suipoxvirus)、兔痘病毒屬 (Leporipoxvirus)、軟體動物痘病毒屬 (Molluscipoxvirus) 、雅塔痘病毒屬(Yatapoxvirus)、禽痘病毒屬 (Avipoxvirus)、副痘病毒屬 (Parapoxvirus),其中兩屬(Orthopoxvirus及Parapoxvirus)為人類的重要病原。同一屬病毒的血清反應常有廣泛的交叉反應,但各屬間僅有一種共同抗原,此抗原一般認為是一種核蛋白複合體。天花的病原Variola病毒為Orthopoxvirus屬,此病毒曾經摧殘全人類,目前已被撲滅,然最近常被認為為一種有可能之再新興之疾病。其他的痘病毒如駱駝痘(camel pox)及ectromelia造成低等動物的皮膚發疹。天花(Variola)及傳染性軟疣(molluscum contagiosum)病毒只對人類有病原性。
ポックスウイルス科(Family Poxviridae)はウイルスの分類における1科である。この科に属するウイルスは線状の2本鎖DNAをゲノムとして持つDNAウイルスであり、そのビリオンは220~450nm×140~260nm×140~260nmの煉瓦状ないし卵形で、エンベロープを有する。通常のエンベロープを持つウイルスとは異なりエーテル耐性のものも存在する(オルトポックスウイルス属、アビポックスウイルス属)。また、ウイルスの増殖は他のDNAウイルスと異なり宿主細胞の細胞質内で行われる。感染経路としては経気道や接触によるものが多く、節足動物による機械的伝播も起こりやすい。ポックスウイルス科による病気の特徴は皮膚の発疹様斑紋である。ポックスウイルス科のウイルスによる疾患としては天然痘が有名である。
ポックスウイルスはレンガ型あるいは卵形をしたウイルスであるが、全長が220~450nmもある(→インフルエンザウイルスやヒト免疫不全ウイルスではだいたい100nmほど)大きなDNAウイルスである。また、通常のDNAウイルスは感染細胞の酵素を借りて核内で増殖するが、ポックスウイルスはDNA複製に必要な酵素のほとんどを自前で持っているので細胞質内で増殖を行う。
ポックスウイルス科(Family Poxviridae)はウイルスの分類における1科である。この科に属するウイルスは線状の2本鎖DNAをゲノムとして持つDNAウイルスであり、そのビリオンは220~450nm×140~260nm×140~260nmの煉瓦状ないし卵形で、エンベロープを有する。通常のエンベロープを持つウイルスとは異なりエーテル耐性のものも存在する(オルトポックスウイルス属、アビポックスウイルス属)。また、ウイルスの増殖は他のDNAウイルスと異なり宿主細胞の細胞質内で行われる。感染経路としては経気道や接触によるものが多く、節足動物による機械的伝播も起こりやすい。ポックスウイルス科による病気の特徴は皮膚の発疹様斑紋である。ポックスウイルス科のウイルスによる疾患としては天然痘が有名である。











