Els planctomicets (Planctomycetes) són un filum d'eubacteris aquàtics que han estat descoberts en mostres d'aigua salabrosa, marina i dolça. Es reprodueixen per gemmació. Quant a la seva estructura, són organismes ovoides dotats d'un rizoide a l'extrem no reproductor, que els permet agafar-se l'un a l'altre durant la gemmació.
Thomas Cavalier-Smith ha postulat que els planctomicets es troben dins del clade dels planctobacteris, dins del clade més gran dels gracilicuts.
Els planctomicets (Planctomycetes) són un filum d'eubacteris aquàtics que han estat descoberts en mostres d'aigua salabrosa, marina i dolça. Es reprodueixen per gemmació. Quant a la seva estructura, són organismes ovoides dotats d'un rizoide a l'extrem no reproductor, que els permet agafar-se l'un a l'altre durant la gemmació.
Thomas Cavalier-Smith ha postulat que els planctomicets es troben dins del clade dels planctobacteris, dins del clade més gran dels gracilicuts.
Planctomycetes („planktomycéty“) jsou kmen[1] bakterií, které se vyskytují v sladké, brakické i slané vodě. Rozmnožují se pučením. Tyto bakterie mají oválný tvar a na povrchu cytoplazmy i zvláštní tubulární útvary, zvané stébla, která jim pomáhají připevnit se k ostatním jedincům v procesu pučení.
Podle Cavalier-Smithe se Planctomycetes řadí do vývojové větve Planctobacteria, která dále patří do velké skupiny Gracilicutes.
Tyto bakterie postrádají peptidoglykan ve své buněčné stěně, který u většiny bakterií slouží jako jakási ochranná vnější kostra buňky. Místo peptidoglykanu obsahují glykoprotein, bohatý zejména na glutamát. Také vnitřní struktury jsou v rámci prokaryot výjimečné. Přestože nemají pravé buněčné jádro, jako eukaryota, jejich DNA (nukleoid) může být u Planctomycetes uzavřena v dvojité membráně a takto vzniklý „váček“ se nazývá pirerulozom.[2][3]
Životní cyklus obsahuje střídání mezi přisedlými a bičíkatými fenotypy. Přisedlé buňky totiž vypučí v pohyblivé buňky s bičíky, které plavou a posléze přilnou k povrchu a rozdělí se.
V tomto článku byl použit překlad textu z článku Planctomycetes na anglické Wikipedii.
Planctomycetes („planktomycéty“) jsou kmen bakterií, které se vyskytují v sladké, brakické i slané vodě. Rozmnožují se pučením. Tyto bakterie mají oválný tvar a na povrchu cytoplazmy i zvláštní tubulární útvary, zvané stébla, která jim pomáhají připevnit se k ostatním jedincům v procesu pučení.
Podle Cavalier-Smithe se Planctomycetes řadí do vývojové větve Planctobacteria, která dále patří do velké skupiny Gracilicutes.
Die Planctomycetes (Planctomyceten) bilden eine gut abgrenzbare Abteilung (Divisio), auch als Phylum bezeichnet, innerhalb der PVC-Gruppe in der Domäne der Bakterien. Man zählt die Klasse der Planctomycea (vorher: Planctomycetacia) mit der Ordnung der Planctomycetales und der Familie der Planctomycetaceae sowie die Klasse der Phycisphaerae dazu. Planctomyceten wurden in den 1970er Jahren entdeckt. Seither hat man sie fast überall in der Umwelt nachweisen können. Viele Arten leben aquatisch, sowohl im Salz- wie im Süßwasser. Einige kommen in hypersalinen (stark salzhaltigen) Gewässern vor, andere auch im Boden oder in Klärschlamm. Die wichtigsten Gattungen sind Planctomyces, Pirellula, Isosphaera und Gemmata.
Die Bezeichnung Planctobacteria wird manchmal im engeren Sinne alternativ zu Planctomycetes verwendet, kann aber auch im weiteren Sinne die gesamte PVC-Gruppe bezeichnen.
Die Planctomyceten sind in mehrfacher Hinsicht einzigartig. Die Zellen von Planctomyces sind extrem stark kompartimentiert und besitzen oft – ähnlich wie die Eukaryoten – eine Membran, die die DNA umgibt. Die Zellen sind gestielt und zeigen einen dimorphen Lebenszyklus, der auch von Caulobacter, einer Art der Alpha-Proteobakterien, bekannt ist: Eine sessile Zelle schnürt eine Tochterzelle ab, die am entgegengesetzten Pol eine Geißel bildet. Die Schwärmerzelle ist mobil, wirft die Geißel aber nach einiger Zeit ab und bildet nun ihrerseits einen Stiel, mit dem sie sich an eine feste Oberfläche anheftet. Der Stiel von Planctomyces-Zellen ist anders aufgebaut als der von Caulobacter, beide Gattungen sind nicht verwandt.
Planktomyceten sind auch deshalb ungewöhnlich, Bis vor einigen Jahren galten Planktomyceten auch deshalb als ungewöhnlich, weil in der Zellwand kein Murein gefunden wurde. Stattdessen war nur ein proteinreicher S-Layer bekannt. Im Jahr 2015 wurde allerdings festgestellt, dass bei einigen Planctomyceten, z. B. bei der Art Gammeta obscuriglobus, auch Peptidoglycan in ihren Zellwänden vorhanden ist.[1][2]
Die Bakterien leben chemoorganotroph und sind fakultativ aerob, neben gestielten existieren auch filamentöse Formen. So ist die Art Isosphaera pallida ein filamentöses, sich gleitend fortbewegendes Bakterium, das in heißen Quellen bei Temperaturen zwischen 35 und 55 °C vorkommt.
Einige Planctomyceten bilden Assoziationen mit Tieren, wie z. B. Insekten. Einige nicht näher identifizierte Planctomyceten wurden mit Hilfe von 16S-rRNA-Analysen in Verdauungstrakten der Termiten Cubitermes ugandensis und Cubitermes orthognathus gefunden.[3][4] Ein Planktomycet vermutlich der Gattung Pirellula wurde in dem Wasserfloh Daphnia pulex gefunden. Eine weitere Art wurde aus der Garnele Penaeus monodon isoliert. Auch in den Schwämmen Aplysina cavernicola und Aplysina aerophoba wurden Planctomyceten nachgewiesen.
In welcher Verbindung die Bakterien mit den Wirten stehen (Symbiose oder Parasitismus) und welche Funktion sie im Verdauungstrakt ausführen, ist noch unklar.
Des Weiteren konnten Planctomyceten auf der Oberfläche einer Vielzahl von Makroalgen nachgewiesen werden. Als Teil des Biofilms nutzen sie von den Algen produzierte Moleküle als Nährstoffe. Einige Taxa der Planctomyceten wurden bisher in keinem anderen Habitat beobachtet. Häufig stellen sie nur einen geringen Anteil des Mikrobioms, auf der Braunalge Laminaria hyperborea jedoch nehmen sie einen Anteil von bis zu 53 % ein.[5]
Analysen von 16S rRNA Gensequenzen und weitere Untersuchungen haben eine enge Verwandtschaft der Planctomycetes und den Abteilungen der Verrucomicrobia und Chlamydiae gezeigt. Aufgrund dessen bilden sie zusammen das sogenannten PVC-Superphylum.[6]
Mitglieder (Stand 17. März 2022):[7]
Für einige der Ordnungen folgt eine Auswahl an Mitgliedern (Familien und Gattungen):
Die Planctomycetes (Planctomyceten) bilden eine gut abgrenzbare Abteilung (Divisio), auch als Phylum bezeichnet, innerhalb der PVC-Gruppe in der Domäne der Bakterien. Man zählt die Klasse der Planctomycea (vorher: Planctomycetacia) mit der Ordnung der Planctomycetales und der Familie der Planctomycetaceae sowie die Klasse der Phycisphaerae dazu. Planctomyceten wurden in den 1970er Jahren entdeckt. Seither hat man sie fast überall in der Umwelt nachweisen können. Viele Arten leben aquatisch, sowohl im Salz- wie im Süßwasser. Einige kommen in hypersalinen (stark salzhaltigen) Gewässern vor, andere auch im Boden oder in Klärschlamm. Die wichtigsten Gattungen sind Planctomyces, Pirellula, Isosphaera und Gemmata.
Die Bezeichnung Planctobacteria wird manchmal im engeren Sinne alternativ zu Planctomycetes verwendet, kann aber auch im weiteren Sinne die gesamte PVC-Gruppe bezeichnen.
Ang Planctomycetes ay isang uri ng phylum sa bakterya kahariang Protista. Hindi malaman kung saan ito maikakategorya, sa Gram-positive bacteria o Gram-negative bacteria.
![]() Ang lathalaing ito na tungkol sa Bakterya ay isang usbong. Makatutulong ka sa Wikipedia sa nito.
Ang lathalaing ito na tungkol sa Bakterya ay isang usbong. Makatutulong ka sa Wikipedia sa nito.
Planctomycetes es un phylo de Negibacteria.
Планктомицетите (Planctomycetes) се колено на водни бактерии кои живеат во морски, слатки и мешани води. Се размножуваат со пупење. По форма се овоидни (јајцевидни), на дел од клеточното тело имаат тенок цилиндричен продолжеток кој на својот врв има прицврстувач кој помага меѓусебно да се прилепат за време на пупењето.
Долго време бактерии кои припаѓаат на оваа група се сметаше дека немаат пептидогликан (муреин) во клеточниот ѕид, кој е важен хетерополимер присутен во повеќето бактериски клеточни ѕидови и служи како заштитна компонента. Наместо тоа се претпоставуваше дека нивните клеточни ѕидови се изградени од гликопротеин кој е богат со глутамат. Сепак, неодамна, беа пронајдени претставниците од сите три таксони во рамките на Planctomycetes кои поседуваат пептидогликан во клеточниот ѕид.[1][2]
Планктомицетите имаат карактеристична морфологија со интрацелуларно мембрански-оградени простори кои се нарекуваат: paryphoplasm (простор во кој нема рибозоми), pirellulosome (простор во кој има рибозоми) и nucleoid (простор кој содржи кондензирана нуклеинската киселина одвоен со двојна мембрана).[3][4] Се до скорешното откривање на Poribacteria, планктомицетите беа единствените познати бактерии со внатрешно-клеточни мембрански прегради.[5] Тродимензионална електронско томографска реконструкција на репрезентативниот вид, Gemmata obscuriglobus, има дадено разни интерпретации за опсервацијата на внатрешно-клеточните простори. Студија од 2013 година, најде дека појавата на внатрешните прегради се должи на густо инвагинирана, но континуирана единечна мембрана, заклучувајќи дека се присутни само два клеточни простори типични за Грам-негативните бактерии - цитоплазма и периплазма. Како и да е, вишокот на мембрана тројно ја зголемува површината на клетката во однос на нејзиниот волумен, што можеби е поврзано со способноста на родот Gemmata да биосинтетизира стероли.[6] Меѓутоа понова студија од 2014 година со употреба на слични методи го поддржува претходниот став дека мембрански-оградените внатрешни простори се од затворен тип.[7]
Неодамна се покажа дека видот Gemmata obscuriglobus е способен да внесува големи молекули во клетката преку процес кој наликува на еукариотската ендоцитоза.[8][9]
Иако Planctomycetes се познати по своите необични клеточни карактеристики, нивната специфичност во однос на сите други бактерии дополнително е поддржана од два CSIs (анг. conserved signature indels).[10] Овие CSIs ја одвојуваат оваа група од соседните колена во рамките на PVC суперколеното.[11]
Еден CSI го споделуваат сите членови на PVC групата, вклучувајќи ги и Planctomycetes. Planctomycetes, исто така, поседуваат CSP (анг. conserved signature protein), за кој е откриено дека врши важна функција која ексклузивно е присутна кај членовите на PVC групата.[12]
Животниот циклус на многу планктомицети е наизменична смена меѓу сесилни и подвижни форми. Неподвижните сесилни клетки пупат и формираат клетки со камшиче кои пливаат, за по некое време да се закачат и поминат во сесилна форма која повторно ја отпочнува репродукцијата.
Филогенијата се заснова на проектот Проектот "Животно дрво на сите видови" (анг. The All-Species Living Tree Project).[13]
Моментално прифатената таксономија е врз основа на Листата на имиња на прокариотите со евиденција во номенклатурата (анг. List of Prokaryotic names with Standing in Nomenclature - LPSN)[14] и Националниот центар за биотехнолошки информации (анг. National Center for Biotechnology Information - NCBI).[15]
♠ Видовите се наоѓаат во NCBI, но не се наведени во LSPN.
♪ Прокариоти кои не се изолирани од чисти култури или не се достапни, бидејќи не можат да се одгледуваат во култура или не можат да се одржат во култура.
Планктомицетите (Planctomycetes) се колено на водни бактерии кои живеат во морски, слатки и мешани води. Се размножуваат со пупење. По форма се овоидни (јајцевидни), на дел од клеточното тело имаат тенок цилиндричен продолжеток кој на својот врв има прицврстувач кој помага меѓусебно да се прилепат за време на пупењето.
The Planctomycetota are a phylum of widely distributed bacteria, occurring in both aquatic and terrestrial habitats.[5] They play a considerable role in global carbon and nitrogen cycles, with many species of this phylum capable of anaerobic ammonium oxidation, also known as anammox.[5][6] Many Planctomycetota occur in relatively high abundance as biofilms,[7] often associating with other organisms such as macroalgae and marine sponges.[8]
Planctomycetota are included in the PVC superphylum along with Verrucomicrobiota, Chlamydiota, Lentisphaerota, Kiritimatiellaeota, and Candidatus Omnitrophica.[9][10] The phylum Planctomycetota is composed of the classes Planctomycetia and Phycisphaerae. First described in 1924, members of the Planctomycetota were identified as eukaryotes and were only later described as bacteria in 1972.[5] Early examination of members of the Planctomycetota suggested a cell plan differing considerably from other bacteria, although they are now confirmed as Gram-negative bacteria, but with many unique characteristics.
Bacteria in the Planctomycetota are often small, spherical cells, but a large amount of morphological variation is seen.[11] Members of the Planctomycetota also display distinct reproductive habits, with many species dividing by budding, in contrast to all other free-living bacteria, which divide by binary fission.[5][12][13]
Interest is growing in the Planctomycetota regarding biotechnology and human applications, mainly as a source of bioactive molecules.[14] In addition, some Planctomycetota were recently described as human pathogens.[8]
The species Gemmata obscuriglobus has been identified specifically as comprising bacteria with unique characteristics among the Planctomycetota,[15][16] such as their ability to synthesize sterols.[5][17][15]

The distinct morphological characteristics of bacteria in the Planctomycetota have been discussed extensively.[6] The common morphology is often spherical cells roughly 2 μm in diameter, as observed in the species Aquisphaera giovannonii. However, the diversity in cell shape often varies greatly in them. Ovoid and pear-shaped cells have been described in some species, and often occur in rosettes of three to 10 cells.[11] Gemmata obscuriglobus is a well studied species in the Planctomycetota with spherical cells. In contrast, bacteria in the species Planctopirus limnophila have ovoid cells.[15]
Many Planctomycetota species display structures and appendages on the outer surface of the cell. Flagella, common in most bacteria, have also been observed in the species P. limnophila.[5][11][18] Many Planctomycetota also have a holdfast, or stalk, which attaches the cell to a surface or substrate.[5][18] Members of some species, though, such as Isosphaera pallida lack a holdfast.[5]
Unique appendages known as crateriform structures have been observed[5][11][18] in species of Planctomycetota belonging to the class Planctomycetia.[13] The outer surface of cells in the species P. limnophila display both large and small crateriform structures. Large crateriform structures often cover the cell surface, while small crateriform structures are often only at the end of the cell. Light microscopy demonstrated fibers of both stalk and pili type in P. limnophila and G. obscuriglobus. The pili fibers in both these species were often associated with large crateriform structures; in contrast, the stalk fibers were associated with small crateriform structures.[18]
Early examination of the Planctomycetota suggested that their cell plan differed considerably from both Gram-positive and Gram-negative bacteria.[5] Until recently, bacteria in the Planctomycetota were thought to lack peptidoglycans in their cell walls, and were instead suggested to have proteinaceous cell walls. Peptidoglycan is an essential polymer of glycans, present in all free-living bacteria, and its rigidity helps maintain integrity of the cell. Peptidoglycan synthesis is also essential during cell division. Recently, those in the species G. obscuriglobus were found to have peptidoglycan in their cell walls.[5][18]
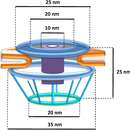
Planctomycetota were once thought to display distinct compartmentalization within the cytosol.[5][18] Three-dimensional electron tomography reconstruction of G. obscuriglobus displayed varying interpretations of this suggested compartmentalization.[16] The cytosol was suggested to be separated into compartments, both the paryphoplasm and pirellulosome, by an intracytoplasmic membrane. This interpretation has since been demonstrated to be incorrect. In fact, the intracytoplasmic membrane is well known to be the cytoplasmic membrane which displays unique invaginations, giving the appearance of compartmentalization within the cytosol.[5][16][18] Planctomycetota therefore display the two compartments typical of Gram-negative bacteria, the cytoplasm and periplasm.
The excess membrane observed in G. obscuriglobus triples the surface area of the cell relative to its volume, which is suggested to be associated with sterol synthesis.[16]
Many Planctomycetota species display pink or orange coloring, suggested to result from the production of carotenoid pigments. Carotenoids are produced by plants and fungi, and by some heterotrophic bacteria to protect against oxidative stress. Three different carotenoid pigments have been identified in two different strains of the Planctomycetota.[19]
In marine environments, Planctomycetota are often suspended in the water column or present as biofilms on the surface of macroalgae, and are often exposed to harmful ultraviolet radiation. More highly pigmented species of the Planctomycetota are more resistant to ultraviolet radiation, although this is not yet well understood.[20]
Bacteria in the Planctomycetota that are anammox-capable form the order Brocadiales.[21] The cells of anammox bacteria are often coccoid with a diameter of about 0.8 μm,[7] and are suggested to contain three compartments, each surrounded by a membrane. The outer membrane encloses the cell and the protoplasm and the innermost membrane surrounds the anammoxosome, the central structure of anammox bacteria.[18][22] The anammoxosome membrane is largely composed of unusual ladderane-based lipids.[22]
Planctomycetota species grow slowly, when compared to other bacteria,[5][10][7][23] often forming rosette structures of 3-5 cells.[5][23] The species P. limnophila is suggested to be relatively fast growing,[5][24] with a doubling time of roughly 6-14 days. In contrast, some other Planctomycetota have doubling times of around 30 days.[24] Their high abundance in many ecosystems is surprising, given their slow growth rates.[7][10]
_and_in_current_PVC_species_(Right).jpg)
Planctomycetota often perform a lifestyle switch between both a sessile stalked stage and a free-swimming stage.[23] Members of the species P. limnophila perform a lifestyle switch that is often associated with cell division. The sessile mother cell produces a free-swimming daughter cell. The daughter cell must then attach to a surface before starting the cycle over again. However, not all of the Planctomycetota have a motile stage, and the lifestyle switch observed in many species may not be common among all Planctomycetota.[5]
.jpg)
The current understanding of bacterial cell division is based on model organisms such as Escherichia coli.[15] The dominant form of reproduction observed in almost all bacteria is cell division by binary fission, which involves the synthesis of both peptidoglycans and proteins known as FtsZ.[15][25] In contrast, many bacteria in the Planctomycetota divide by budding.[5][12][13]
FtsZ proteins are suggested to be similar in structure to that of tubulin, the protein present in eukaryotes,[26] and is essential for septal formation during cell division.[5][6] The lack of FtsZ proteins is often lethal.[5] Peptidoglycan also play a considerable role in cell division by binary fission.[25]
Planctomycetota is one of the only known phyla whose members lack FtsZ proteins.[5][25][26] Bacteria in the Chlamydiales, also a member of the PVC superphylum, also lack FtsZ.[26] Although bacteria in the Planctomycetota lack FtsZ, two distinct modes of cell division have been observed.[5] Most Planctomycetota divide by binary fission, mainly species of the class Phycisphaerae. In contrast, species of the class Planctomycetia divide by budding.[5][12][13]
The mechanisms involved in budding have been described extensively for yeast cells. However, bacterial budding observed in Planctomycetota is still poorly understood.[15] Budding has been observed in both radial symmetric cells, such as bacteria in the species P. limnophila, and axially symmetric cells.[13] During cell division in members of P. limnophila, the daughter cells originate from the region opposite to the pole with the holdfast or stalk.
Considerable diversity has been observed in cell division among bacteria in the Planctomycetota.[12][13] During cell division in Fuerstia marisgermanicae, a tubular structure is connected from the bud to the mother cell.[5][21] The species Kolteria novifilia forms a distinct clade of Planctomycetota, and is the only known species to divide by lateral budding at the middle of the cell. Lastly, members of the clade Saltatorellus are capable of switching between both binary fission and budding.[12][13]
Planctomycetota are known for their unusual cellular characteristics, and their distinctness from all other bacteria is additionally supported by the shared presence of two conserved signature indels (CSIs).[27] These CSIs demarcate the group from neighboring phyla within the PVC group.[28] An additional CSI has been found that is shared by all Planctomycetota species, with the exception of Kuenenia stuttgartiensis. This supports the idea that K. stuttgartiensis forms a deep branch within the Planctomycetota phylum.
A CSI has also been found to be shared by the entire PVC superphylum, including the Planctomycetota.[27][28] Planctomycetota also contain an important conserved signature protein that has been characterized to play an important housekeeping function that is exclusive to members belonging to the PVC superphylum.[29]
The genome size of Rhodopirellula baltica has been estimated to be over 7 million bases, making it one of the largest prokaryotic genomes sequenced. Extensive genome duplication takes up about 25% of the genome sequence.[6] This may be a way for the organism to adapt to mutations, allowing for redundancy if a part of the genome is damaged. The polymerase chain reaction primer used often mismatches with the genes, creating difficulty when sequencing the genome.[9]
When comparing under a microscope, a defining characteristic for some Planctomycetota is that a single unlinked rRNA operon can be identified near the origin. The changes of genetic material is through internal chromosomal inversion, and not through lateral gene transfer. This creates a way of diversification in the Planctomycetota variants as multiple transposon genes in these regions have reverse orientation that transfers to rearrangements.
Some Planctomycetota thrive in regions containing highly concentrated nitrate,[6] and have genes that are required for heterotactic acid fermentation. The enzyme lactate dehydrogenase plays a key role in this process. The genetic process also has ultraviolet radiation protection response, and is associated with the genes recA, lexA, uvrA, uvrB, and uvrC, in addition to a photolyase gene that is expressed when the environment offers excessive ultraviolet radiation stress. Other stress responses include the decomposition of hydrogen peroxide and oxidation.
Many Planctomycetota also express sulfatase genes. The genome of Pirellula sp. strain 1 incorporates 110 genes that contribute to encoding proteins that produce sulfatase enzymes. In comparison with a different species of prokaryotic, Pseudomonas aeruginosa, only 6 sulfatases occur and the genes that express these proteins are contained as two to five pairs, usually clustered in 22 groups.[6]
Planctomycetota originate from within the Bacteria and these similarities between proteins in Planctomycetales and eukaryotes reflect convergent evolution. Gained protein families in Gemmataceae, a subgroup within Planctomycetota, have low sequence similarity to eukaryotic proteins; however, they show highest sequence similarity to other Gemmataceae protein families.[30]
There is massive emergence of novel protein families within the Gemmataceae. More than one thousand protein families were acquired by duplications and domain rearrangements. The new paralogs function in signal transduction, regulatory systems, and protein interaction pathways. They are related to the functional organisation of the cell, which can be interpreted as an adaptation to a more complex lifestyle.[30] The protein length is longer in the Gemmataceae than in most other bacteria and the genes have linkers. There is an overlap between the longest proteins in Planctomycetales and the shortest proteins in eukaryotes. In the terms of gene paralogy, protein length, and protein domain structures, prokaryotes and eukaryotes do not have sharp boundaries.[30]
Originally classified as a eukaryote due to morphology, the advent of genetic sequencing allowed researchers to agree that the Planctomycetota belong to the domain Bacteria.[5] Within that domain, Planctomycetota are classified as their own phylum, however, other researchers have argued they could also be categorized as part of a larger superphylum entitled PVC, which would encompass the phyla Verrucomicrobia, Chlamydiae and Lentisphaerae, and the candidate phylum "Candidatus Omnitrophica".[9] Within this superphylum, its members have been found to be closely related through the creation of 16S rRNA trees. Both the Planctomycetota and Chlamydiota encode proteins for nucleotide transporters, and the Verrucomicrobiota have also been found to have features common among eukaryotic cells. Thus, a common ancestor of this superphylum may have been the start of the eukaryotic lineage.[9] While this is one possible explanation, because PVC is not the start of the bacterial tree,[31] the existence of eukaryotic traits and genes is more likely explained through lateral gene transfer, and not a more recent eukaryotic ancestor.[9]
Members of the Planctomycetota are found in a diverse range of environments, both geographically and ecologically,[38] and occur in both aquatic and terrestrial habitats.[5] In aquatic environments, they are found in both freshwater and marine systems.[38] Planctomycetota were originally believed to exist exclusively in aquatic environments, but they are now known to be also abundant in soils[39] and hypersaline environments.[40] They are widespread on five continents, including Antarctica and Australia.[39][38]
Fluorescence in situ hybridization was used to detect Planctomycetota in various environments, and Planctomycetota are found in abundance in sphagnum bogs. Some Planctomycetota were found in the digestive systems of marine lifeforms, while others tend to live among eukaryotes.[9]
Planctomycetota account for roughly 11% of prokaryotic communities in marine systems, and their vast distribution demonstrates their ability to inhabit many different environments. They can also adapt to both aerobic and anaerobic conditions. Many factors can affect their distribution, such as humidity, oxygen levels, and pH levels. Planctomycetota diversity and abundance are strongly associated with relative humidity. The effects of oxygen levels demonstrate the energy needs of the individual. Many species of Planctomycetota are chemoheterotrophic, including G. obscuriglobus. Thermostilla marina, a thermophilic anaerobic species occupying hydrothermal vent regions, can use elemental sulfur to generate sulfide and respire with nitrate. Planctomycetota can also inhabit regions with ranges in pH levels from 4.2 to 11.6.[8]
Planctomycetota have a significant impact on global biogeochemistry and climate, with their ability to mineralize and break down detritus particles in the water column.[6][20]
Planctomycetota play a considerable role in the global carbon cycle.[5][6][13][41] As both obligate and facultative aerobic chemoheterotrophs, the primary source of carbon used by Planctomycetota is from carbohydrates. Many Planctomycetota have the ability to breakdown extremely complex carbohydrates, making these nutrients available to other organisms. This ability to recycle carbon has been linked to specific C1 metabolism genes observed in many Planctomycetota and are suggested to play a significant role, but this area of research is still poorly understood.
Planctomycetota also display many sulfatase enzymes, which are capable of breaking down sulfated heteropolysaccharides, which are produced by many groups of macroalgae. The breakdown of these sulfated heteropolysaccharides by Planctomycetota are then used as an energy source. Some Planctomycetota are suggested to be capable of breaking down carrageenan.[41]

Planctomycetota have often been observed in association with many organisms, including, macroalgae, microalgae, marine sponges, and plants such as lichens and bryophytes.[8] They have also been observed inhabiting deep-sea cold seeps, where they are dominant organisms living on tube worms.[5]

Planctomycetota are often associated with marine surfaces high in nutrients. They occur as biofilms on algal surfaces in relatively high abundance.[7] Macroalgae such as the kelps Laminaria hyperborea and Ecklonia radiata are suggested to be an important habitat for Planctomycetota.[5][42] Roughly 70% of the bacterial community on Ecklonia radiata were Planctomycetota.[5][10] Almost 150 Planctomycetota species have been isolated from the biofilms of macroalgae, and these communities associated with macroalgae are mainly independent of changes in geographical distribution. This would suggest a symbiotic relationship.[8]
Kelp forests dominate the rocky coastlines of temperature regions, and provide habitat, shelter, and food for many organisms, including the Planctomycetota.[5] Given the considerable role of kelp forests in coastal primary productivity, the association of the Planctomycetota with kelp could indicate their significant role in coastal habitats.[43] Planctomycetota also play an important role as components of detritus in the water column, also known as marine snow,[5][43] given their ability to attach to surfaces.[44]
As the climate continues to warm, the abundance of Planctomycetota associated with macroalgae might increase. The seaweed Caulerpa taxifolia was incubated under higher CO2 conditions, and the abundance of Planctomycetota increased substantially, as much as 10 times in some species.[5]
While macroalgae are well known substrates for Planctomycetota communities, their abundance has also been known to correlate with blooms of microalgae such as diatoms.[43][5] Blooms of cyanobacteria, diatoms, and dinoflagellates provide nutrients for Planctomycetota, which could explain the association.[8]
Planctomycetota species are often associated with the surfaces of marine sponges.[8][44] They interact with sponges either by attachment with a holdfast, or through a symbiotic relationship. A high diversity of Planctomycetota is present as biofilms on sponges. The symbiotic relationship among sponges and Planctomycetota contributes to the health of the sponge, and the sponge often provides suitable habitat and nutrients to the Planctomycetota.[8]
Planctomycetota were found to be highly abundant in lichen communities throughout northwestern Siberia and displayed extremely high diversity. Planctomycetota have also been associated with lichen communities and Sphagnum wetlands. Sphagnum wetlands store large amounts of carbon, contributing to the global carbon cycle. Planctomycetota play a considerable role in the degradation of sphagnum, accounting for roughly 15% of the bacterial community.[8]
Planctomycetota display associations with other bacterial communities, mainly Alphaproteobacteria, Bacteroidota, Gemmatimonadota, and Verrucomicrobiota. The growth of many Planctomycetota is often supported by the essential nutrients provided by other bacteria within the community, and some Planctomycetota rely strongly on symbiotic relationships with other bacteria.[8]
The existence of membrane coat proteins near the intracytoplasmic membrane could be used for an endocytosis-like uptake system, which would be the first instance this function has been found outside of the eukaryotic domain. However, now that the existence of a rigid peptidoglycan cell wall has been confirmed, these vesicles to be able to pass through this cell wall seems unlikely. Additionally, deletion of one of these membrane coat proteins within P. limnophila found no decrease in macromolecule uptake.[18] In addition, with the use of cryoelectron tomography-based three-dimensional reconstruction of Planctomycetota has found that what were originally thought to be vesicles being held in the periplasm are actually just folds in the cytoplasmic membrane.[5] Yet it has been demonstrated that the Planctomycetota can survive on high-molecular-weight polysaccharides as their only source of carbon, meaningthey must have the ability to incorporate complex carbon substrates into their cytoplasm. Three hypotheses have been put forth: First, the Planctomycetota excrete an enzyme which, outside of the cell wall, degrades the complex substrates into smaller monosaccharides, which can more easily be transported through the different membranes. Second, the complex substrates become anchored to the outside of Planctomycetota, which are then able to slowly break down these substrates into oligosaccharides, which are able to be transported into the periplasm of Planctomycetota by specialized proteins. The third hypothesis involves the crateriform structures found on the outside of Planctomycetota cell walls. These structures have fibers lining their pits that may be able to absorb whole polysaccharides into the periplasm, where they would then be digested.[18]
Almost all bacteria have a cytosol following the outer shape of their peptidoglycan cell wall. Eukaryotes are different in that they have their cytosol divided into multiple compartments to create organelles such as a nucleus. Planctomycetota are unique in that they have large invaginations of their cytoplasmic membrane, pulling away from the peptidoglycan cell wall and leaving room for the periplasm. Traditionally, the cytoplasmic membrane has been thought to be responsible for controlling the osmotic pressure of bacterial cells. Yet due to the folds in the cytoplasmic membrane, and the existence of large spaces of periplasm within Planctomycetota, their peptidoglycan acts as an osmotic barrier with the periplasm being isotonic to the cytosol.[5]
Anammox is the process of oxidizing ammonium where nitrite acts as the electron acceptor. This process creates energy for the organism performing the reaction in the same way humans gain energy from oxidizing glucose.[45] In a marine environment, this ultimately removes nitrogen from the water, as N2 gas cannot be used by phytoplankton and is released into the atmosphere. Up to 67% of dinitrogen gas production in the ocean can be attributed to anammox[46] and about 50% of the nitrogen gas in the atmosphere is thought to be produced from anammox.[47] Planctomycetota are the most dominant phylum of bacteria capable of performing anammox, thus the Planctomycetota capable of performing anammox play an important role in the global cycling of nitrogen.[48]
The synthesis of sterols, often observed in eukaryotes and uncommon among bacteria, has been observed very rarely in Planctomycetota.[5][15] The synthesis of sterols such as lanosterol has been observed in G. obscuriglobus. Lanosterol is common in eukaryotes and two other groups of bacteria, both methylotrophic Pseudomonadota and myxobacteria. The synthesis of sterols observed in G. obscuriglobus is unique within Planctomycetota. Sterol synthesis is suggested to be associated with regulation of membrane fluidity in Planctomycetota,[15] and has been described as essential to the proper growth and reproduction of G. obscuriglobus.[17]
Recently, interest has arisen in examining the Planctomycetota regarding their potential roles in biotechnology, mainly as a source of bioactive molecules,[8][14] of interest mainly to the pharmaceutical industry. Bioactive compounds are mainly present as secondary metabolites,[14] although little is known about Planctomycetota secondary metabolites.[49] This is unexpected, as the Planctomycetota have several key features as other known producers of bioactive molecules, such as the Myxobacteria.[49] However, a number of ongoing studies serve as various first steps in including Planctomycetota in small-molecule drug development for humans.
Planctomycetota species are worthwhile considerations in challenging the current models for the origin of the nucleus, along with other aspects of origin and evolution of the eukaryotic endomembrane system.[40]
The impacts of research on Planctomycetota and their uses might be of global significance with regards to nutrient cycling processes and assist in furthering understanding for global marine biogeochemistry. However, with Planctomycetota's growing influences on metabolic processes involving water and air, it may also have a role in interchanges between oceans and atmosphere, potentially affecting climate change.[40]
Planctomycetota species were recently identified as being an opportunistic human pathogen, but a lack of culture media limits studies on the bacteria in the Planctomycetota as pathogens of humans.[8]
The Planctomycetota are a phylum of widely distributed bacteria, occurring in both aquatic and terrestrial habitats. They play a considerable role in global carbon and nitrogen cycles, with many species of this phylum capable of anaerobic ammonium oxidation, also known as anammox. Many Planctomycetota occur in relatively high abundance as biofilms, often associating with other organisms such as macroalgae and marine sponges.
Planctomycetota are included in the PVC superphylum along with Verrucomicrobiota, Chlamydiota, Lentisphaerota, Kiritimatiellaeota, and Candidatus Omnitrophica. The phylum Planctomycetota is composed of the classes Planctomycetia and Phycisphaerae. First described in 1924, members of the Planctomycetota were identified as eukaryotes and were only later described as bacteria in 1972. Early examination of members of the Planctomycetota suggested a cell plan differing considerably from other bacteria, although they are now confirmed as Gram-negative bacteria, but with many unique characteristics.
Bacteria in the Planctomycetota are often small, spherical cells, but a large amount of morphological variation is seen. Members of the Planctomycetota also display distinct reproductive habits, with many species dividing by budding, in contrast to all other free-living bacteria, which divide by binary fission.
Interest is growing in the Planctomycetota regarding biotechnology and human applications, mainly as a source of bioactive molecules. In addition, some Planctomycetota were recently described as human pathogens.
The species Gemmata obscuriglobus has been identified specifically as comprising bacteria with unique characteristics among the Planctomycetota, such as their ability to synthesize sterols.
Los planctomicetos (Planctomycetota, anteriormente Planctomycetes) son un filo de bacterias Gram negativas acuáticas, flageladas con al menos una fase móvil, encontradas en agua dulce, salobre y marina. Tienen forma ovoide o esférica debido a la ausencia de pared celular.
Los organismos pertenecientes a este grupo carecen de mureína en su pared celular. La mureína es un importante heteropolímero presente en la mayoría de las paredes celulares bacterianas que sirve como componente protector en el esqueleto de la pared celular. En su lugar, las paredes se componen de una glicoproteína rica en glutamato.
Los Planctomycetota tienen estructuras internas más complejas que las encontradas en el resto de los procariontes típicos. Aunque no tienen núcleo en el sentido eucariota clásico, el material nuclear está contenido a veces dentro de una doble membrana. Además del compartimento del cuerpo nuclear que contiene el ADN, hay otros dos compartimientos separados por membranas: uno es el riboplasma (o pirelulosoma) que contiene ribosomas con las proteínas asociadas y el otro es el parifoplasma que no contiene ribosomas (Glockner, 2003).[1]
La secuenciación del ARN muestra que las relaciones de los Planctomycetota con las demás bacterias son distantes. Algunas cadenas esenciales no se organizan en operones, lo que es inusual para una bacteria (Glockner, 2003). A través de comparaciones de secuencias, se han descubierto algunos genes que son similares a los que poseen los eucariones. Un ejemplo de ello es la significativa homología de una secuencia genética de Gemmata obscuriglobus con la integrina alfa-V, una proteína importante para la transdución de señales transmembranosas en los eucariontes (Jenkins y otros, 2002).
Cavalier-Smith considera que Planctomycetota puede incluirse en el clado Planctobacteria y este a su vez en el clado Gracilicutes.
Los planctomicetos (Planctomycetota, anteriormente Planctomycetes) son un filo de bacterias Gram negativas acuáticas, flageladas con al menos una fase móvil, encontradas en agua dulce, salobre y marina. Tienen forma ovoide o esférica debido a la ausencia de pared celular.
Les planctomycètes (ou Planctomycetes) forment un phylum de bactéries aquatiques, qui se trouvent dans des terrains saumâtres, marins ou d'eau douce, mais aussi de bactéries terrestres qui jouent un rôle important dans le processus anammox. Ils se reproduisent par bourgeonnement. La structure des organismes de ce groupe est ovoïde avec une tige, à la terminaison non-reproductive, qui les aide à se joindre les uns aux autres au cours du bourgeonnement.
Cavalier-Smith a postulé que le groupe des Planctomycetes est dans le clade Planctobacteria, dans les grandes clade Gracilicutes.
Les organismes appartenant à ce groupe n'ont pas de maillage en peptidoglycane dans leur paroi cellulaire. Les peptidoglycane sont des hétéropolymères importants présent dans la plupart des parois des cellules bactériennes, qui constitue un élément de protection dans la paroi squelettique. Au lieu de cela, ils sont constitués de glycoprotéines riches en glutamate. Les planctomycètes ont des structures internes plus complexes que ce qu'on attend généralement chez les procaryotes. Même si elles n'ont pas un noyau dans le sens eucaryote, le matériel nucléaire peut parfois être enfermé dans une double membrane. En plus de ce nucléoïde, il existe deux autres membrane séparées : le riboplasme (ou pirellulosome) qui contient le ribosome et les protéines, et le paryphoplasme qui ne contient pas de ribosomes (Glockner, 2003).
Le séquençage de l'ARN ribosomique montre que les Planctomycètes sont assez éloignés des autres eubacteria. Les groupes les plus proches semblent être les Verrucomicrobia et Chlamydiae[1]. Un certain nombre de gènes impliqués dans des voies essentielles ne sont pas organisés en opéron, ce qui est assez inhabituel chez les bactéries (Glöckner, 2003). Un certain nombre de gènes semblables à des gènes eucaryotes ont été mis en évidence à l'aide de comparaison de séquences. Par exemple un gène de Gemmata obscuriglobus présente une similarité significative par rapport au gène de l'intégrine alpha-V, qui est une protéine qui joue un rôle important dans la transduction du signal transmembranaire chez les eucaryotes (Jenkins et al., 2002).
Le cycle de vie est constitué de deux phases où alternent une cellule sessile (immobile, accrochée à une paroi) et une cellule à flagelle. Les cellules sessiles bourgeonnent pour former des cellules à flagelle, qui iront nager un certain temps avant de se rattacher à leur tour à une paroi et commencer leur reproduction.
Selon NCBI (10 janv. 2011)[2] :
Les planctomycètes (ou Planctomycetes) forment un phylum de bactéries aquatiques, qui se trouvent dans des terrains saumâtres, marins ou d'eau douce, mais aussi de bactéries terrestres qui jouent un rôle important dans le processus anammox. Ils se reproduisent par bourgeonnement. La structure des organismes de ce groupe est ovoïde avec une tige, à la terminaison non-reproductive, qui les aide à se joindre les uns aux autres au cours du bourgeonnement.
Cavalier-Smith a postulé que le groupe des Planctomycetes est dans le clade Planctobacteria, dans les grandes clade Gracilicutes.
Os planctomicetos (Planctomycetes) son un filo de bacterias esencialmente acuáticas, que se atopan en mostras de augas mariñas, salobres e doces, ou en chans. Reprodúcense por xemación. En estrutura, son organismos de forma oval e teñen unha especie de prolongación fina chamada talo ou pedúnculo situado no seu extremo non reprodutor, que lles axuda a fixarse durante a reprodución. Estes filamentos que forman parécense aos que forman os fungos e por esta característica foron nomeados: Planctomycetes = "fungos flotantes" (pero non son fungos).
Cavalier-Smith postulou que os Planctomycetes estaban dentro do clado das Planctobacteria do gran clado Gracilicutes.
Os organismos que pertencen a este grupo carecen de peptidoglicano nas súas paredes celulares. O peptidoglicano é un importante heteropolímero presente na maioría das paredes celulares das bacterias que serve como un compoñente básico da súa estrutura. En Planctomycetes as paredes están feitas de glicoproteínas ricas en glutamato, polo que son máis ben do tipo das capas S. Como non teñen peptidoglicano son insensibles á penicilina. Teñen apéndices con forma de talo (prostecas), que serven para a fixación.
Os Planctomycetes teñen estruturas internas que son máis complexas do que normalmente se agardaría nos procariotas. Algúns planctomicetos teñen membranas internas. Presentan dous compartimentos separados por membranas, que son o pirelulosoma ou riboplasma, que contén os ribosomas e proteínas relacionadas, e por fóra del está o parifoplasma sen ribosomas en situación periférica. Carecen de núcleo no sentido eucariota, pero o material xenético (zona do nucleoide) pode ás veces, como ocorre en Gemmata, estar rodeado por unha dobre membrana especial, e toda a estrutura está dentro do compartimento do pirelulosoma ou riboplasma.[1] Algúns planctomicetos que oxidan amoníaco en anaerobiose aínda teñen outro compartimento máis que contén encimas, o anamoxosoma. Só o grupo dos Epixenosomas, un grupo de bacterias relacionadas con Verrucomicrobia, teñen unha estrutura máis complexa entre as bacterias.
Este compartimento nuclear dalgúns planctomicetos apoia a teoría da orixe autóxena do núcleo eucariota (orixinado por membranas internas), fronte á teoría endosimbiótica (formado pola endosimbiose con outra célula), aínda que isto non quere dicir que os planctomicetos estean emparentados cos eucariotas.
O material xenético do nucleoide está máis concentrado ca o doutras bacterias, que xeralmente está moi espallado por todo o centro da célula.
Recentemente, viuse que a especie de Planctomycetes Gemmata obscuriglobus pode captar grandes moléculas do exterior por un proceso que lembra á endocitose que presentan os eucariotas.[2][3]
A secuenciación do ARN mostra que os planctomicetos están relacionados cos Verrucomicrobia e posiblemente con Chlamydiae.[4] Varias vías metabólicas esenciais non están reguladas por operóns, o cal é pouco usual en bacterias.[1] Atopáronse varios xenes por comparación das secuencias, que son similares a xenes atopados en eucariotas. Un exemplo é unha secuencia xénica de Gemmata obscuriglobus, que ten unha homoloxía significativa coa do xene da integrina alfa-V, unha proteína que é importante na transdución de sinais transmembrana en eucariotas.[5]
O ciclo biolóxico de vida de moitos planctomicetos presenta alternancia entre unha fase de células sésiles e outra de células flaxeladas nadadoras. As células sésiles experimentan xemación formando as células flaxeladas, que nadan durante un tempo antes de que se asentan e se unen para empezar a reprodución.
No cladograma móstrase a taxonomía actualmente aceptada deste grupo baseada na LPSN (List of Prokaryotic names with Standing in Nomenclature) [6] e a filoxenia baseada no ARNr 16S da LTP 106 do Proxecto The All-Species Living Tree.[7]
Phycisphaera mikrensis Fukunaga et al. 2010
Planctomycetales?Candidatus Nostocoida limicola III
Brocadiaceae Candidatus Scalindua?Candidatus S. marina
?Candidatus S. arabica Woebken et al. 2008
Candidatus S. wagneri Schmid et al. 2003
Candidatus S. sorokinii Kuypers et al. 2003
Candidatus S. brodae Schmid et al. 2003
Candidatus Kuenenia stuttgartiensis Schmid et al. 2000
Candidatus BrocadiaCandidatus B. anammoxidans Jetten et al. 2001
Candidatus B. brasiliensis Araujo et al. 2011
Candidatus B. caroliniensis
Candidatus B. fulgida Kartal et al. 2004
Candidatus B. sinica Hu et al. 2010
Candidatus Anammoxoglobus propionicus Kartal et al. 2006
Candidatus Jettenia asiatica Quan et al. 2008
PlanctomycetaceaeIsosphaera pallida (ex Woronichin 1927) Giovannoni et al. 1995
SingulisphaeraS. acidiphila Kulichevskaya et al. 2008
S. mucilaginosa ♠ Zaicnikova et al. 2011
Gemmata obscuriglobus Franzmann and Skerman 1985
Zavarzinella formosa Kulichevskaya et al. 2009
Pirellula staleyi Schlesner and Hirsch 1987
Blastopirellula marina (Schlesner 1987) Schlesner et al. 2004
Rhodopirellula baltica Schlesner et al. 2004
Planctomyces?P. bekefii ♪ Gimesi 1924
?P. guttaeformis ♪ (ex Hortobágyi 1965) Starr and Schmidt 1984
?P. stranskae ♪ (ex Wawrik 1952) Starr and Schmidt 1984
P. brasiliensis Schlesner 1990
P. maris (ex Bauld and Staley 1976) Bauld and Staley 1980
P. limnophilus Hirsch and Müller 1986
Schlesneria paludicola Kulichevskaya et al. 2007
Notas:
♠ Cepas atopadas no NCBI pero non na LSPN.
♪ Procariotas dos que non existen cultivos puros (axénicos) illados ou dispoñibles, é dicir, que non se puideron cultivar ou que o cultivo non dura moito tempo.
Os planctomicetos (Planctomycetes) son un filo de bacterias esencialmente acuáticas, que se atopan en mostras de augas mariñas, salobres e doces, ou en chans. Reprodúcense por xemación. En estrutura, son organismos de forma oval e teñen unha especie de prolongación fina chamada talo ou pedúnculo situado no seu extremo non reprodutor, que lles axuda a fixarse durante a reprodución. Estes filamentos que forman parécense aos que forman os fungos e por esta característica foron nomeados: Planctomycetes = "fungos flotantes" (pero non son fungos).
Cavalier-Smith postulou que os Planctomycetes estaban dentro do clado das Planctobacteria do gran clado Gracilicutes.
De Planctomycetales zijn een orde van obligaat (verplicht) aerobe bacteriën die voorkomen in brak, zout en zoet water. Het zijn eivormige bacteriën, die een steel aan het niet-reproductieve eind hebben. Ze vermeerderen zich door knopvorming.
Deze bacteriën behoren tot de groep die geen mureïne in hun celwand hebben zitten. Mureïne is een belangrijk copolymeer, dat als een beschermstof in veel celwanden van bacteriën voorkomt. In plaats daarvan bestaan hun celwanden uit glycoproteïne, dat rijk is aan glutamaat. Planctomycetes hebben inwendige structuren die meer complex zijn dan men zou verwachten bij prokaryoten. Ofschoon ze geen celkern, zoals die voorkomt bij eukaryoten, hebben, kan kernmateriaal soms omgeven zijn door een dubbelmembraan. Naast dit nucleoïde zijn er twee andere door membranen omgeven compartimenten; het pirellulosoom of riboplasma, dat het ribosoom bevat met de verwante proteïnen en het ribosoomvrije paryphoplasma (Glockner, 2003).
RNA-sequencing laat zien dat de planctomycetes verre verwanten zijn van de andere eubacteria. Een aantal onontbeerlijke pathways (een reeks van biochemische processen) zijn niet georganiseerd in operons hetgeen voor bacteriën ongewoon is (Glockner, 2003). Een aantal genen zijn gelijk aan die in eukaryoten. Een voorbeeld hiervan is de gensequentie in Gemmata obscuriglobus dat een significante homologie vertoont met het integrine alpha-V, een proteïne dat van belang is voor de transmembrane signaal transductie in eukaryoten (Jenkins et al., 2002).
De levenscyclus bestaat uit een afwisseling van cellen, die zich ergens aan vasthechten, met zwermcellen, die flagellen hebben. De eerstgenoemde cellen vormen door knopvorming de zwermcellen met flagellen. Deze zwermcellen zwemmen een poosje alvorens ze zich vasthechten en zich gaan vermeerderen.
De Planctomycetales zijn een orde van obligaat (verplicht) aerobe bacteriën die voorkomen in brak, zout en zoet water. Het zijn eivormige bacteriën, die een steel aan het niet-reproductieve eind hebben. Ze vermeerderen zich door knopvorming.
Planctomycetes er en serie av vannlevende bakterier og har blitt påvist i brakkvann, sjø- og ferskvannsprøver. De formerer seg ved okulasjon.
Planctomycetes er en serie av vannlevende bakterier og har blitt påvist i brakkvann, sjø- og ferskvannsprøver. De formerer seg ved okulasjon.
Planctomycetes é um filo de bactérias aquáticas encontradas em água doce, salobra e marinha. Têm forma ovóide e reproduzem-se por gemulação. O ciclo biológico implica a alternância entre células sésseis e flageladas. As células sésseis formam células flageladas por gemulação, as quais nadam durante certo tempo antes de estabelecerem-se e começarem a reprodução.
Embora inicialmente se pensasse que os organismos pertencentes a este grupo careciam de mureína na sua parede celular estudos recentes revelam que os Planctomycetes apresentam uma estrutura fina de mureína[1]. A mureína é um importante heteropolímero presente na maioria das paredes celulares bacterianas, que serve como componente protector no esqueleto da parede celular. As paredes são principalmente compostas por uma glicoproteína rica em glutamato. Planctomycetes possui estruturas internas mais complexas que as que existem nos procariontes típicos. Ainda que não possuam núcleo, no sentido eucariota, o material nuclear está contido, por vezes, dentro de una membrana dupla. Para além deste nucleóide, existem dois outros compartimentos separados por membranas: o riboplasma (oo pirelulossoma) que contém ribossomas com as proteínas associadas e o parifoplasma que não contém ribossomas (Glockner, 2003).
A sequenciação do ARN mostra que as relações dos Planctomycetes com as demais bactérias são distantes. Algumas cadeias essenciais não se organizam em operões, o que não é usual para uma bacteria (Glockner, 2003). Através de compações de sequências, descobriram-se alguns genes que são similares aos que possuem os eucariones. Um exemplo disto é a significativa homologia de uma sequência genética de Gemmata obscuriglobus com a integrina alfa-V, uma proteína importante para a transdução de sinais transmembranares nos eucariontes (Jenkins y otros, 2002).
Cavalier-Smith considera que Planctomycetes pode ser incluída no clado Planctobacteria, e este por sua vez no clado Gracilicutes.
Planctomycetes é um filo de bactérias aquáticas encontradas em água doce, salobra e marinha. Têm forma ovóide e reproduzem-se por gemulação. O ciclo biológico implica a alternância entre células sésseis e flageladas. As células sésseis formam células flageladas por gemulação, as quais nadam durante certo tempo antes de estabelecerem-se e começarem a reprodução.
Plancomycetes là nhóm vi khuẩn thuỷ sinh hiếu khí bắt buộc (cần ôxy để phát triển). Nhóm vi khuẩn này sinh sản bằng hình thức nảy chồi. Về mặt cấu trúc, các vi khuẩn trong nhóm này có hình ovan (hình trứng) và có phần chân ống (stalk) nằm ở cực dinh dưỡng (ngược với cực sinh sản) để bám vào giá thể trong quá trình nảy chồi.
Các vi khuẩn thuộc nhóm này không có chất murein trong thành tế bào. Thay vào đó thành tế bào của Plancomycetes được tạo thành bới các phân tử glycoprotein giàu glutamate. Các phân tích RNA ribosome cho thấy Plancomycetes thuộc nhóm vi khuẩn thực (eubacteria) chứ không phải là nấm như phân loại trước kia. Mặc dù vậy tên của nhóm vi khuẩn này vẫn được giữ như trước và có vẻ như làm một nhóm nấm
Nhóm vi khuẩn Planctomycetes lại là vi khuẩn kỵ khí bắt buộc, chúng không thể sống trong môi trường có nhiều oxy tự do. Vi khuẩn anammox được cung cấp bởi công ty Công ty Meidensa, Nagoya, Nhật Bản. Vi khuẩn Planctomycetes dạng hạt màu nâu đỏ, bổ sung từ 20–50 mg hạt, làm nhuyễn thành dung dịch lỏng rồi cho vào bể phản ứng. Bơm lưu lượng sẽ bơm nước thải từ dưới lên đưa vi khuẩn Anammox bám vào vật liệu mang. Sau khi bổ sung vi khuẩn Anammox trên lớp vật liệu mang sẽ hình thành lớp màng sinh học có chứa vi khuẩn Anammox.
Жизненный цикл Planctomycetes состоит из чередования сидячей стадии, прикрепляющейся к субстрату при помощи стебелька, и подвижной стадии, у которой стебелёк является не продолжением клетки, а отдельным белковым придатком[16][17].
Сравнительный анализ нуклеотидных последовательностей 16S рРНК показывает, что тип имеет высокий уровень гомологии с тремя другими типами — Verrucomicrobia, Chlamydiae и Lentisphaerae[en], образуя надтип[18]. Изучение геномов представителей типа Planctomycetes позволяет пролить свет на происхождение метаногенеза и метилотрофии[19]. Уникальной особенностью геномов представителей типа Planctomycetes является отсутствие оперонной структуры генов некоторых важных метаболических путей[en]*[12], что также не характерно для прокариот. Сравнение нуклеотидных последовательностей некоторых генов показывает больший уровень гомологии с таковыми у эукариот: например, один ген Gemmata obscuriglobus проявляет большой уровень гомологии с геном, кодирующим белок интегрин альфа-V, играющим большую роль в трансмембранной передаче сигнала у эукариот[20].
Представители Planctomycetes обитают в пресных и солёных водах[21][22], являются важными членами почвенных сообществ микроорганизмов[23]. Кроме того, они обнаруживаются в торфяных сфагновых болотах[24], обитают также в анаэробных источниках, богатых соединениями серы[25].
Ниже приведена кладограмма Planctomycetes с учётом номенклатуры, рекомендованной List of Prokaryotic names with Standing in Nomenclature[en] (LSPN)[26]. Филогенетические связи определены на основе анализа 16S рРНК, представленного в 111 релизе (2013 год) проекта The All-Species Living Tree[en][27].
Phycisphaera mikrensis Fukunaga et al. 2010
?Candidatus Nostocoida limicola III
?Candidatus S. arabica Woebken et al. 2008
?Candidatus S. marina Van de Vossenberg et al. 2007
?Candidatus S. profunda Van De Vossenberg et al. 2008
?Candidatus S. richardsii Fuchsman et al. 2012
Candidatus S. wagneri[en] Schmid et al. 2003
Candidatus S. sorokinii Kuypers et al. 2003
Candidatus S. brodae[en] Schmid et al. 2003
Candidatus Kuenenia stuttgartiensis Schmid et al. 2000
Candidatus B. anammoxidans Jetten et al. 2001
Candidatus B. brasiliensis Araujo et al. 2011
Candidatus B. caroliniensis
Candidatus B. fulgida[en] Kartal et al. 2004
Candidatus B. sinica Hu et al. 2010
Candidatus Anammoxoglobus propionicus Kartal et al. 2006
Candidatus Jettenia asiatica Quan et al. 2008
?P. bekefii ♪ Gimesi 1924 (type sp.)
?P. guttaeformis ♪ (ex Hortobágyi 1965) Starr and Schmidt 1984
?P. stranskae ♪ (ex Wawrik 1952) Starr and Schmidt 1984
P. brasiliensis Schlesner 1990
P. maris (ex Bauld and Staley 1976) Bauld and Staley 1980
P. limnophilus Hirsch and Müller 1986
Schlesneria paludicola Kulichevskaya et al. 2007
R. baltica Schlesner et al. 2004 (type sp.)
R. europaea ♠ Frank 2011
R. maiorica ♠ Frank 2011
R. sallentina ♠ Frank 2011
Blastopirellula marina (Schlesner 1987) Schlesner et al. 2004
Pirellula staleyi Schlesner and Hirsch 1987
Gemmata obscuriglobus[en] Franzmann and Skerman 1985
Telmatocola sphagniphila Kulichevskaya et al. 2012
Zavarzinella formosa Kulichevskaya et al. 2009
Isosphaera pallida (ex Woronichin 1927) Giovannoni et al. 1995
Aquisphaera giovannonii Bondoso et al. 2011
?S. mucilaginosa ♠ Zaicnikova et al. 2011
S. acidiphila Kulichevskaya et al. 2008 (type sp.)
S. rosea Kulichevskaya et al. 2012
Обозначения:
♠ Штаммы, имеющиеся в базе данных Национального центра биотехнологической информации (NCBI), но отсутствующие в базе данных LSPN
♪ Прокариоты, для которых не существует чистых культур, то есть некультивируемые штаммы или штаммы, культуры которых нестойки.
Жизненный цикл Planctomycetes состоит из чередования сидячей стадии, прикрепляющейся к субстрату при помощи стебелька, и подвижной стадии, у которой стебелёк является не продолжением клетки, а отдельным белковым придатком.
浮黴菌門(Planctomycetes)是一小門水生細菌,在海水、半咸水、淡水中都可被發現。
其中浮黴菌屬(Planctomyces)和小梨形菌屬(Pirellula)等都是專性好氧菌。它們通過出芽法繁殖。形態上,它們通常是卵形,不用來繁殖的一端有柄,可以用來附着。它們的生活史分爲固着細胞和有鞭毛的游動細胞,類似α-變形菌綱的柄桿菌屬。
浮黴菌門的細胞壁中含有糖蛋白而不含胞壁質,因此它們可以通過青黴素等破壞細胞壁的抗生素來選擇性富集。最爲奇特的一點是,浮黴菌門細胞具有複雜的胞内膜結構,甚至有些屬(如出芽菌屬(Gemmata))的染色質被膜包圍且緊縮,類似真核生物的細胞核,這在原核生物中是僅有的。
此外,在浮黴菌門中還有一類和浮黴菌屬等關係較遠的細菌,如(Candidatus Brocadia)、(Candidatus Kuenenia)和(Candidatus Scalindua)屬,它們至今未能成功分離得到純菌株,因此尚未獲得正式命名和分類。它們能夠在缺氧環境下利用亞硝酸鹽(NO2-)氧化銨離子(NH4+)生成氮氣來獲得能量,因此稱作厭氧氨氧化菌(anaerobic ammonium oxidation, Anammox),對全球氮循環具有重要意義,也是污水處理中重要的細菌。
浮黴菌門(Planctomycetes)是一小門水生細菌,在海水、半咸水、淡水中都可被發現。
其中浮黴菌屬(Planctomyces)和小梨形菌屬(Pirellula)等都是專性好氧菌。它們通過出芽法繁殖。形態上,它們通常是卵形,不用來繁殖的一端有柄,可以用來附着。它們的生活史分爲固着細胞和有鞭毛的游動細胞,類似α-變形菌綱的柄桿菌屬。
浮黴菌門的細胞壁中含有糖蛋白而不含胞壁質,因此它們可以通過青黴素等破壞細胞壁的抗生素來選擇性富集。最爲奇特的一點是,浮黴菌門細胞具有複雜的胞内膜結構,甚至有些屬(如出芽菌屬(Gemmata))的染色質被膜包圍且緊縮,類似真核生物的細胞核,這在原核生物中是僅有的。
此外,在浮黴菌門中還有一類和浮黴菌屬等關係較遠的細菌,如(Candidatus Brocadia)、(Candidatus Kuenenia)和(Candidatus Scalindua)屬,它們至今未能成功分離得到純菌株,因此尚未獲得正式命名和分類。它們能夠在缺氧環境下利用亞硝酸鹽(NO2-)氧化銨離子(NH4+)生成氮氣來獲得能量,因此稱作厭氧氨氧化菌(anaerobic ammonium oxidation, Anammox),對全球氮循環具有重要意義,也是污水處理中重要的細菌。
プランクトミケス門 (Planctomycetes、プランクトマイセス門) は、グラム陰性細菌の小型の門で、幾つかの水生従属栄養細菌を含む。この門には2綱2門2科12属17種が属している。系統的にはウェルコミクロビウムやクラミジア(他にレンティスパエラ)に比較的近縁だが、他の細菌群とは離れている。分類体系によっては、この3系統を合わせPVC群(あるいはプランクトバクテリア)を設置する場合がある[1]。 タイプ属のPlanctomycesは、ギリシャ語のπλαγκτός(プランクトス/漂う)+μύκες(ミュケス/菌)をラテン語化したもので、「(水中で)浮遊する菌」との意を持つ。
原核生物の中では最も複雑な構造と生活環を持つグループの1つである。形状は大まかに卵形であるが、柄を持つなど変わったところがあり、出芽によって増殖する点でも特異である。細胞壁は通常の細菌と異なりペプチドグリカンを含まないと考えられてきたが、最近になってプランクトミケス門細菌の細胞壁にもペプチドグリカンが存在することが確認された[2]。また、細胞内に核膜のような構造を形成することでも知られる。原核生物においてこの構造は、プランクトミケスを含むPVC系統と、古細菌であるIgnicoccusにしか発見されていない。Gemmata obscuriglobusなどでは特に顕著で、細胞壁エンドサイトーシスに似たタンパク質の取り込み機構が報告されている[3]。
これまでに知られている種は殆どが好気性の従属栄養生物であるが、嫌気的アンモニア酸化反応(Anammox)を行う系統も存在する。この細菌類は亜硝酸を電子受容体としてアンモニアを窒素に酸化する特異な代謝系を備えている。これらは何れも未記載であるが、暫定的な系統分類としてBrocadia, Kuenenia, Anammoxoglobus, Scalindua, Jetteniaの5属約20種が提案されている[4]。Anammox細菌は出芽ではなく分裂により増殖する点、細胞質内にエネルギー代謝を行う細胞内小器官アナモキソソームを有する点でも他のプランクトミケス門細菌と異なっている[5]。
近年では水系だけでなく、土壌などからもプランクトミケス門の16S rRNA配列が見つかっている。
プランクトミケス門 (Planctomycetes、プランクトマイセス門) は、グラム陰性細菌の小型の門で、幾つかの水生従属栄養細菌を含む。この門には2綱2門2科12属17種が属している。系統的にはウェルコミクロビウムやクラミジア(他にレンティスパエラ)に比較的近縁だが、他の細菌群とは離れている。分類体系によっては、この3系統を合わせPVC群(あるいはプランクトバクテリア)を設置する場合がある。 タイプ属のPlanctomycesは、ギリシャ語のπλαγκτός(プランクトス/漂う)+μύκες(ミュケス/菌)をラテン語化したもので、「(水中で)浮遊する菌」との意を持つ。
原核生物の中では最も複雑な構造と生活環を持つグループの1つである。形状は大まかに卵形であるが、柄を持つなど変わったところがあり、出芽によって増殖する点でも特異である。細胞壁は通常の細菌と異なりペプチドグリカンを含まないと考えられてきたが、最近になってプランクトミケス門細菌の細胞壁にもペプチドグリカンが存在することが確認された。また、細胞内に核膜のような構造を形成することでも知られる。原核生物においてこの構造は、プランクトミケスを含むPVC系統と、古細菌であるIgnicoccusにしか発見されていない。Gemmata obscuriglobusなどでは特に顕著で、細胞壁