
The following description of pin cherry provides characteristics that may be relevant to fire ecology, and is not meant for identification. Keys for identification are available (e.g. [44,104,115,127]).
Pin cherry is a shrub or small tree [19,63,96,109,127]. It generally has a straight trunk and a narrow, round-topped crown [5,53,59,66,109,127], though it may form thickets [109]. Branches, at first ascending, become more or less horizontal and spreading with age. Pin cherry generally grows 15 to 50 feet (5-15 m) tall and 4 to 20 inches (10-51 cm) in diameter [5,53,59,66,109,127]. However, trees up to 100 feet tall (30 m) have been found in the southern Appalachians [19,53], with the largest size attained on western slopes of the Great Smoky Mountains [53]. In western North America, pin cherry (P. p. var. saximontana) may be generally smaller with an arching shrub form, growing 5 to 15 feet (1.5-4.5 m) tall and spreading 5 to10 feet (1.5-3 m) [23].
Pin cherry has thin foliage [63], with leaves 1.5 to 4.3 inches (4-11 cm) long [59,66,109,127] and 0.5 to 1.75 inches (1-4.5 cm) wide [59,66,127]. Flowers grow in small clusters of 5 to 7 with individual flowers 0.4 inch (1 cm) across. Fruit are drupes 0.15 to 0.3 inch (4-8 mm) across with 1 large seed [19,53,59,66,109,127,143]. Seeds are 0.15 to 0.24 inch (4-6 mm) in diameter with a thick seed coat. There are 13,600 to 22,700 seeds per pound (30,000-50,000 seeds/kg) [19,143].
Pin cherry has a shallow root system [53,143]. In New England, root systems in 4- to 14-year-old stands were less than 14 inches (36 cm) deep and had many lateral branches. In West Virginia, root systems of wind-thrown trees 25 years old were confined to the upper 24 inches (61 cm) of soil. Once a seedling reaches a height of about 3 feet (1 m), lateral roots begin rapid growth. [143]. Though Prunus spp. have been reported to be ectomycorrhizal, studies by Malloch and Malloch [85,86] in Ontario found no evidence of ectomycorrhizae in pin cherry roots. Endomycorrhizae were generally scarce in pin cherry roots, and most often absent altogether [85,86].
Pin cherry is short lived [63,109,127], maturing rapidly and dying off at 20 to 40 years [5,53,55,75,87]. Stands may begin to have a pulse of dead pin cherry wood deposition 15 years after disturbance. Pin cherry allocates relatively less carbon to structural stem issues and to fine roots than do other hardwoods, contributing to the common occurrence of stem snap at the root crown. Possible low production of defense compounds may encourage damage by insects and pathogens [68].
Fire adaptations: Pin cherry has adapted to fire by establishing a soil seed bank that germinates rapidly following fire and other disturbance [32,116,133,144]. Pin cherry seed survives most forest fires, and seeds which may be dormant for many years are stimulated to germination by the altered conditions after fire [48,87,144]. Combined with the rapid initial growth of pin cherry seedlings [48,53,109], this characteristic enables pin cherry to dominate many burned-over areas, particularly in the northern hardwood forest [48]. For more information on pin cherry establishment and seedling growth see Botanical and Ecological Characteristics. Pin cherry sprouts after cutting [71], and though not documented, it may also sprout after fire. Further research is needed on postfire regeneration of pin cherry.
FIRE REGIMES: Pin cherry occurs in plant communities and ecosystems that historically experienced a range of FIRE REGIMES. In oak-hickory communities, for example, fires were typically frequent (<35-year mean), low-severity surface fires. Conversely, northern maple-beech-birch communities experienced long intervals between fire, and fire was usually stand-replacing [138]. White-red-jack pine communities experienced a mixed-severity regime with low to moderate severity at frequent intervals (20-40 years) and stand-replacing fires over longer intervals (100-300 years) [45]. FIRE REGIMES for plant communities and ecosystems in which pin cherry occurs are summarized below. Find further fire regime information for the plant communities in which this species may occur by entering the species name in the FEIS home page under "Find FIRE REGIMES".
Community or Ecosystem Dominant Species Fire Return Interval Range (years) grand fir Abies grandis 35-200 [6] maple-beech-birch Acer-Fagus-Betula > 1,000 silver maple-American elm A. saccharinum-Ulmus americana < 35 to 200 sugar maple A. saccharum > 1,000 sugar maple-basswood A. saccharum-Tilia americana > 1,000 sugarberry-America elm-green ash Celtis laevigata-U. americana-Fraxinus pennsylvanica 138] curlleaf mountain-mahogany* Cercocarpus ledifolius 13-1,000 [10,120] beech-sugar maple Fagus spp.-A. saccharum > 1,000 black ash Fraxinus nigra 138] Rocky Mountain juniper Juniperus scopulorum < 35 tamarack Larix laricina 35-200 [102] western larch L. occidentalis 25-350 [7,16,38] yellow-poplar Liriodendron tulipifera 138] Great Lakes spruce-fir Picea-Abies spp. 35 to > 200 northeastern spruce-fir Picea-Abies spp. 35-200 [45] southeastern spruce-fir Picea-Abies spp. 35 to > 200 [138] Engelmann spruce-subalpine fir P. engelmannii-A. lasiocarpa 35 to > 200 [6] black spruce P. mariana 35-200 [45] blue spruce* P. pungens 35-200 [6] red spruce* P. rubens 35-200 jack pine Pinus banksiana 45] Rocky Mountain lodgepole pine* P. contorta var. latifolia 25-340 [15,16,131] western white pine* P. monticola 50-200 Pacific ponderosa pine* P. ponderosa var. ponderosa 1-47 [6] interior ponderosa pine* P. ponderosa var. scopulorum 2-30 [6,13,77] red pine (Great Lakes region) P. resinosa 10-200 (10**) [45,51] red-white-jack pine* P. resinosa-P. strobus-P. banksiana 10-300 [45,62] pitch pine P. rigida 6-25 [28,64] eastern white pine P. strobus 35-200 eastern white pine-eastern hemlock P. strobus-Tsuga canadensis 35-200 eastern white pine-northern red oak-red maple P. strobus-Quercus rubra-Acer rubrum 35-200 [138] eastern cottonwood Populus deltoides 102] aspen-birch Populus tremuloides-Betula papyrifera 35-200 [45,138] quaking aspen (west of the Great Plains) Populus tremuloides 7-120 [6,58,94] black cherry-sugar maple Prunus serotina-Acer saccharum > 1,000 [138] Rocky Mountain Douglas-fir* Pseudotsuga menziesii var. glauca 25-100 [6,8,9] oak-hickory Quercus-Carya spp. < 35 northeastern oak-pine Quercus-Pinus spp. 10 to < 35 white oak-black oak-northern red oak Q. alba-Q. velutina-Q. rubra < 35 northern pin oak Q. ellipsoidalis < 35 bear oak Q. ilicifolia < 35 bur oak Q. macrocarpa < 10 chestnut oak Q. prinus 3-8 northern red oak Q. rubra 10 to 138] western redcedar-western hemlock Thuja plicata-Tsuga heterophylla > 200 [6] eastern hemlock-yellow birch Tsuga canadensis-Betula alleghaniensis > 200 [138] mountain hemlock* T. mertensiana 35 to > 200 [6] elm-ash-cottonwood Ulmus-Fraxinus-Populus spp. 45,138] *fire return interval varies widely; trends in variation are noted in the species summaryPin cherry has an elevational range from nearly sea level to over 9,000 feet (2,740 m) [19,24,31,42,53,75,105,117,129,143]. In the northeastern part of its range, pin cherry is found from 100 to 4,080 feet (30-1,240 m) [24,75]. In the southern Appalachians it generally occurs between 2,500 and 4,500 feet (762-1,370 m) [31,53,105,117,129,143]. In the western states, pin cherry is found between 5,000 and 9,300 feet (1,520-2,830 m) [42].
Throughout pin cherry's range, the number of days of snow cover ranges from 1 to 10 days in the South to 120+ days in the North [53]. Mean annual total snowfall ranges from 24 inches (610 mm) in the southern Appalachians to 100 inches (2,540 mm) in the northern part of the range. Average annual precipitation ranges from 16 inches (410 mm) in the West to 30 inches (760 mm) in Canada and 80 inches (2,030 mm) in the Great Smoky Mountains; average growing seasons vary from 100-210 days [23,53,118,143]. In the southern Appalachians, the average number of days with minimum temperatures below freezing is 90; in the northern and western part of pin cherry's range the number of days is more than 180 [143]. In Colorado, pin cherry is cold hardy to -50 oF (-45 oC) [23]. As demonstrated below, normal daily temperatures vary widely throughout the range of pin cherry (data are in oF/oC) [118,143]:
Southeast Northeast and West January max. 50/10 30/-1 January min. 25/-4 -10/-23 July max. 80/27 85/29 July min. 50/10 40/4Pin cherry grows on a wide range of soils and drainage classes [19,55,143]. Generally found on moderately coarse to coarse soils [61,109,127,137], pin cherry grows on sites varying from rocky ledges and sandy plains to moist loamy soils [19,23,55,143,143]. Pin cherry sites can be characterized as water-shedding (rocky ridges, cliffs, dry woods, clearings) or water-receiving (sandy and gravelly banks, shores of rivers and lakes) [109,127,137]. Pin cherry grows well on somewhat dry sites and shallow organic layers relatively low in nutrients [23,53,61,113,143]. Soils very low in moisture may result in a shrub form of pin cherry [66]. Though commonly found on mesic sites with nutrient-rich soil [25,53], pin cherry is generally absent from wet sites [109,143]. Optimum pH for pin cherry is 5.0 to 6.0 [53], though is also grows on more acidic soils [67].
Pin cherry is shade intolerant [18,25,27,53,75,91,109]. A study of shelterwood harvest in the northern hardwood forest of Vermont found that pin cherry increased in abundance with <60% canopy cover [60]. Pin cherry establishes in canopy gaps and proliferates with removal of overstory, increasing in density and cover [33,41,91]. It is frequent in burned areas, clearings, disturbed areas, roadsides, fencerows, and along forest margins [53,66,109,127,137]. A New Hampshire study found that pin cherry was relatively more abundant in recently created (24 years) gaps, large gaps (1,052-2,428 m2), and gap centers. Pin cherry abundance increased with gap size [91]. In a New England hardwoods forest, pin cherry growth exceeded that of other pioneer species near the center of clearcut openings, while other species sometimes grew faster near the side-shade zone at the edges of openings [126].
Pin cherry is important winter moose browse in the Great Lake states and boreal forest region [4,92,101,109]. Deer browse it moderately, especially in late winter and spring [3,53,109]. Pin cherry foliage and bark are important winter foods for snowshoe hares and cottontail rabbits [39,53,109], and the bark is preferred fall and winter food for porcupines [132].
Pin cherries are important to wildlife; they are eaten in summer and fall by at least 25 nongame birds (e.g. American robins, bluebirds), several upland game birds (e.g. ruffed grouse), large and small mammals (e.g. black bears, raccoons), and game animals [17,53,109,114,119,136,143].When birds eat the cherries, they disgorge the stones after the pulp is ingested [17]. Upland game birds, especially sharp-tailed and ruffed grouse, eat pin cherry buds [53,143].
Palatability/nutritional value: Pin cherry is characterized as preferred moose browse in Newfoundland [43,69]. Pin cherry palatability for white-tailed deer is low in fall and winter months, but improves from April to September [65,69].
Average mid-summer nutrient concentrations of pin cherry leaves, stems, and roots, based on 4 years of sampling in New Hampshire hardwoods, follow [97]:
Dry mass (%) N P K Ca Mg leaves 3.57 0.221 1.685 0.61 0.287 stems 0.918 0.076 0.838 0.4 0.059 roots 0.728 0.065 0.634 0.342 0.063Nutrient composition (in %) of pin cherry fruits from northern Ontario is presented below [135]:
Moisture content Dry matter Fat Protein Soluble carbohydrate based on glucose 70.22 29.78 1.99 1.53 11.45Though deer browse pin cherry, its calcium:phosphorus ratio may be too high for deer nutrition [53]. Pin cherry has nutritional value similar to that of chokecherry [96]. The foliage of pin cherry contains hydrocyanic acid, and livestock browsing the leaves may be poisoned [93,109,143]; however, the toxicity of pin cherry leaves is lower than that of most other cherry species [143].
Cover value: Pin cherry generally provides fair to good cover for mule deer, white-tailed deer, and pronghorn, but provides poor cover for elk [42]. It offers good hiding cover for small mammals and fair to good nesting cover for nongame birds [42,53,109,143]
In the northern part of its range, pin cherry occurs in virtually every forest
type. It is characteristically a short-lived, successional tree
in eastern hemlock (Tsuga
canadensis), northern hardwood, and spruce (Picea spp.)-fir (Abies
spp.) forests [53]. Pin cherry may also occur in pure stands, or more commonly, represent a majority of
stocking [55].
Pin cherry is found in the boreal forest region with white spruce (P. glauca),
black spruce (P. mariana), balsam fir (A. balsamea), paper
birch (Betula papyrifera), quaking aspen (Populus tremuloides),
chokecherry (Prunus virginiana), mountain maple (Acer spicatum),
speckled alder (Alnus rugosa), American green alder (A. crispa ssp.
crispa), oblongfruit serviceberry (Amelanchier bartramiana), roundleaf
serviceberry (A. sanguinea), California hazel (Corylus cornuta
var. californica), Bebb willow (Salix bebbiana), northern mountain-ash (Sorbus
decora), red raspberry (Rubus idaeus), skunk currant (Ribes glandulosum), wild sarsaparilla
(Aralia nudicaulis), fireweed (Epilobium angustifolium), whorled
wood aster (Oclemena acuminata),
bunchberry (Cornus canadensis), red-osier dogwood (C. sericea),
highbush cranberry (Viburnum edule), starflower (Trientalis borealis), and goldthread (Coptis
groenlandica) [20,33,49,139].
In the northern hardwoods region, frequent associates of pin cherry include quaking and bigtooth aspen
(Populus grandidentata); paper and
yellow birch (B. alleghaniensis); striped, red, and sugar maple (Acer
pensylvanicum, A. rubrum, and A. saccharum); American beech (Fagus
grandifolia), basswood (Tilia americana), eastern hemlock, northern red oak
(Quercus rubra), balsam
fir, and red spruce (Picea rubens) [24,36,55,143]. Prior to crown closure, raspberries (Rubus
spp.) and red elderberry (Sambucus racemosa ssp. pubens) are also abundant. Following stand closure,
these species are replaced by
shade-tolerant shrubs such as hobblebush (V. lantanoides) and Canada yew
(Taxus canadensis). Other common understory
plants include dwarf raspberry (R. arcticus ssp. acaulis), wild sarsaparilla, whorled wood
aster, bracken fern (Pteridium aquilinum), spinulose woodfern (Dryopteris
carthusiana), and shining clubmoss (Huperzia lucidula) [55,143]. In the Central and Lake States,
chokecherry (Prunus virginiana) and black
cherry (Prunus serotina) are particularly common associates [36,55,143].
In the southern and mid-Appalachian mountains, common pin cherry associates
are eastern hemlock; red spruce; Fraser fir (Abies fraseri); yellow and sweet birch (B. lenta);
American beech; pignut hickory (Carya glabra); sugar, red, mountain, and
striped maple; black cherry; American mountain-ash
(Sorbus americana); and northern red oak. Understory associates include
downy serviceberry (A. arborea), Allegheny serviceberry (A. laevis), Canadian serviceberry
(A. canadensis), flowering and alternate-leaf dogwood (Cornus florida and C. alternifolia),
red elderberry, southern bush honeysuckle (Diervilla
sessilifolia), mountain
holly (Ilex montana), mountain-laurel (Kalmia latifolia),
hobblebush, huckleberry (Vaccinium spp.), thornless blackberry (R. canadensis),
red raspberry, and Appalachian gooseberry (Ribes
rotundifolium) [24,30,31,34,55,81,105,143].
Pin cherry is also common in
heath balds adjacent to southern spruce-fir forests, occurring in thickets with
American mountain-ash, hawthorn (Crataegus spp.), mountain maple,
treefern (Dicksonia spp.), currants (Ribes spp.), raspberries (Rubus
spp.), sedges (Carex spp.), Catawba rosebay (Rhododendron catawbiense),
rosebay (R. maximum), minniebush (Menziesia pilosa), and
mountain-laurel [24,31,34,73].
Classifications identifying pin cherry as a plant community dominant are
listed below:
Due to its short life span and thin foliage, pin cherry is not usually considered
a serious competitor to commercially valuable hardwoods. However, high
densities of pin cherry may reduce the establishment and growth of commercial
hardwoods (e.g. black cherry, white ash, and sugar maple) [63]. Following clearcutting,
control plots in a northern hardwood forest study
demonstrated pin cherry dominance in stands aged 6 to 23 years. Basal area and
leaf area of pin cherry peaked at about year 17, followed by a steady decline in
pin cherry dominance. Fertilization treatments (combined N, P, K, Ca, Mg)
increased and prolonged pin cherry dominance, indicating that nutrient
limitation may accelerate the demise of pin cherry during the 2nd and 3rd
decades of stand development. In response to fertilization,
pin cherry had more leaf area pre unit stem basal area, and proportionally more
of this leaf area was in the upper canopy [14]. Demchik and Sharpe
[40], however, found little response of pin cherry to N-P-K
fertilizer following cutting. Frequency of pin cherry tends to increase after
stand harvest because seed stores well in the seed bank and is stimulated to
germinate when the canopy is opened [40]. Following cutting in a Massachusetts northern
hardwood forest, initial pin cherry density was approximately 40,000 stems/acre
(100,000 stems/ha).
Within 20 years, density decreased to 720 stems/acre (1,800 stems/ha), and
dropped to 5.6 stems/acre (14 stems/ha) after 40 years. Prior to cutting no pin cherry was documented
in the stand, illustrating the volume of seeds that may be buried in the seed
bank [5]. Due to the prolonged viability of pin cherry seeds
in the soil, harvest rotations of 120 years are recommended to allow sufficient
depletion of the pin cherry seed bank and stabilize population size. A study of
northern hardwood stands in New Hampshire found seed bank depletion to be
negligible in stands aged 40-60 years, and moderate (30% over 20 years) in
stands aged 95-115 years. Short harvest rotations (e.g. 60 years) may triple the
size of the pin cherry seed bank, resulting in greater proliferation
post-harvest [133]. Shading from slash cover may prevent pin
cherry germination and/or survival of new germinants [111].
Pin cherry responds well to clipping and/or browsing [3,29,40,53]; a study in the Lake States region found
annual growth increased by 88 to 121% following 4 years of "heavy"
clipping [3]. A study in western Ontario found cutting of pin cherry stems
initially decreased density by 25% after 1 year. However, stem density in the
2nd and 3rd years exceeded precut levels by 5 and 8%, respectively. Stem thinning by
natural mortality in the untreated control plots was 41% to 69% over 4 years [84]. A New England study found that moose and
white-tailed deer preferred browsing taller pin cherry plants. Browsed plants
had higher relative
height growth following browsing (compensatory growth) than unbrowsed plants at
"low" and "intermediate" densities. Compensatory growth decreased with increased
density. Despite browser preference for taller plants, there was a clear
net growth advantage for pin cherry of initial large size, when the effects
of competition, browsing and compensatory growth were combined [121]. Though it
generally does well under moderate to heavy browsing [53], heavy browsing of pin cherry can nearly eliminate it from a site [126].
A Pennsylvania study found that white-tailed deer browsing significantly reduced (p<0.05) pin
cherry density [90]. Heavy barking of stems by snowshoe hares can cause pin
cherry mortality [39].
Pin cherry is easily controlled by 2,4-D, 2,4,5-T, and triclopyr
[29,98], and moderately controlled by hexazinone
[29,146] and glyphosate [29,83,98,103,130].
Yarborough and Bhowmik [146] found hexazinone significantly
reduced (p<0.05) cover and frequency of pin cherry. Raymond and others [107]
report a single application of glyphosate resulted in an 82% reduction
in pin cherry browse by posttreatment year 2. Mallik and others [84] found
that both single and multiple applications of glyphosate significantly reduced
(p=0.035) pin cherry stem density, achieving 90% mortality by the 3rd year
following treatment.
Pin cherry currently has little commercial value [63], though recent interest in commercial production of pin cherry fruit has emerged [122]. The fruit is edible and can be used in jams and preserves [109,137].
Wood Products: Pin cherry wood is light, moderately soft, porous, and low in strength [66,143] giving it little commercial value. In general, pin cherry is not used for lumber and is considered a noncommercial species. It occurs in abundance, however, over a wide range of sites and produces large quantities of biomass in a relatively short time. The species has been described as well adapted to intensive management and chip harvesting on short rotations for fiber and fuel [143]
Pin cherry regenerates by both seed and sprouts. However, in a 25-year-old Maine hardwood forest 1 year after clearcutting, pin cherry was represented primarily by seedlings, not sprouts [144].
Breeding system: The flowers of nearly all Prunus species are bisexual [57].
Pollination: Prunus species are insect-pollinated [57].
Seed production: Pin cherry produces abundant seed at early ages [63]. Sexual maturity may be reached as early as 2 years [87,133,143], though large quantities of fruit are generally not produced for several years later [143]. Fruit is produced annually with large crops occurring every 2 to 3 years [87]. By the time pin cherry dies off (20 to 40 years), sufficient numbers of seeds have been produced and disseminated in a dormant condition for the reestablishment of pin cherry following disturbance [87,133]. Despite its short life span, fruit production of pin cherry is high. In New Hampshire, annual fruit production of 15-year-old, open-grown trees in pure stands was estimated at 1,118,000 fruits/acre (2,762,500/ha). For 25-year-old pin cherry stands in the same area, annual seed production was 940,700 seeds/acre (2,324,500/ha) [143]. Seed production peaks between 10 and 25 years, declining thereafter [22,133].
Seed dispersal: Pin cherry seeds are dispersed by birds, small mammals [53,55,87,143], and gravity [53,143].
Seed banking: As part of its reproductive strategy, pin cherry maintains a bank of seeds in the soil that remain viable for many years [32,133]. Seeds accumulate over prolonged periods (e.g. 50 years) [12,87,89], and most pin cherry seeds available for establishment following disturbance in mature stands (~100 years) are the result of seed banking rather than recently dispersed seed [55,89]. In a clearcut Maine hardwood forest, pin cherry seedlings were numerous although occurrence in surrounding stands and presumably in the preharvest stand was low [144]. Estimates of pin cherry seed contained in the duff of middle-aged northeastern deciduous forests range from 100,000 to >1.8 million per acre (250,000-4.5 million/ha) [11]. In 2 areas in New Hampshire, the average number of viable pin cherry seeds in the forest floor ranged from 139,676/acre (345,000/ha) to 200,000/acre (494,000/ha). In other New Hampshire stands, depending on stand age, the number of viable seeds in the forest floor ranged from 4,050 to 450,000/acre (10,000-1,110,500/ha) [143]. The distribution of pin cherry seeds in the forest floor is influenced by prior colonization of the site by pin cherry; dissemination of fruits by birds; and small mammal caches that tend to shift the distribution of seeds toward more or less discrete aggregations [32,87]. Longevity of buried pin cherry seeds has been estimated at 50 to 100 years [55,87]; pin cherry establishment after the removal of older stands relies primarily on seed dispersal into disturbed areas [55]. Seeds lose viability over time, so the pin cherry seed bank is eventually depleted in the absence of large-scale disturbance. The seed bank may persist at least 30 years after production ceases with little depletion, but is depleted gradually thereafter [22,133]. Seed bank losses are the result of seed rot and consumption by small mammals [32].
Germination: Pin cherry germination requires 1) the aging of the endocarp, either to break down an inhibitor or to increase permeability; and 2) an altered microclimate conducive to germination [87]. Dormancy of pin cherry seeds is caused by an initial physiological inhibition for several years followed by a secondary dormancy that remains until germination is stimulated by disturbance [133]. Because pin cherry seeds require open conditions for germination, there is often a prolonged delay between seed dispersal and germination [32]. Pin cherry seed banks may germinate in response to changes in soil temperature, light, or other triggering factors associated with the formation of a large gap due to disturbance [12,32,87,110]. However, the specific aspect of the changed environment that triggers germination is not known [32]. A study of stand development in Nova Scotia found higher incidence of pin cherry regeneration on soil disturbed by logging machinery than on undisturbed soil [139]. Laboratory experiments indicate that germination in open areas may be related to more extreme temperature fluctuations [76,143]. Germination may also be promoted by increased soluble nitrogen concentrations in the soil, based on increases in pin cherry germination following nitrogen (especially nitrate) fertilization [11].
The presence of occasional pin cherry seedlings beneath the canopy of undisturbed forest suggests that at least some germination occurs on a regular or annual basis [32,87,143]; however, seedlings have been reported to survive only in large openings where light and moisture were more available [143].
Seedling establishment/growth: Pin cherry establishment is directly related to open stand conditions. In a study of spruce-balsam fir forest canopy disturbance (spruce budworm outbreak), pin cherry established in greater abundance with 100% canopy removal than with partial canopy removal [99]. When established in high density, pin cherry grows quickly with early attainment of canopy closure [87]. When pin cherry occurs in high density (more than one 5-foot stem/43 ft2 (1.5-m/4 m)), it may live longer than when occurring at lower density (less than one 5-foot stem/43 ft2), because high pin cherry density early in stand development delays the time when shade-tolerant species reach a stable proportion of the total basal area. Pin cherry develops an early height advantage over other species, and as pin cherry density increases, growth and survival of other hardwood (black cherry, red maple, sugar maple) seedlings decreases. A study of an Allegheny hardwood stand found that survival of black cherry, red maple, and sugar maple at age 15 decreased as the density of pin cherry >5 feet (1.5 m) tall at age 3 increased. The height of black cherry and white ash (Fraxinus americana) also decreased [110]. In dense stands, the pin cherry canopy closes in about 3 years, shading out many of the other early intolerant species. After 25-30 years, sugar maple, beech and balsam fir are the seral species. At intermediate densities, pin cherry may codominate with yellow birch, paper birch, and quaking aspen. At low densities, dominance is shared by many species including blackberries, striped maple, paper and yellow birch, quaking aspen, and stump sprouts of cut trees [143].
Pin cherry exhibits a strong self-thinning tendency. On a jack pine (Pinus banksiana) plantation in Ontario, pin cherry density (sprouting) was 1,440 stems/acre (3,600 stems/ha) 4 years after harvest and fell to 488 stems/acre (1,220 stems/ha) 6 years after harvest [82]. In a study of Allegheny hardwoods, pin cherry seedlings ranged from 1,400 to 78,000 stems/acre (3,500-195,000 stems/ha) 1 year after overstory removal. First-year seedlings were less than 5 feet (1.5 m) tall. By year 3, stands averaged 6,800 stems/acre (17,000 stems/ha), and trees were more than 5 feet tall. After year 5, the number of pin cherry stems declined rapidly and only 1,600 stems/acre (4,000 stems/ha) remained at year 15. The 15-year-old stems exceeded 39 feet (12 m) in height [110].
Pin cherry seedlings grow rapidly, especially when young [53,109]. It is not uncommon for pin cherry growing on good sites in the central Appalachians to reach 8 to 10 inches (20-25 cm) in diameter in 25 years. Rapid growth is directly related to the amount of light received [143]. In preliminary results, Roberts [112] found height growth of pin cherry seedlings was significantly greater (p<0.05) under partial-canopy (4.4 feet (1.3 m) tall) and open-canopy (9.8 feet (3 m) tall) conditions than under closed-canopy conditions (1.2 feet (0.4 m) tall).
Asexual regeneration: Once established, pin cherry seedlings may reproduce by sprouting and may form thickets [53]. Pin cherry has a high potential for stem sprouting and root suckering after cutting [71,71,143]. In a Quebec study most of the suckers forming a clone emerged 1 to 2 years after the parent stem was cut, with the number of suckers per clone ranging from 11 to 32. The largest clone studied covered 153 square feet (14.25 m2) [71].
Pin cherry is an early to mid-seral species, important in both primary and secondary succession after fire or harvest [12,20,32,35,105,145]. Pin cherry is common in rock- and soil-slide succession, but is usually far more numerous after windthrow, logging, or light fires because its presence is highly dependent on seed buried in the forest floor [29,143]. Pin cherry does, however, colonize through "seeding in" [29,87]. It is generally restricted to sites that supported forest vegetation prior to disturbance; it is not common in abandoned pastures or old fields. Pin cherry is further restricted to colonization of open areas larger than about 0.25 acre (0.1 ha) [87].
Pin cherry is a particularly common successional species in northern hardwood and spruce-fir forest ecosystems, dying out relatively quickly as seral stands develop [24,49,87,139]. For example, pin cherry that established following a landslide in the northeastern spruce-fir zone was no longer present in the stand after 40 years [50]. In northern hardwood forests, pin cherry grows quickly after stands are cut and is often the tallest species from the 2nd to the 6th year of stand regrowth [27]. Pin cherry can be an effective competitor on disturbed sites [63,109] because it employs a vertical competition strategy, competing with other species for light by overtopping them due to rapid height growth [82]. Pin cherry often dominates regenerating northern hardwood or spruce stands (up to 70% of stems) for 15-20 years after disturbance [48,53,80,133,145], though it may die out sooner [48]. Pin cherry may also codominate with aspen, black cherry, red maple, and white or gray birch [53].
The pin cherry forest type develops rapidly, forming a closed canopy in 3 to 7 years and reaching maturity in 20 to 30 years [55]. Dominance shifts to mature forest species after approximately 30 years, and pin cherry rarely persists in stands after 40 years due to a combination of nutrient limitation, shade intolerance, and the relatively short life span of pin cherry [5,11,55,133]. A study in New England hardwoods found pin cherry was initially prominent following clearcutting, especially near the center of openings, but had died or become moribund after 20 years [126]. The initial density of pin cherry stands, determined by available buried seed, is an important determinant of succession. Where pin cherry is very dense, canopy closure limits the establishment and growth of other early successional species. Succession occurs in 2 distinct phases: the 1st is characterized by an almost pure pin cherry overstory and sparse understory; the 2nd phase involves the relatively abrupt dominance by shade tolerant species like sugar maple, beech, and balsam fir. At intermediate densities, pin cherry may codominate with other fast-growing species (e.g. yellow birch, paper birch, quaking aspen, bigtooth aspen), with shade-tolerant species gaining dominance gradually because shade-intolerant species die and are replaced at different times [55,63,87]. Successional patterns in low-density stands of pin cherry are quite variable, primarily because the rate of canopy closure is slower [87]. Repeated disturbance may result in persistent dominance of pin cherry or codominance of pin cherry and aspens [80].
At lower elevations in New England, pin cherry succeeds to aspens, white pine, or white pine-northern red oak-red maple. In the southern Appalachians, succession is to red spruce, red spruce-Fraser fir, or northern hardwoods [55]. Twenty-five years after a fire in the Southern Appalachian spruce-fir forest, pin cherry density was 182 trees per acre (455 trees/ha), comprising 12.2% of total basal area, while Fraser fir had reached 531 stems per acre (1,327 stems/ha) and comprised 63.8% of total basal area [117,118]. This seral community was still several decades from a mature, closed-canopy spruce-fir forest. Pin cherry may persist longer on sites where severe fire, steep slopes, and shallow soil combine to slow development of mature stands [117].
Pin cherry sequesters nutrients that might otherwise be lost from the ecosystem during early stages of succession [88,109,133]. Rapid establishment and growth of pin cherry minimizes nutrient loss by 1) channeling water from runoff to evapotranspiration, reducing erosion and nutrient loss; 2) reduction in rates of decomposition through moderation of the microclimate during the growing season so that the supply of soluble ions available for loss in drainage water is reduced; and 3) incorporation into the rapidly developing biomass of nutrients that do become available and that may otherwise might be lost from the system [88].
Propagation: Soaking the seeds is helpful for pin cherry propagation, but scarification is unnecessary [53]. Seed germination rate can be improved by drastic temperature fluctuation treatments. Effective treatment involves a 24-hour soak followed by 30 days of stratification with 5 days at 41 oF (5 oC) alternating with 5 days at 86 oF (30 oC). The next step is 60 to 90 days of stratification at 41 oF, followed by a 10-day germination period with 12 hours at 41 oF alternating with 12 hours at 86 oF. This procedure can be expected to achieve at least 75% germination [76]. Belcher [19] recommends prechilling 60 to 90 days at 37 to 41 oF (3-5 oC) and encouraging germination with temperatures from 68 to 86 oF (20-30 oC). Air-dried seeds can be stored at 37 to 41 oF for 3-5 years [19]. Pin cherry seeds have retained viability for up to 10 years when stored in sealed containers at 34 to 38 oF (1-3 oC) [53].
Pin cherry suckers readily and is assumed by some authors to grow well from root cuttings [143]. However, propagation of pin cherry from cuttings has proved difficult with little success reported [23]. Pin cherry is used as grafting stock for sour cherry (Prunus cerasus) [53,143].
Restoration: Pin cherry grows naturally on unreclaimed coal mine spoils [53,67]
Planhigyn blodeuol sy'n frodorol o Hemisffer y Gogledd yw Coeden geirios Pensylfania sy'n enw benywaidd. Mae'n perthyn i'r teulu Rosaceae. Yr enw gwyddonol (Lladin) yw Prunus pensylvanica a'r enw Saesneg yw Pin cherry.[1]
Mae'r teulu Rosaceae yn perthyn i'r genws Rosa (rhosyn) fel ag y mae'r cotoneaster a'r eirinen. Prif nodwedd y teulu yw ei ffrwythau amrywiol a phwysig i economi gwledydd.[2] Ceir 5 sepal, 5 petal ac mae'r briger wedi'u gosod mewn sbeiral sy'n ffurfio llestr tebyg i gwpan o'r enw hypanthiwm.
Planhigyn blodeuol sy'n frodorol o Hemisffer y Gogledd yw Coeden geirios Pensylfania sy'n enw benywaidd. Mae'n perthyn i'r teulu Rosaceae. Yr enw gwyddonol (Lladin) yw Prunus pensylvanica a'r enw Saesneg yw Pin cherry.
Mae'r teulu Rosaceae yn perthyn i'r genws Rosa (rhosyn) fel ag y mae'r cotoneaster a'r eirinen. Prif nodwedd y teulu yw ei ffrwythau amrywiol a phwysig i economi gwledydd. Ceir 5 sepal, 5 petal ac mae'r briger wedi'u gosod mewn sbeiral sy'n ffurfio llestr tebyg i gwpan o'r enw hypanthiwm.
Prunus pensylvanica, also known as bird cherry,[3] fire cherry,[3] pin cherry,[3] and red cherry,[3] is a North American cherry species in the genus Prunus.
Prunus pensylvanica grows as a shrub or small tree, usually with a straight trunk and a narrow, round-topped crown. It grows 5–15 metres (16–49 feet) tall and 10–51 centimetres (4–20 inches) in diameter. Trees up to 30 m (98 ft) tall have been found growing in the southern Appalachians, with the largest found on the western slopes of the Great Smoky Mountains. Its foliage is thin,[5] with leaves 4–11 cm (1+1⁄2–4+1⁄4 in) long and 1–4.5 cm (3⁄8–1+3⁄4 in) wide. Flowers occur in small groupings of five to seven with individual flowers 1 cm (1⁄2 in) across. The fruit are drupes, ranging from 4–8 millimetres (1⁄8–3⁄8 in), each with a single seed 4–6 mm (3⁄16–1⁄4 in) in diameter contained within a hard "stone".[6][7]

Young trunk

Mature trunk

Trunk with peeling bark
.jpg)
Blossom

Tree in early summer
_-_Newfoundland_2019-08-21.jpg)
Specimen in summer

Branches with fruit
.gif)
Foliage and fruit

Leaf close-up
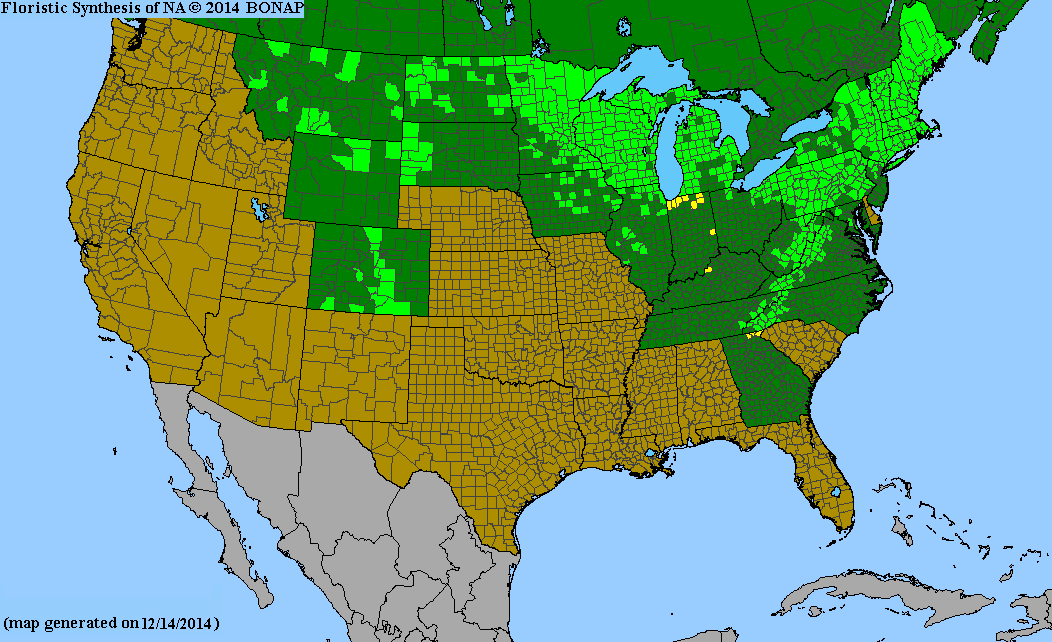
The species is widespread across much of Canada from Newfoundland and southern Labrador to British Columbia and the southern Northwest Territories. Additionally it is very common in New England and the Great Lakes region. It can also be found in the Appalachian Mountains as far south as northern Georgia and eastern Tennessee. Scattered growth of the pin cherry also occurs in the Rocky Mountains, south to Colorado as well as in the Black Hills of South Dakota.[8]
The pin cherry can regenerate by seed and sprout. Its flowers are bisexual and pollinated by insects. Seeds are dispersed by birds, small mammals, and gravity. As part of its reproductive strategy, pin cherries seeds can remain viable in the soil for many years. Seeds accumulate over prolonged periods, and soil seed banks may be viable for 50–100 years. Asexual reproduction is achieved by sprouting, and often thickets of pin cherry plants form.[6]
The pin cherry is rather short lived, having a lifespan of only 20 to 40 years following a rapid maturation. Its root system is shallow, with roots tending to grow laterally. It is an important food source for many animals. Winter moose browse it in the Great Lake states and boreal forest region.[6]
Though they are documented to sprout following cutting, individual pin cherry thickets are often killed if exposed to fire. Nonetheless, they have adapted as a species by the establishment of their seed banks which are protected from the most severe heat by their soil cover and fed by the nutrients in the resultant ash residue. Following a fire or other disturbance, seeds which may be dormant for years will germinate rapidly, stimulated by the altered conditions after fire. Combined with the rapid initial growth of seedlings, these characteristics enable groupings of Pin cherry thickets to dominate many burned-over areas, particularly in the northern hardwood forest.
The pin cherry serves as food for various Lepidoptera.
Pin cherry currently has little commercial value, though recent interest in commercial production of pin cherry fruit has emerged. The fruit is edible and can be used in jams, jellies, and preserves.[9]
Pin cherry wood is light, moderately soft, porous, and low in strength giving it little commercial value. In general, it is not used for lumber and is considered a noncommercial species. It occurs in abundance, however, over a wide range of sites and produces large quantities of biomass in a relatively short time. The species has been described as well adapted to intensive management and chip harvesting on short rotations for fiber and fuel.
{{cite web}}: CS1 maint: others (link) Note that this website has been superseded by World Flora Online Prunus pensylvanica, also known as bird cherry, fire cherry, pin cherry, and red cherry, is a North American cherry species in the genus Prunus.
Prunus pensylvanica, "pin cherry" o cerezo de fuego, es una especie perteneciente al género Prunus.
Se puede encontrar a partir de la Isla de Terranova y al sur del Labrador, cruzando Canadá al oeste y hasta en la Columbia Británica y al sur de los Territorios del Noroeste. Además es muy común en Nueva Inglaterra y los Estados del los Lagos del Sur, pero no muy común en Pennsylvania, donde solo se encuentra esporádicamente en los Montes Apalaches al norte de Georgia (Estados Unidos) y al este de Tennessee. Grandes poblaciones del Cerezo Pin también se producen en las Montañas Rocosas, al sur de Colorado y el sureste de la Black Hills en Dakota del Sur.
Crece como arbusto o árbol pequeño. El cerezo Pin generalmente tiene un tronco recto y estrecho, y remata en la corona. Crece de 5-15 m de altura y su tronco mide entre 10-51 cm de diámetro. Se han encontrado árboles de hasta 30 m de altura cada vez mayores en los Apalaches del Sur, con el ejemplar más grande encontrado en las laderas occidentales de la Grandes Montañas Humeantes. Su follaje es delgado, con hojas de 4-11 cm de largo y entre 1 y 4.5 cm de ancho. Manifiesta pequeños grupos de cinco a siete flores, de 1 cm de ancho cada una. Los frutos son drupas, que van desde los 4 a los 8 mm, que contienen semillas de 4-6 mm de diámetro con una cubierta de la semilla de buen espesor. El sistema de raíces de la planta es poco profundo, ya que las raíces tienden a crecer lateralmente.
Prunus pensylvanica es más bien de corta duración, con una vida útil de solo 20 a 40 años, después de una rápida maduración.
Prunus pensylvanica se regenerara por semillas y brotes. Sus flores son bisexuales y polinizadas por insectos. Las semillas son dispersadas por aves, pequeños mamíferos, y por acción de la gravedad. Como parte de su estrategia reproductiva, este cerezo mantiene un banco de semillas en el suelo que permanecen viables durante muchos años. Las semillas se acumulan durante largos períodos, de hasta 50 años, y los bancos de semillas pueden ser viables durante uno 50 o 100 años. La reproducción asexual se logra mediante la germinación de semillas, y, a menudo se forman matorrales de esta especie en forma de arbustos.
A pesar de que los matorrales del Cerezo Pin vuelven a brotar si se los corta, esta especie a menudo mueren si es expuesta al fuego. No obstante, se ha adaptado como especie por el establecimiento de sus bancos de semillas que están protegidas del calor más grave, gracias a su cobertura del suelo, y al ser alimentados por los nutrientes de los residuos de las cenizas, después de un incendio u otro tipo de perturbación. Las semillas que se mantienen latentes durante años, germinan rápidamente una vez estimuladas por las nuevas condiciones después de un incendio. Combinado con el crecimiento inicial rápido de las plántulas, estas características le permiten a las agrupaciones de matorrales del Cerezo Pin dominar muchas áreas quemadas, en particular en los bosque frondosos del norte.
El Cerezo Pin es una importante fuente de alimento para muchos animales. En invierno los alces migran debido a ella en los estados de los Grandes Lagos y la región boreal de la selva.
Este cerezo tiene en la actualidad poco valor comercial, aunque el reciente interés en la producción comercial de la cereza pin ha resurgido. El fruto es comestible y se puede utilizar en mermeladas, jaleas y confituras.
La madera de este cerezo es liviana, moderadamente porosa, y de poca fuerza que le da poco valor comercial. En general, no se usa para madera y se considera una especie no comercial. Se produce en abundancia, sin embargo, en una amplia gama de sitios y produce grandes cantidades de biomasa en un tiempo relativamente corto. La especie ha sido descrita como bien adaptado a la gestión y aprovechamiento intensivo del suelo en rotaciones cortas para su uso como fuente de leña.
Prunus pensylvanica sirve como planta base para muchas especies de Lepidoptera.
Prunus pensylvanica fue descrita por Carlos Linneo el Joven y publicado en Supplementum Plantarum 252–253. 1781[1782].[1]
Prunus: nombre genérico que proviene de un antiguo nombre griego (προύνη), y luego latino (prūnus, i) del ciruelo. Ya empleado por, entre otros, Virgilio (Geórgicas, 2, 34) y Plinio el Viejo (Historia naturalis,13, XIX, 64)[2][3]
pensylvanica: epíteto geográfico que alude a su localización en Pelsinvania.
Prunus pensylvanica, "pin cherry" o cerezo de fuego, es una especie perteneciente al género Prunus.
Pilvikirsikka (Prunus pensylvanica) on Pohjois-Amerikan pohjoisosista ja vuoristoalueilta kotoisin oleva kesävihanta, yksikotinen pensas tai pieni puu, joka kuuluu Prunus-sukuun ja ruusukasvien heimoon.[2][3]
Pilvikirsikka on juurivesallinen, oraton pensas tai pieni puu, joka kasvaa 5–15 metriä korkeaksi ja rinnankorkeusläpimitaltaan 10–50 senttimetriä paksuksi.[1][2] Pohjois-Amerikan länsiosissa se jää yleensä matalaksi ja pensasmaiseksi, kun taas Appalakkien länsirinteiltä on löydetty jopa 30 metrin korkuisia yksilöitä.[2]
Pilvikirsikalla on matalat juuret, suora runko, kapean pyöreä latvus ja yläviistot haarat, jotka alkavat iän myötä siirottaa.[1][2] Runkoa ja oksia peittää punaruskea ja poikkijuovainen kuori ja nuori haararanka on kalju. Lehdet ovat varrella kierteisesti ja lehtiruodit ovat 1–3 senttimetrin pituiset ja nystyiset. Lehtilapa on 8–12 senttimetriä pitkä, puikea, tyvestä leveäsuippuinen, kärjestä pitkäsuippuinen ja sahalaitainen. Se on yleensä molemmin puolin kalju mutta joskus alta suonihangoista karvainen.[1]
Pilvikirsikka kukkii maalis–heinäkuussa heti lehtien puhjettua ja kantaa hedelmää heinä–syyskuussa.[1][2] Kukinto on 4–5-kukkainen sarja. Yksittäiset kukat ovat säteittäisiä ja maljakehäisiä ja niissä on 1–1,5 senttimetrin pituinen kukkaperä. Verhiö on jakautunut 5 tylppään liuskaan ja teriössä on 5 valkoista, 6 millimetrin pituista terälehteä. Kukat ovat kaksineuvoisia ja niissä on paljon heteitä mutta vain yksi emi.[1][3] Hedelmä on punainen, kitkerä, 1-siemeninen luumarja, jonka läpimitta on 4–8 millimetriä.[1][2][3] Siemen on paksukuorinen ja halkaisijaltaan 4–6 millimetriä.[2]
Pilvikirsikkaa tavataan laajalla alueella Pohjois-Amerikan hemiboreaalisella vyöhykkeellä.[1][2] Kanadassa sen levinneisyysalue ulottuu Newfoundlandista ja Etelä-Labradorista Brittiläiseen Kolumbiaan ja Luoteisterritorioiden eteläreunalle. Yhdysvalloissa se on yleinen Uudessa Englannissa ja Suurten järvien ympäristössä. Etelämpänä se menestyy vain vuoristoalueilla: Black Hillsissä Etelä-Dakotassa, Kalliovuorilla Coloradoon asti ja Appalakeilla Georgiaan ja Tennesseehen asti.[2]
Pilvikirsikka on sekä kuivaan että kosteaan maaperään sopeutunut, lyhytikäinen pioneeripuu, joka valtaa nopeasti metsäpalojen tuhoamia alueita. Sitä tavataan keskiravinteisissa metsissä, metsänreunuksissa, hakkuu- ja kuloaloilla sekä tienvarsilla.[1]
Pilvikirsikan metsätaloudellinen arvo on vähäinen, sillä sen puuaines on kevyttä, huokoista ja lujuusominaisuuksiltaan heikkoa. Se on kuitenkin tärkeä ravinnonlähde villieläimille: hirvieläimet, lumikenkäjänikset, pumpulihäntäkaniinit ja puupiikkisiat käyttävät talviravintonaan sen kuorta ja lehtiä ja monet lintu- ja nisäkäslajit syövät sen marjoja.[2]
Pilvikirsikka (Prunus pensylvanica) on Pohjois-Amerikan pohjoisosista ja vuoristoalueilta kotoisin oleva kesävihanta, yksikotinen pensas tai pieni puu, joka kuuluu Prunus-sukuun ja ruusukasvien heimoon.
Cerisier de Pennsylvanie
Le cerisier de Pennsylvanie[1],[2] (Prunus pensylvanica[1] L. f.[2]) est un arbuste ornemental de la famille des Rosaceae. Aussi connu comme le merisier, le petit merisier[2], le cerisier d'été[2] ou l'arbre à petites merises[2] au Québec[3], on le trouve dans toute l'Amérique du Nord : Terre-Neuve, sud du Labrador, ouest du Canada jusqu'à la Colombie-Britannique et Territoires du Nord-Ouest.
L'espèce est très commune en Nouvelle-Angleterre. Elle l'est beaucoup moins au sud de la Pennsylvanie : on ne la retrouve que dans les Appalaches, au nord de la Géorgie et à l'est du Tennessee.
Elle est sporadique dans les montagnes Rocheuses, au sud du Colorado et le sud-est des Black Hills et du Dakota du Sud.
Le système racinaire est peu profond et s'étend latéralement.
Son tronc est droit et cylindrique, de 5 à 15 mètres de haut et 10 à 51 centimètres de diamètre. Des arbres de 30 mètres sont avérés dans les Appalaches. Ses feuilles sont étroites, de 4 à 11 centimètres de long pour 1 à 4,5 centimètres de large.
Les fleurs sont regroupées par inflorescence de 5 à 7 fleurs. La corolle d'une fleur fait environ un centimètre.
Le fruit est une drupe assez petite et peu juteuse.
Un individu a une espérance de vie de 20 à 40 ans.
Les fleurs sont hermaphrodites et la pollinisation est entomogame (par les insectes).
La dissémination des graines est faite par des oiseaux ou de petits mammifères. Sinon, les graines tombées au pied peuvent germer et former des haies. Les graines au sol non germées peuvent rester viables pendant plusieurs dizaines d'années. Cette réserve de graines est un élément déterminant dans la capacité de résistance au feu qu'à cette espèce.
Une multiplication végétative est également possible.
L'espèce est consommée par beaucoup d'animaux. Les élans en font une recherche systématique en l'hiver, les cerfs également, en moindre mesure, durant l'hiver et le printemps suivant.
Le feuillage et l'écorce sont une source importante de nourritures, en hiver, pour le lièvre d'Amérique et le lapin d'Amérique. L'écorce est recherchée par les porcs-épics, en hiver toujours.
Le fruit est comestible et peut être employé en confiture. Son commerce n'a jamais été réellement important jusqu'à présent.
Le bois de cette espèce est peu doux, assez lumineux mais poreux. Sa valeur marchande est faible. Il ne peut pas être utilisé en bois de charpente.
Cerisier de Pennsylvanie
Le cerisier de Pennsylvanie, (Prunus pensylvanica L. f.) est un arbuste ornemental de la famille des Rosaceae. Aussi connu comme le merisier, le petit merisier, le cerisier d'été ou l'arbre à petites merises au Québec, on le trouve dans toute l'Amérique du Nord : Terre-Neuve, sud du Labrador, ouest du Canada jusqu'à la Colombie-Britannique et Territoires du Nord-Ouest.
L'espèce est très commune en Nouvelle-Angleterre. Elle l'est beaucoup moins au sud de la Pennsylvanie : on ne la retrouve que dans les Appalaches, au nord de la Géorgie et à l'est du Tennessee.
Elle est sporadique dans les montagnes Rocheuses, au sud du Colorado et le sud-est des Black Hills et du Dakota du Sud.
Prunus pensylvanica, hay còn gọi là anh đào chim, anh đào lửa, anh đào đỏ, là một loại cây bụi thuộc chi Mận mơ ở vùng Bắc Mỹ[1][2].
P. pensylvanica có mặt trên khắp Canada, từ đảo Newfoundland và nam Labrador đến tận bang British Columbia và các Lãnh thổ Tây Bắc. Nó cũng phổ biến tại New England và vùng Ngũ Đại Hồ. Nó cũng nằm rải rác ở dãy Appalachian, dãy Rocky, bắc Georgia, đông Tennessee, dãy Black Hills (South Dakota)[3].
P. pensylvanica là một loại cây bụi rụng lá, thân thẳng với tán tròn và hẹp. Nó có chiều cao khoảng từ 5 đến 15 m, đường kính thân từ 10 đến 50 cm. Tuy nhiên, những cây P. pensylvanica cao tới 30 mét đã được tìm thấy tại phía nam của bang Appalachia, tập trung nhiều nhất tại sườn tây dãy núi Great Smoky. Lá của P. pensylvanica mỏng, dài 4 - 11 cm và rộng 1 - 4,5 cm. Hoa mọc thành cụm từ 5 tới 7 hoa nhỏ, đường kính 1 cm. Quả hạch, đường kính 4 - 8 mm, hạt cứng[4][5].
P. pensylvanica sinh sản bằng hạt và chồi. Hoa lưỡng tính và được thụ phấn bởi côn trùng. Hạt được phân tán bởi chim, động vật có vú nhỏ và gió. Hạt có thể tồn tại trong lòng đất từ 50 đến 100 năm. Tuy nhiên, tuổi thọ của loài P. pensylvanica khá ngắn, chỉ từ 20 đến 40 năm. Rễ của nó cạn, có xu hướng phát triển theo chiều ngang[5]. Quả của cây là nguồn thức ăn quan trọng cho nhiều loài động vật, nhất là loài nai sừng tấm trong các khu rừng taiga[5].
Quả của P. pensylvanica ít có giá trị thương mại. Gần đây, sự quan tâm đến việc sản xuất loại quả này đã xuất hiện. Chúng được dùng làm mứt, thạch và có thể ăn sống. Gỗ của loài này mềm và xốp, không được sử dụng nhiều.
Prunus pensylvanica, hay còn gọi là anh đào chim, anh đào lửa, anh đào đỏ, là một loại cây bụi thuộc chi Mận mơ ở vùng Bắc Mỹ.
Prunus pensylvanica L.f.
СинонимыЧерёмуха пенсильва́нская (лат. Prúnus pensylvánica) — вид рода Слива семейства Розовые. Родина — Северная Америка.
Небольшое лиственное дерево высотой от 1 до 6 метров, иногда вырастает до 12 метров.
Кора блестящего тёмно-вишневого цвета. Ветки тонкие и гибкие, крона воздушная[уточнить].
Цветёт в конце мая, цветки белые, собраны в соцветия, практически не имеют запаха.
В августе созревают небольшие красные съедобные плоды. Вкус у них кисло-сладкий или кислый.
Черёмуха пенсильванская светолюбива, нетребовательна к почве, зимостойка.
Черёмуха пенсильва́нская (лат. Prúnus pensylvánica) — вид рода Слива семейства Розовые. Родина — Северная Америка.



.jpg)





.jpg)

{kind=link}