Tetrahymena és un gènere de protozous ciliats de vida lliure no patogènics. Són comuns en aigua dolça. Les espècies de Tetrahymena que s'utilitzen com a organismes model en les recerques biomèdiques són T. thermophila i T. pyriformis.
Tetrahymena je označení pro skupinu prvoků z kmene nálevníci, žijících volně, běžně ve stojatých vodách. Preferují parazitický, či komenzální způsob života. Často bývají používáni jako modelový organismus (často druhy T. thermophila a T. pyriformis).[1]
V tomto článku byl použit překlad textu z článku Tetrahymena na anglické Wikipedii.
{{Cite book}} označená jako k „pouze dočasnému použití“. Tetrahymena je označení pro skupinu prvoků z kmene nálevníci, žijících volně, běžně ve stojatých vodách. Preferují parazitický, či komenzální způsob života. Často bývají používáni jako modelový organismus (často druhy T. thermophila a T. pyriformis).
Tetrahymena er en slægt af fritlevende ciliater. Det er mikrober som kan skifte mellem levemåderne kommensal og patogen. Tetrahymena er almindelig i ferskvandsdamme og er 30-60 mikrometer store. De arter af Tetrahymena, der anvendes som modelorganismer i biomedicinsk forskning, er Tetrahymena thermophila og Tetrahymena pyriformis.[1]
Tetrahymena thermophila har to cellekerner. Tetrahymena thermophila eksisterer i syv forskellige køn, som kan reproducere i 21 forskellige kombinationer - og en enkelt Tetrahymena thermophila kan ikke reproducere sig seksuelt med sig selv.
Tetrahymena er en slægt af fritlevende ciliater. Det er mikrober som kan skifte mellem levemåderne kommensal og patogen. Tetrahymena er almindelig i ferskvandsdamme og er 30-60 mikrometer store. De arter af Tetrahymena, der anvendes som modelorganismer i biomedicinsk forskning, er Tetrahymena thermophila og Tetrahymena pyriformis.
Tetrahymena ist eine Gattung eukaryotischer Einzeller, die dem Stamm der Wimperntierchen angehört. Arten dieser Gattung kommen in Meeren, im Süßwasser und in feuchter Erde vor. Aufgrund der besonderen Ausprägung ihres Mundapparats (Cytostom) werden sie zu den Hymenostomatida (gr. für ‚Haut-Münder‘) gezählt.
Die Arten der Gattung Tetrahymena sind 30 bis 60 μm groß und allseitig von einer holotrichen Ciliatur bedeckt. Das heißt, der ganze Körper ist von Wimpern überzogen, wodurch auch ihr Übergattungsname Wimperntierchen (von Cilien alias Wimpern) zu erklären ist. Die Ciliatur dient der Fortbewegung und ist extrem empfindlich gegenüber sensorischen (Sinnes-)Reizen. Der Organismus weist den gleichen Oralapparat (Zellmund, Cytostom) wie das Pantoffeltierchen auf, der erkennbar ein Viertel der vorderen Seite einnimmt. Mikroskopisch sichtbar sind die parallel angelegten, von Cilien bedeckten, vier Häutchen, die den Mund kennzeichnen. Die Tetrahymena haben dieser Struktur ihren Namen zu verdanken (gr.: tetra- = ‚vier‘).
Wie andere Wimperntierchen besitzt auch diese Gattung den, zur Exkretion benötigten, Cytoprokt (oder Zellafter). Tetrahymena-Arten weisen die typischen Merkmale einer eukaryotischen Zelle auf, wie beispielsweise endoplasmatisches Reticulum, Lysosom und Ribosom.[1][2]
Die für die Gattung Tetrahymena wie für alle Wimpertierchen kennzeichnende genetische Besonderheit ist der Kerndimorphismus. Die Organismen besitzen jeweils einen Mikro- und einen Makronukleus. Während im diploiden Mikronukleus die DNA sitzt, beinhaltet der Makronukleus die neu geordnete RNA-Interferenz (RNAi).[3]
Die Atmungskette in den Mitochondrien (zumindest) der Spezies T. thermophila weicht von der anderer Eukaryoten ab.[4]
Die Arten Tetrahymena thermophila and T. pyriformis werden als Modellorganismen genutzt.[5] T. thermophila wurde beispielsweise genutzt, um Tubulin-Hemmstoffe zu entwickeln. Diese Eiweiße sind für die Zellteilung der Eukaryoten nötig. Man hofft, solche Hemmstoffe als Mittel gegen Parasiten aus der Gruppe der Apicomplexa (etwa die Malaria-Erreger Plasmodium sp. und Toxoplasmose-Erreger Toxoplasma gondii), aber auch gegen menschliche Tumorzellen verwenden zu können.[6]
Artenliste (Auswahl):
Tetrahymena ist eine Gattung eukaryotischer Einzeller, die dem Stamm der Wimperntierchen angehört. Arten dieser Gattung kommen in Meeren, im Süßwasser und in feuchter Erde vor. Aufgrund der besonderen Ausprägung ihres Mundapparats (Cytostom) werden sie zu den Hymenostomatida (gr. für ‚Haut-Münder‘) gezählt.
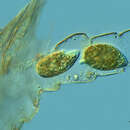
Tetrahymena, a unicellular eukaryote, is a genus of free-living ciliates.[1] The genus Tetrahymena is the most widely studied member of its phylum.[2]: 59 It can produce, store and react with different types of hormones. Tetrahymena cells can recognize both related and hostile cells.[3]
They can also switch from commensalistic to pathogenic modes of survival. They are common in freshwater lakes, ponds, and streams.[2]: 277
Tetrahymena species used as model organisms in biomedical research are T. thermophila and T. pyriformis.[4]

As a ciliated protozoan, Tetrahymena thermophila exhibits nuclear dimorphism: two types of cell nuclei. They have a bigger, non-germline macronucleus and a small, germline micronucleus in each cell at the same time and these two carry out different functions with distinct cytological and biological properties. This unique versatility allows scientists to use Tetrahymena to identify several key factors regarding gene expression and genome integrity. In addition, Tetrahymena possess hundreds of cilia and has complicated microtubule structures, making it an optimal model to illustrate the diversity and functions of microtubule arrays.
Because Tetrahymena can be grown in a large quantity in the laboratory with ease, it has been a great source for biochemical analysis for years, specifically for enzymatic activities and purification of sub-cellular components. In addition, with the advancement of genetic techniques it has become an excellent model to study the gene function in vivo. The recent sequencing of the macronucleus genome should ensure that Tetrahymena will be continuously used as a model system.
Tetrahymena thermophila exists in 7 different sexes (mating types) that can reproduce in 21 different combinations, and a single tetrahymena cannot reproduce sexually with itself. Each organism "decides" which sex it will become during mating, through a stochastic process.[5][6]
Studies on Tetrahymena have contributed to several scientific milestones including:

The life cycle of T. thermophila consists of an alternation between asexual and sexual stages. In nutrient rich media during vegetative growth cells reproduce asexually by binary fission. This type of cell division occurs by a sequence of morphogenetic events that results in the development of duplicate sets of cell structures, one for each daughter cell. Only during starvation conditions will cells commit to sexual conjugation, pairing and fusing with a cell of opposite mating type. Tetrahymena has seven mating types; each of which can mate with any of the other six without preference, but not its own.
Typical of ciliates, T. thermophila differentiates its genome into two functionally distinct types of nuclei, each specifically used during the two different stages of the life cycle. The diploid germline micronucleus is transcriptionally silent and only plays a role during sexual life stages. The germline nucleus contains 5 pairs of chromosomes which encode the heritable information passed down from one sexual generation to the next. During sexual conjugation, haploid micronuclear meiotic products from both parental cells fuse, leading to the creation of a new micro- and macronucleus in progeny cells. Sexual conjugation occurs when cells starved for at least 2hrs in a nutrient-depleted media encounter a cell of complementary mating type. After a brief period of co-stimulation (~1hr), starved cells begin to pair at their anterior ends to form a specialized region of membrane called the conjugation junction.
It is at this junctional zone that several hundred fusion pores form, allowing for the mutual exchange of protein, RNA and eventually a meiotic product of their micronucleus. This whole process takes about 12 hours at 30 °C, but even longer than this at cooler temperatures. The sequence of events during conjugation is outlined in the accompanying figure.[10]
The larger polyploid macronucleus is transcriptionally active, meaning its genes are actively expressed, and so it controls somatic cell functions during vegetative growth. The polyploid nature of the macronucleus refers to the fact that it contains approximately 200–300 autonomously replicating linear DNA mini-chromosomes. These minichromosomes have their own telomeres and are derived via site-specific fragmentation of the five original micronuclear chromosomes during sexual development. In T. thermophila each of these minichromosomes encodes multiple genes and exists at a copy number of approximately 45-50 within the macronucleus. The exception to this is the minichromosome encoding the rDNA, which is massively upregulated, existing at a copy number of approximately 10,000 within the macronucleus. Because the macronucleus divides amitotically during binary fission, these minichromosomes are un-equally divided between the clonal daughter cells. Through natural or artificial selection, this method of DNA partitioning in the somatic genome can lead to clonal cell lines with different macronuclear phenotypes fixed for a particular trait, in a process called phenotypic assortment. In this way, the polyploid genome can fine-tune its adaptation to environmental conditions through gain of beneficial mutations on any given mini-chromosome whose replication is then selected for, or conversely, loss of a minichromosome which accrues a negative mutation. However, the macronucleus is only propagated from one cell to the next during the asexual, vegetative stage of the life cycle, and so it is never directly inherited by sexual progeny. Only beneficial mutations that occur in the germline micronucleus of T. thermophila are passed down between generations, but these mutations would never be selected for environmentally in the parental cells because they are not expressed.[11]
Free swimming cells of Tetrahymena are attracted to certain chemicals by chemokinesis. The major chemo-attractants are peptides and/or proteins.[12]
A 2016 study found that cultured Tetrahymena have the capacity to 'learn' the shape and size of their swimming space. Cells confined in a droplet of a water for a short time were, upon release, found to repeat the circular swimming trajectories 'learned' in the droplet. The diameter and duration of these swimming paths reflected the size of the droplet and time allowed to adapt.[13]
It is common among protists that the sexual cycle is inducible by stressful conditions such as starvation.[14] Such conditions often cause DNA damage. A central feature of meiosis is homologous recombination between non-sister chromosomes. In T. thermophila this process of meiotic recombination may be beneficial for repairing DNA damages caused by starvation.
Exposure of T. thermophila to UV light resulted in a greater than 100-fold increase in Rad51 gene expression.[15] Treatment with the DNA alkylating agent methyl methanesulfonate also resulted in substantially elevated Rad 51 protein levels. These findings suggest that ciliates such as T. thermophila utilize a Rad51-dependent recombinational pathway to repair damaged DNA.
The Rad51 recombinase of T. thermophila is a homolog of the Escherichia coli RecA recombinase. In T. thermophila, Rad51 participates in homologous recombination during mitosis, meiosis and in the repair of double-strand breaks.[16] During conjugation, Rad51 is necessary for completion of meiosis. Meiosis in T. thermophila appears to employ a Mus81-dependent pathway that does not use a synaptonemal complex and is considered secondary in most other model eukaryotes.[17] This pathway includes the Mus81 resolvase and the Sgs1 helicase. The Sgs1 helicase appears to promote the non-crossover outcome of meiotic recombinational repair of DNA,[18] a pathway that generates little genetic variation.
Many species of Tetrahymena are known to display unique response mechanisms to stress and various environmental pressures. The unique genomic architecture of the ciliates (presence of a MIC, high ploidy, large number of chromosomes, etc.) allows for differential gene expression, as well as increased genomic flexibility. The following is a non-exhaustive list of examples of phenotypic and genotypic plasticity in the Tetrahymena genus.
T. vorax is known for its inducible trophic polymorphisms, an ecologically offensive tactic that allows it to change its feeding strategy and diet by altering its morphology.[19] Normally, T. vorax is a bacterivorous microstome around 60 μm in length. However, it has the ability to switch into a carnivorous macrostome around 200 μm in length that can feed on larger competitors. If T. vorax cells are too nutrient starved to undertake transformation, they have also been recorded as transforming into a third “tailed”-microstome morph, thought to be a defense mechanism in response to cannibalistic pressure. While T. vorax is the most well studied Tetrahymena that exhibits inducible trophic polymorphisms, many lesser known species are able to undertake transformation as well, including T. paulina and T. paravorax.[20] However, only T. vorax has been recorded as having both a macrostome and tailed-microstome form.
These morphological switches are triggered by an abundance of stomatin in the environment, a mixture of metabolic compounds released by competitor species, such as Paramecium, Colpidium, and other Tetrahymena. Specifically, chromatographic analysis has revealed that ferrous iron, hypoxanthine, and uracil are the chemicals in stomatin responsible for triggering the morphological change.[21] Many researchers cite “starvation conditions” as inducing the transformation, as in nature, the compound inducers are in highest concentration after microstomal ciliates have grazed down bacterial populations, and ciliates populations are high. When the chemical inducers are in high concentration, T. vorax cells will transform at higher rates, allowing them to prey on their former trophic competitors.
The exact genetic, and structural mechanisms that underlie T. vorax transformation are unknown. However, some progress has been made in identifying candidate genes. Researchers from the University of Alabama have used cDNA subtraction to remove actively transcribed DNA from microstome and macrostome T. vorax cells, leaving only differentially transcribed cDNA molecules.[22] While nine differentiation-specific genes were found, the most frequently expressed candidate gene was identified as a novel sequence, SUBII-TG.
The sequenced region of SUBII-TG was 912 bp long and consists of three largely identical 105 bp open-reading frames. A northern blot analysis revealed that low levels of transcription are detected in microstome cells, while high levels of transcription occur in macrostome cells. Furthermore, when the researchers limited SUBII-TG expression in the presence of stomatin (using antisense oligonucleotide methods), a 55% reduction in SUBII-TG mRNA correlated with a 51% decrease in transformation, supporting the notion that the gene is at least partially responsible for controlling the transformation in T. vorax. However, very little is known about the SUBII-TG gene. Researchers were only able to sequence a portion of the entire open-reading frame, and other candidate genes have not been investigated thoroughly. mRNA and amino acid sequencing indicate that ubiquitin may play a crucial role in allowing transformation to take place as well. However, no known genes in the ubiquitin family have been identified in T. vorax.[23] Finally, the genetic mechanisms of the “tailed” microstome morph are completely unknown.
Other related species exhibit their own unique responses to various stressors. In T. thermophila, chromosome amplification and gene expansion are inducible responses to common organometallic pollutants such as Cadmium, Copper, and Lead.[24] Strains of T. thermophila that were exposed to large quantities of Cd2+ over time were found to have a 5-fold increase of MTT1, and MTT3 (metallothionein genes that code for Cadmium and Lead binding proteins) as well as CNBDP, an unrelated gene that lies just upstream of MTT1 on the same chromosome. The fact that a non-metallothionein gene on the same locus as MTT1 and MTT3 increased copy number indicates that the entire chromosome had been amplified, as opposed to just specific genes. Tetrahymena species are 45-ploid for their macronucleus, meaning that the wild type of T. thermophila normally contains 45 copies of each chromosome. While the actual number of unique chromosomes are unknown, the number is thought to be around 187 in the MAC, and 5 in the MIC.[25] Thus, the Ca2+ adapted strain contained 225 copies of the specific chromosome in question. This resulted in a nearly 28 times increase in detected expression levels of MTT1, and slightly less in MTT3.
Interestingly, when researchers grew a sample of the T. thermophila population in normal growth medium (lacking Cd2+) for 1 month, the number of MTT1, MTT3, and CNBDP genes decreased to an average of 3 copies (135C). By 7 months in normal growth medium, the T. thermophila cells were found reduced to just the wild type copy number (45C). When researchers returned cells from the same colony to Cd2+ medium, within a week MTT1, MTT3, and CNBDP genes increased to 3 copies once again (135C). Thus, the authors argue that chromosome amplification is an inducible and reversible mechanism in the Tetrahymena genetic response to metal stress.
Researchers also used gene-knockdown experiments, where the copy number of another metallothionein gene on a different chromosome, MTT5, was dramatically reduced. Within a week, the new strain was found to have developed 4 novel genes from at least 1 duplication of MTT1. However, chromosome duplication had not taken place, as indicated by the wild-type ploidy and the normal quantity of other genes on the same chromosomes. Rather, researchers believe that the duplication resulted from homologous recombination events, producing transcriptionally active, upregulated genes that carry repeated MTT1.
T. thermophila also undergoes phenotypic changes when faced with limited resource availability. Cells are capable of changing their shape and size, along with behavioral swimming strategies in response to starvation.[26] The more motile cells that change in response to starvation are known as dispersers, or disperser cells. While rates and levels of phenotypic change differ between strains, disperser cells form in nearly all strains of T. thermophila when faced with starvation. Dispersers, and non-dispersing cells both become dramatically thinner and smaller, increasing the basal body and cilia density, allowing them to swim between 2 and 3 times faster than normal cells.[27] Some strains of T. thermophila have also been found to develop a single, non-beating, enlarged cilia that assists the cell in steering or directing movement. While the behavior has been shown to correlate with faster dispersal and form as a reversible trait in Tetrahymena cells, little is known about the genetic or cellular mechanisms that allow for its development. Furthermore, other studies show that when genetically variable populations of T. thermophila were starved, dispersal cells actually increased in cell length, despite still becoming thinner.[28] More research is needed to determine the genetic mechanisms that underlie disperser formation.
Species in this genus include.[1]
Cornell University offers a National Institutes of Health (NIH) funded program through the Science Education Partnership Award (SEPA) Program called Advancing Secondary Science Education thru Tetrahymena (ASSET).[29] The group develops stand-alone labs or lessons using Tetrahymena as training modules that teachers can use in classes.
Tetrahymena, a unicellular eukaryote, is a genus of free-living ciliates. The genus Tetrahymena is the most widely studied member of its phylum.: 59 It can produce, store and react with different types of hormones. Tetrahymena cells can recognize both related and hostile cells.
They can also switch from commensalistic to pathogenic modes of survival. They are common in freshwater lakes, ponds, and streams.: 277
Tetrahymena species used as model organisms in biomedical research are T. thermophila and T. pyriformis.
Tetrahymena es un género de protozoos ciliados, que se encuentran naturalmente en aguas dulces. Las especies T. termophila y T. pyriformis son ampliamente empleadas como modelos de investigaciones biomédicas.[1]
Como protozoo ciciliado, T. thermophila exhibe un marcado dimorfismo nuclear. Posee dos tipos de núcleo celular: un gran macronúcleo, y un pequeño micronúcleo, que coexisten en una sola célula y cumplen diversas funciones con propiedades bioquímicas y citológicas diferentes. Esta versatilidad única permite a los científicos usar la Tetrahymena para identificar factores clave relacionados con la expresión de los genes y la integridad del genoma. Además, posee cientos de cilias y complicadas estructuras de microtúbulos. Esto lo convierte en el modelo ideal para elucidar la diversidad de funciones de los sistemas de microtúbulos.
Ya que Tetrahymena puede ser fácilmente cultivada en grandes cantidades en los laboratorios, ha sido por años una gran fuente de análisis de actividades enzimáticas importantes y para la purificación de componentes subcelulares. También se han desarrollado técnicas de genética molecular avanzadas, incluyendo la transformación mediada por ADN, el knock-out y knock-in de genes por recombinación homóloga, marcado de epítopos, y la expresión de genes inducible y reprimible. Estas características lo convierten en un excelente modelo para el estudio de funciones genéticas in vivo. Recientemente se secuenció completamente el genoma macronuclear, lo que promete que se continúe usando la Tetrahymena como sistema modelo en la era genómica y post-genómica.
Los estudios de este microorganismo han contribuido en varios avances científicos:
Científicos argentinos habrían encontrado un método para emplear la Tetrahymena para eliminar el 90 % del colesterol de la leche y huevos, y convertir un 5 % en pro-vitamina D. De esta forma podrían crearse nuevos productos nutracéuticos (alimentos que posee un adicional beneficio a la salud), por el bajo colesterol por un lado, y por la inclusión de pro-vitamina D por el otro.[2][3]
Tetrahymena es un género de protozoos ciliados, que se encuentran naturalmente en aguas dulces. Las especies T. termophila y T. pyriformis son ampliamente empleadas como modelos de investigaciones biomédicas.
Tetrahymena est un genre de ciliés pouvant être commensaux ou pathogènes. Les espèces sont très communes dans l'eau douce. Ses dimensions normales sont de 50 µm de longueur et de 20 µm de largeur[1].
Les chercheurs en biochimie utilisent les espèces Tetrahymena thermophila[2] et Tetrahymena pyriformis[3] comme organisme modèle. On a par exemple montré que cette espèce dispose de récepteurs hormonaux, dont l'un induit la production de mélatonine par le protozoaire, quand il est exposé à de faibles dose de cette hormone, notamment impliqué dans la photoadaptation[4] avec des effets d'attractivité vers la lumière le jour[5], de chemotactisme la nuit[5] et autres[5].
Tetrahymena thermophila a la particularité d'exister en 7 sexes (types sexuels) différents, ce qui donne 21 différentes combinaisons possibles d'accouplement, chacun de ces sexes étant incompatible avec lui-même. Chaque organisme "décide" via un processus stochastique du sexe qu'il aura durant l'accouplement[6].
Chez le poisson guppy, la Tetrahymena est responsable d'une maladie[7] nommée Guppy Killers.
Le cycle de vie de la Tetrahymena est en enchaînement de stades sexués et asexués. En milieu riche en nutriments, pendant la croissance végétative, les cellules se reproduisent asexuellement par fission binaire. Ce type de division cellulaire se produit par une séquence d'évènements morphogénétiques qui aboutit au développement d'ensembles de structures cellulaires en double, un pour chaque cellule fille. Ce n'est que pendant les conditions de famine que les cellules s'engagent dans la conjugaison sexuelle, l'appariement et la fusion avec une cellule de type d'accouplement opposé. La tetrahymena a sept types d'accouplement ; chacun d'entre eux peut s'accoupler avec l'un des six autres sans préférence, mais pas avec son propre type.
Typique des ciliés, T. thermophila différencie son génome en deux types de noyaux fonctionnellement distincts, chacun utilisé spécifiquement pendant les deux différentes étapes du cycle de vie. Le micronoyau de la lignée germinale diploïde est silencieux sur le plan de la transcription et ne joue un rôle que pendant les étapes de la vie sexuelle. Le noyau germinal contient 5 paires de chromosomes qui codent les informations héréditaires transmises d'une génération sexuelle à l'autre. Lors de la conjugaison sexuelle, les produits méiotiques des micro-nucléaires haploïdes des deux cellules parentales fusionnent, entraînant la création d'un nouveau micro et macro-noyau dans les cellules de la progéniture. La conjugaison sexuelle se produit lorsque des cellules affamées pendant au moins 2 heures dans un milieu pauvre en nutriments rencontrent une cellule de type complémentaire. Après une brève période de costimulation d'environ 1 heure, les cellules affamées commencent à s'accoupler à leurs extrémités antérieures pour former une région spécialisée de la membrane appelée jonction de conjugaison.
Deux cellules de tetrahymena qui sont de types d'accouplement complémentaires s'accouplent pour échanger leurs noyaux pendant la conjugaison sexuelle.
C'est à cette zone de jonction que se forment plusieurs centaines de pores de fusion, permettant l'échange mutuel de protéines, d'ARN et éventuellement d'un produit méiotique de leur micro-noyau. Tout ce processus prend environ 12 heures à 30 °C, mais encore plus longtemps à des températures plus basses. La séquence des évènements pendant la conjugaison est décrite en anglais dans la figure ci-jointe (figure "cycle du tetrahymena")[8].
Selon ITIS:
Tetrahymena est connu comme étant un cas exceptionnel en biologie moléculaire. En effet, l'épissage des introns de certains ARN précurseurs est catalysé par l'ARN transcrit lui-même. Chez Tetrahymena, le précurseur de l'ARN ribosomique contient un intron qui est capable de s'épisser seul, en l'absence de facteurs externes. Dans l'immense majorité autres eucaryotes, c'est le splicéosome qui remplit cette fonction en trans. On a trouvé depuis ce type d'introns auto-épissables chez d'autres ciliés, dans des gènes mitochondriaux et chez certains virus comme le bactériophage T4). Cette découverte a permis de démontrer que, pour la première fois, une molécule d'ARN pouvait, dans certains cas, agir comme une enzyme et catalyser une réaction biochimique spécifique[9]. Ces ARN à action enzymatique ont été appelés ribozymes. Le ribosome (l'ARN ribosomique de la grande sous unité) est un ribozyme.
La tetrahymena a été un modèle utile pour la recherche fondamentale notamment parce qu'elle est facile à cultiver et qu'elle présente une série de processus complexes, le tout au sein d'une seule cellule. Pour ces mêmes raisons, tetrahymena a montré un énorme potentiel en tant qu'outil d'enseignement des principes fondamentaux de la biologie à de multiples niveaux d'enseignement scientifique[10].
L'élaboration spéciale de certains mécanismes eucaryotes de base du tetrahymena ont facilité des découvertes notamment :
- Première cellule dont la division a été synchronisée, ce qui a permis de découvrir l'existence de mécanismes de contrôle du cycle cellulaire[11].
- Identification et purification du premier moteur cytosquelettique, la dynéine, et détermination de l'activité directionnelle[11].
- Participation à la découverte des lysosomes et des peroxysomes.[11]
- L'une des premières descriptions moléculaires du réarrangement programmé du génome somatique[11].
- Découverte de la structure moléculaire des télomères, de l'enzyme télomérase, du rôle de modèle de l'ARN télomérase et de leurs rôles dans la sénescence cellulaire et la guérison des chromosomes.[11]
- Co-découverte d'ARN catalytique (ribozymes), récompensée par le prix Nobel[11]
- Découverte de la fonction d'acétylation des histones dans la transcription[11].
Tetrahymena est un genre de ciliés pouvant être commensaux ou pathogènes. Les espèces sont très communes dans l'eau douce. Ses dimensions normales sont de 50 µm de longueur et de 20 µm de largeur.
Les chercheurs en biochimie utilisent les espèces Tetrahymena thermophila et Tetrahymena pyriformis comme organisme modèle. On a par exemple montré que cette espèce dispose de récepteurs hormonaux, dont l'un induit la production de mélatonine par le protozoaire, quand il est exposé à de faibles dose de cette hormone, notamment impliqué dans la photoadaptation avec des effets d'attractivité vers la lumière le jour, de chemotactisme la nuit et autres.
Tetrahymena thermophila a la particularité d'exister en 7 sexes (types sexuels) différents, ce qui donne 21 différentes combinaisons possibles d'accouplement, chacun de ces sexes étant incompatible avec lui-même. Chaque organisme "décide" via un processus stochastique du sexe qu'il aura durant l'accouplement.
Chez le poisson guppy, la Tetrahymena est responsable d'une maladie nommée Guppy Killers.
Tetrahymena é un xénero de protozoos ciliados de vida libre. En certos caos pode tamén adoptar unha forma de vida comensal ou patoxénica; por exemplo, no peixe guppy Tetrahymena pode causar unha doenza mortal[1]. Son comúns en estanques de auga doce. Algunhas especies de Tetrahymena utilízanse como organismos modelo en investigacións biomédicas, como T. thermophila e T. pyriformis.[2] Entre os descubrimentos realizados estudando Tetrahymena salientaremos o da estrutura dos telómeros e a telomerase, o autosplicing do ARN, as primeiras proteínas motoras e a regulación xénica por acetilación das histonas.
Tetrahymena ten dous tipos de núcleos (dimorfismo nuclear), un pequeno (micronúcleo), diploide usado para a reprodución e outro máis grande (macronúcleo) poliploide para o funcionamento da célula. O macronúcleo divídese por amitose e periodicamente debe ser rexenerado a partir do micronúcleo. Ten tamén reprodución por conxugación.
As especies do xénero Tertrahymena teñen un tamaño de 30-60 microns e están cubertas completamente por unha ciliatura típica de holotrico. A ciliatura sérvelle para a locomoción e é moi sensible aos estímulos sensoriais. O organismo ten un aparato oral de grande tamaño, comparable ao do paramecio. As catro membranas que ten na zona oral danlle o nome a este xénero (tetra = catro, hymen = membrana); estas catro membranas están situadas a esquerda e dereita, tres membranas de tipo membranela a un lado e unha membrana ondulante no outro. A célula presenta un citoprocto para a excreción de residuos.
Tetrahymena é un xénero de protozoos ciliados de vida libre. En certos caos pode tamén adoptar unha forma de vida comensal ou patoxénica; por exemplo, no peixe guppy Tetrahymena pode causar unha doenza mortal. Son comúns en estanques de auga doce. Algunhas especies de Tetrahymena utilízanse como organismos modelo en investigacións biomédicas, como T. thermophila e T. pyriformis. Entre os descubrimentos realizados estudando Tetrahymena salientaremos o da estrutura dos telómeros e a telomerase, o autosplicing do ARN, as primeiras proteínas motoras e a regulación xénica por acetilación das histonas.
Tetrahymena ten dous tipos de núcleos (dimorfismo nuclear), un pequeno (micronúcleo), diploide usado para a reprodución e outro máis grande (macronúcleo) poliploide para o funcionamento da célula. O macronúcleo divídese por amitose e periodicamente debe ser rexenerado a partir do micronúcleo. Ten tamén reprodución por conxugación.
As especies do xénero Tertrahymena teñen un tamaño de 30-60 microns e están cubertas completamente por unha ciliatura típica de holotrico. A ciliatura sérvelle para a locomoción e é moi sensible aos estímulos sensoriais. O organismo ten un aparato oral de grande tamaño, comparable ao do paramecio. As catro membranas que ten na zona oral danlle o nome a este xénero (tetra = catro, hymen = membrana); estas catro membranas están situadas a esquerda e dereita, tres membranas de tipo membranela a un lado e unha membrana ondulante no outro. A célula presenta un citoprocto para a excreción de residuos.
Tetrahymena è un genere di protisti ciliati comuni in acqua dolce. Le due specie tetrahymena thermophila e tetrahymena pyriformis sono utilizzate come organismi modello nella ricerca biomedica.[1]
A differenza degli altri eucarioti e come per gli altri ciliati, tetrahymena ha due tipi differenti di nuclei (dimorfismo nucleare), uno piccolo (micronucleo), diploide usato per la riproduzione e uno più grande (macronucleo) poliploide per il funzionamento della cellula. Il macronucleo è prodotto dal micronucleo per ampliamento del genoma. La sua divisione avviene per amitosi. Periodicamente il macronucleo deve essere rigenerato dal micronucleo. In molti casi questo avviene attraverso la riproduzione sessuale che avviene usualmente per coniugazione.
Come protozoo ciliato, Tetrahymena thermophila esibisce dimorfismo nucleare: due tipi di nuclei cellulari. Essi hanno un macronucleo più grande, non della linea germinale, e un piccolo micronucleo, della linea germinale, in ciascuna cellula allo stesso tempo ed entrambi eseguono funzioni diverse con distinte proprietà citologiche e biologiche. Questa versatilità unica permette agli scienziati di usare Tetrahymena per identificare parecchi fattori chiave riguardanti l'espressione genica e l'integrità genomica. Inoltre, Tetrahymena possiede dozzine di ciglia ed ha complicate strutture microtubulari, che lo rendono un modello ottimale per illustrare la diversità e le funzioni delle disposizioni microtubulari.
Poiché Tetrahymena può essere coltivata facilmente in grande quantità nel laboratorio, essa è da anni una grande fonte per le analisi biochimiche, specificamente per le attività enzimatiche e per la purificazione dei componenti sub-cellulari. Inoltre, con l'avanzamento delle tecniche genetiche è diventata un eccellente modello per studiare la funzione genica in vivo. Il recente sequenziamento del genoma del macronucleo dovrebbe garantire che Tetrahymena continuerà ad essere utilizzata come sistema modello.
Tetrahymena thermophila può esistere in 7 diversi sessi che possono riprodursi in 21 combinazioni diverse, e una singola tetrahymena non può riprodursi sessualmente con sé stessa.
Gli studi su Tetrahymena hanno contribuito a parecchie pietre miliari scientifiche tra cui:
Tetrahymena è un genere di protisti ciliati comuni in acqua dolce. Le due specie tetrahymena thermophila e tetrahymena pyriformis sono utilizzate come organismi modello nella ricerca biomedica.
A differenza degli altri eucarioti e come per gli altri ciliati, tetrahymena ha due tipi differenti di nuclei (dimorfismo nucleare), uno piccolo (micronucleo), diploide usato per la riproduzione e uno più grande (macronucleo) poliploide per il funzionamento della cellula. Il macronucleo è prodotto dal micronucleo per ampliamento del genoma. La sua divisione avviene per amitosi. Periodicamente il macronucleo deve essere rigenerato dal micronucleo. In molti casi questo avviene attraverso la riproduzione sessuale che avviene usualmente per coniugazione.

Tetrahymena – rodzaj orzęsków. Występuje u niego 7 płci (oznaczanych I, II, III, IV, V, VI i VII), z których każda może współżyć z każdą inną. Orzęski różnej płci wyglądają tak samo, ale nie występują równie często.
U Tetrahymena płeć kontroluje gen zwany "mat". Każdy jego allel nie decyduje o płci, tylko określa jej prawdopodobieństwo. W sumie jest co najmniej 14 alleli, podzielonych na 2 główne grupy: A i B. W grupie A możliwa jest każda płeć poza IV i VII, w B każda inna od I.
Tetrahymena ma dwa jądra komórkowe: małe (mikronukleus) i duże (makronukleus). Makronukleus kontroluje codzienne funkcje życiowe, mikronukleus odpowiada za życie seksualne. Podczas połączenia dwóch orzęsków, zwanego koniugacją, mikronukleusy wymieniają geny. Po tej wymianie powstają nowe makronukleusy. Później zmodyfikowane koniugacją orzeski mogą się rozmnażać przez podział.
Jest pierwszym organizmem, u którego wykryto (w 1982 roku), a następnie opisano mechanizm autokatalitycznego wycinania intronów (tzw. self-splicing)[1].
Tetrahymena – rodzaj orzęsków. Występuje u niego 7 płci (oznaczanych I, II, III, IV, V, VI i VII), z których każda może współżyć z każdą inną. Orzęski różnej płci wyglądają tak samo, ale nie występują równie często.
U Tetrahymena płeć kontroluje gen zwany "mat". Każdy jego allel nie decyduje o płci, tylko określa jej prawdopodobieństwo. W sumie jest co najmniej 14 alleli, podzielonych na 2 główne grupy: A i B. W grupie A możliwa jest każda płeć poza IV i VII, w B każda inna od I.
Tetrahymena ma dwa jądra komórkowe: małe (mikronukleus) i duże (makronukleus). Makronukleus kontroluje codzienne funkcje życiowe, mikronukleus odpowiada za życie seksualne. Podczas połączenia dwóch orzęsków, zwanego koniugacją, mikronukleusy wymieniają geny. Po tej wymianie powstają nowe makronukleusy. Później zmodyfikowane koniugacją orzeski mogą się rozmnażać przez podział.
Jest pierwszym organizmem, u którego wykryto (w 1982 roku), a następnie opisano mechanizm autokatalitycznego wycinania intronów (tzw. self-splicing).
Tetrahymena é um gênero de protozoários ciliados não patogênicos de vida livre cujos integrantes são encontrados principalmente em água fresca. As espécies Tetrahymena thermophila e Tetrahymena pyriformis são amplamente utilizadas em pesquisas medicinais e biológicas.
O ciclo de vida da T. thermophila é constituído por uma alternância entre fases sexuadas e assexuadas. Em ambientes favoráveis e ricos em nutrientes, as células reproduzem-se por bipartição de forma assexuada. Esse tipo de divisão celular resulta no desenvolvimento de conjuntos duplicados de estruturas celulares, um para cada célula-filha. Apenas quando há escassez de alimento é que as células irão reproduzir-se sexuadamente conjuntamente com outra célula de sexo oposto. A T. thermophila possui sete tipos de sexos diferentes, cada um dos quais capaz de se reproduzir com qualquer um dos outros seis, sem preferência.
T. thermophila diferencia o seu genoma em dois tipos de núcleos, o macronúcleo, responsável por regular o metabolismo celular, e o micronúcleo, cujo genoma permanece silenciado a maior parte do tempo, servindo apenas para a reprodução sexual. O micronúcleo contém 5 pares de cromossomas que codificam a informação genética que é transmitida pela célula para os seus descendentes. Durante a conjugação sexual, os micronúcleos haplóides de cada célula, decorrentes da divisão meiótica de cada micronúcleo original, fundem-se, levando à criação de novos micronúcleos e macronúcleos em cada uma das células. As duas células começam por formar poros na sua zona de junção, permitindo a troca mútua de proteínas, RNA e, eventualmente os produtos da meiose dos micronúcleos. O processo leva cerca de 12 horas a 30°C, mas ainda mais que isso a temperaturas mais baixas.[1]
Tetrahymena é um gênero de protozoários ciliados não patogênicos de vida livre cujos integrantes são encontrados principalmente em água fresca. As espécies Tetrahymena thermophila e Tetrahymena pyriformis são amplamente utilizadas em pesquisas medicinais e biológicas.
Tetrahymena thermophila là một sinh vật lạ, nhân chuẩn đơn bào hình trứng, được tìm thấy trong môi trường nước ngọt. T. thermophila có đến 7 giới tính khác nhau, khi giao phối, có bảy loại gen ghép cặp quy định giới tính. Ngoài ra, sinh vật này cũng có thể sinh sản vô tính bằng phương pháp tự nhân đôi.[1]
T.thermophila là một tế bào có hai nhân quan trọng: nhân nhỏ chứa vật liệu cần thiết cho sự sinh sản; nhân to tạo tính mềm dẻo cho vi sinh vật, chứa ít nhất 25.000 gen năm trên khoảng 200 nhiễm sắc thể.
Mỗi tế bào T.thermophila đều có 2 gen, một gen kiểm soát hoạt động hằng ngày của vi sinh vật, gen còn lại đóng vai trò như buồng trứng và tinh hoàn ở động vật. Gen thứ 2 chứa các cặp gen không hoàn chỉnh của 6 hoặc 7 giới tính. Khi hai vi khuẩn giao phối, tùy theo kết quả gép đôi của các gen quy định giới tính sẽ quyết định giới tính ở thế hệ sau. Giới tính của thế hệ sau cũng có thể mang giới tính như cha/mẹ, hoặc khác.
Mỗi con Tetrahymena đều có giới tính riêng, mô hình giao phối riêng ở trên vỏ tế bào. Khác với các sinh vật đơn bào khác, sinh vật này có khả năng kết đôi đặc biệt giúp nhằm tăng cơ hội sinh sản trong tự nhiên. T.thermophila không có thói quen quan hệ tình dục đồng giới, sinh sản bằng phương pháp phân đôi và giao phối khác giới. Hầu hết trường hợp, T.thermophila sinh sản vô tính, một tế bào mẹ phân chia thành 2 tế bào con. Khi nguồn thức ăn khan hiếm, chúng có khuynh hướng kết đôi để sinh sản. Khi 2 tế bào kết hợp, tạo ra ADN chung, thì 2 tế bào còn lại rã đám. “Đó là trường hợp giao phối mà không sinh sản”, theo chuyên gia Orias. Phát hiện này đã giúp các nhà khoa học hiểu thêm về các tế bào khác bao gồm cả các tế bào trong cơ thể con người.
Khi T.thermophila giao phối, có bảy loại gen ghép cặp quy định giới tính, do đó giới tính của thế hệ sau khác so với bố mẹ và có thể có tới 7 giới tính khác nhau. Sự hình thành giới tính đời sau được sắp đặt hoàn toàn ngẫu nhiên.
Khi giao phối, có bảy loại gen ghép cặp quy định giới tính, các loại gen ghép cặp sẽ thông qua quá trình chọn lọc và cuối cùng chỉ giữ lại một loại gen chuẩn nhất, sáu loại khác sẽ bị đào thải và ADN được điều chỉnh lại và sẽ quyết định giới tính của đời sau.[2]
Tetrahymena thermophila là một sinh vật lạ, nhân chuẩn đơn bào hình trứng, được tìm thấy trong môi trường nước ngọt. T. thermophila có đến 7 giới tính khác nhau, khi giao phối, có bảy loại gen ghép cặp quy định giới tính. Ngoài ra, sinh vật này cũng có thể sinh sản vô tính bằng phương pháp tự nhân đôi.
Tetrahymena
ВидыТетрахимены[источник не указан 24 дня] (лат. Tetrahymena) — род преимущественно свободноживущих пресноводных ресничных инфузорий, включающий около 40 валидных видов. Обычны в прудах среди гниющей листвы на дне водоёмов, но найдены также в водотоках и горячих источниках. Большинство видов — микрофаги, питающиеся бактериями, однако есть также хищные виды, питающиеся другими инфузориями. Для некоторых видов описан переход к комменсализму или факультативному (возможно, иногда и облигатному) паразитизму на пресноводных беспозвоночных — улитках, личинках комаров-звонцов и др. Некоторые виды рода Tetrahymena используются как модельные организмы в биологических и медицинских исследованиях, например, T. thermophila и T. pyriformis[1].
Как и всем инфузориям, Tetrahymena thermophila свойственен ядерный дуализм: у неё имеется два типа ядер — большое, соматическое (макронуклеус), и малое, половое (микронуклеус), которые находятся в клетке одновременно и имеют различные функции. Tetrahymena также имеет тысячи ресничек и сложные структуры цитоскелета (пелликулу, инфрацилиатуру и др.), что делает её идеальным модельным объектом для изучения систем цитоскелета.
Так как модельные виды Tetrahymena легко выращивать в больших количествах в лабораторных условиях, это отличный объект для биохимического анализа ферментов и выделения компонентов клетки. Разработаны молекулярно-генетические методы, которые позволяют модифицировать ДНК, убирать и встраивать гены путём гомологичной рекомбинации, индуцировать и репрессировать экспрессию генов, что делает тетрахимену идеальным объектом для изучения функции генов in vivo. После полного секвенирования генома макронуклеуса («начерно» осуществлённого в 2006 г.) Tetrahymena может быть использована как модельная система и в постгеномный период молекулярной биологии.
Изучение Tetrahymena внесло вклад в многие разделы биохимии и молекулярной биологии и позволило сделать ряд открытий:
Тетрахимены[источник не указан 24 дня] (лат. Tetrahymena) — род преимущественно свободноживущих пресноводных ресничных инфузорий, включающий около 40 валидных видов. Обычны в прудах среди гниющей листвы на дне водоёмов, но найдены также в водотоках и горячих источниках. Большинство видов — микрофаги, питающиеся бактериями, однако есть также хищные виды, питающиеся другими инфузориями. Для некоторых видов описан переход к комменсализму или факультативному (возможно, иногда и облигатному) паразитизму на пресноводных беспозвоночных — улитках, личинках комаров-звонцов и др. Некоторые виды рода Tetrahymena используются как модельные организмы в биологических и медицинских исследованиях, например, T. thermophila и T. pyriformis.
T hegewischi
T. hyperangularis
T. malaccensis
T. pigmentosa
T. pyriformis
T. thermophila
T. vorax
四膜蟲(Tetrahymena)是自由游離生活的(非寄生性的)纤毛原生動物,也可以從偏利共生轉變為病原生存模式。 它們在淡水池中很常見。在生物醫學研究中用作模式生物的四膜蟲屬是嗜熱棲熱菌(T. thermophila)和熱厭氧桿菌(T. pyriformis)[1]。
四膜蟲是一種單細胞真核生物,分布在全球的淡水水域中,屬於囊泡蟲類 (Alveolata)纖毛蟲门(Ciliophora),與一般人所熟知的草履蟲(Paramecium)在型態生理上十分相似。四膜蟲外觀呈橢圓長梨狀,體長約50微米,全身佈滿數百根長約4—6微米長的纖毛,纖毛排列成數十條縱列,是不同種間纖毛蟲分類的特徵之一。四膜蟲身體前端具有口器(oral apparatus),有三組三列的口部纖毛,早期在光學顯微鏡下觀察時看似有四列膜狀構造,因此據以命名。
四膜蟲主要是游離生活的異營生物,以攝取水中的細菌與其他有機質維生,尚未發現會造成人體疾病或對人類健康造成危害。
四膜蟲與草履蟲等其他纖毛蟲一樣,具有雙元核型(英语:nuclear dimorphism)(nuclear dimorphism):在一個細胞中有兩種类型的细胞核,小核負責生殖功能,一般生長時小核的基因並不會表現/表達;和大核(英语:macronucleus)(macronucleus)負責維持細胞生長營養所需,可觀察到旺盛的基因轉錄。
四膜蟲易於在實驗室裡培養,這歸功於研究人員已經找出可以適於四膜蟲生長所需的液態培養基成分,因此四膜蟲從早年開始即是一種實驗生物學上所使用的模式生物(model organism),用這種生物當作範例與工具,研究各種基礎生物學的現象。由於可以大量培養四膜蟲,所以它適於作為生化純化分析的材料來源。現代的分子生物技術與分子遺傳操作法也已經成功地使用在四膜蟲上,研究人員可以把DNA轉殖入四膜蟲細胞中,這些DNA可經由同源重組互換的方式將染色體上的基因剔除(knock-out),或在特定的基因座上將基因置入(knock-in),因此四膜蟲也適於藉由遺傳工程技術來解析基因的功能。近年來,四膜蟲大核的基因體(genome)也已經完成定序,所以在進入基因體時代的今日與後基因體時代,生物學家仍可以持續以四膜蟲為材料進行研究。
四膜虫交配时,其后代的性别可能和其父母的性别都不一样——有7种可能的性别。研究人员发现了四膜虫中决定其后代性别的复杂DNA特性,并确认了性别形成是随机的[2]。该结果发表在3月26日的《科学公共图书馆 — 生物学》上。
每一只四膜虫都有其自身性别或交配模式的基因——存在于其大核(或称体核)中,同时也带有另一个仅用于繁殖的小核。这种“生殖核”不完全包含7种交配型基因,经过剪切、粘贴的过程,最终会留下1个完整的基因,其他6种会被淘汰。重新调整过的DNA成为四膜虫下一代体核的一部分,并会决定其交配模式。
在四膜虫通过接合产生子代时,子代的体核在发育过程中需要在6种残缺的基因对中挑出一种,并组装成完整的基因对,才能使子代呈现对应的交配型。研究者发现,这一交配型选择过程是随机的——在子代的体核发育过程中,上述串联的交配型基因序列会发生一系列随机的剪切-连接反应。6种残缺的基因对中的5种会随着反应的进行被删掉,而剩下的一对则会与II型的MTA末端序列以及III型的MTB末端序列组装,形成具有功能的完整MTA/MTB基因对 。
四膜蟲(Tetrahymena)是自由游離生活的(非寄生性的)纤毛原生動物,也可以從偏利共生轉變為病原生存模式。 它們在淡水池中很常見。在生物醫學研究中用作模式生物的四膜蟲屬是嗜熱棲熱菌(T. thermophila)和熱厭氧桿菌(T. pyriformis)。

本文参照
テトラヒメナ(学名:Tetrahymena)は水中に生息する繊毛虫の属の一つ。洋梨型の形態をしており、体長は 30-100 μm 程度。多数の繊毛が生えており、これによって運動する。細胞内には収縮胞がある。細胞核は大核と小核にわかれており、小核は生殖核とも呼ばれる。小核は通常の染色体数をもち、細胞分裂のたびに受け継がれてゆく。大核は多細胞生物でいうと体細胞のような役割をしており、多倍体として生活に必要な遺伝子のコピー数を増やしている。大核には染色体の末端が多数存在するため、テロメア研究のモデル生物として用いられた。また触媒機能をもつ RNAリボザイムもテトラヒメナにおいて初めて発見された。
テトラヒメナ属の種
테트라하이메나(Tetrahymena)는 섬모충류에 속하는 원생동물이다. 편리 공생에서 병원체로 전환할 수도 있다. 이들은 일반적으로 민물에 산다. 생물의학 연구에서 모델 생물로 사용되는 테트라하이메나 종들은 T. thermophila와 T. pyriformis가 있다.[1]
Tetrahymena thermophila의 경우 7개의 성이 있으며 21개의 다른 조합으로 번식할 수 있다. 성은 성적(性的)으로 자가 생식하여 번식할 수 없다.
|volume=에 templatestyles stripmarker가 있음(위치 1) (도움말) 테트라하이메나(Tetrahymena)는 섬모충류에 속하는 원생동물이다. 편리 공생에서 병원체로 전환할 수도 있다. 이들은 일반적으로 민물에 산다. 생물의학 연구에서 모델 생물로 사용되는 테트라하이메나 종들은 T. thermophila와 T. pyriformis가 있다.
Tetrahymena thermophila의 경우 7개의 성이 있으며 21개의 다른 조합으로 번식할 수 있다. 성은 성적(性的)으로 자가 생식하여 번식할 수 없다.














