
Fires typically reduce the number of alderleaf mountain-mahogany shrubs, and as fire frequency increases the number
of shrubs in a stand decreases [102]. Prefire and postfire site conditions as well as fire behavior affect the
recovery of alderleaf mountain-mahogany on burned sites. In the following studies, alderleaf mountain-mahogany increased,
decreased, or remained the same on burned plots compared to unburned plots. Several studies related postfire
recovery to prefire and/or postfire site conditions while others correlated recovery with fire severity and/or
seasonality. Other studies lacked information on fire behavior or comparisons of burned and unburned or preburn
abundance.
Studies lacking information on fire behavior or burned and unburned comparisons:
Alderleaf mountain-mahogany sprouts required over 2 months recovery time after a July fire in southern California's
Anza-Borrego Desert State Park. Sprout and shrub number and productivity of C. m. var. glaber were
evaluated at postfire months 2, 4, 7, and 10. An estimated 25% of alderleaf mountain-mahogany shrubs suffered mortality.
There were no sprouts in postfire month 2, and the density of sprouts and productivity of shrubs decreased between
the 4th and 7th postfire sampling months. Production and shrub and sprout density increased substantially in the
growing season. Results from the study are summarized below [286]:
When individual alderleaf mountain-mahogany shrubs were burned with a kerosene flame gun, current annual growth
increased with dormant season burning and decreased with growing season burning [326,327]. The degree to
which this study imitated natural burning conditions is unknown. Curtin [49] reported that alderleaf mountain-mahogany
was significantly (p-value not reported) lower on burned than unburned sites 2 years following wildfires in
the Malpai Borderlands. Fire behavior was not described.
North
Pre- and postfire coverage of alderleaf mountain-mahogany was greater in younger Colorado pinyon-Utah
juniper stands with lower canopy cover than in older stands in northeastern Utah's Daggett
County. The behavior, season, or severity of the prescription fire were not described. Researchers
recommended that a fire frequency that maintains Colorado pinyon-Utah juniper cover below 25% is
best for the maintenance of alderleaf mountain-mahogany. Study findings are presented below [122]:
Postfire soils:
Available nitrogen and extractable phosphorus increased by 14 and 4 times, respectively, after a
fire in shrublands dominated by shrub live oak, alderleaf mountain-mahogany, and pointleaf manzanita in
central Arizona's Prescott National Forest. Before the fire, available nitrogen in the soil under true
mountain-mahogany shrubs was 5.4 mg/kg and phosphorus was 5.8 mg/kg. Relative humidity ranged from 10%
to 30%, temperatures were 61 to 84 °F (16-29 °C), and wind speeds were 5 to 18 miles (8-29 km)/hour when
the fire burned. Alderleaf mountain-mahogany fuel moisture was 61% to 72% at the time of the fire. At its most
"intense," fire flame heights were 10 feet (4 m), and fire spread was 39 feet (12 m)/minute.
Approximately 70% of available fuels were consumed. Following the fire, soils under alderleaf mountain-mahogany
had 71.6 mg/kg of available nitrogen and 22.6 mg/kg of available phosphorus [213]. Another study of postfire
soil nutrients in Arizona chaparral suggested that available nitrogen was unchanged and extractable
phosphorus increased following a fall prescription fire. Researchers noted, though, that the distribution
of available nitrogen and extractable phosphorus made it susceptible to erosion. Fire characteristics were
very similar in the 2 studies [320].
This description provides characteristics that may be relevant to fire ecology, and is not meant for identification. Keys for identification are available (e.g. [30,47,64,112,114,115,133,178,301,313,317]).
Aboveground description: Alderleaf mountain-mahogany is a shrub or less often a small tree 3 to 20 feet (1-7 m) tall [30,86,114]. Dayton [53] suggests that on "favorable" sites alderleaf mountain-mahogany can assume small tree stature. Few to many branches are produced, and the form is erect to spreading [112,178,200,262]. Stems measure 0.2 to 2 inches (0.5-5 cm) in diameter and are sturdy and stiff [151,301]. The main stem is typically less than 30 inches (80 cm) in diameter [112]. Researchers noted lignotubers on some shrubs in Nuttall's shrub oak-dominated chaparral vegetation in southern California [164]. Alderleaf mountain-mahogany bark is thin and smooth [7,47,104]. Alderleaf mountain-mahogany is considered long-lived [151], and stem diameter has been used to successfully age shrubs in Wyoming and Utah [34,37].
Deciduous, persistent, "mostly deciduous," and "occasionally evergreen" are all used to describe alderleaf mountain-mahogany [78,151,178,245,317]. Deciduousness is latitude and/or variety dependent. Little [172] indicates that C. m. var. var. paucidentatus and C. m. var. glaber are both evergreen, while Sampson and Jesperson [245] describe C. m. var. glaber as "occasionally evergreen." In the Sierra Nevada foothills of Tehama County, California, true mountain-mahogany loses 25% to 75% of its leaves each fall. Researchers think leaf loss may be a response to summer drought conditions and not freezing [27]. Leaves of alderleaf mountain-mahogany live 11.4 to 12 months in southern California [1]. In central Utah, alderleaf mountain-mahogany leaves have a 5- to 6-month lifespan [242].
Leaves are simple, alternate, and vary from thin to somewhat thick and firm [53,83,114,115,134,177]. Clusters of leaves are common on alderleaf mountain-mahogany's short spur-like branches [107,262]. Leaf shape is variable and ranges from lanceolate to obovate [114,115,200]. Often leaves are broadest at their midpoint [53]. Leaf blades are small and commonly length is less that 2 times the width [134]. Length ranges from 0.4 to 2 inches (1-5 cm), and width ranges from 0.2 to 1 inch (0.5-2.5 cm) [47,78,114,115,151,200]. Margins are entire or toothed [107,301]. Many describe margins that are toothed only on the upper 50% or 33% of the leaf [30,50,200]. Two to four teeth per centimeter of margin is described by Stephens [262]. Leaves are only slightly, if at all, revolute [83].
Alderleaf mountain-mahogany flowers are perfect and without petals [114,115,262]. Flowers occur singly or in small clusters of up to 12 on the upper portions of 2-year-old stems [50,112,151]. Clustered flowers are common in the axils of the short spur-like branches and 2 to 3 flowers together are most common [86,133,200,301]. Flower tubes measure 0.1 to 0.7 inch (3-17 mm) long [114,115,317]. The fruit is a hard, cylindrical achene coated with short hairs [83,86,114,115,151]. Achenes measure 0.3 to 0.5 inch (8-12 mm) long [47,104,262]. The average mass of alderleaf mountain-mahogany seed collected in southern California was 18 mg [138]. As is characteristic for the genus, true mountain-mahogany achenes have a persistent, somewhat twisted, hairy style or tail [7,50,83]. Styles are 1 to 4 inches (3-10 cm) long [78,107,112,317].
Underground description: The alderleaf mountain-mahogany root system is described as "extensive" [153]. Typically there are substantial tap and lateral roots [151]. Vegetative regeneration occurs from the root crown and from rhizomes [47,146]. Likely root penetration and spread are related to soil and site characteristics. Baker and Korstian [16] describe 1 to several taproots extending almost straight down with minimal spread. Rootlets on the taproots persisted to "great" depths. Taproots reached over 8 feet (2.5 m) deep. Based on data collected in 5 separate studies, alderleaf mountain-mahogany roots averaged 44.5 inches (113 cm) and ranged from 15.7 to 59.8 inches (40-152 cm) deep in northern New Mexico. The large stem base gave rise to "stout lateral roots" [74].
In California's San Gabriel Mountains, root systems of 2 young alderleaf mountain-mahogany shrubs that averaged 5 feet (1.5 m) tall had few branches and spread widely. The site had shallow soils overlaying rock. The majority of roots were concentrated in the shallow soil layer. The longest root measured 12.3 feet (3.75 m); maximum penetration was 5 feet (1.5 m), and maximum radial spread was 9 feet (2.7 m) [109]. From a 3-foot (1 m)-deep excavation in Nuttall's scrub oak-dominated chaparral in the Cleveland National Forest in California, researchers found that a substantial number of alderleaf mountain-mahogany roots grew vertically beyond 3 feet (1 m). Sixty-three percent of the roots were 16 to 39 inches (40-100 cm) deep. The site burned 9 years before and received its last herbicide application 5 years before the study. Researchers claimed that treated and untreated sites were not visually different [164].
Alderleaf mountain-mahogany roots growing in 3 different soil types in central Colorado's Manitou Experimental Forest showed little difference in maximum penetration and lateral spread. In Chubbs stony loam soils described as calcareous, loose, and fertile, a 4-foot (1 m) tall alderleaf mountain-mahogany shrub had roots penetrating to a maximum depth of 5 feet (1.5 m) and spreading laterally to a maximum of 10 feet (3 m). In Edole soils that had weak profile development, low fertility, and hard granite at 4.5- to 6-foot (1.4-1.8 m) depths, roots of a 4.5-foot (1.4 m) tall alderleaf mountain-mahogany penetrated to a maximum depth of 5 feet (1.5 m) and spread a maximum of 8 feet (2.4 m). In skeletal soils described as "little more than disintegrated parent rock," a 5-foot (1.5 m)-tall shrub had maximum root penetration of 3.5 feet (1.1 m) and maximum root spread of 5 feet (1.5 m). Based on provided root system sketches, most lateral roots were concentrated in the top 1 to 1.5 feet (0.3-0.5 m) of soil. Edole soil sites were dominated by ponderosa pine and Douglas-fir with some scattered quaking aspen (Populus tremuloides). Chubbs soil sites were ponderosa pine dominated, and Skeletal soil sites were Douglas-fir dominated [24].
Alderleaf mountain-mahogany plants can support nitrogen-fixing bacteria in root nodules. Young plants (most under 12 years) excavated from sites in southeastern and central Wyoming had nodules deep below the soil surface (>10 inches (30 cm)). Nodulated plants were limited to areas where soils were low in nitrate (< 1ug/g) and organic matter (<1%) [322]. From 2 plants in northern New Mexico, Aldon [3] identified 3 endomycorrhizal species and determined that 53% of the roots supported endomycorrhizae.
Variability: All the alderleaf mountain-mahogany varieties are described and a key for distinguishing varieties is available in [177]. The following references provide descriptions of 2 or more varieties.
Variety Reference C. m. var. argenteus [223,301] C. m. var. blancheae [112] C. m. var. glaber [112,172,223] C. m. var. macrourus [112] C. m. var. montanus [128,178] C. m. var. paucidentatus [128,172,178,223,301]Alderleaf mountain-mahogany may hybridize with curlleaf mountain-mahogany and littleleaf mountain-mahogany when distributions overlap. Alderleaf mountain-mahogany à curlleaf mountain-mahogany hybrids result only when alderleaf mountain-mahogany provides the ovule. Curlleaf mountain-mahogany typically flowers 2 weeks before alderleaf mountain-mahogany in overlapping habitats, and hybrids are only found in alderleaf mountain-mahogany stands. Rarely do alderleaf mountain-mahogany à curlleaf mountain-mahogany hybrids produce offspring [274,306]. True mountain-mahogany and littleleaf mountain-mahogany hybrids may be reciprocal in southeastern Utah. True mountain-mahogany à littleleaf mountain-mahogany hybrids are found in areas dominated by littleleaf mountain-mahogany [306].
Alderleaf mountain-mahogany à curlleaf mountain-mahogany hybrids grow in a tree-like form and have persistent leaves characteristic of curlleaf mountain-mahogany that are larger and have teeth like those characteristic of alderleaf mountain-mahogany in Utah's Uinta Basin [83,274,317]. Seed of alderleaf mountain-mahogany à curlleaf mountain-mahogany hybrids can be produced on the current year's growth, whereas alderleaf mountain-mahogany seed is produced exclusively on 2-year-old branches [306]. Alderleaf mountain-mahogany à curlleaf mountain-mahogany hybrids are susceptible to frost and drought, and often winter dieback is 50% or more [273]. Rarely do true mountain-mahogany à littleleaf mountain-mahogany hybrids produce flowers, and these hybrids are considered sterile [274].
Alderleaf mountain-mahogany occurs over a large portion of the western United States and northern Mexico. Alderleaf mountain-mahogany's distribution extends from southern Montana west to southwestern Oregon south to Baja California Norte and northern Mexico and north through the Texas panhandle, extreme western portions of Nebraska, and South Dakota [86,262]. Alderleaf mountain-mahogany is uncommon in Nevada and Idaho [124,133,260] and occurs only in the easternmost Great Basin ranges [47]. Only a single record exists for alderleaf mountain-mahogany in Kansas [262].
Distribution of varieties: Of the alderleaf mountain-mahogany varieties, C. m. var. montanus is the most widespread [151]. Cercocarpus montanus var. blancheae occurs on the Channel Islands and in the Western Traverse Range in California and is considered rare [112,177]. Plants Database provides a distributional map of alderleaf mountain-mahogany and its varieties. See Powell [223] for descriptions of the distributions of C. m. var. paucidentatus, C. m. var. glaber, and C. m. var. argenteus in the Trans-Pecos region of Texas. Kearney and Peebles [134] provide information on the distributions of C. m. var. paucidentatus and C. m. var. montanus in Arizona.
Fire adaptations: Often alderleaf mountain-mahogany is top-killed by fire. However, alderleaf mountain-mahogany quickly recolonizes burned sites through root crown or rhizome sprouts [47,146]. Postfire sprouting is described as "vigorous" and "rapid" [33] and is common under a range of FIRE REGIMES [1]. Alderleaf mountain-mahogany seed is typically killed by "intense" heat, and seedlings are rare in early postfire communities [146]. Hanes and Jones [102] report that seedlings may establish from off-site seed sources in early postfire communities, although Halsey [97] indicates that alderleaf mountain-mahogany seed germination is best in mature stands of chaparral. Fires typically reduce the number of alderleaf mountain-mahogany shrubs, and as the fire frequency increases, the number of shrubs in a stand decreases [102].
FIRE REGIMES: The alderleaf mountain-mahogany fire regime is dictated by its associated vegetation and distribution, both of which vary widely. In singleleaf pinyon, California juniper, and western juniper (Juniperus occidentalis) woodlands of the San Bernardino Mountains where alderleaf mountain-mahogany is common the fire cycle is an estimated 480 years [309]. Alderleaf mountain-mahogany is also a common shrub in mixed conifer and pinyon-juniper forests of New Mexico's El Malpais National Park where low-severity surface fires were common from ~1700 to 1880. An evaluation of 9 sites revealed an average fire return interval of 5 to 12 years [91].
Numerous studies of past and present FIRE REGIMES have been conducted in alderleaf mountain-mahogany habitats. Presented below is information on the fire history, fuel availability, and ignition sources and/or ignition probability in pinyon-juniper woodlands, mountain shrublands, mixed conifer forests, ponderosa pine forests, oak woodlands, and chaparral communities.
California chaparral: The FIRE REGIMES of southwestern chaparral, especially in southern California, have been investigated extensively. The lack of fire scars to reconstruct the fire history of these shrublands requires the use of alternate study methods. Fuel studies, ignition probabilities, postfire recovery of the vegetation, and ethnocentric histories are typically used to reconstruct the FIRE REGIMES. Using these methods there are still competing schools of thought about fire in chaparral systems.
Mature chaparral vegetation in southern and central California burns in stand-replacing fires. Rarely are herbaceous fuels sufficient to support surface fires, except maybe in the case of recently burned shrublands [142]. Ethnohistories indicate that the native people of California burned mixed chaparral vegetation primarily in the fall, but some spring burning occurred as well. "Spot burning" was routine and likely created and maintained openings and ecotones. A range of successional stages and a complex of habitats were probably normal under these burning practices [170].
California chaparral vegetation is comprised of a mixture of species, including those that sprout following fire but require a fire-free period for successful seedling establishment and those that require fire for seed scarification and successful seedling establishment. While a fire return interval of 20 years would be fine for species that germinate with fire, it is at odds with species requiring a fire-free period for the recruitment of new individuals. This conundrum suggests that the "long-term stability of chaparral communities may require a mosaic of fire frequencies" [139]. Not only does the fire return interval affect species recruitment; it may also affect species recovery. When chaparral vegetation burns frequently (~25 year fire return interval), the number of dead shrubs is typically low and fire severity will likely be low. Under these conditions, the potential for postfire sprouting is high and there are only small openings available for seedlings. When chaparral vegetation experiences long fire-free intervals (~100 years), there are more dead shrubs at time of the fire, and fire severity increases. Fewer postfire sprouts and increased mortality create larger openings on which seedlings may capitalize [136].
Postfire root crown and rhizome sprouting allow alderleaf mountain-mahogany to tolerate fire return intervals of 10 to 100 years. However, a fire-free interval of 100 years or more may be required for the successful recruitment of new individuals [137]. When considering all the species present in chaparral communities, Keeley and others [147] indicate that long fire-free periods are tolerated. Based on biological requirements and fire adaptations of chaparral species, Zedler [334] suggests that there is not an "urgent need to reduce the time between fires" in chaparral systems. See Fire management considerations for recommendations on managing fire in chaparral communities.
Competing philosophies of thought on California chaparral FIRE REGIMES: There are 2 prevailing philosophies about the best way to view and estimate natural FIRE REGIMES in California chaparral. One "deterministic" approach proposes that fire frequency is driven by fuel loadings and that ignitions are not limiting. Under this "school of thought," chaparral vegetation may burn every 15 to 20 years. The other philosophy suggests that FIRE REGIMES are dictated by a series of stochastic events, especially unpredictable ignitions. Proponents of this philosophy believe that "chaparral is resilient to a wide range of fire frequencies" [148].
Minnich, a proponent of the "deterministic" theory, suggests that FIRE REGIMES in southern California chaparral have changed with 20th century fire exclusion. While the fire return interval of approximately 40 to 60 years has not changed considerably with fire exclusion practices, fire size and behavior have changed [186]. Minnich describes an "erratic smolder and run" fire behavior before the 20th century. Behaviors of 3 chaparral fires that burned in 1896, 1898, and 1900 in the San Gabriel Mountains were studied from written accounts. Fires burned for 2 to 3 months during summer droughts. However, fire size was small (5,800 to 15,800 acres (2,300-6,390 ha)), suggesting that fire spread was slow, and fireline intensity was low. The smolder and run behavior that created a mosaic burn pattern and a community of fragmented fuels was created through the flare up of smoldering logs, trees, or other large fuels and the movement of embers into areas beyond the fire zone [185]. With the exclusion of fires and greater fuel uniformity, fires are now large and severe [185]. Since 20th century fire exclusion, fires are brief (< a few days) and extensive (>25,000 acres (10,000 ha)) [186]. Minnich and Bahre [187] found further support for these fire regime theories in a study comparing chaparral vegetation on the California-Baja California Norte.
Keeley, a supporter of the "stochastic" theory, suggests evidence that young southern California chaparral stands effectively suppress fire spread is lacking. Weather, according to stochastic theory supporters, is the primary controller of fire spread [142]. Santa Ana winds of southern California can exceed 60 miles (100 km)/hour and may have humidity levels below measurable levels. These winds create extreme fire conditions [97,142] that are not mitigated by stand age and/or species composition. Early 1900 fire exclusion practices have not decreased the fire frequency in southern California shrublands. Fire frequency has increased with increased human population. The current fire return interval for southern California shrublands is 30 to 40 years [142]. A study in San Diego County mapped 20th century fire data on top of corresponding vegetation types. The resulting picture supported Keeley's thoughts that fire size has decreased while amount of area burned has increased over time; it did not support Minnich's theory that the number of fires per decade has decreased while fire intensity and size have increased [316].
Regardless of what past FIRE REGIMES in chaparral may have been, human influences on FIRE REGIMES and disturbance patterns in heavily populated areas are now a dominant factor shaping southern California shrublands. In Pinnacles National Monument, the fire record from 1934 to 1978 indicates that the majority of fires were human caused, and in southern California in general, 75% of fires were human caused [2]. Keeley and Keeley [145] report that approximately 5% of California chaparral ignitions are natural. The introduction of persistent annual Mediterranean grasses has increased the probability of frequent fires [335]. For more on how humans, as ignition sources and fire suppressors, fit into the 2 prevailing theories on chaparral FIRE REGIMES, see [148].
Arizona chaparral: Researchers evaluated the fire history of a shrub live oak-alderleaf mountain-mahogany chaparral community from 7 fire-scarred stumps from a surrounding 215 (87 ha)-acre ponderosa pine forest in central Arizona's Bradshaw Mountains. There were no living trees with fire scars, and all living trees were less than 100 years old. Stumps recorded 92 fires in the 174-year period from 1700 to 1874, suggesting that ~1 fire burned every ~2 years. However, fire spread is poor in this chaparral type when stands are less than 10 years old and grass cover is scant. Flammability increases each year without fire. Researchers suggest that, although the surrounding ponderosa pine forest burned on average every 2 years, chaparral communities likely did not burn at an interval of less than 15 to 20 years. Fire protection and grazing likely both played a part in the dramatically decreased fire frequency since the late 1800s in the area [58].
Oak woodlands: The fire history of oak woodlands in the Yosemite Wilderness was determined for the 1930 to 1983 time period. Woodlands with an overstory of canyon live oak, interior live oak, and gray pine and alderleaf mountain-mahogany common in the understory contributed 4.2% to the wilderness' vegetation. Just 1.9% of Yosemite's fires occurred in this vegetation type. The average size of fires greater than 10 acres (4 ha) was 177.5 acres (71.8 ha). While weather conditions could likely support a fire in any year, lightning strikes were infrequent. The average number of fires in this vegetation type for the 53-year period was 22 fires per year per million acres. Fires were considered "intense" [297].
Parsons [215] summarized the recent fire history of foothill chaparral and oak woodland communities found below 6,000 feet (1,830 m) in Sequoia National Park. From 1920 to 1929, a total of 10,350 acres (4,189 ha) of the approximately 69,000 (28,000 ha)-acre study area burned. From 1930 to 1978, a total of 105 lightning fires burned 1,460 acres (592 ha), and 107 man-caused fires burned 6,946 acres (2,811 ha). The majority of fires (91%) were less than 7 acres (3 ha). Oak savannah and chaparral had the most burned area from 1920 to 1978 [215].
Ponderosa pine forests: The fire history of Kings Canyon National Park in California was reconstructed from 10 fire-scarred trees over a 400 (160 ha)-acre study area. Alderleaf mountain-mahogany was an occasional species in the ponderosa pine- and Jeffrey pine (Pinus jeffreyi)-dominated park. From 1775 to 1909, a fire burned somewhere in the study area on average once every 3.5 years, and the average fire interval per individual tree was approximately 11 years. There were no fire scars recorded on any tree after 1909 [310]. In Yosemite National Park, the number of lightning strikes from 1985 to 1990 in low- and mid-elevation sites, where chaparral and ponderosa pine are typical, was significantly (p<0.05) lower than what would be expected for low- and mid-elevation sites based on chance [296].
Mixed dry conifer forests: An extensive study of El Malpais National Park in New Mexico revealed that frequent low-severity surface fires were common in the presettlement period of ~1700 to 1880. Alderleaf mountain-mahogany was a common shrub in the mixed Douglas-fir, ponderosa pine, and pinyon-juniper forests. Nine sites were studied, and on average, sites burned every 5 to 12 years. The minimum and maximum fire return intervals were 1 and 55 years. From before 1700 to 1782, fires were frequent, patchy, and occurred predominantly in the late season during drought years. From 1783 to 1794, there were almost no fires, which was likely due to increased rainfall during this period. From 1795 to 1880, fire intervals were longer than in the pre-1700 to 1782 time period. Fire size was larger, and early season fires were most typical. Fires were uncommon or absent again from 1881 to 1892, and while ignitions occurred from 1893 to 1939 fire spread was limited. Intense domestic sheep grazing, which reduced fine fuel availability, was likely partially responsible for fewer fires from the late 1800s to 1939. From 1940 to 1992, the fire-free intervals were the longest in the past 500 years. Grazing, fire exclusion, logging, and/or road building affected the likelihood of fire after 1940. Large fires were reported in 1976 and 1989. The researcher indicated that the lack of fire in this area has created fuel and vegetation conditions that will likely support uncharacteristic high-severity, stand-replacing fires [91].
Dry Douglas-fir/alderleaf mountain-mahogany habitats in Utah likely did not burn frequently due to the discontinuity of fine fuels. However, when fires did burn they were likely severe [33]. Before fire exclusion practices, areas in the San Bernardino Mountains dominated by bigcone Douglas-fir were characterized by low-severity, summer-long fires. The interval between stand-replacing fires was likely long. Present stand replacement due to fires is ~26% and seemingly "excessive" when compared with the historic spatial extent of stand establishment [186].
Pinyon-juniper/petran chaparral: Using stand age structure sampling, researchers reconstructed the fire history for Colorado pinyon-Utah juniper-Rocky Mountain juniper, and petran chaparral for the 1850 to 2000 time period in Colorado's Mesa Verde National Park. Gambel oak, Utah serviceberry, and alderleaf mountain-mahogany are typical in petran chaparral. The sampling method allowed detection of only large severe fires. The fire cycle for the petran chaparral was an estimated 100 years. The fire cycle for the pinyon-juniper vegetation was ~400 years. Fire cycles for the early 1900s were long (~200 years for chaparral and ~1200 years for pinyon-juniper), likely due to livestock grazing, successful exclusion efforts, and/or a moist climate that prevailed in the southwest during this time. The fire regime since ~1950 more approximates that of the late 1800s. Changes in the fire regime may have been due to the removal of grazing and/or failed suppression efforts during severe weather conditions. From 1926 to 1977, the park averaged 8 fires per year; 90% of the fires were lightning ignited. Only 2% of the fires were greater than 10 acres (4 ha) but account for greater than 95% of the area burned in the 1900s. Large stand-replacing fires occurred during periods of prolonged drought and high wind conditions. Low-severity surface fires have been of limited extent in Mesa Verde Park [72].
FIRE REGIMES in singleleaf pinyon, California juniper, western juniper forests of the San Bernardino Mountains have not changed considerably with the exclusion of fire. Fire return intervals were long before and after exclusion. Long fire return intervals are attributed to low productivity and fuel build up rates [186]. Researchers estimate the fire cycle for these pinyon-juniper woodlands at 480 years. Fires were stand replacing and typically small. Postfire succession proceeds slowly in these woodlands [309].
Mountain shrublands: Bare ground and sparse grass and forb growth in most alderleaf mountain-mahogany stands in Wyoming's Laramie Basin [34] suggest that fires are uncommon.
The following table provides fire return intervals for plant communities and ecosystems where true mountain-mahogany is important. Find fire regime information for the plant communities in which this species may occur by entering the species name in the FEIS home page under "Find FIRE REGIMES".
Community or Ecosystem Dominant Species Fire Return Interval Range (years) California chaparral Adenostoma and/or Arctostaphylos spp. <35 to <100 sagebrush steppe Artemisia tridentata/Pseudoroegneria spicata 20-70 [217] Wyoming big sagebrush Artemisia tridentata var. wyomingensis 10-70 (x=40) [300,328] coastal sagebrush Artemisia californica <35 to <100 [217] plains grasslands Bouteloua spp. <35 [217,324] California montane chaparral Ceanothus and/or Arctostaphylos spp. 50-100 [217] curlleaf mountain-mahogany* Cercocarpus ledifolius 13-1,000 [13,248] mountain-mahogany-Gambel oak scrub Cercocarpus ledifolius-Quercus gambelii <35 to <100 Ashe juniper Juniperus ashei <35 western juniper Juniperus occidentalis 20-70 Rocky Mountain juniper Juniperus scopulorum <35 pinyon-juniper Pinus-Juniperus spp. <35 [217] Rocky Mountain bristlecone pine P. aristata 9-55 [62,63] Mexican pinyon Pinus cembroides 20-70 [192,276] Rocky Mountain lodgepole pine* Pinus contorta var. latifolia 25-340 [19,20,279] Sierra lodgepole pine* Pinus contorta var. murrayana 35-200 [10] Colorado pinyon Pinus edulis 10-400+ [72,85,136,217] interior ponderosa pine* Pinus ponderosa var. scopulorum 2-30 [10,15,168] Arizona pine Pinus ponderosa var. arizonica 2-15 [15,45,249] quaking aspen (west of the Great Plains) Populus tremuloides 7-120 [10,93,183] mountain grasslands Pseudoroegneria spicata 3-40 (x=10) [9,10] Rocky Mountain Douglas-fir* Pseudotsuga menziesii var. glauca 25-100 [10,11,12] coastal Douglas-fir* Pseudotsuga menziesii var. menziesii 40-240 [10,197,238] California oakwoods Quercus spp. <35 [10] oak-juniper woodland (Southwest) Quercus-Juniperus spp. <35 to <200 [217] canyon live oak Quercus chrysolepis <35 to 200 blue oak-foothills pine Quercus douglasii-P. sabiniana <35 Oregon white oak Quercus garryana <35 [10] California black oak Quercus kelloggii 5-30 [217] interior live oak Quercus wislizenii <35 [10] little bluestem-grama prairie Schizachyrium scoparium-Bouteloua spp. <35 [217] *fire return interval varies widely; trends in variation are noted in the species reviewFlammability: Using a combustion chamber and pelletized fuel samples from alderleaf mountain-mahogany tissue collected in Claremont, California's botanical garden, Montgomery [196] reported that alderleaf mountain-mahogany flammability was lower than most other chaparral species tested. Average ignition delay was 13.3 seconds, ignition temperature was 812.8 °F (433.8°C), and flame height was 2 inches (5.1 cm).
Fuels and fire behavior: A prescribed fire study in chaparral of San Diego County showed that 45% of live and 95% of dead true mountain-mahogany fuels were consumed when burning occurred in the spring. Sites were treated with a desiccant to encourage burning. At the time of the fire, twigs that were 0.125 to 1 inch (0.3-2.5 cm) in diameter had 68% to 82% moisture, and fuels that were 2.5 to 3 inches (6.3-7.6 cm) in diameter had 47% moisture. Live fuels averaged 29 tons/acre, and dead fuels averaged 19 tons/acre. Minimum relative humidity was ~30%, maximum temperatures were 77 to 80 °F (25-27 °C), and winds were variable up to 10 miles (16 km)/hour with gusts of 12 to 15 mi (19-24 km)/hour when the fire burned. A fire temperature of 1,250 °F (677 °C) was recorded at 31 inches (79 cm) above the soil surface but likely does not represent a maximum temperature [87].
After a fire, alderleaf mountain-mahogany fuel structure is a large number of fairly uniform diameter sprouts, and as time without fire or aboveground destruction increases, stem mortality occurs, and eventually just a few large diameter stems persist. Based on the analysis of 9 shrubs between 4 and 18 years of age, 66% of woody fuels were less than 0.4 inch (1 cm) in diameter, and dead fuels were uncommon [305].
Prescription fire: Prescription and wildfire behavior are similar in chaparral vegetation; both produce crown fires. However, the severity of chaparral fires can vary. Low-, moderate-, and high-severity fires produce surface soil temperatures of ~480 °F (250 °C), 800 °F (425 °C), and 1,300 °F (700 °C), respectively. Temperatures at 1 inch (2.5 cm) depths are 210 °F (100 °C), 350 °F (175 °C), and 390 °F (200 °C) for low-, moderate-, and high-severity fires [54].
Green [88] provides information regarding the planning and execution of prescription fires in chaparral vegetation. However, the use of prescribed fire in chaparral vegetation is not considered necessary or useful by some. Keeley and Fotheringham [142] indicate that "there are few places where fire-dependent shrubs are threatened by a lack of fire" in southern California, and that the use of prescription fire as a means to mitigate future fire hazards is unwarranted. Prescribed burning is not likely to prevent fire spread under severe weather conditions, and expense would be great while resource benefit would be small. Zedler [334], too, suggests the prevention of fire through fire use is unrealistic and adds there is no "urgent need to reduce the time between fires" in southern California chaparral. He recommends that fire suppression efforts concentrate only on recently burned southern California shrublands, that controlled burning be discontinued, and that wildfires be allowed to burn, unless there is evidence of harm to an endangered species. He indicates that fires should be allowed to burn even in very old shrublands even under the "most severe circumstances" [334].
Postfire management: Heavy browsing of alderleaf mountain-mahogany postfire sprouts can kill shrubs. This warning came from observations made in Lake County, California [26].
Evaluating fire severity: Keeley and others [147] evaluated fire severity by measuring alderleaf mountain-mahogany postfire shrub heights in chaparral vegetation in the Sierra Nevada.
Alderleaf mountain-mahogany occurs on dry slopes, hills, ridges, mesas, desert foothills, and rocky outcrops. It is associated with chaparral, northern oak woodlands, pine (Pinus spp.)-oak woodlands, mixed coniferous forests, alpine shrub, mountain brush, sagebrush (Artemisia spp.), aspen, and pinyon-juniper (Pinus-Juniperus spp.) vegetation [7,47,104,112,114,200,234,262,301,317,321].
In southeastern Arizona, Mouat [198] found that alderleaf mountain-mahogany was restricted to high-elevation sites with hilly or mountainous terrain and a high density of drainages (mostly >6 mi/mi²). Slopes occupied were steep (averaging >45%), and north aspects were preferred [198]. In the Wasatch mountains of north-central Utah, alderleaf mountain-mahogany stands occurred on slopes ranging from 25% to 60%. True mountain-mahogany stands on south-facing slopes were consistently found at higher elevations than stands on north-facing slopes, suggesting that a minimum moisture level requirement may exist in this area [36]. Alderleaf mountain-mahogany was significantly (p<0.01) associated with north aspects in the San Gabriel and San Bernardino mountains of southern California [99].
Climate: Alderleaf mountain-mahogany's large distribution suggests a wide tolerance of climatic conditions but limits an exhaustive review of climatic tolerance. Kitchen [151] indicates that the annual precipitation in alderleaf mountain-mahogany habitats ranges from 9.8 to 26 inches (250-660 mm). However, the Oregon white oak/alderleaf mountain-mahogany type in interior southwestern Oregon receives only 1.6 inches (40 mm) of precipitation per year [234]. Climate regimes for several alderleaf mountain-mahogany populations are provided.
California: San Mateo, coastal California, experiences a Mediterranean climate. The mean July and January temperatures are 68.2 °F (20.1 °C) and 48 °F (9.1 °C), respectively. Freezing occurs only occasionally. The average annual precipitation is 25.7 inches (654 mm), and a severe dry season of approximately 5 months (May to September) receives less than 0.59 inch (15 mm) of rainfall per month [1].
Colorado: Forty years of data collection in Mesa Verde, Colorado, revealed average annual precipitation of 18.3 inches (465 mm). Normal annual snow levels average 78 inches (2,000 mm), and May and June are dry months. In the late summer and early fall, brief intense storms are typical. The average monthly maximum temperature for a 40-year period was 102 °F (39 °C), and the minimum was -20 °F (-29 °C) [67].
New Mexico: In Carlsbad Caverns National Park in southern New Mexico, the climate is semiarid continental. Seventy-eight percent of annual precipitation occurs from May to October, and the average annual precipitation is 14 inches (360 mm). An extreme low of annual precipitation was 4.3 inches (110 mm) in 1951, and an extreme high of 43 inches (1,100 mm) was recorded in 1941. Droughts are common in southern New Mexico [155].
Utah: In alderleaf mountain-mahogany habitats of southeastern Utah, the climate is semiarid. Humidity levels are low, and precipitation is variable. The range in diurnal and annual temperatures is great; the average January and July temperatures are 27 °F (-3 °C) and 72.3 °F (22.4 °C), respectively. Annual precipitation averages 12.8 inches (324 mm) [206].
Wyoming: In Wyoming's Laramie Basin, the frequency of wind speeds greater than 2 miles (3 km)/hour in true mountain-mahogany stands are 77% to 87%, and wind speeds average 2.7 miles (4.3 km)/hour and 5.5 miles (8.9 km)/hour for 2 sites [34].
Elevation: Alderleaf mountain-mahogany typically occupies habitats between 4,000 and 10,000 feet (1,200-3,000 m) throughout its range [53]. Narrower elevational ranges by state and variety are provided below:
State/region Variety, if applicable Elevation in feet Arizona 4,500-7,000 C. m. var. glaber 3,000-6,500 C. m. var. paucidentatus 5,000-8,000 [134] California under 8,200 [112,200] C. m. var. var. blancheae below 2,000 C. m. var. glaber* under 8,200 C. m. var. macrourus 2,600-5,900 [112] Colorado 4,000-8,500 [104] Nevada 6,500-8,000 [133] New Mexico 4,500-9,500 [78] C. m. var. argenteus 6,000-8,000 C. m. var. montanus 6,000-8,000 C. m. var. paucidentatus 6,000-8,500 [178] Texas (Trans-Pecos region) C. m. var. argenteus 7,500-8,500 C. m. var. glaber 3,400-7,300 C. m. var. paucidentatus 4,600-8,000 [223] Utah 4,600-9,200 [317] Utah (Unita Basin) 6,000-8,500 [83] Intermountain West 3,900-9,800 [47] Southwest 3,500-9,000 C. m. var. argenteus (NM and TX) 4,000-8,500 C. m. var. paucidentatus (AZ, NM, TX, and Mexico) 4,500-8,500 [301] *C. m. var. glaber frequency increases with increased elevation in the San Gabriel and San Bernardino mountains [99].
Soils: Alderleaf mountain-mahogany occupies sites with well-drained, coarse, poorly developed, very shallow to moderately deep soils with almost neutral pH [34,151,260,271]. In the north-central Plains, alderleaf mountain-mahogany occurs on eroded hillsides with "badland" soils [262]. Habitats in north-central Utah have rocky, shallow soils with poor development [318]. In southeastern Arizona, alderleaf mountain-mahogany occurs predominantly on nonalluvial soils where limestone is the primary parent material. Soils are often very shallow and rocky [198,311]. Alderleaf mountain-mahogany was also absent from alluvial soils in the Laramie Basin of Wyoming. Alderleaf mountain-mahogany is rare or absent on serpentine soils in California's Santa Ana Mountains [302]. In Idaho alderleaf mountain-mahogany is found on poor shallow soils on rocky ridgelines but also on moister and deeper soils in the canyon bottoms [260]. Based on studies in the Intermountain West, Brotherson [35] reports that alderleaf mountain-mahogany habitats have an average soil depth of 9.7 inches (24.5 cm), pH of 7.3, soluble salt levels of 426 ppm, and clay, silt, and sand percentages of 33.5, 37.7, and 28.7, respectively.
In the Wasatch Mountain range in north-central Utah, stands where alderleaf mountain-mahogany was a dominant or codominant occurred on sites with soil depths of 3 to 19 inches (8-48 cm). Soils were classified as clay to sandy clay loams, and pH was neutral to slightly alkaline. Density of alderleaf mountain-mahogany was positively correlated with the percent clay and silt in the soil and negatively correlated with the percent sand [36]. In another study involving 20 north-central Utah sites, percent clay was positively correlated with true mountain-mahogany height, density, frequency, and cover. This finding together with observations of true mountain-mahogany at higher elevation sites on south slopes than on north slopes suggested that water retaining properties were important in dictating alderleaf mountain-mahogany's distribution [8]. In Utah's Uinta Basin, soil depth beneath alderleaf mountain-mahogany plants averaged 7.5 inches (19 cm), and pH averaged 7.7. Soils were 75% sand, 11% clay, and 14% silt [90].
Alderleaf mountain-mahogany provides food and habitat for numerous wildlife species and most classes of livestock. It is an important forage for mule and white-tailed deer, and the amount of literature on deer utilization of alderleaf mountain-mahogany is extensive. Utilization of alderleaf mountain-mahogany foods and habitats however are not exclusive to deer although this relationship has indeed received the most attention. For a more complete list of vertebrate wildlife species that use ponderosa pine and chaparral habitats, where alderleaf mountain-mahogany is common, in central Arizona's Sierra Ancha Experimental Forest, see [232].
Dittberner and Olson [61] rate alderleaf mountain-mahogany forage as good for elk, fair to good for mule and white-tailed deer, poor for pronghorn, and fair for upland game birds, small nongame birds, and small mammals.
Livestock: Alderleaf mountain-mahogany is rated good to excellent forage for all classes of livestock [53,78]. Forage is considered somewhat better for domestic sheep than cows [53].
Domestic goats in southern California consumed nearly 100% of the available browse of 1- and 5-year-old alderleaf mountain-mahogany shrubs. Researchers suggested that goats could be used to maintain fuel breaks to aid in the control of wildfires [89]. When domestic goats are used to control juniper (Pinchot juniper and Ashe juniper (Juniperus ashei)) in Texas, heavy grazing of the area is required, and if true mountain-mahogany is present on browsed sites it will likely decrease since it is more palatable than juniper [203].
Response of alderleaf mountain-mahogany to browsing is provided in the Response to browsing section below.
Livestock and wildlife: Utilization of alderleaf mountain-mahogany by both wildlife and livestock requires that habitats be managed to avoid over use. Alderleaf mountain-mahogany may be consumed by both livestock and wildlife species any time these species occupy the same area. Mule deer, cattle, and domestic sheep utilized 70% of leaves and annual stem growth of alderleaf mountain-mahogany on "properly" stocked areas on northern Arizona's Kaibab National Forest [130]. Alderleaf mountain-mahogany is referred to as a "superior feed" in California [245]. Alderleaf mountain-mahogany/sideoats grama habitat type in the Black Hills National Forest provides late spring and early summer livestock range and big game winter range. Researchers indicate that proper grazing management is essential to maintain deer winter range [116]. Colorado pinyon-alligator juniper/alderleaf mountain-mahogany-gray oak/mountain muhly vegetation in western New Mexico has high early-season grazing potential and receives moderate big game use [113]. On south-central Utah's Oak Creek range, alderleaf mountain-mahogany utilization was high in areas where either deer or cattle sign were plentiful. Use of alderleaf mountain-mahogany areas available only to deer was 34.4%, but on areas used by both deer and cattle, use was 53.6%. Areas with both deer and cattle averaged 1.7 times as many deer as did areas used by deer alone [131].
When alderleaf mountain-mahogany grows into a small tree form, much of its available browse is out of the reach of livestock and big game animals [123]. Following fire, heavy browsing of new true mountain-mahogany sprouts can kill shrubs [26].
Deer: Mule and white-tailed deer consume alderleaf mountain-mahogany and utilize alderleaf mountain-mahogany habitats year round throughout its range [39,78,209,210]. Degree of use may depend on phenology, season, and/or weather.
Alderleaf mountain-mahogany is typically rated as highly valuable or excellent forage [39,78,209,210]. Mule deer diet information that was summarized from studies in the western United States indicated that alderleaf mountain-mahogany received moderate use in the spring and fall and heavy use in the winter and summer based on 8 and 15 studies, respectively [163].
Contents of 96 mule deer stomachs taken from the Tehama winter range in northern California revealed that consumption of alderleaf mountain-mahogany was greatest in April and May. Usage was lowest in the fall and winter. The frequency of alderleaf mountain-mahogany in mule deer stomachs was 28.5% for this time [169]. Based on 328 fecal samples collected in southeastern Wyoming's Medicine Bow National Forest, the frequency of alderleaf mountain-mahogany in mule deer diets was 13% in the summer, 8% in the fall, 50% in the winter, and 46% in the spring. Winter and spring samples were collected in an area dominated by alderleaf mountain-mahogany [84]. In central Arizona's Sierra Ancha Experimental Forest, utilization by mule and white-tailed deer was greatest in the spring, 36%, and low to none until November when utilization was 13% [231].
Monitoring data from 22 radio-collared adult female mule deer indicated that alderleaf mountain-mahogany stands were "preferred" for feeding and for resting in the daylight hours. Monitoring was done from mid-November through March for 3 years in Colorado's Front Range [161]. Based on the volume of alderleaf mountain-mahogany in mule and white-tailed deer stomachs and its availability in the Three Bar Wildlife Area of south-central Arizona, researchers determined that alderleaf mountain-mahogany was not necessarily a preferred food source. Utilization did, however, increase in late winter [181]. In Guadalupe National Park in Texas, researchers determined that the relative density of alderleaf mountain-mahogany was 13% and frequency was 74% in annual mule deer diets. Fecal and rumen analysis and observations were used to assess consumption [158].
In the eastern part of Carlsbad Caverns National Park, mule deer fed on alderleaf mountain-mahogany for 7 of the 222.5 minutes of observations in a "good growing season" and in 6 of the total 158 observations. Feeding on alderleaf mountain-mahogany was not observed in a "poor growing season." In the Park's Scoggins Corner area, the frequency of alderleaf mountain-mahogany in 6 deer analyzed in a "good growing season" was 33% and in a poor growing season from 12 deer was 17% [155]. Alderleaf mountain-mahogany made up only 0.01% of the vegetative cover on New Mexico's Fort Stanton Experimental Ranch, but percent composition of mule deer diets in the area as determined through fecal analyses were 6.9%, 5.0%, and 5.8% in June, September, and March, respectively [174].
Several studies that involved the use of tame deer were not included in the above discussion. If interested, see [21,48,201].
Management issues related to deer use: Heavy winter browsing of alderleaf mountain-mahogany in California has caused severe shrub damage [245]. Chaparral vegetation on the San Joaquin deer winter range in Madera County, California, that was too tall or dense to provide deer browse was smashed by a bull dozer and then burned in early spring. After 6 years of intense deer browsing (utilization of 35% to 85%) of postfire sprouts, only a few grew out of deer's reach. On 22-year-old burned and browsed sites, alderleaf mountain-mahogany grew into a tree-like form largely beyond the reach of deer. Even with prolonged intense use, sprouts can become unavailable after time [80,81]. For more information on alderleaf mountain-mahogany and its response to browsing pressure, see Response to browsing.
Deer and sheep: In Palo Duri Canyon, Texas, researchers found a high degree of overlap in the diets of Barbary sheep and mule deer. The relative density of alderleaf mountain-mahogany was 12.5% and 11.9% in Barbary sheep and mule deer diets, respectively, based of the analysis of feces collected from February 1977 to January 1979. Sand shinnery oak (Quercus havardii) made up bulk of both species diets and contributed most to diet overlap [159].
Bighorn sheep: Alderleaf mountain-mahogany is consumed primarily in the summer by bighorn sheep. Alderleaf mountain-mahogany made up the greatest content in 3 bighorn sheep stomachs taken from Texas in the early 1900s (Bailey in [52]). No alderleaf mountain-mahogany was recovered from bighorn sheep feces collected in the fall or winter in the Sheep Creek-Trickle Mountain Area of Saguache County, Colorado. Bighorn sheep did feed on alderleaf mountain-mahogany in the summer and spring. Alderleaf mountain-mahogany made up a trace of spring diets and 12.1% of the relative density of feces collected in the summer [285].
Bighorn sheep in Waterton Canyon, Colorado, preferred mountain shrub habitats between east and southwest aspects. Alderleaf mountain-mahogany was the dominant shrub on south slopes [239]. Based on analysis of collected feces and direct feeding observations, researchers found that alderleaf mountain-mahogany leaves dominated the summer diet of bighorn sheep. From fecal analyses, alderleaf mountain-mahogany made up 74% to 90% of the relative densities of diets from June through September. Observations generally agreed with fecal analyses findings [240].
Deer and elk: Alderleaf mountain-mahogany is thought to be consumed more heavily by deer than elk [295], but both game animals utilize alderleaf mountain-mahogany habitats and browse. However, season of consumption may differ. Colorado pinyon/alderleaf mountain-mahogany/nodding brome vegetation in western New Mexico is considered excellent deer habitat but receives elk use, too [113]. Usage of Colorado pinyon-alligator juniper vegetation where alderleaf mountain-mahogany was important, as determined by deer and elk pellet counts, increased as shrub coverage increased in the Gila National Forest of southern New Mexico [230].
Researchers compared the diets of mule deer and elk in Colorado pinyon-oneseed juniper and ponderosa pine woodlands in north-central New Mexico. Based on year-round feces collections that spanned 2 years, alderleaf mountain-mahogany was 5% and 16% of annual elk and mule deer diets, respectively. Utilization of alderleaf mountain-mahogany was greatest in the summer for elk when it made up 12% the diets, and mule deer diets were 23% to 24% alderleaf mountain-mahogany in the summer and fall [246]. Both elk and mule deer fed heavily on alderleaf mountain-mahogany in the Fort Baynard Watershed in New Mexico, but season of heaviest use differed. Alderleaf mountain-mahogany made up 17% to 39% of 43 elk rumens. The low of 17% was found in the winter and the high of 39% was in the spring. For mule deer, stomachs contained 15% to 45% true mountain-mahogany. The highest amount of alderleaf mountain-mahogany was from collections made in the fall and winter, and the lowest was from summer collections [253].
Neither deer nor elk utilized alderleaf mountain-mahogany from January through March on northern Utah winter ranges. Based on fecal analyses, alderleaf mountain-mahogany was absent from mule deer diets and was less than 1% of the relative density of elk diets [199].
Elk: Based on feeding observations in Wind Cave National Park in the northern Great Plains, elk use of true mountain-mahogany was greatest in the spring. Fall, winter, spring, and summer use was 0%, 3.4%, 6.8%, and 1.1%, respectively [325].
Pronghorn: Researchers have observed pronghorn feeding on alderleaf mountain-mahogany and detected alderleaf mountain-mahogany in pronghorn feces. In north-central Colorado, researchers observed pronghorn feeding on alderleaf mountain-mahogany, but percentage of the diet or preference were not discussed [224]. In Utah the maximum percentage of true mountain-mahogany in pronghorn diets was 15% based on rumen or feces collected in the spring or summer [254].
Coyotes: In Las Animas County, Colorado, researchers found that 72 radio-collared coyotes used canyon shrub-grassland communities dominated by skunkbush sumac and alderleaf mountain-mahogany more than expected (p<0.005) based on habitat availability. Of 18 occupied dens, 7 were in the shrub-grassland type [79].
Collared peccaries: Researchers found that alderleaf mountain-mahogany was one of the most common shrubs found at javelina bed sites in urban areas of Prescott, Arizona. Bed sites were located by tracking 8 collared peccaries from 6 different herds [284].
Small mammals: Many small mammals feed on alderleaf mountain-mahogany and utilize its habitats. In the Platte River Watershed in Colorado, prairie voles and deer mice were captured most often in the true mountain-mahogany-dominated vegetation type [211]. Brush mice utilized alderleaf mountain-mahogany-dominated vegetation in the Granite Basin area of Arizona's Yavapai County. Nearly 30% of all trappings occurred in alderleaf mountain-mahogany-dominated stands. Alderleaf mountain-mahogany stands averaged 4.3 captures/trap night [65].
In California's San Gabriel Mountains, steep rocky slopes dominated by alderleaf mountain-mahogany and Nuttall's scrub oak were habitat for brush mouse, California mouse, and dusky-footed woodrat populations. In a single acre of Nuttall's scrub oak-alderleaf mountain-mahogany shrublands near Jackson Lake, there were 13 occupied dusky-footed woodrat nests [298]. Researchers examined the food contents in dusky-footed woodrat nests on the San Dimas Experimental Forest in Los Angeles County, California. The frequency of alderleaf mountain-mahogany leaves in nests was as high as 100% on some sites, and the maximum distance leaves were carried was 45 feet (14 m). Alderleaf mountain-mahogany is an important summer food. Over 0.02 pound (7 g) of alderleaf mountain-mahogany was recovered from nests in July and September, but alderleaf mountain-mahogany occurred in nests in all seasons [120].
Alderleaf mountain-mahogany is typical in oak chaparral and oak-juniper habitats in Brewster County, Texas, utilized by white-ankled mice, Merriam's pocket mice, brush mice, cactus mice, yellow-nosed cotton rats, western white-throated woodrats, desert cottontails, black-tailed jackrabbits, rock squirrels, Texas antelope squirrels, raccoons, ringtails, and gray foxes. Specific use of alderleaf mountain-mahogany was not discussed [55].
Based on fecal analyses made from summer collections from Colorado pinyon-Utah juniper woodlands of Colorado's Piceance Basin, researchers found that alderleaf mountain-mahogany made up 1.1% and 2.2% of the relative density of deer mouse and least chipmunk habitats, respectively [106].
Birds: Alderleaf mountain-mahogany provides cover and habitat to many game and song birds. Scaled quail in Oldham County, Texas, used alderleaf mountain-mahogany habitats in the summer [269]. One of seven located Montezuma quail nests in the Huachuca Mountains of Cochise County, Arizona, occurred on hot, dry limestone ridges where cover included alderleaf mountain-mahogany [307]. Seventy night time scaled quail roost sites were found in the Rolling Plains of northwestern Texas in the spring and winter. The alderleaf mountain-mahogany-yucca (Yucca spp.)-fragrant sumac vegetation type was selected by scaled quail based on calculated electivity ratios. Eighteen of the located roosts were in this vegetation type [270].
Between 19 and 32 species of breeding birds occurred in upland forests of east-central Taos County, New Mexico. Alderleaf mountain-mahogany was a common shrub in these forests. However, specific use of alderleaf mountain-mahogany was not mentioned [259]. There were 9 species of breeding birds and a total of 103 breeding pairs/40 ha in shrub live oak-alderleaf mountain-mahogany chaparral vegetation in central Arizona's Tonto National Forest. The following bird species occurred in a greater numbers in shrub live oak-alderleaf mountain-mahogany chaparral vegetation than in nearby grassland or riparian sites: Gambel's quail, common poorwill, bridled titmouse, Bewick's wren, crissal thrasher, northern cardinal, and eastern towhee [277].
Alderleaf mountain-mahogany was common in Virginia's warbler breeding habitats in the southwestern Black Hills of South Dakota. Researchers often observed Virginia's warblers foraging in alderleaf mountain-mahogany shrubs. It is likely that alderleaf mountain-mahogany was important nest cover, too. The density of breeding Virginia's warblers was 8.3 birds/km² [275]. In the Sandia Mountains of north-central New Mexico, a crissal thrasher nest was located 2 feet (0.6 m) above the ground in a alderleaf mountain-mahogany shrub in Embudo Canyon [280].
Palatability/nutritional value: Alderleaf mountain-mahogany is considered a "highly preferred," "highly palatable" browse species [123,202]. Stems are considered palatable year round [290]. Palatability rates excellent to very good for deer, bighorn sheep, and domestic goats, good to very good for cattle and goats, and good to poor for horses [128,245,271]. Alderleaf mountain-mahogany is considered an excellent source of nitrogen for livestock and big game animals [14]. Forage collected in the winter near Logan, Utah, was fed to captive deer. Based on browsing time and forage consumed, alderleaf mountain-mahogany à curlleaf mountain-mahogany hybrids were considered more palatable than alderleaf mountain-mahogany species [255].
Alderleaf mountain-mahogany in vitro digestibility, crude protein, and phosphorus average 26.5%, 7.8%, and 0.13%, respectively, in the winter season. These averages are based on values reported in the literature. Given these values, a diet of only alderleaf mountain-mahogany is deficient in meeting crude protein, total digestible nutrients, and phosphorus requirements of wintering mule deer but exceeds calcium and carotene requirements [314]. Based on the nutrient requirements for cattle, domestic sheep, horses, deer, elk, small mammals, and birds, a diet of only alderleaf mountain-mahogany in the winter would be deficient for all but cattle during gestation, lactation, growth, and maintenance periods. For cattle, a diet of only alderleaf mountain-mahogany would be adequate only during maintenance and gestation periods [315]. In the Uinta Mountains of Utah, researchers reported nearly toxic levels of copper in alderleaf mountain-mahogany. Poisoning would depend on the amount of alderleaf mountain-mahogany consumed [38].
The current year's growth of mountain-mahogany (Cercocarpus spp.) leaves and stems were higher in digestible organic matter and crude protein only in the 1st postfire growing season when browse on control, 1-, 2-, 3-, and 6-year-old burned sites was compared in Guadalupe Mountains of New Mexico [156].
Many studies have investigated the nutritional content of alderleaf mountain-mahogany leaves and stems. Some studies analyzed stems and leaves separately while other did not discriminate. Some were interested in seasonal fluctuations while others were season specific. Some report that crude protein levels do not fluctuate seasonally [22,294], while others indicate that crude protein decreases from spring to winter [231]. Refer to the studies listed below for information on alderleaf mountain-mahogany nutritional and/or chemical content.
Study area Plant portion collected Season of collection Notes Reference Arizona current year's growth; stems and leaves analyzed separately late September, late November, early January, early March provides in vitro digestibility for mule and white-tailed deer separately [294] Arizona, Mogollon Plateau leaves June, August [293] Arizona, Sierra Ancha Experimental Forest terminal 2 inches (5 cm) of sprout growth year round [231] California current year's growth stems and leaves year round crude protein only [25] California, North Coast Range current year's growth year round comparisons for 2-, 3-, and 5-year-old burned sites [278] Colorado, western current year's growth January [162] New Mexico, north-central in Jemez Mountains current year's growth January-April 3-year-old burned site [244] New Mexico, southwest in Fort Baynard Watershed current year's growth year round [253] Wyoming current year's growth growing season [98] Utah, central leaves and stems analyzed separately April, July, September, November collections in April were before leaf out; in July relatively little growth; in November temperatures freezing [242] Utah, Uinta Basin current year's growth; stem base, middle, and tip analyzed separately early October [35] Utah, Uinta Mountains stems within easy reach of large herbivores early October [38]Cover value: Alderleaf mountain-mahogany likely provides cover for many wildlife species. The oak chaparral vegetation of Brewster County, Texas, where alderleaf mountain-mahogany is common, provides good cover for black-tailed jackrabbits and desert cottontails [55]. Alderleaf mountain-mahogany provides excellent hiding or escape cover and fawning cover and good thermal cover for mule deer in Wyoming [209]. Alderleaf mountain-mahogany is considered fair hiding/escape and thermal cover and good fawning cover for white-tailed deer in Wyoming [210]. Dittberner and Olson [61] rate alderleaf mountain-mahogany cover as good for mule and white-tailed deer, fair for elk, poor for pronghorn, and fair to good for upland game birds, small nongame birds, and small mammals.
Alderleaf mountain-mahogany is a dominant species in the following vegetation classifications:
General/Southwest:
Alligator juniper-oneseed juniper (Juniperus deppeana-J. monosperma)/true
mountain-mahogany-desert ceanothus (Ceanothus greggii)
Colorado pinyon (Pinus edulis)/alderleaf mountain-mahogany type; Gambel oak (Quercus gambelii),
sandpaper oak (Q. pungens), Alligator juniper, and oneseed juniper phases recognized
Mexican pinyon (P. cembroides)-alderleaf mountain-mahogany-evergreen sumac (Rhus
virens var. choriophylla)
Colorado pinyon/alderleaf mountain-mahogany/spreading phlox (Phlox diffusa) in southeastern Great
Basin [318]
Arizona:
alderleaf mountain-mahogany type associated with Mexican pinyon in Chiricahua National
Monument [228]
alderleaf mountain-mahogany scrub in the Santa Catalina Mountains [319]
chaparral [229]
desert ceanothus-alderleaf mountain-mahogany-littleleaf
sumac (R. microphylla) on Fort Bowie National Historic Site [311]
Colorado pinyon/alderleaf mountain-mahogany in northern Arizona [166] and in southeastern
Arizona
gray oak (Q. grisea)/alderleaf mountain-mahogany in southeastern Arizona [272]
shrub live oak (Q. turbinella)-alderleaf mountain-mahogany [40]
Toumey oak (Q. toumeyi)/grassland in Chiricahua National Monument
[228]
California:
alluvial scrub vegetation in coastal southern California [101]
desert chaparral in San Bernardino Mountains [184]
Nuttall's scrub oak (Q. dumosa) chaparral [117] in San Bernardino Mountains
[119]
scrub oak (Quercus spp.)/alderleaf mountain-mahogany chaparral type in the Liebre
Mountains [32]
island chaparral in canyons and on slopes of interior southeastern Santa Catalina
[218]
mixed mesic form chaparral on northern aspects in southern California [97]
alderleaf mountain-mahogany/Eastern Mojave buckwheat (Eriogonum fasciculatum) chaparral
in the Liebre Mountains [32]
blue oak-gray pine/wedgeleaf ceanothus (Q. douglasii-Pinus sabiniana/
C. cuneatus)-alderleaf mountain-mahogany along the coast [4,5]
blue oak/alderleaf mountain-mahogany/hoary
bowlesia-San Francisco woodland-star (Bowlesia incana-Lithophragma affine) in
southern San Luis Obispo and northern Santa Barbara counties [31]
interior live oak-canyon live oak (Q. wislizenii-Q. chrysolepis) chaparral
woodland (Horton, in [100]
island oak (Quercus spp.) woodland
cismontane California juniper (J. californica) woodland and
scrub
southern interior cypress (Arizona cypress, Tecate cypress (Cupressus
arizonica, C. forbesii)) forest
bigcone Douglas-fir (Pseudotsuga macrocarpa)-canyon live oak forest [117]
Colorado:
Utah serviceberry-Wyoming big sagebrush (Amelanchier utahensis-Artemisia tridentata ssp.
wyomingensis)-alderleaf mountain-mahogany-antelope bitterbrush-mountain snowberry (Purshia
tridentata-Symphoricarpos oreophilus)
Utah serviceberry
-alderleaf mountain-mahogany [17]
alderleaf mountain-mahogany-skunkbush sumac (Rhus trilobata) [17,42]
alderleaf mountain-mahogany/bluebunch wheatgrass (Pseudoroegneria spicata) in northwestern Moffat County [18]
Gambel oak-alderleaf mountain-mahogany [17]
shrub live oak-alderleaf mountain-mahogany [247]
Rocky Mountain juniper (J. scopulorum)/alderleaf mountain-mahogany in Roosevelt
National Forest [110]
Utah juniper (J. osteosperma)/alderleaf mountain-mahogany-squaw apple (Peraphyllum ramosissimum)
in White River National Forest [111]
Utah juniper-Colorado pinyon/alderleaf mountain-mahogany/bluebunch
wheatgrass in northwestern Moffat County [18]
oneseed juniper/alderleaf mountain-mahogany-wax current (Ribes cereum)
Colorado pinyon/alderleaf mountain-mahogany [247]
Colorado pinyon-Utah juniper/alderleaf mountain-mahogany
in White River National Forest [111]
ponderosa pine (Pinus ponderosa)/alderleaf mountain-mahogany [247] in Roosevelt
National Forest [110]
ponderosa pine/alderleaf mountain-mahogany/big bluestem (Andropogon gerardii)
[247]
Idaho:
alderleaf mountain-mahogany-antelope bitterbrush in Wasatch Mountains [227]
Montana:
Rocky Mountain juniper-limber pine (P. flexilis) in south-central part of the
state [216]
Nebraska:
alderleaf mountain-mahogany/sideoats grama (Bouteloua curtipendula) [247]
Nevada:
Utah serviceberry-alderleaf mountain-mahogany shrublands;
rare type [204]
ponderosa pine/mountain shrub and mountain shrub types near Yucca Mountain [287]
New Mexico:
alderleaf mountain-mahogany-serviceberry-Mexican cliffrose-littleleaf mock orange
(Amelanchier spp.-Purshia mexicana-Philadelphus microphyllus)-sparse
shrub
alderleaf mountain-mahogany-littleleaf mock orange-fendlerbush (Fendlera rupicola)-skunkbush
sumac-sparse shrub
alderleaf mountain-mahogany-shrub live oak-skunkbush sumac-sacahuista-Wright
silktassel (Nolina microcarpa-Garrya wrightii)-sparse shrub
alderleaf mountain-mahogany-Pinchot juniper (J. pinchotii)-desert ceanothus-shrub live
oak-Gambel oak à shrub live oak/mixed grass
alderleaf mountain-mahogany-gray oak-shrub live oak-skunkbush sumac-sparse shrub
alderleaf mountain-mahogany-shrub live oak-turpentine bush (Ericameria laricifolia)-Wright
silktassel-Schott's yucca (Yucca schottii)-sparse shrub [57]
alderleaf mountain-mahogany/sideoats grama [247]
gray oak-true
mountain-mahogany in southern New Mexico [272]
gray oak-alderleaf mountain-mahogany-banana yucca (Y. baccata)/mixed forb [57]
shrub live oak-alderleaf mountain-mahogany
oneseed juniper/alderleaf mountain-mahogany [247]
Colorado pinyon-oneseed juniper/alderleaf mountain-mahogany/big bluestem [57] in Guadalupe,
Jicarrilla, and Sacramento mountains of Lincoln National Forest [149]
Mexican pinyon/Toumey oak-alderleaf mountain-mahogany-pointleaf
manzanita (Arctostaphylos pungens)/mixed grass [57]
Colorado pinyon/alderleaf mountain-mahogany/nodding brome (Bromus anomalus) high elevation
woodland in the Gila National Forest; Gambel oak phase recognized
Colorado juniper-alligator juniper/alderleaf mountain-mahogany-gray
oak/mountain muhly (Muhlenbergia montana) in western portion of state; Parry's agave (Agave
parryi) phase recognized [113]
Oklahoma:
oneseed juniper/alderleaf mountain-mahogany
Colorado pinyon/alderleaf mountain-mahogany [247]
Oregon:
Oregon white oak (Q. garryana)/alderleaf mountain-mahogany, a rare type in the interior valleys of
southwestern Oregon [234]
South Dakota:
western chaparral climax in the Black Hills [182]
alderleaf mountain-mahogany/sideoats grama [247] in Black Hills National Forest [116]
alderleaf mountain-mahogany-fragrant sumac (Rhus aromatica) in Black Hills National Forest
[261]
ponderosa pine-Rocky Mountain juniper/alderleaf mountain-mahogany-common snowberry/little bluestem
(S. albus/Schizachyrium scoparium)
in southwestern Black Hills [281]
Texas:
alderleaf mountain-mahogany-desert ceanothus-Gambel oak à shrub live oak-sandpaper oak
smooth-leaf sotol (Dasylirion leiophyllum)-alderleaf mountain-mahogany in Guadalupe
Mountains National Park (Warnock in [207])
sandpaper oak-alderleaf mountain-mahogany in Trans Pecos region [56]
Utah:
alderleaf mountain-mahogany-antelope bitterbrush in Wasatch Mountains [227]
Colorado pinyon/alderleaf mountain-mahogany/Indian ricegrass (Achnatherum hymenoides)
in the central Great Basin of southern Utah [318]
Colorado pinyon-Utah juniper/alderleaf mountain-mahogany in Manti-LaSal National Forest [283]
Douglas-fir (Pseudotsuga menziesii)/alderleaf mountain-mahogany in southern portions of Abajo
Mountains and Aquarius Plateau [329]
Wyoming:
alderleaf mountain-mahogany/Idaho fescue (Festuca idahoensis) [42]
alderleaf mountain-mahogany/sideoats grama [247] in Black Hills National Forest [116]
alderleaf mountain-mahogany-antelope bitterbrush in Wyoming Basin [42]
alderleaf mountain-mahogany-fragrant sumac in Black Hills National Forest [261]
alderleaf mountain-mahogany/soapweed yucca(Y. glauca) in Cheyenne River Basin [282]
Utah juniper/alderleaf mountain-mahogany in southwest
Rocky Mountain juniper-ponderosa pine/alderleaf mountain-mahogany in Black Hills [42]
ponderosa pine/alderleaf mountain-mahogany [247]
Mexico:
Baja California Norte:
mixed chaparral vegetation north of Ensenada [188]
Response to browsing:
While alderleaf mountain-mahogany tolerates "close" browsing [290], it can be killed by heavy use [295].
It has been referred to as both a decreaser and increaser in the literature. Parker [214] considers true
mountain-mahogany a "decreaser" when browsed yearlong by domestic sheep, cattle, and deer but
refers to alderleaf mountain-mahogany as an "increaser" on upland or mountain sites when only cattle
are the nonnative browsers. Zacek and others [330] indicate that alderleaf mountain-mahogany decreases in relative
abundance with continued heavy use. Likely site conditions, grazing timing, and climatic stress all play a role
in the browsing response.
Alderleaf mountain-mahogany cover was lower after 18 years of light and moderate use by cattle in Colorado's
Manitou Experimental Forest. Before cattle were on the sites, alderleaf mountain-mahogany cover was 0.12% and
0.05% on lightly and moderately used sites, respectively. After the introduction of cattle, coverage was
0.05% on lightly used and 0.02% on moderately used sites [256]. In Rio Blanco County, Colorado, 3 relatively
homogeneous enclosures were utilized at different levels by different herbivores. Enclosure 1 supported 2
domestic sheep/acre/month during the spring and fall and 2.3 deer/acre/month in the winter. Enclosure 2 was
utilized at 15.4 cows/acre/month in the spring and fall and 2.3 deer/acre/month in the winter. Enclosure 3
had 1.6 deer/acre/month in the winter. Percent composition of alderleaf mountain-mahogany was lowest in enclosure
1, and sheep browsing had decreased plant "vigor" and killed some plants. Composition or frequency
of alderleaf mountain-mahogany in enclosures 2 and 3 was not different, but cattle and deer browsing on true
mountain-mahogany was heavy. Heavy use of alderleaf mountain-mahogany by sheep may decrease browse available to
deer using the same area [237].
Researchers compared alderleaf mountain-mahogany growth inside and outside an exclosure in the Medicine Bow
National Forest of Wyoming. Vegetation within the exclosure was protected for 40 years. Alderleaf mountain-mahogany
density and canopy cover were similar inside and outside of the exclosure. Production was significantly (p=0.1)
higher outside (242.3 kg/ha) than inside (107.8 kg/ha), and the amount of dead alderleaf mountain-mahogany material
was significantly greater inside (43.5%) than outside (18.7%). Researchers suggested that some level of browsing
may be necessary to maintain an "optimum sustained yield" of alderleaf mountain-mahogany [312].
Browsed and unbrowsed alderleaf mountain-mahogany shrubs in the Wasatch and Uinta mountains of Utah were more
affected by resource availability than by previous year's use. Regardless of use, annual twig growth
and flower and seed production were greatest when precipitation levels were greatest. Utilization of
shrubs ranged from <21% to over 300% (consumption of annual growth and previous year's growth). Shrubs
compensated but did not over compensate for losses; browsed shrubs were smaller than unbrowsed
shrubs. Flower
and seed production were much greater on unbrowsed than browsed shrubs. Herbivores in the area were moose,
elk, deer, antelope, and cattle [288,289].
Browsing simulated by clipping:
Clipped shrubs produced more biomass than unclipped shrubs in Logan, Utah. Ninety percent of the previous
year's growth of 4-year-old shrubs was clipped in either December or April. When 90% of the distal buds were
removed in the spring, alderleaf mountain-mahogany produced significantly (p<0.05) more biomass than control
plants. Alderleaf mountain-mahogany also averaged 1 new sprout/plant following the treatment. Experiments were
conducted in manipulated environments where nutrients and light availability were high, unlike natural conditions
[308].
Season of clipping affected the response of alderleaf mountain-mahogany in Ft. Collins, Colorado. Researchers
removed 80% and 40% of stems at ~2 inches (5 cm) above the ground. Current annual growth of clipped plants
was higher than controls when plants were clipped in the dormant season and lower when plants were clipped
during the growing season. Differences, however, were not significant [327]. Twelve years of clipping true
mountain-mahogany in southwestern Colorado suggests that shrubs are resistant to browsing levels of 60% to
80%. No significant differences were found between the current annual growth or mean stem length of control
shrubs and shrubs with 20% to 100% of their current annual growth removed. Plants receiving 100% removal
treatments were monitored for 10 years. Seed production was lowest for heavily clipped shrubs. Shrubs that
had all current annual growth removed and old wood to 0.2 inch (4 mm) removed produced new sprouts early in
the treatments, but none survived over 6 years of this treatment [251].
Predictive equations:
Predictive equations for estimating alderleaf mountain-mahogany fuels in southern California chaparral are
provided in [304]. Brotherson and others [37] used diameter and age data to develop predictive equations
for populations in the Laramie Basin of Wyoming and the Uinta Mountains of Utah. Combined data from both
populations produced more accurate estimates for either population, and researchers suggested that similar
alderleaf mountain-mahogany growth rates made equations useful to both areas. Researchers used twig length and
diameter measurements of alderleaf mountain-mahogany current year's growth, including leaves, in a regression
equation to predict production and utilization. Measurements were obtained from shrubs in the Organ Mountains
Recreation Lands in southern New Mexico. Predictive power for other populations was not assessed [175].
Methods for increasing production:
Dimeo [60] provides a review of the mechanical tools used (successfully and unsuccessfully) to increase
alderleaf mountain-mahogany browse.
Methods to control:
The control of postfire alderleaf mountain-mahogany sprouts as a means to increase herbaceous species production
in chaparral vegetation in San Gabriel Mountains is described in [129].
Native people utilized alderleaf mountain-mahogany in the past. Wood was used by native people of southern California in making arrow tips, digging tools, spears, and fighting clubs [41,50]. Navajo people used alderleaf mountain-mahogany to make their weaving tools and other tool handles. Alderleaf mountain-mahogany wood was used in the construction of sweat houses and prayer sticks utilized in the Mountain Chant Ceremony [66,299]. Hopi people used alderleaf mountain-mahogany bark to make a red-brown dye for leather [271]. This type of dye was also utilized by the Ramah Navaho of northwestern New Mexico to color buckskin, wool, and basket materials [299]. Kawaisu natives of south-central California carved pipe heads from alderleaf mountain-mahogany [336].
Alderleaf mountain-mahogany was also used medicinally. Navajo people took alderleaf mountain-mahogany extracts to treat stomach problems, such as sickness from overeating venison, and to speed up postpartum recovery. Chewing alderleaf mountain-mahogany leaves from a deer-browsed plant was thought to bring a Navajo hunter good luck [66,299]. Native people of southeastern Utah reportedly utilized alderleaf mountain-mahogany as a medicine but what it treated is unknown [206]. Kawaisu natives of south-central California boiled the root of alderleaf mountain-mahogany and drank it to treat coughing [336].
More recently alderleaf mountain-mahogany has been suggested as a landscape or garden plant in the northwestern and southwestern U.S. and in the southern Great Plains [95,126,157]. It is suggested for dry, well-drained sites [157] and has been recommended for water-wise landscaping because of its drought and heat tolerance [95].
Alderleaf mountain-mahogany may also have a future use. Arizona chaparral species including true mountain-mahogany showed promise as "feedstocks for pyrolytic conversion to liquid fuels." For more information, see [51].
Wood Products: Wood produced by alderleaf mountain-mahogany is hard and brittle [107,290].
Generally, alderleaf mountain-mahogany flowers from April to early July and fruits ripen and disperse from July to October. Some varieties in the southern United States and Mexico flower as late as November. Site conditions including latitude, elevation, and aspect affect the timing of flower and fruit production [151,153,271]. Flowering and fruiting dates are presented by state or region and by variety in the tables below.
Flowering and fruit production dates for alderleaf mountain-mahogany varieties are summarized below [177]:
C. m. var. argenteus June-July C. m. var. blancheae March-June C. m. var. glaber February-June; rarely September-October C. m. var. macrourus June C. m. var. minutiflorus March-May C. m. var. montanus May-July C. m. var. paucidentatus May-NovemberThe following table summarizes the phenology of alderleaf mountain-mahogany based on 9 years of data collected at 2 elevations in Utah's Ephraim Canyon. Presented are the average dates of appearance [46]:
ElevationAlderleaf mountain-mahogany sprouts from the root crown and/or from rhizomes following fire [47,146]. Sprouts from rhizomes may appear ~3 feet (1 m) away from the main stem [146]. Postfire sprouting is often described as "vigorous" and "rapid" [33] and is common under a range of FIRE REGIMES [1]. Fire severity, degree of soil heating, soil and duff moisture, and the physiological stage of plant development will likely affect the postfire sprouting response [190].
Seed is typically killed by fire, and it is rare to find seedlings in early postfire communities [146]. Biswell and Gilman [27] indicate that late season fires are most detrimental to seedling production on burned sites. Hanes and Jones [102] report that seedlings may establish from off-site seed sources in early postfire communities, although Halsey [97] indicates that alderleaf mountain-mahogany seed germination is best in mature stands of chaparral.
Postfire sprouting: Fire timing, fire severity, site conditions, and ecotypic variation may affect alderleaf mountain-mahogany postfire sprouting. Ferguson [70] suggests that the alderleaf mountain-mahogany postfire sprouting response may be ecotype dependent. Hanes [99] indicates that alderleaf mountain-mahogany sprouts are typically taller on burned desert than on burned coastal sites.
Production of postfire sprouts, however, is not guaranteed. On 1- to 2-year-old burned sites in the San Gabriel Mountains density of sprouting alderleaf mountain-mahogany shrubs was 2/acre and nonsprouting shrub density was 3/acre. Information about the fire was lacking. Fire severity, seasonality, or behavior may have affected sprouting [150].
Sprouting was vigorous after a fire in the western foothills of the Sierra Nevada Mountains of Tehama County. The fire did not kill alderleaf mountain-mahogany shrubs, and postfire sprouts produced seed in the 5th postfire year. Researchers noted that fire timing affects alderleaf mountain-mahogany's sprouting response. If burned in a late season fire (late September or after), postfire sprouts may not appear until the following spring. If burned during the growing season, postfire sprouts typically appear by postfire week 3. Heavy browsing of postfire spouts frequently kills alderleaf mountain-mahogany [27].
Sixty-four percent of alderleaf mountain-mahogany shrubs sprouted ~4.5 months after an "extremely hot" July fire in the San Gabriel Mountains. A greater percentage of plants were sprouting on the low-elevation site (2,100 feet (640 m)) than at the high-elevation sites (3,100 (940 m) and 4,100 feet (1,200 m)). Some areas had been sprayed with herbicide prior to the fire; whether or not this affected postfire sprouting was not discussed [220].
Researchers found that alderleaf mountain-mahogany produced more and taller spouts in areas burned at low severity than on unburned or high-severity burned sites following the Hi Meadow Fire in Colorado. Basal and epicormic sprouts occurred on low-severity burned sites. Only basal sprouts occurred on high-severity burned sites and only epicormic sprouts on unburned sites [171]. Similar results were found following a January fire in Carlsbad Caverns National Park. More and longer sprouts were produced by "lightly" burned or scorched true mountain-mahogany shrubs than by shrubs that were "burned to char." Sprouts averaged 2 inches (5 cm) in postfire month 2 and 8 inches (20 cm) in postfire month 6. Shrubs regained their prefire height of 2.5 feet (0.76 m) three and a half years following the fire [155].
Seed survival and postfire seedling production: Heat treatments decrease alderleaf mountain-mahogany germination, but seeds have survived fire and seedlings have appeared in early postfire communities. However, the recruitment of new alderleaf mountain-mahogany shrubs may require a fire-free interval of 100 years or more [138], and seeds are thought to germinate best in mature chaparral stands [97]. For information on heat treatment effects on alderleaf mountain-mahogany seed see Heat treatments.
Seeds in the canopy of tree-like alderleaf mountain-mahogany plants survived fire in the foothills of Madera County, California. Fourteen- to sixteen-foot-tall trees burned in a prescription fire in July of 1949. Some trees were heavy with seed at the time of the fire. Trees were killed but seeds were not. Three days after the fire, true mountain-mahogany seedlings were dense. In the summer of 1951, there was on average a 2-year-old seedling per every 3 ft² (0.3 m²). Researchers indicated that seed survival in the canopy would be unlikely during an "intense summer" fire [28].
Following fall fires in southern California, seedlings were present as soon as the 2nd postfire year on chaparral sites. A total of 200± 100 (sx) alderleaf mountain-mahogany seedlings occurred on burned chaparral sites. No seedlings occurred on burned sage-scrub sites. No seedlings occurred in the 1st or 3rd postfire years. Approximately 33% of the total number of seedlings occurred equally in the 2nd, 4th, and 5th postfire years. Seedling survival was not reported [143].
Alderleaf mountain-mahogany cover was reduced, but seedling density increased following fire in a mixed chaparral/oak community with an annual herbaceous understory. Before the fire, interior live oak, which covered 60% of the plot, was selectively crushed and piled. This site preparation decreased alderleaf mountain-mahogany cover from 3% to 2%, and 1 and 2 years following the fire, cover was nearly zero. Seedlings were absent before the fire but 2 years after the fire there were 13 alderleaf mountain-mahogany seedlings in the plot. Fire season, conditions, or characteristics were not described [76].
Eighty-three percent of the postfire alderleaf mountain-mahogany population was sprouts on a low-elevation 1,800 feet (560 m) site in southern California. Postfire populations were 100% sprouts on mid- (3,300 feet (1,000 m)) and high- (5,480 feet (1,670 m)) elevation sites in the same area. Average postfire sprout heights were 27 inches (69 cm), 18 inches (45 cm), and 12 inches (31 cm) at the low-, mid-, and high-elevation sites, respectively. Seedling heights on the low-elevation site were not reported [143].
See Fire Studies in FEIS for further information on prescribed fire and postfire responses of many plant species. These Fire Studies are available for birchleaf mountain-mahogany:
Alderleaf mountain-mahogany reproduces by sexual and asexual means. Establishment from seed requires favorable conditions and is periodic [222,236]. Vegetative sprouts are produced following damage [1,47,251,332] but are also produced in the absence of damage [135].
Pollination: Flowers are wind-pollinated [151,153].
Breeding system: Alderleaf mountain-mahogany is monoecious [115,262].
Seed production: Seed production is variable in time and space and can be affected by browsing animals and climatic conditions. While Plummer and others [222] report that alderleaf mountain-mahogany produces abundant seed, Kitchen [151] indicates that a high percentage of seed produced is not viable. Seed crops according to Biswell and Gilman [27] are heavy some years and scant in others. Alderleaf mountain-mahogany was considered a "prolific" seed producer in Madera County, California, where in an undisturbed mature stand 1,900,000 seedlings/acre emerged in a single season [81].
Resources can be partitioned by alderleaf mountain-mahogany. One branch may produce a large amount of seed, while an adjacent branch produces little or no seed but produces considerable vegetative growth [306].
Unbrowsed alderleaf mountain-mahogany shrubs produced many more flowers and seeds than browsed shrubs in the Wasatch and Uinta mountains of Utah. Flower and seed production for both browsed and unbrowsed shrubs were greatest when precipitation was greatest [288].
Seed dispersal: Seeds are dispersed by wind and small mammals [1,33,151]. Wind-dispersed seed can travel up to 450 feet (140 m) from the parent plant [27].
High-quality alderleaf mountain-mahogany seeds are typically those 1st detached from the plant [194]. Seed shed earliest may be most vulnerable to predation. Likely the pattern of seed shed affects seedling germination and establishment.
Seed banking: A soft, thin seed coat and lack of seed dormancy suggest that alderleaf mountain-mahogany seed does not persist in the seed bank [1,27,28]. The absence of alderleaf mountain-mahogany seedlings on burned sites led researchers to suggest that seeds are not adapted to long-term survival in soil and/or are not resistant to fire [333]. Alderleaf mountain-mahogany was present in vegetation in the Huachuca Mountains of Cochise County, Arizona, but did not emerge from soils collected in same area. Abundance of true mountain-mahogany on the site was not reported. A small area of 3.04 inches² (19.6 cm²) of soil was sampled [233]. On 10-year-old burned sites in southern California mixed chaparral, C. m. var. glaber occurred irregularly and had low live cover (<10%) but did not emerge from soil collected on the site [331].
Seed stored in an open warehouse in Utah was evaluated periodically for viability. Temperature extremes in the warehouse during the 25-year period ranged from a low of -21.8 °F (-29.9 °C) to a high of 101 °F (38.3 °C). Stored seed was collected from Utah's Ephraim Canyon. In the 1st five years of storage, germination ranged from 61% to 68%. Germination decreased to 46%, 25%, and 3% after 7 years, 10 years, and 15 years of storage, respectively. No seed germinated after 20 and 25 years of storage [266]. Viability of alderleaf mountain-mahogany seed was significantly (p<0.05) better in cold than in hot or variable hot and cold environments. Germination of alderleaf mountain-mahogany seed after 6 years of storage at -5 to -10°F (-21 to -23 °C) was 84.4 %, at 36 to 44 °F (2.2-6.74 °C) was 81.6%, at 55 to 95 °F (13-35 °C) was 61.6%, and in a metal container kept outdoors at -15 to 105 °F (-26 to 41 °C) was 42% [258].
Germination: Requirements for successful seed germination differ depending on the locality of production. Adequate moisture is often correlated with successful germination. In chaparral vegetation, germination is best in mature stands [97]. Alderleaf mountain-mahogany seed produced in California germinates readily [1,27]. Seed collected in California germinated soon after planting in pure river sand kept in a greenhouse at 60 to 70 °F (16-21 °C) [189]. However, some indicate that alderleaf mountain-mahogany seed requires 1 to 3 months of cold stratification [125]. Germination of seed collected at more northerly locations in New Mexico increased with increased duration of stratification, and the improvement was greatest for seeds collected from the 2 highest elevation sites. Improvement was least for seed collected in the furthest south of the 6 northern sites. Stratification did not improve germination of seed collected from the southernmost latitudes [243].
Moisture, climate, and site conditions: Increased germination of seed collected in Wyoming, Colorado, and Utah was linked to moisture in several independent studies. Seeds collected in Wyoming's Laramie Basin had 80% germination after 48 hours of soaking in water. The germination percentage was lower (57%) after 96 hours at 41 °F (5 °C) but without soaking [34]. Alderleaf mountain-mahogany seed germination was poor with alternating moist and dry conditions. Seed was collected in native habitats that were not described. Germination was 10% under the moist-dry treatments, much lower than the maximum germination of 65% using other tests, which were not described [92].
Smith [257] found that site and climate affected alderleaf mountain-mahogany seed germination and seedling establishment in ponderosa pine/bunchgrass communities in Colorado's Pike National Forest. Greenhouse, nursery, and field studies were conducted. Seed was collected from sites with different soils. Precipitation between the beginning of April and the end of August affected seed viability. Achenes collected from northwestern slopes were heavier, had a greater percent filled, and had higher germination percentages than seed collected on southwestern slopes. The largest and most viable seeds were collected from 2 sites with the least fertile soils. Average germination was 33% in a dry year and 80% when precipitation was normal or above. Although shrubs on infertile sites produced the most viable seed, a higher percentage of seedlings survived in fertile than infertile soils. Survival was nearly equal on north and south aspects, but seedlings grew fastest and were most "vigorous" on southern slopes. Pocket gophers and cottontail rabbits were important seedling predators [257].
Germination of alderleaf mountain-mahogany seed collected from 9 populations in Utah and Colorado correlated with winter precipitation but not elevation. Germination was 15% without any pretreatment. Germination increased with moist, cold treatments. After 2 weeks of moist chilling, germination averaged 28.6%; after 4, 6, 8, and 12 weeks germination was 57.0%, 83.4%, 89.7%, and 98%, respectively. Researchers suggested that snowpack levels of the sites may have affected the length of moist chilling required for seeds [152].
Seed size: Large-sized seeds germinated best out of 5 seed lots collected in northern New Mexico. Seeds less than 0.3 inch (0.8 cm) long were considered small, seeds 0.4 inch (0.9-1 cm) long were classified as medium-sized, and large seeds were greater than 0.4 inch (1 cm) long. Seeds were stratified at 38 to 42 °F (3.3-5.6 °C) for 28 days, then kept moist at room temperature. Germination of small-, medium-, and large-sized seeds averaged 37%, 54%, and 69%, respectively. Germination differences were significant (p≤0.05). Seed size alone did not explain germination differences. The seed lot with the largest number of large-sized seeds had the lowest average germination percentage [219].
Heat treatments: Alderleaf mountain-mahogany seed collected below 6,600 feet (2,000 m) in southern California germinated readily on moist filter paper after 1 month of stratification but was significantly (P>0.001) reduced by 5 minute heat treatments of 250 °F (120 °C). Light and dark treatments produced very similar germination percentages, and seeds moistened with a solution of charred chamise (Adenostoma fasciculatum) were not adversely affected [138].
Seedling establishment/growth: Successful alderleaf mountain-mahogany seedling establishment is often correlated with adequate moisture. In chaparral communities, seedling establishment is sporadic and likely restricted to high moisture years [236]. Seedlings are vulnerable to frost and drought, and on harsh sites, establishment occurs only during "favorable" conditions [222]. Keeley [137] indicates that seedlings require mesic conditions and a well-developed litter layer. During a wet winter in San Diego County, alderleaf mountain-mahogany seedlings were abundant although they were absent the previous 5 years [333]. The number of seedlings and saplings varied from 0 to 36,530/ha in California chaparral vegetation that was unburned for 56 to 118 years. For more on this study, see Succession in chaparral [140]. Brooks [34] found no alderleaf mountain-mahogany seedlings in vegetation types adjacent to alderleaf mountain-mahogany stands in Wyoming's Laramie Basin and suggested that soil differences between the distinct vegetation types may have affected seedling establishment. In undisturbed alderleaf mountain-mahogany stands in Madera County, California, 1,900,000 seedlings/acre were produced in a single season. Mortality, however, was 98% after 2 years [81].
Moisture and site conditions affect establishment and growth: Woodmansee [323] studied seedling establishment in northwestern and north-central New Mexico. Germination occurred in 1967 when summer rainfall was above normal but not in 1968 when summer rainfall was again above normal. Timing of the rainfall may have been a factor. Spring emergent seedlings did not survive. Seeds germinated on or just below the soil or litter surface. Just a small number of seedlings were observed from 1967 to 1969, but survival was high (50%-100%) for the 1st year. Surviving seedlings grew in areas that were shaded for most of the day by associated vegetation and where herbaceous vegetation was scarce.
In northwestern Nebraska, aspect affected seedling establishment. Seedlings established in populations on southwestern but not on northeastern slopes. When populations were visited 10 years later, the researcher indicated that plant growth was slow and that the number of stems per plant decreased with plant age [205]. Alderleaf mountain-mahogany seedling production, survival, and growth were monitored on an herbaceous-dominated site in the western foothills of Tehama County, California. Grasses were predominantly annuals and included soft chess (Bromus hordeaceus), red brome (B. rubens), and foxtail fescue (Vulpia myuros). Twenty-one C. m. var. glaber seedlings emerged in the spring of 1948; of those, 3 survived to the spring of 1960, and seedling height averaged 7.2 inches (18 cm) [27]. Additional information from Smith [257] on seedling survival as it related to site conditions is available in Germination.
Postfire seedling establishment: Many report that alderleaf mountain-mahogany seedlings are rare in the early postfire years [139,146] and that seedling establishment is most typical in mature stands [97]. Keeley [140] indicates that although seed production soon following fire can be substantial, seedlings are uncommon in chaparral communities less than 50 years old. For more information on postfire seedling production, see Seed survival and postfire seedling production.
Mature plant growth: Annual twig growth of browsed and unbrowsed shrubs was greatest when precipitation levels were greatest in Utah's Unita and Wasatch mountains [288].
Vegetative regeneration: Alderleaf mountain-mahogany regenerates through asexual means following aboveground stem removal from browsing and/or fire; however, asexual sprouting also occurs in the absence of aboveground damage. Researchers noted lignotubers on some C. m. var. glaber plants in Nuttall's scrub oak-dominated chaparral vegetation in southern California [164]. Root crown sprouting following fire is considered "vigorous" and "rapid" [1,33]. Sprouts from rhizomes (~3 feet (1 m) from the main stem) occur as well [146]. Fire severity may affect postfire sprouting. Researchers found more and taller spouts in areas burned at low severity than on unburned sites or sites burned in high-severity fires in Colorado. Root crown and epicormic sprouts occurred on the low-severity burned sites. Only root crown sprouts occurred on high-severity burned sites, and only epicormic sprouts occurred on unburned sites [171].
Plants that received over 300% utilization, meaning current annual growth and previous year's growth were removed, compensated but did not overcompensate for removal. Browsing occurred for 4 years, but utilization of 300% was typical for only 1 year. Alderleaf mountain-mahogany cover was often lower on browsed than protected sites, but the height of browsed and unbrowsed shrubs was similar. Animals using unprotected sites were cattle, moose, elk, antelope, and deer [288,289]. In southwestern Colorado, the current annual growth and old wood to 0.2 inch (4 mm) above ground were removed from alderleaf mountain-mahogany shrubs. Early in the study, shrubs produced new sprouts, but no plants survived more than 6 years of the severe clipping treatments [251].
Sites in the Silverado Canyon in the Santa Ana Mountains of California, which were unburned for at least 60 years, revealed stem age structures that suggested new stems sprouted in the absence of fire [135]. True mountain-mahogany produced sprouts from root crowns and from rhizomes ~3 feet (1 m) or more away from the main trunk in California chaparral that was unburned for 56 to 120 years. The largest number of rhizome sprouts occurred on sites overtopped by other vegetation [140]. For additional information on sexual and asexual recruitment in this area see Keeley [141] in Succession in chaparral.
Alderleaf mountain-mahogany grows best in full sun [151], tolerates early-seral site conditions, and is dominant in several late-seral or climax communities [330]. For additional information on late-seral or climax communities where alderleaf mountain-mahogany is dominant, see Habitat types and plant communities. Following disturbances in areas where true mountain-mahogany is established, colonization by this species is rapid.
In southern California's San Sevaine area, alderleaf mountain-mahogany occurs in both pioneer and mature alluvial scrub vegetation. Pioneer communities, in inactive stream channels or recently scoured stream beds, are characterized by sparse vegetation and low species diversity. Mature communities are dominated by shrubs and subshrubs [101]. Alderleaf mountain-mahogany is a common understory species in old-growth ponderosa pine forests in southwestern Colorado's San Juan National Forest. Old-growth stands are those that have passed through the stand initiation stage and stem exclusion stage of development [241]. In Utah's Wasatch chaparral, alderleaf mountain-mahogany vegetation occurs on rocky ridges and represents a stage in xeric site succession and not a late-seral or climax vegetation type [108].
Succession in chaparral: Succession following disturbances in chaparral vegetation involves little or no compositional change [333]. The typically self-replacing recovery in this vegetation type has been termed autosuccessional. Shrubs present in mature stands, including alderleaf mountain-mahogany, are normally present in the 1st postfire year [99].
Several studies report abundance of alderleaf mountain-mahogany in different aged chaparral communities. In chaparral communities in the San Gabriel and San Bernardino mountains, alderleaf mountain-mahogany cover was 6.5% in 2- to 8-year-old stands, 15.5% in 9- to 21-year-old stands, 4.6% in 22- to 40-year-old stands, and 4.2% in 41- to 96-year-old stands [99]. Researchers suggested that alderleaf mountain-mahogany may be reduced by taller shrubs such as Nuttall's scrub oak, hollyleaf redberry (Rhamnus ilicifolia), and/or hollyleaf cherry (Prunus ilicifolia) when chaparral stands approach 50 years old [102]. However, alderleaf mountain-mahogany density was greater in ancient (130-150 years unburned) than mature (50-60 years unburned) chaparral vegetation in the Sierra Nevada. Density in mature stands was 209/ha and in over-mature stands was 487/ha [147].
Succession in pinyon-juniper: Alderleaf mountain-mahogany typically appears in the shrub/grass stage of postfire succession in pinyon-juniper communities. This stage can appear immediately following fire and cover may reach prefire levels within 7 years if pinyon-juniper cover was low (10% to 15%) when burned. If pinyon-juniper cover was 60% or greater when burned, sprouting shrub cover may be less than 2% in the 10th postfire year [82]. In Daggett County, Utah, alderleaf mountain-mahogany coverage was compared in different-aged Colorado pinyon-Utah juniper stands. Alderleaf mountain-mahogany cover decreased with increased pinyon-juniper canopy cover which generally increased with stand age. A summary of the results is presented below [122]:
Seral stage Mid Mid Late Late Approximate stand age (years) 70 150 215 140 Colorado pinyon-Utah juniper cover (%) 18.5 29.2 53.9 62.1 Alderleaf mountain-mahogany cover (%) 29.6 12.7 0.9 0Postfire succession: Many studies have compared alderleaf mountain-mahogany abundance on sites with different fire histories and increasing time since fire. Alderleaf mountain-mahogany abundance may vary with time since fire, fire history, site conditions, and/or prefire stand characteristics.
Alderleaf mountain-mahogany coverage varied on sites with different recent fire histories and substrates in California's Santa Monica Mountains. All sites burned 4 years before measurements were made. Substrates included andesite, granite, and sandstone. On sites that had burned 22 years earlier, the highest coverage was 82.2%. On sites that burned 75 years before the most recent fire, the highest coverage was 29%. The inclusion of different substrates and lack of prefire or unburned comparisons makes assessing the importance of fire regime difficult [176].
In cis-montane chaparral vegetation of San Diego County, California, alderleaf mountain-mahogany frequency and cover were relatively unchanged from 1930 to 2001. Coverage and frequency were 3% and 10%, respectively, in 1930. Between 1930 and 2001, 41% of measured plots burned once and 40% burned 2 or more times. In 2001, alderleaf mountain-mahogany coverage was 3% to 4% and frequency was 13% [75].
Alderleaf mountain-mahogany was present on all sites in a chronosequence ranging from 4-year-old burned sites to a 400-year-old climax Colorado pinyon-Utah juniper/mountain brush woodland in Mesa Verde National Park, Colorado. Alderleaf mountain-mahogany cover, 14%, and frequency, 28%, were greatest in ~400-year-old climax stands. Alderleaf mountain-mahogany cover and frequency were 1% and 6% on 30-year-old burned sites and 2% and 10% on 90-year-old burned sites. Presence was indicated on 4-year-old burned sites, but values were not reported. Most sites burned in summer fires, but other fire characteristics were not reported [67,68].
Importance of alderleaf mountain-mahogany generally increased as time since fire increased in mixed conifer forests in the White Mountains of south-central New Mexico. Importance values were the sum of relative density, relative basal area, and absolute frequency. Alderleaf mountain-mahogany was absent from sites burned 3 years earlier, but importance values were 50, 104, 75, and 104 on 16-, 21-, 27-, and 80-year-old burned sites, respectively. Researchers reported that shrub stages could persist for up to 80 years and that substantial increases in conifers took 50 to 100 years [103].
The successional recruitment of alderleaf mountain-mahogany in California chaparral was studied in 50- to 120-year-old stands. The percentage of dead basal cover was greatest (34.1%) at the oldest site. Alderleaf mountain-mahogany basal coverage and density were high in both the 74- and 83-year-old stands. Seedling and sapling production were absent in the oldest and youngest stands. Seedling density was by far the greatest in the 83-year-old stand, which authors reported was open and disturbed. Time since fire was not a good predictor for seedling and/or sapling growth. Data collected during this study are summarized below [140,141].
Site Pacific View Bartlett Spring Sequoia Silverado Time since fire (years) 56 74 83 118 Latitude 34° 05' 39° 09' 36° 28' 33° 45' Elevation (m) 375 1,075 1,200 950 Basal coverage (m²/ha) 3.24 8.47 13.92 1.32 Density (genet/ha) 610 820 670 480 Ramets/genet 65.8 17.5 23.2 7.3 Seedlings, saplings/ha 0 50 36,530 0 Stems produced in 1st decade after fire (%) <1 0 4 0 Stems produced in most recent decade (%) 92 67 85 61The scientific name of alderleaf mountain-mahogany is Cercocarpus montanus Raf. (Rosaceae)
[30,47,64,115,132,133,313,317].
Many varieties are recognized. Throughout this review, varieties will be identified using their
scientific names.
Cercocarpus montanus var. argenteus (Rydb.) Martin [86,178,223], silverleaf mountain-mahogany
Cercocarpus montanus var. blancheae (Schneider) Martin [177], island mountain-mahogany
Cercocarpus montanus var. glaber (Wats.) Martin [86,114,115,133,223], birchleaf mountain-mahogany
Cercocarpus montanus var. macrourus (Rydb.) Martin [177], Klamath mountain-mahogany
Cercocarpus montanus var. minutiflorus (Abrams) Martin [177], smooth mountain-mahogany
Cercocarpus montanus var. montanus [47,178], alderleaf mountain-mahogany
Cercocarpus montanus var. paucidentatus (Wats.) Martin [178,223], hairy mountain-mahogany
Many consider alderleaf mountain-mahogany a valuable component in revegetation or reclamation efforts. Seed and/or plants are available commercially [118,291]. Alderleaf mountain-mahogany provides soil stabilization [125,179], has fair to good drought tolerance [125], fixes nitrogen [3,322], and provides important wildlife browse and cover [264].
Site conditions and community types within the Intermountain West that are best suited for true mountain-mahogany revegetation are provided in [267]. Guidelines on how and where to plant true mountain-mahogany in New Mexico are given in [6]. Alderleaf mountain-mahogany seed collection, cleaning, and storage information is available in [127], and guidelines for producing alderleaf mountain-mahogany container seedlings is summarized in [165].
Alderleaf mountain-mahogany is difficult to establish from seed [221], and low seedling "vigor" and "slow initial growth rates" are typical [193]. Seed may be locally adapted and may not do well without proper stratification. Using seed collected near the proposed planting site is recommended [152,195]. Seeding is typically done in the fall; for information on seedbed preparation and seeding rates, see [195]. Alderleaf mountain-mahogany seed needs to be planted in the soil at depth of ~0.5 inch (1 cm); broadcast seeding is seldom successful. Researchers recommend against planting alderleaf mountain-mahogany with herbaceous species. For more seeding and/or planting tips, see [250]. If planting conditions are favorable, establishment and survival are not compromised by low initial growth rates [193]. Successful establishment of true mountain-mahogany from bareroot stock is considered moderate. Container-grown plants do not establish as well [263]. Due to the slow establishment of alderleaf mountain-mahogany (4-6 years), revegetated sites should be protected from livestock grazing for at least 4 years, and protection from native browsers may also be necessary [265].
Results of revegetation projects: Researchers seeded and planted container grown alderleaf mountain-mahogany on 40-year-old burned and unburned open ponderosa pine stands in South Dakota's Black Hills National Forest. Plantings were protected from browsers and monitored for 10 years. Seeded alderleaf mountain-mahogany survival averaged 4.2% on burned and 3.6% on unburned open stands in postfire year 10. Container grown plantings survived better; survival averaged 6.7% on burned and 18.2% in unburned open stands [59]. Survival of alderleaf mountain-mahogany was as high as 95% six years after planting container-grown plants in southwestern Utah [71].
Results of mine reclamation projects: Alderleaf mountain-mahogany survival was 25% two years after planting on a surface-mine site in Oak Creek, Colorado. Survival did not differ on overburden sites and sites with topsoil [29]. Alderleaf mountain-mahogany survival was 0% on surface coal mined reclamation sites in grassland vegetation of Colorado but was 25% on mine sites in mixed shrub vegetation sites [121].
Cercocarpus montanus is a North American species of shrub or small tree in the family Rosaceae native to northern Mexico and the western United States. It is known by the common names alder-leaf mountain-mahogany, alder-leaf cercocarpus, and true mountain-mahogany.[2][5] The variety argenteus is commonly known as silverleaf mountain-mahogany.[2]
Cercocarpus montanus is common in chaparral scrub, on mesas, the lower foothills of the Rocky Mountains, and the Great Plains in the United States.[6] Its range extends from Montana, Idaho, and South Dakota south as far as Sonora, Durango, and Nuevo León.[2][7][8]
Cercocarpus montanus often remains under 1 metre (3.3 ft) in height because of browsing by elk and deer, but can reach 20 feet (6.1 m). It has thin and smooth bark.[5] The species is considered to be long lived.[9]
It is also eaten by yellow-haired porcupine.[10]
Alder-leaf mountain-mahogany is sometimes grown as a drought tolerant garden plant, particularly in its native range. It is relatively unobtrusive in most seasons, but is noted for the beauty of is seed in the fall. The wildflower writer Claude A. Barr said that the shiny hairs on the curled plumbs attached to the are particularly attractive in the sun.[11]

Flowers appear red when they first open

Flowers are yellow when fully expanded
Cercocarpus montanus is a North American species of shrub or small tree in the family Rosaceae native to northern Mexico and the western United States. It is known by the common names alder-leaf mountain-mahogany, alder-leaf cercocarpus, and true mountain-mahogany. The variety argenteus is commonly known as silverleaf mountain-mahogany.
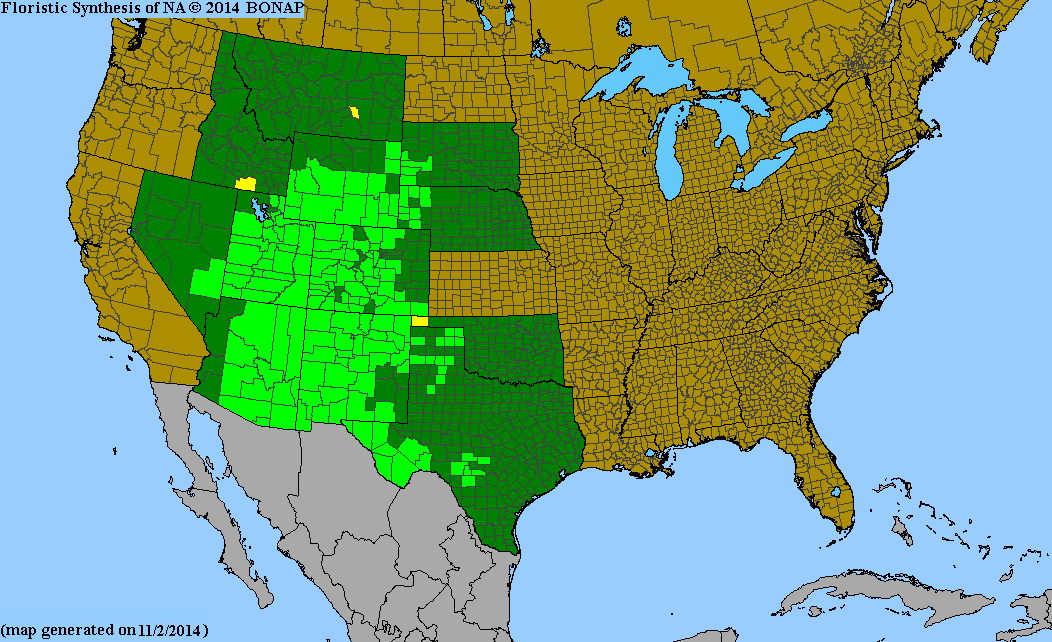{kind=link}