Comprehensive Description
provided by Smithsonian Contributions to Zoology
Planaria torva (Müller)
This is another eurythermic species with a wide distribution in Europe. Stankovi (1960:253) reports that one specimen was observed at the mouth of a brook at Studenište, but none in the lake itself. I have not collected any mature specimens that could be assigned with certainty to this species.
Phagocata Leidy
The genus Phagocata in the wider sense, as indicated in my index (Kenk, 1974:40), is admittedly a very heterogeneous genus, widely distributed over the Northern Hemisphere. Most of the species of Phagocata described up to the early part of this century were placed by their authors in the genus Planaria. Komárek (1926a:9–10) was the first to attempt a subdivision of the European species of that genus when he arranged the representatives of Planaria into several genera and establish Fonticola and Albiplanaria for several white species of planariids in which the common oviduct opens into the genital atrium; the male atrium is not surrounded by radial muscle plates; and adenodactyls are lacking. Kenk (1930:293) and de Beauchamp (1932:272) did not accept Albiplanaria as a separate genus and included it in Fonticola. The genus, as defined above, today comprises a great number of species inhabiting Europe, Asia (including the Japanese islands), and North America. Hyman (1937:302) pointed out that one of the American species of the genus, Planaria gracilis Haldeman, had been placed by Leidy (1847:248) into a new subgenus, Phagocata, which predates Komárek's Fonticola and should therefore be applied to all species placed into Fonticola. This suggestion was followed by Dahm (1949), Gourbault (1972), and Kenk (1974) but was objected to by several European investigators, particularly Komárek (1953a:275) and Reisinger (1960:289–290).
It is true that the about 50 species, now included in Phagocata because of the scheme of organization of their reproductive systems, represent a great variety of external features as well as specific anatomical characters. It would be very desirable if one could divide them at least into subgenera that would reflect natural (evolutionary) groups that would make sense also from a zoogeographical standpoint (see Ball, 1976:417–418). The three North American polypharyngeal species (Phagocata gracilis, P. woodworthi, and P. nordeni) with almost identical external habit but clearly different anatomical features, could be united in a subgenus Phagocata sensu stricto, as suggested by Reisinger (1960:290). Fonticola was used as a subgenus by Gourbault (1972:29) who, however, did not formally define it. Atrioplanaria de Beauchamp (1932:334), with five or six European species, may be considered a valid genus because of morphological as well as karyological peculiarities (Gourbault and Benazzi, 1977). Livanov and Zabusova (1940:96) established a separate genus Penecurva for four Asian species of different external aspects, in which the ejaculatory duct opens on the ventral side of the penis papilla rather than on its tip. Such a condition is observed also in several American species that have little else in common with the Asian forms. Thus, Penecurva is hardly tenable even as a subgenus.
When Fonticola is established formally as a subgenus or genus with a clear definition, it will comprise all Phagocata species of the Ohrid region, since the type-species of Fonticola, Planaria olivacea O. Schmidt, 1961, is a member of this species group.
The European species of Phagocata, generally designated under Fonticola, eliminating Atrioplanaria, are very similar in their external appearance: unpigmented (white, with one exception), two-eyed planariids with a truncate head, lacking prominent auricular appendages and anterior adhesive organs. The interspecific differences concern mainly their anatomical characters. Reisinger (1960: 289–291), a meticulous investigator of this group, lists the following features as distinctive attributes: presence or absence of a spermatopositor (a stiff tube attached to the spermatophore); number and arrangement of the excretory pores; epithelial conditions at the transition from the copulatory bursa to the bursa stalk; number of retinal clubs in the eyes; differences in the postembryonic development of the copulatory apparatus; and characteristic behavior of the animals in the search for food or for a copulatory mate. Not all these characters can be identified in preserved specimens; some must be investigated in living animals by special techniques.
The following species may be considered to be well established and readily distinguishable morphologically: Phagocata vitta (Dugès), P. albissima (Vejdovský), P. olivacea (Schmidt), P. paravitta (Reisinger), P. bosniaca (Stankovi), P. undulata (Stankovi), and P. leptophallus (Reisinger). On the other hand, the affinities of four species, distributed chiefly on the Balkan Peninsula, P. macedonica (Stankovi), P. dalmatica (Stankovi and Komárek), P. ochridana (Stankovi and Komárek), and P. illyrica (Komárek), have been questioned, with good reason, by de Beauchamp (1932), Dahm (1958, 1967), and even by one of their authors, Komárek (1953a:277; 1955:170). Not all the criteria enumerated by Reisinger can be applied to these species. I shall retain them for the time being until further detailed investigations can be carried out.
Phagocata ochridana (Stankovi and Komárek)
Fonticola ochridana Komárek, 1926a:9 [nomen nudum].
Fonticola ochridana Stankovi and Komárek, 1927:643.
Fonticola ohridana.—Stankovi, 1932:577.
Fonticola albissima var. dalmatica.—de Beauchamp, 1932:319 [in part].
Phagocata (? = Fonticola) ochridana.—Dahm, 1964:486.
Phagocata (Fonticola) ochridana.—Gourbault, 1972:32.
Phagocata ochridana.—Kenk, 1974:42.
MATERIAL DEPOSITED.—Sagittal serial sections of 7 specimens on 9 slides, USNM 55299–55305.
A good description of this species was given by Stankovi and Komárek, based on materials from springs and streams in the Ohrid region.
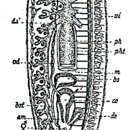
EXTERNAL FEATURES (Figure 15).—This is a rather slender species, sexually mature specimens measuring up to 8 mm in length and 0.8 mm in width. The anterior end is truncated, with a convex frontal margin and rounded lateral corners. There is no narrowing or neck behind the head. The lateral body margins diverge only slightly behind the anterior end, then run parallel up to the level of the copulatory complex where they begin to converge to form a bluntly pointed or nearly rounded tail end. The two eyes are far removed from the frontal margin and lie close together at a distance of one-fifth to one-fourth the width of the head. The body is unpigmented, white. The pharynx is inserted at about the middle of the body or slightly behind it and measures one-seventh the body length. The copulatory apparatus occupies the anterior two-fifths of the postpharyngeal region. The anterior ramus of the intestine ends at, or somewhat behind, the level of the eyes. The animals move by gliding only, never by “crawling.”
ANATOMY.—The testes extend from the level of the ovaries backward to near the posterior end. They are situated, in general, nearer to the ventral than to the dorsal body wall, though a few testes may be placed in a more dorsal position and individual testes may occupy almost the entire dorsoventral diameter of the body. The structure and situation of the ovaries show no pecularities. Vitellaria or yolk glands are found in the mesenchyme of the entire body, except the head, and are particularly numerous in the spaces lateral to the zone of testes.
In the copulatory apparatus (Figure 32), the genital opening leads directly into the genital atrium which is not divided into the usual two chambers. In several specimens, however, histological differences were seen in the epithelium lining the atrial cavity. The anterior part, which would correspond to a male atrium, had a normal, cuboidal or cylindrical, nucleate epithelium, while in the posterior section, the common atrium, the epithelium was infranucleate.
The size, shape, and position of the male copulatory organ appear, in the majority of my specimens, as shown in Figure 32. The bulb of the penis is of moderate size and hemispherical. Its musculature is rather weak, the muscle fibers being densest near the periphery of the bulb. The free penis or penis papilla is conical or finger shaped, somewhat curved ventrally, its pointed tip reaching to the dilated genital pore. The papilla is covered with a cuboidal or flattened epithelium, beneath which is a well-developed layer of circular muscle fibers, followed by a thinner layer of longitudinal muscles. The two vasa deferentia enter the penis bulb laterally and acquire a coat of circular muscles. They first pass toward the midline of the organ, then turn posteriorly, enter the base of the penis papilla, and open closely together, but separately, into the penis lumen. This lumen is a straight canal, the ejaculatory duct, running in the axis of the papilla and opening at its tip. Histologically, it is differentiated into three sections. A short anterior portion is lined with very tall cells, the distal parts of which are bent posteriorly and form a villus or plug filling the lumen of the canal. This part has been likened to a seminal vesicle by Stankovi and Komárek. The following section of the canal has a rather flattened epithelium, the cells of which are pierced by numerous gland ducts with a granular, strongly eosinophilic secretion. The third, most distal region of the penial lumen is usually somewhat widened and lined with taller cells with apparently secretory function. It narrows again at the tip of the papilla. At least the anterior two sections of the penis lumen have a thin coat of circular muscle fibers. A considerable variation in the length and shape of the penis was observed even in animals collected in the same locality. These differences are, undoubtedly, due to muscular contractions of the organ. The two main muscular systems of the penis, i.e., the peripheral layer of the penis bulb and the powerful circular muscles of the papilla, both effect, when contracted, an elongation of the penis and its protrusion through the gonopore; much of the soft parenchymal tissue of the bulb is pressed into the base of the penis papilla. On the other hand, a relaxed state of the muscles will result in a shorter and broader shape of the organ; such a retraced penis has a more voluminous bulb. The outer covering of the papilla appears to be a cuboidal epithelium in the shortened penis, a flattened one in the elongated organ. The histological differentiation of the penial lumen is related to its function of forming a spermatophore during (or before?) copulation. The most anterior section, though it is not widened in this species, may play the role of a seminal vesicle as Stankovi and Komárek (1927:625) assume. A comparison with closely related species indicates that the middle part of the lumen produces the chitinoid stalk or spermatopositor, and the distal section the capsule of the spermatophore (cf. “Fonticola albissima var. illyrica” in de Beauchamp, 1932:317).
The two oviducts, running above the nerve cords, are equipped with accessory seminal receptacles or modified yolk funnels, as Reisinger (1963:686–687) has demonstrated. In the region of the copulatory complex they ascend dorsally and medially, the left one passing between the genital atrium and the bursal stalk. They unite dorsally to the atrium to form a rather long, somewhat curved common oviduct that opens into the posterior part of the atrium. The end parts of the paired oviducts and the greater part of the common oviduct are connected with numerous eosinophilic shell glands. The copulatory bursa varies in size and shape according to the state of maturity. Its lumen frequently contains remainders of one or more spermatophores and/or spermatopositors. The bursal stalk proceeds from the bursa dorsally to the penis, then shifts to the left of the midline, increasing gradually in its outer and inner diameters, and joins the genital atrium from the left side near the gonopore. Sometimes a differentiation of the epithelium lining of the duct is seen. The anterior part, adjoining the bursa, always has a normal, nucleate, cuboidal epithelium, while the posterior section, connecting with the atrium, may show partly or wholly infranucleate cells.
Numerous eosinophilic glands open into a large area of the epidermis surrounding the genital aperture, the function of which is not quite evident. Their secretions may serve for the attachment of the egg capsules (cement glands) or else may play a role during copulation.
DISTRIBUTION AND ECOLOGY.—Phagocata ochridana occurs in the Ohrid region in a great variety of habitats. Stankovi and Komárek found it originally in springs near Sveti Naum and in the large limestone spring Šum. Later, Stankovi (1934:167; 1938:9; 1955b:288) extended its known occurrence first to the littoral zone of Lake Ohrid and to littoral springs of Lake Janina (Ioannina) in the Greek Epirus, and then to the sublitoral and profundal zones of Lake Ohrid. Obviously, Stankovi's interpretation of the species includes a form that was designated by Reisinger (1960:274) as a separate species, P. stankovici.
I collected Phagocata ochridana in the following localities: Studenište, at the Hydrobiological Station, in cold springs and warmer pools, under stones, and also on water plants; Bej-Bunar, northeast of the station, where it is rare in the cold spring itself, more common in the creek and in ditches with warmer water, and under stones; a small spring near Elešec (north of the village of Peštani), together with Dugesia gonocephala; springs at the monastery of Sveti Naum; spring Šum (about 1 km south of Zagraani, near Struga), where the species is absent in the cold springs (8.2° C, 7 September 1935) but occurs in warmer parts (up to 11.7° C), under stones; an irrigation ditch at Teferi, west of Struga; in Lake Ohrid, the species was found in Ohrid Bay and near Kalište from the shoreline down to the shell zone.
A lot of Phagocata ochridana was cultured for two generations in the laboratory, with beef liver offered as food. No asexual reproduction was observed in the cultures.
TAXONOMIC POSITION.—Phagocata ochridana was first recorded by Stankovi (1926:238) as a continental variety of P. olivacea (Schmidt). Stankovi and Komárek (1927:643) described it as a separate species. De Beauchamp (1932:319) considered it to be a probable synonym of P. dalmatica (Stankovi et Komárek), which he cited as “Fonticola albissima var. dalmatica.” This assumption was accepted also by Komárek (1953a:278) and by the majority of later authors who made reference to the species. On the other hand, Stankovi (1960:177) preferred to keep the species separate “for reasons of biogeographical order.” Reisinger (1963:687) pointed out that accessory seminal receptacles on the oviducts have been found only in a few species of “Fonticola,” F. ochridana, F. maculata, and F. leptophallus Reisinger, but apparently he did not examine P. dalmatica for their occurrence. Reisinger (1960:274) also separated from Stankovi's P. ochridana a form inhabiting the deeper zones of Lake Ohrid, which he designated as a new species, P. stankovici.
Considering that these three forms are still open questions, for the time being I am keeping the common Ohrid Phagocata as a separate species to avoid a possible confusion when a more thorough comparison can be made, an investigation that should include experiments on the possibility of interbreeding. It seems to me quite probable that all three will unltimately prove to be one species, Phagocata dalmatica (Stankovi and Komárek).
- bibliographic citation
- Kenk, Roman. 1978. "The planarians (Turbellaria, Tricladida Paludicola) of Lake Ohrid in Macedonia." Smithsonian Contributions to Zoology. 1-56. https://doi.org/10.5479/si.00810282.280