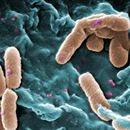
Pseudomonas is genus of Gram-negative aerobic bacteria that form large colonies when plated on assays, bear a motile tail or flagella, and have a slightly curved rod-shaped body. They are found living in almost every microbial ecosystem with a wide distribution due to their metabolism (Palleroni, 2010). Some Pseudomonas species are used in agricultural settings to protect seedling roots from fungus (Chin-A-Woeng et al., 2000), others are the culprits for much of the food spoilage in the the food industry, and a few can act as opportunistic pathogens as in the case of P. aeruginosa. They have an particularly dense cell wall that allows them to survive in unusual environments, are naturally resistant to penicillins, and form super sticky plaques or biofilms on surfaces (Hassett et al., 2002). Suitable habitats include soil, plastics, sterilized hospital structures, subzero temperature environments, and chemical containers such as alcohol bottles. Their cell walls have an additional structure called a porin that allows them to alter their surrounding by pumping out foreign substances like antibiotics, and organic compounds to resist environmental changes.
Pseudomonas és un gènere de bacils rectes o lleugerament corbats, gramnegatius, oxidasa positius, aeròbics estrictes encara que en alguns casos poden utilitzar el nitrat com acceptor d'electrons. El catabolisme dels glúcids es realitza per la ruta d'Etner-Doudoroff i el cicle dels àcids tricarboxílics. Alguns membres del gènere són psicròfils, mentre que altres sintetitzen sideròfors fluorescents de color groc-verdós amb gran valor taxonòmic. És comuna la presència de plasmidi si no formen espora.Pseudomonas literalment vol dir «falsa unitat», derivat del grec pseudo (ψευδο 'fals') i mones (μονάς / μονάδα 'una sola unitat'). El terme «monada» es feia servir en la microbiologia antiga per anomenar els organismes unicel·lulars.
Amb el recent anàlisi de seqüències del RNAr 16S s'han definit la taxonomia de moltes espècies bacterianes i com a resultat, el gènere Pseudomonas inclou algunes soques classificades anteriorment dins de les Chryseomonas i Flavimonas. Altres soques classificades prèviament en el gènere Pseudomonas, ara són agrupades en els gèneres Burkholderia i Ralstonia.
Pseudomonas (též pseudomonáda) je rod gramnegativních bakterií ze skupiny Gammaproteobacteria. V přírodě jsou velice hojné a díky své přizpůsobivosti osídlují velmi rozmanitá stanoviště. Některé druhy způsobují i onemocnění rostlin a živočichů včetně člověka; nejnebezpečnějším původcem lidských onemocnění mezi nimi je Pseudomonas aeruginosa. Co se týká vlastností, jsou pohyblivé a tyčinkovitého tvaru, obvykle aerobní a netvořící spory.
V nedávné době proběhla revize tohoto a dalších příbuzných rodů pomocí analýzy genů pro 16S rRNA.[1] Některé druhy, dříve řazené k rodům Chryseomonas a Flavimonas, jsou dnes řazeny mezi pseudomonády,[2] a naopak, jisté druhy původně považované za pseudomonády dnes vědci spíše řadí do rodů Burkholderia či Ralstonia.
Pseudomonad doslova v překladu znamená „zdánlivá jednotka“ a pochází z řeckých slov pseudo (ψευδο – zdánlivý, falešný) a monas (μονάς či μονάδα – jednotka, jednotlivina). Termín monáda se používal dříve v přírodovědě pro jednobuněčné organizmy, které první mikrobiologové spatřili pod mikroskopem.
Bakterie rodu Pseudomonas jsou v přírodě velmi hojné a vyskytují se například ve vodě či uvnitř těl rostlin, a proto byly objeveny již prvními mikrobiology. Rodový název Pseudomonas byl nejprve v roce 1894 definován poměrně volně jako označení pro gramnegativní tyčinkovité bakterie s polárním bičíkem. Vbrzku začalo množství druhů narůstat, neboť byly pseudomonády nacházeny v nejrůznějších prostředích. Až použití nových metod umožnilo vytvoření modernější klasifikace.[3] Po roce 2000 byly přečteny kompletní genomy několika kmenů bakterií z rodu Pseudomonas: např. P. aeruginosa kmen PAO1, P. putida kmen KT2440, P. fluorescens kmen Pf-5, P. syringae patovar tomato DC3000, P. syringae patovar syringae B728a, P. syringae patovar phaseolica 1448A, P. fluorescens PfO-1 a P. entomophila L48.[3]
Pro rod Pseudomonas jsou podle Bergeyho manuálu typické následující vlastnosti:[4]
Dalšími poměrně typickými rysy pseudomonád je (až na výjimky) vylučování pyoverdinu (fluoresceinu), žlutozelené fluorescenční látky, které umožňuje bakteriím získávat železo v prostředí chudém na tento prvek (tzv. siderofor).[5] K dalším vylučovaným sideroforním látkám patří pyocyanin u Pseudomonas aeruginosa[6] a thiochinolobaktin u Pseudomonas fluorescens.[7] Obvykle jsou pseudomonády pozitivní při oxidázovém testu, bývají beta-hemolytické (na krevním agaru), indol negativní, při aplikaci methylové červeni i při Voges–Proskauerovu testu jsou negativní, ale zase jsou citrát–pozitivní.
V rámci rodu je možné pozorovat velkou metabolickou rozmanitost, což umožňuje těmto bakteriím osídlovat široké rozpětí různých prostředí.[8] Snadná kultivace ve skle (in vitro) a dostupnost přečtených genomů z nich činí výborné modelové organizmy pro vědecký výzkum; nejčastěji se zkoumá P. aeruginosa jako významný lidský patogen, P. syringae jako patogen rostlin, půdní bakterie P. putida a P. fluorescens, jenž stimuluje růst rostlin.
Poměrně velké množství pseudomonád je schopno tvorby biofilmů. Spočívá to ve vylučování lepkavých polysacharidů do okolí buněk. Díky tomu mají bílé krvinky větší problém se pseudomonád v těle zbavit – fagocytóza v prostředí biofilmu je náročnější.[9] Navíc tyto bakterie lépe přilnou k povrchu a je například mnohem náročnější omýt takový biofilm z povrchu kuchyňských nástrojů. Díky tvorbě biofilmů mohou pseudomonády přežít i například v nádobách, v nichž se vyrábí léčiva. Mezi další podobně pozoruhodná místa, kde by možná málokdo čekal bakterie, jsou nádoby s antiseptickými kvartérními amoniovými sloučeninami nebo láhve s balenou minerální vodou.
Pseudomonády jsou gramnegativní, což je přirozeně činí odolné k takovým antibiotikům, jako jsou penicilin a ostatní beta-laktamová antibiotika, ale na druhou stranu jsou citlivé k piperacilinu, imipenemu, tobramycinu a ciprofloxacinu.[9] Odolnost k některým antibiotikům je často dána tím, že jsou schopné velmi rychle vypumpovat tato antibiotika z buňky. Z toho plynoucí rezistence vůči většině antibiotik je značný problém u patogenní bakterie Pseudomonas aeruginosa. Tato bakterie vlastní charakteristické geny pro odolnost – nazývají se mexAB-oprM, mexXY a další.[10] Jsou navíc velmi přizpůsobivé a často u nich mutací vznikne odolnost k antibiotiku, jež na ně doposud účinkovalo. Mnohdy si geny pro odolnost bakterie předávají takzvaně horizontálně. Navíc jim pomáhá život v biofilmech, kde jsou před antibiotiky lépe chráněny.[3]
V napadeném těle bakterie rodu Pseudomonas produkují řadu látek, z nichž mnohé jsou toxické. Patří k nim pyocyanin (váže se na flavoproteiny v elektronovém transportním řetězci), lecitináza (způsobuje rozklad lecitinu a tím zejména rozklad krvinek), kolagenáza (rozkládá kolagen a tím narušuje zejména vazivovou tkáň), lipázy (rozklad tuků), hemolysiny (rozklad krvinek) a fluorescein. Výsledkem souhry těchto a dalších jedovatých látek je v lidském těle chudokrevnost, nekróza (odumírání) tkání a poškození nervové soustavy.[11]
Pseudomonády, zejména P. aeruginosa, ale i např. P. oryzihabitans[12] a P. plecoglossicida,[13] jsou původci významných infekčních onemocnění člověka nebo jiných živočichů. Zejména infekce P. aeruginosa u člověka působí velké problémy v nemocničním prostředí (těžko léčitelné tzv. nozokomiální infekce). Vyvolává široké spektrum chorob: zánět průdušek, zánět plic, záněty různých ran a popálenin na kůži, infekce související s katetry v močové trubici, infekce ucha a očí, ale i celotělní často fatální záněty (bakteriémie). Zvláště u pacientů s cystickou fibrózou nebo s jinými onemocněními je obvykle infekce pseudomonádami smrtelná.[14]
Infekce způsobují u člověka i blízce příbuzné rody, z nichž některé byly dříve řazeny mezi pseudomonády. Proto se uvádí nemoc vozhřivka, způsobená bakterií Pseudomonas mallei (dnes Burkholderia mallei) a vzácně přenosná na člověka, nebo melioidóza, způsobená bakterií Pseudomonas pseudomallei (dnes Burkholderia pseudomallei).[11]
Zejména P. syringae je významným rostlinným patogenem. Rozlišuje se více než 50 různých patovarů, tedy jakýchsi kmenů s typickými patogenními vlastnostmi. Mnohé jsou například specializované na výhradního rostlinného hostitele a jiné napadají zřídka. Existují i další druhy pseudomonád schopné vyvolat onemocnění rostlin, a to zejména v podskupině pseudomonád kolem P. syringae.
Mimo to působí problémy v zemědělství i druh P. tolaasii, jenž je zajímavý tím, že způsobuje hnití pěstovaných hub.[15] P. agarici způsobuje zase nákazu v lupenech pěstovaných druhů hub.[16]
Pseudomonády jsou schopné růst za nízkých teplot a za nestandardních podmínek prostředí, takže nepřekvapí, že mnohé z nich mohou způsobovat i kažení potravin. Mléčné produkty napadá bakterie P. fragi,[17] vejce kolonizuje např. P. taetrolens and P. mudicolens,[18] zatímco P. lundensis způsobuje kažení mléka, sýru a masa včetně rybího.[19]
Od poloviny 80. let 19. století jsou pseudomonády využívány v biokontrole či také biologickém boji se škůdci zemědělských plodin. Nejlépe jsou v tomto ohledu známy výsledky postřiků obsahujících různé kmeny P. fluorescens (např. CHA0 nebo Pf-5). Přesto není jasné, jak P. fluorescens pomáhá růstu plodin. Podle jedné z hypotéz vyvolávají bakterie jakousi imunitní odpověď v rostlině, čímž zlepšují její následné schopnosti boje se skutečnými bakteriemi. Další možností je, že se pseudomonády po postřiku dostanou do půdy a vytlačí z ní patogenní bakterie, například pomocí sideroforních sloučenin „nasávajících“ z půdy železo. Možné také je, že pseudomonáda produkuje antibakteriální sloučeniny, jako jsou fenazinová antibiotika nebo kyanovodík. Všechny zmíněné teorie jsou do jisté míry podporovány experimentálními daty, ale situace není zcela vyřešena.[20]
Dalšími pseudomonádami se schopností biokontroly jsou P. chlororaphis (produkuje antibiotika ničící některé patogenní houby)[21] nebo jí blízce příbuzný druh P. aurantiaca (produkuje jistý fluoroglucid, antibiotikum proti grampozitivním mikrobům).[22]
Někteří zástupci pseudomonád jsou schopní přijímat z prostředí jisté znečišťující látky a metabolizovat je. Díky tomu mohou být použity v bioremediaci znečištěných půd či vod. Mezi druhy, jež je v této souvislosti možné zmínit, patří:
Moderní studie zabývající se taxonomií tohoto složitého rodu poněkud pokulhávaly za tradičními morfologickými postupy používanými v lékařské praxi, dokud se nenaskytla možnost srovnávat jedince pomocí stavby jejich ribozomální RNA (a dalších genů bakterií).[1] Tyto postupy spolehlivě prokázaly, že rod Pseudomonas tak, jak byl dříve chápán, je nepřirozený shluk mnoha různých rodů. Od té doby došlo k velkému pokroku a původní pseudomonády byly pomocí rRNA roztříděny do 5 skupin podle příbuznosti. Výzkumníci díky tomu byli schopni vyřešit problém s hromaděním druhů v rodu Pseudomonas. V první polovině 20. století byl tento rod zahlcen obrovským množstvím zástupců, od té doby se však počet uznávaných druhů zmenšil více než desetkrát. Jen velmi málo druhů přežilo toto období revize a většina pseudomonád v dnešním slova smyslu jsou druhy popsané v posledních letech.[3]
Druhy, které byly vyčleněny z rodu Pseudomonas, jsou uvedeny níže. Interní odkazy vedou na současné převládající taxonomické označení – povšimněte si, že mnohé dnes patří např. do rodů Burkholderia a Ralstonia.
V tomto článku byl použit překlad textu z článku Pseudomonas na anglické Wikipedii.
Pseudomonas (též pseudomonáda) je rod gramnegativních bakterií ze skupiny Gammaproteobacteria. V přírodě jsou velice hojné a díky své přizpůsobivosti osídlují velmi rozmanitá stanoviště. Některé druhy způsobují i onemocnění rostlin a živočichů včetně člověka; nejnebezpečnějším původcem lidských onemocnění mezi nimi je Pseudomonas aeruginosa. Co se týká vlastností, jsou pohyblivé a tyčinkovitého tvaru, obvykle aerobní a netvořící spory.
V nedávné době proběhla revize tohoto a dalších příbuzných rodů pomocí analýzy genů pro 16S rRNA. Některé druhy, dříve řazené k rodům Chryseomonas a Flavimonas, jsou dnes řazeny mezi pseudomonády, a naopak, jisté druhy původně považované za pseudomonády dnes vědci spíše řadí do rodů Burkholderia či Ralstonia.
Pseudomonas (fem.: die Pseudomonas, Plural für die der Gattung angehörigen Organismen: Pseudomonaden, von altgriechisch ψεῦδος pseúdos, deutsch ‚Täuschung‘, μονάς monás, deutsch ‚Einheit‘) ist eine Gattung stäbchenförmiger, mit polaren Geißeln sich aktiv bewegender, immer anaerob oder aerob veratmend, nie fermentierender, gramnegativer Bakterien. Sporen werden nicht gebildet. Das Genom der Art P. aeruginosa wurde vollständig sequenziert.[1]
Die Größe variiert zwischen 0,5–1,0 × 1,5–5,0 µm. Sie können sich mit einer oder mehreren polaren Geißeln aktiv bewegen (sie sind motil). Sie reagieren im Voges-Proskauer-Test negativ, im Katalase-Test positiv. Bei den meisten Arten fällt der Oxidase-Test positiv aus, z. B. bei Pseudomonas fluorescens, diese Arten besitzen somit ein c-Typ-Cytochrom.
Pseudomonaden sind aerob, das heißt, sie brauchen in der Regel Sauerstoff für ihren Energiestoffwechsel, um zu wachsen. Sie nutzen also einen oxidativen Energiestoffwechsel mit Sauerstoff als Elektronenakzeptor. Zucker werden dabei über den Entner-Doudoroff-Weg zur Energiegewinnung abgebaut. Nitrat kann bei den meisten Arten als alternativer Elektronenakzeptor statt Sauerstoff bei der Atmung dienen (Nitratatmung). In diesem Fall wird Nitrat (NO3−) zu elementarem, molekularem Stickstoff (N2) reduziert (Denitrifikation). Da für die Nitratatmung kein Sauerstoff nötig ist, sind einige Arten auch in der Lage, unter Ausschluss frei gelösten Sauerstoffs (anoxisch) zu wachsen, doch dies ist eher selten der Fall. Als Vertreter der nitratreduzierenden Bakterien werden die Arten Pseudomonas aeruginosa, Pseudomonas stutzeri und Pseudomonas denitrificans bei der Abfallbehandlung zur Nitratelimination eingesetzt. Eine der wenigen Arten, die nicht zur Nitratatmung fähig sind, ist Pseudomonas fluorescens.
Fermentation wird von dieser Gattung nicht als Energiestoffwechsel genutzt, alle Arten sind sogenannte Nonfermenter (Nichtfermentierer).
Mehrere Pseudomonas-Arten produzieren bei Eisenmangel Siderophore; mindestens 50 verschiedene Pyoverdine wurden gefunden, außerdem Ferrioxamine, die sonst nur in Actinobakterien vorkommen.[2]
Kulturen von P. aeruginosa zeichnen sich durch eine blau-grüne Färbung des Kulturmediums und süßlich-aromatischen Geruch aus.
Pseudomonaden sind ubiquitär, also allgegenwärtig in der Umwelt („Pfützenkeim“). Man findet sie im Boden, Wasser sowie in oder auf Pflanzen und Tieren. Pseudomonaden leben oft saprotroph, also von abgestorbenem organischen Material. Man findet sie auch häufig in der Rhizosphäre, diese von den Pflanzen beeinflusste Umgebung stellt ein gutes Habitat für Pseudomonaden dar, sie sind gut an die Rhizosphäre angepasst.[3] So bewegen sie sich unter anderem aktiv, sind chemotaxisch und resistent gegen spezielle Abwehrstoffe der Pflanzen. Obwohl einige pflanzenpathogen sind, tragen viele Arten auch zum Schutz der Pflanzen bei, indem sie innerhalb der Rhizosphäre andere Mikroorganismen verdrängen. Des Weiteren sind einige Arten wie Pseudomonas putida mit arbuskulären Mykorrhizapilzen assoziiert und fördern somit sogar das Wachstum der Pflanzen.[4] Man spricht von plant-growth promoting rhizobacteria (PGPR), zu Deutsch pflanzenwachstumsfördernde Rhizobakterien.
Die Pseudomonaden sind physiologisch hoch flexibel und können auch als opportunistische Krankheitserreger bei bereits geschwächten Pflanzen und Tieren auftreten. Von vielen Pseudomonas-Arten existieren pflanzenpathogene Stämme, so genannte Pathovare (Phytopathogene). Die Art Pseudomonas marginalis verursacht, meist an Sprossachsen, die Weichfäule. Pseudomonas syringae subsp. savastanoi pv. oleae verursacht die Tuberkelkrankheit beim Olivenbaum. Pseudomonas syringae pv. aesculi ist neben der Kastanienminiermotte und seltener Phytophthora-Arten (Oomycetes) ein bedeutender Verursacher des Kastaniensterbens, der den Baum so stark schädigt, dass er anfällig für opportunistische Pilze wird.[5][6] Bei Tieren kann u. a. die Fleckenseuche der Süßwasserfische von Pseudomonas verursacht werden. Arten wie Pseudomonas aeruginosa können für Menschen mit geschwächtem Immunsystem gefährlich sein.
Die Virulenzgene sind meistens mobil (Plasmide) und können leicht von einer Pseudomonas-Art auf eine andere, aber auch auf entfernter verwandte Bakterien übertragen werden. So besitzen die Pseudomonaden oft ein sogenanntes R-Plasmid, welches der Übertragung der Resistenzgene dient.
Das hochgefährliche Nervengift Tetrodotoxin (TTX) wird von einigen Pseudomonas-Arten produziert. Auch anhydro-TTX, eine im molekularen Aufbau leicht abweichende Variante, wurde bei dieser Gattung nachgewiesen. Pseudomonas und auch verschiedene Vibrionen werden für die starke Giftigkeit verschiedener Kugelfische verantwortlich gemacht.[7] Der Fisch nimmt die Bakterien vermutlich durch die Nahrung (zum Beispiel verschiedene Krebstiere, Würmer und Rotalgen) auf. Pseudomonas wurde bei dem Kugelfisch Takifugu poecilonotus (Synonym: Fugu poecilonotus) nachgewiesen.[8] Man geht hierbei von einer Symbiose zwischen dem Kugelfisch und den Bakterien aus. Der Kugelfisch setzt das Gift für die Verteidigung bzw. Abschreckung gegen Räuber ein und liefert den Bakterien wiederum ein geeignetes Habitat. Die Fähigkeit der Bakterien, insbesondere der Vibrionen, Tetrodotoxin zu bilden, wurde allerdings wieder in Frage gestellt.[9][10]
Die Gattung ist klinisch besonders wichtig, weil die Mehrheit ihrer Mitglieder Resistenzen gegen Antibiotika aufweist. Außerdem sind sie fähig, bei höherer Zelldichte Biofilme (Schleime) zu bilden, die sie gegen Fresszellen und Antibiotika schützen. Während Bakterien der Gattung Pseudomonas bei Menschen mit intaktem Immunsystem selten Krankheiten verursachen, können sie bei Patienten, deren Immunsystem bereits geschwächt ist (beispielsweise in Krankenhäusern, sogenannte Hospitalismuskeime), die Infektion von Wunden, Atem- und Harnwegen, Lungenentzündung sowie Sepsis und Herzerkrankungen verursachen. Wundinfektionen durch P. aeruginosa zeichnen sich durch ihre blau-grüne Färbung und ihren süßlich-aromatischen Geruch aus. Besonders gefährdet sind Patienten mit der Erbkrankheit Cystische Fibrose (Mukoviszidose), bei denen Lungenentzündungen durch Pseudomonas die häufigste Todesursache sind.
Zur Therapie eignen sich Acylaminopenicilline wie Piperacillin, Cephalosporine ab der dritten Generation (insbesondere Ceftazidim und Cefepim; Ceftriaxon und Cefotaxim hingegen sind nicht wirksam), Fluorchinolone, Aminoglykoside sowie Carbapeneme.
Klinische Studien zu Pseudomonas-Impfungen werden seit einiger Zeit durchgeführt, etwa vom österreichischen Biotech-Unternehmen Intercell, das seinen Impfstoff bereits in Phase III testet.[11] Die referenzierte Studie kam nicht zum gewünschten Ergebnis.[12]
Pseudomonas (fem.: die Pseudomonas, Plural für die der Gattung angehörigen Organismen: Pseudomonaden, von altgriechisch ψεῦδος pseúdos, deutsch ‚Täuschung‘, μονάς monás, deutsch ‚Einheit‘) ist eine Gattung stäbchenförmiger, mit polaren Geißeln sich aktiv bewegender, immer anaerob oder aerob veratmend, nie fermentierender, gramnegativer Bakterien. Sporen werden nicht gebildet. Das Genom der Art P. aeruginosa wurde vollständig sequenziert.
Pseudomonas is a genus of Gram-negative bacteria belonging to the family Pseudomonadaceae in the class Gammaproteobacteria. The 313 members of the genus[2][3] demonstrate a great deal of metabolic diversity and consequently are able to colonize a wide range of niches.[4] Their ease of culture in vitro and availability of an increasing number of Pseudomonas strain genome sequences has made the genus an excellent focus for scientific research; the best studied species include P. aeruginosa in its role as an opportunistic human pathogen, the plant pathogen P. syringae, the soil bacterium P. putida, and the plant growth-promoting P. fluorescens, P. lini, P. migulae, and P. graminis.[5][6]
Because of their widespread occurrence in water and plant seeds such as dicots, the pseudomonads were observed early in the history of microbiology. The generic name Pseudomonas created for these organisms was defined in rather vague terms by Walter Migula in 1894 and 1900 as a genus of Gram-negative, rod-shaped, and polar-flagellated bacteria with some sporulating species.[7][8] The latter statement was later proved incorrect and was due to refractive granules of reserve materials.[9] Despite the vague description, the type species, Pseudomonas pyocyanea (basonym of Pseudomonas aeruginosa), proved the best descriptor.[9]
Like most bacterial genera, the pseudomonad[note 1] last common ancestor lived hundreds of millions of years ago. They were initially classified at the end of the 19th century when first identified by Walter Migula. The etymology of the name was not specified at the time and first appeared in the seventh edition of Bergey's Manual of Systematic Bacteriology (the main authority in bacterial nomenclature) as Greek pseudes (ψευδής) "false" and -monas (μονάς/μονάδος) "a single unit", which can mean false unit; however, Migula possibly intended it as false Monas, a nanoflagellated protist[9] (subsequently, the term "monad" was used in the early history of microbiology to denote unicellular organisms). Soon, other species matching Migula's somewhat vague original description were isolated from many natural niches and, at the time, many were assigned to the genus. However, many strains have since been reclassified, based on more recent methodology and use of approaches involving studies of conservative macromolecules.[10]
Recently, 16S rRNA sequence analysis has redefined the taxonomy of many bacterial species.[11] As a result, the genus Pseudomonas includes strains formerly classified in the genera Chryseomonas and Flavimonas.[12] Other strains previously classified in the genus Pseudomonas are now classified in the genera Burkholderia and Ralstonia.[13][14]
In 2020, a phylogenomic analysis of 494 complete Pseudomonas genomes identified two well-defined species (P. aeruginosa and P. chlororaphis) and four wider phylogenetic groups (P. fluorescens, P. stutzeri, P. syringae, P. putida) with a sufficient number of available proteomes.[15] The four wider evolutionary groups include more than one species, based on species definition by the Average Nucleotide Identity levels.[16] In addition, the phylogenomic analysis identified several strains that were mis-annotated to the wrong species or evolutionary group.[15] This mis-anotation problem has been reported by other analyses as well.[17]
In 2000, the complete genome sequence of a Pseudomonas species was determined; more recently, the sequence of other strains has been determined, including P. aeruginosa strains PAO1 (2000), P. putida KT2440 (2002), P. protegens Pf-5 (2005), P. syringae pathovar tomato DC3000 (2003), P. syringae pathovar syringae B728a (2005), P. syringae pathovar phaseolica 1448A (2005), P. fluorescens Pf0-1, and P. entomophila L48.[10]
By 2016, more than 400 strains of Pseudomonas had been sequenced.[18] Sequencing the genomes of hundreds of strains revealed highly divergent species within the genus. In fact, many genomes of Pseudomonas share only 50-60% of their genes, e.g. P. aeruginosa and P. putida share only 2971 proteins out of 5350 (or ~55%).[18]
By 2020, more than 500 complete Pseudomonas genomes were available in Genebank. A phylogenomic analysis utilized 494 complete proteomes and identified 297 core orthologues, shared by all strains.[15] This set of core orthologues at the genus level was enriched for proteins involved in metabolism, translation, and transcription and was utilized for generating a phylogenomic tree of the entire genus, to delineate the relationships among the Pseudomonas major evolutionary groups.[15] In addition, group-specific core proteins were identified for most evolutionary groups, meaning that they were present in all members of the specific group, but absent in other pseudomonads. For example, several P. aeruginosa-specific core proteins were identified that are known to play an important role in this species' pathogenicity, such as CntL, CntM, PlcB, Acp1, MucE, SrfA, Tse1, Tsi2, Tse3, and EsrC.[15]
Members of the genus display these defining characteristics:[19]
Other characteristics that tend to be associated with Pseudomonas species (with some exceptions) include secretion of pyoverdine, a fluorescent yellow-green siderophore[20] under iron-limiting conditions. Certain Pseudomonas species may also produce additional types of siderophore, such as pyocyanin by Pseudomonas aeruginosa[21] and thioquinolobactin by Pseudomonas fluorescens,.[22] Pseudomonas species also typically give a positive result to the oxidase test, the absence of gas formation from glucose, glucose is oxidised in oxidation/fermentation test using Hugh and Leifson O/F test, beta hemolytic (on blood agar), indole negative, methyl red negative, Voges–Proskauer test negative, and citrate positive.
Pseudomonas may be the most common nucleator of ice crystals in clouds, thereby being of utmost importance to the formation of snow and rain around the world.[23]
All species and strains of Pseudomonas have historically been classified as strict aerobes. Exceptions to this classification have recently been discovered in Pseudomonas biofilms.[24] A significant number of cells can produce exopolysaccharides associated with biofilm formation. Secretion of exopolysaccharides such as alginate makes it difficult for pseudomonads to be phagocytosed by mammalian white blood cells.[25] Exopolysaccharide production also contributes to surface-colonising biofilms that are difficult to remove from food preparation surfaces. Growth of pseudomonads on spoiling foods can generate a "fruity" odor.
Most Pseudomonas spp. are naturally resistant to penicillin and the majority of related beta-lactam antibiotics, but a number are sensitive to piperacillin, imipenem, ticarcillin, or ciprofloxacin.[25] Aminoglycosides such as tobramycin, gentamicin, and amikacin are other choices for therapy.
This ability to thrive in harsh conditions is a result of their hardy cell walls that contain porins. Their resistance to most antibiotics is attributed to efflux pumps, which pump out some antibiotics before they are able to act.
Pseudomonas aeruginosa is increasingly recognized as an emerging opportunistic pathogen of clinical relevance. One of its most worrying characteristics is its low antibiotic susceptibility.[26] This low susceptibility is attributable to a concerted action of multidrug efflux pumps with chromosomally encoded antibiotic resistance genes (e.g., mexAB-oprM, mexXY, etc.,[27]) and the low permeability of the bacterial cellular envelopes. Besides intrinsic resistance, P. aeruginosa easily develops acquired resistance either by mutation in chromosomally encoded genes or by the horizontal gene transfer of antibiotic resistance determinants. Development of multidrug resistance by P. aeruginosa isolates requires several different genetic events that include acquisition of different mutations and/or horizontal transfer of antibiotic resistance genes. Hypermutation favours the selection of mutation-driven antibiotic resistance in P. aeruginosa strains producing chronic infections, whereas the clustering of several different antibiotic resistance genes in integrons favours the concerted acquisition of antibiotic resistance determinants. Some recent studies have shown phenotypic resistance associated to biofilm formation or to the emergence of small-colony-variants, which may be important in the response of P. aeruginosa populations to antibiotic treatment.[10]
Although gallium has no natural function in biology, gallium ions interact with cellular processes in a manner similar to iron(III). When gallium ions are mistakenly taken up in place of iron(III) by bacteria such as Pseudomonas, the ions interfere with respiration, and the bacteria die. This happens because iron is redox-active, allowing the transfer of electrons during respiration, while gallium is redox-inactive.[28][29]
Infectious species include P. aeruginosa, P. oryzihabitans, and P. plecoglossicida. P. aeruginosa flourishes in hospital environments, and is a particular problem in this environment, since it is the second-most common infection in hospitalized patients (nosocomial infections).[30] This pathogenesis may in part be due to the proteins secreted by P. aeruginosa. The bacterium possesses a wide range of secretion systems, which export numerous proteins relevant to the pathogenesis of clinical strains.[31] Intriguingly, several genes involved in the pathogenesis of P.aeruginosa, such as CntL, CntM, PlcB, Acp1, MucE, SrfA, Tse1, Tsi2, Tse3, and EsrC are core group-specific,[15] meaning that they are shared by the vast majority of P. aeruginosa strains, but they are not present in other Pseudomonads.
P. syringae is a prolific plant pathogen. It exists as over 50 different pathovars, many of which demonstrate a high degree of host-plant specificity. Numerous other Pseudomonas species can act as plant pathogens, notably all of the other members of the P. syringae subgroup, but P. syringae is the most widespread and best-studied.
Although not strictly a plant pathogen, P. tolaasii can be a major agricultural problem, as it can cause bacterial blotch of cultivated mushrooms.[32] Similarly, P. agarici can cause drippy gill in cultivated mushrooms.[33]
Since the mid-1980s, certain members of the genus Pseudomonas have been applied to cereal seeds or applied directly to soils as a way of preventing the growth or establishment of crop pathogens. This practice is generically referred to as biocontrol. The biocontrol properties of P. fluorescens and P. protegens strains (CHA0 or Pf-5 for example) are currently best-understood, although it is not clear exactly how the plant growth-promoting properties of P. fluorescens are achieved. Theories include: the bacteria might induce systemic resistance in the host plant, so it can better resist attack by a true pathogen; the bacteria might outcompete other (pathogenic) soil microbes, e.g. by siderophores giving a competitive advantage at scavenging for iron; the bacteria might produce compounds antagonistic to other soil microbes, such as phenazine-type antibiotics or hydrogen cyanide. Experimental evidence supports all of these theories.[34]
Other notable Pseudomonas species with biocontrol properties include P. chlororaphis, which produces a phenazine-type antibiotic active agent against certain fungal plant pathogens,[35] and the closely related species P. aurantiaca, which produces di-2,4-diacetylfluoroglucylmethane, a compound antibiotically active against Gram-positive organisms.[36]
Some members of the genus are able to metabolise chemical pollutants in the environment, and as a result, can be used for bioremediation. Notable species demonstrated as suitable for use as bioremediation agents include:
One way of identifying and categorizing multiple bacterial organisms in a sample is to use ribotyping.[46] In ribotyping, differing lengths of chromosomal DNA are isolated from samples containing bacterial species, and digested into fragments.[46] Similar types of fragments from differing organisms are visualized and their lengths compared to each other by Southern blotting or by the much faster method of polymerase chain reaction (PCR).[46] Fragments can then be matched with sequences found on bacterial species.[46] Ribotyping is shown to be a method to isolate bacteria capable of spoilage.[47] Around 51% of Pseudomonas bacteria found in dairy processing plants are P. fluorescens, with 69% of these isolates possessing proteases, lipases, and lecithinases which contribute to degradation of milk components and subsequent spoilage.[47] Other Pseudomonas species can possess any one of the proteases, lipases, or lecithinases, or none at all.[47] Similar enzymatic activity is performed by Pseudomonas of the same ribotype, with each ribotype showing various degrees of milk spoilage and effects on flavour.[47] The number of bacteria affects the intensity of spoilage, with non-enzymatic Pseudomonas species contributing to spoilage in high number.[47]
Food spoilage is detrimental to the food industry due to production of volatile compounds from organisms metabolizing the various nutrients found in the food product.[48] Contamination results in health hazards from toxic compound production as well as unpleasant odours and flavours.[48] Electronic nose technology allows fast and continuous measurement of microbial food spoilage by sensing odours produced by these volatile compounds.[48] Electronic nose technology can thus be applied to detect traces of Pseudomonas milk spoilage and isolate the responsible Pseudomonas species.[49] The gas sensor consists of a nose portion made of 14 modifiable polymer sensors that can detect specific milk degradation products produced by microorganisms.[49] Sensor data is produced by changes in electric resistance of the 14 polymers when in contact with its target compound, while four sensor parameters can be adjusted to further specify the response.[49] The responses can then be pre-processed by a neural network which can then differentiate between milk spoilage microorganisms such as P. fluorescens and P. aureofaciens.[49]
Pseudomonas comprises the following species,[50] organized into genomic affinity groups:[51][52][53][54][55][56][57]
P. asplenii Subgroup
P. chlororaphis Subgroup
P. corrugata Subgroup
P. fluorescens Subgroup
P. fragi Subgroup
P. gessardii Subgroup
P. jessenii Subgroup
P. koreensis Subgroup
P. mandelii Subgroup
P. protegens Subgroup
incertae sedis
Recently, 16S rRNA sequence analysis redefined the taxonomy of many bacterial species previously classified as being in the genus Pseudomonas.[11] Species removed from Pseudomonas are listed below; clicking on a species will show its new classification. The term 'pseudomonad' does not apply strictly to just the genus Pseudomonas, and can be used to also include previous members such as the genera Burkholderia and Ralstonia.
α proteobacteria: P. abikonensis, P. aminovorans, P. azotocolligans, P. carboxydohydrogena, P. carboxidovorans, P. compransoris, P. diminuta, P. echinoides, P. extorquens, P. lindneri, P. mesophilica, P. paucimobilis, P. radiora, P. rhodos, P. riboflavina, P. rosea, P. vesicularis.
β proteobacteria: P. acidovorans, P. alliicola, P. antimicrobica, P. avenae, P. butanovora, P. caryophylli, P. cattleyae, P. cepacia, P. cocovenenans, P. delafieldii, P. facilis, P. flava, P. gladioli, P. glathei, P. glumae, P. huttiensis, P. indigofera, P. lanceolata, P. lemoignei, B. mallei, P. mephitica, P. mixta, P. palleronii, P. phenazinium, P. pickettii, P. plantarii, P. pseudoflava, B. pseudomallei, P. pyrrocinia, P. rubrilineans, P. rubrisubalbicans, P. saccharophila, P. solanacearum, P. spinosa, P. syzygii, P. taeniospiralis, P. terrigena, P. testosteroni.
γ-β proteobacteria: P. boreopolis, P. cissicola, P. geniculata, P. hibiscicola, P. maltophilia, P. pictorum.
γ proteobacteria: P. beijerinckii, P. diminuta, P. doudoroffii, P. elongata, P. flectens, P. marinus, P. halophila, P. iners, P. marina, P. nautica, P. nigrifaciens, P. pavonacea,[58] P. piscicida, P. stanieri.
δ proteobacteria: P. formicans.
The following relationships between genomic affinity groups have been determined by phylogenetic analysis:[57][59]
PseudomonasPseudomonas fluorescens group
Pseudomonas syringae group
Pseudomonas lutea group
Pseudomonas putida group
Pseudomonas rhizosphaerae group
Pseudomonas massiliensis group
Pseudomonas anguilliseptica group
Pseudomonas straminea group
Pseudomonas oleovorans group
Pseudomonas pohangensis group
Pseudomonas linyingensis group
Pseudomonas stutzeri group
Pseudomonas resinivorans group
Pseudomonas aeruginosa group
Pseudomonas oryzihabitans group
Halopseudomonas (formerly the P. pertucinogena group)
Cellvibrio japonicus (outgroup)
There are a number of bacteriophages that infect Pseudomonas, e.g.
Pseudomonas is a genus of Gram-negative bacteria belonging to the family Pseudomonadaceae in the class Gammaproteobacteria. The 313 members of the genus demonstrate a great deal of metabolic diversity and consequently are able to colonize a wide range of niches. Their ease of culture in vitro and availability of an increasing number of Pseudomonas strain genome sequences has made the genus an excellent focus for scientific research; the best studied species include P. aeruginosa in its role as an opportunistic human pathogen, the plant pathogen P. syringae, the soil bacterium P. putida, and the plant growth-promoting P. fluorescens, P. lini, P. migulae, and P. graminis.
Because of their widespread occurrence in water and plant seeds such as dicots, the pseudomonads were observed early in the history of microbiology. The generic name Pseudomonas created for these organisms was defined in rather vague terms by Walter Migula in 1894 and 1900 as a genus of Gram-negative, rod-shaped, and polar-flagellated bacteria with some sporulating species. The latter statement was later proved incorrect and was due to refractive granules of reserve materials. Despite the vague description, the type species, Pseudomonas pyocyanea (basonym of Pseudomonas aeruginosa), proved the best descriptor.
Pseudomonas es un género de bacilos rectos o ligeramente curvados, Gram negativos, oxidasa positivos, aeróbicos estrictos aunque en algunos casos pueden utilizar el nitrato como aceptor de electrones. El catabolismo de los glúcidos se realiza por la ruta de Etner-Doudoroff y el ciclo de los ácidos tricarboxílicos. Algunos miembros del género son psicrófilos, mientras que otros sintetizan sideróforos fluorescentes de color amarillo-verdoso con gran valor taxonómico. Es común la presencia de plásmidos y no forman esporas.
Con el reciente análisis de secuencias del RNAr 16S, se han definido la taxonomía de muchas especies bacterianas[1] y como resultado, el género Pseudomonas incluyen algunas cepas clasificadas anteriormente dentro de Chryseomonas y Flavimonas.[2] Otras cepas clasificadas previamente en el género Pseudomonas, ahora son agrupadas en los géneros Burkholderia y Ralstonia.
Pseudomonas literalmente significa «falsa unidad», derivado del griego pseudo (ψευδο 'falso') y monas (μονάς / μονάδα 'una sola unidad'). El término «monada» se usaba en la microbiología antigua para nombrar a los organismos unicelulares.
Debido a su amplia distribución en la naturaleza, las Pseudomonadaceae fueron observadas en los inicios históricos de la microbiología. El nombre genérico Pseudomonas creado para estos organismos estaba definida en términos relativamente vagos en 1894, como el género de bacteria gram negativa, bacilos con flagelo polar. Poco después, un gran número de especies eran asignadas al género. Las pseudomonadaceae eran aisladas de un variado número de nichos ecológicos de modo que un grandísimo número de especies recibían el nombre del género. Nuevas metodologías y la aparición de abordajes basados en los estudios de macromoléculas conservadas entre diversos organismos, han reclasificado a muchas especies.[3]
La Pseudomonas aeruginosa ha estado aumentando como un reconocido patógeno oportunista emergente en la relevancia clínica. Varios estudios epidemiológicos diferentes indican además que la resistencia a antibióticos ha venido incrementando entre muestras clínicas.
En el año 2000, se determinó el genoma completo de una especie de Pseudomonas y más recientemente se han determinado las secuencias de otras especies, incluyendo P. aeruginosa cepa PAO1 (2000), P. putida KT2440 (2002), P. fluorescens Pf-5 (2005), P. syringae pathovar tomato DC3000 (2003), P. syringae pathovar syringae B728a (2005), P. syringae pathovar phaseolica 1448A (2005), P. fluorescens PfO-1, P. entomophila L48.[3] gta y P. balearica DSM 6083T (2014).
Los miembros de este género generalmente son móviles gracias a uno o más flagelos polares que poseen, son catalasa positivos y no forman esporas.[4] Algunas especies sintetizan una cápsula de exopolisacáridos que facilita la adhesión celular, la formación de biopelículas y protege de la fagocitosis, de los anticuerpos o del complemento aumentando así su patogenicidad.
Otras características que tienden a ser asociadas con las especies de Pseudomonas -con algunas excepciones- incluye la secreción de pioverdina (fluorescein), un sideróforo fluorescente de color amarillo verdoso[5] bajo condiciones limitadas de hierro. Algunas especies pueden producir otros sideróforos, tales como la piocianina por la Pseudomonas aeruginosa[6] y tioquinolobactina por Pseudomonas fluorescens.[7] Las especies de Pseudomonas son típicamente oxidasa positivas, con ausencia de formación de gas a partir de glucosa, son hemolíticas (en agar sangre), prueba del indol negativas, rojo de metilo negativas y Voges Proskauer negativas.
El género demuestra una gran diversidad metabólica, y consecuentemente son capaces de colonizar un amplio rango de nichos.[8] Son de fácil cultivo in vitro y ampliamente disponibles en número, por lo que ciertas cepas son excelentes para investigaciones científicas, por ejemplo, P. aeruginosa y su rol como patógeno oportunista de humanos, el patógeno de plantas P. syringae, la bacteria de tierra P. putida y la P. fluorescens que promueve el crecimiento de plantas.
Los factores de virulencia de la estructura celular incluyen antígenos somáticos O[9] y flagelares H, fimbrias y cápsula de polisacáridos. Producen enzimas extracelulares como elastasas, proteasas y dos hemolisinas: fosfolipasa C termolábil y un lipopolisacárido termoestable.[10] La exotoxina A bloquea la síntesis de proteínas responsable de la necrosis tisular. En aves se refiere al uso contaminado de agua.
Las Pseudomonas crecen en medios simples. En caldo crecen abundantemente formando un anillo y un sedimento de color verde azulado. En agar simple forman colonias brillantes, confluentes, de borde continuo y a veces ondulado con un centro opaco. El pigmento (piocianina) se difunde en el medio dándole una tonalidad azul verdosa. Este pigmento tiene cualidades bactericidas sobre otras bacterias Gram positivas y Gram negativas.
Las especies del género Pseudomonas son organismos ubicuos, bacterias gram negativas que se encuadran dentro del grupo γ de las proteobacterias. Se han aislado bacterias de este género tanto en suelos limpios como en suelos contaminados por productos biogénicos y xenobióticos. También son microbiota predominante en la rizosfera y en la filosfera de plantas; del mismo modo, se han aislado de ambientes acuáticos, tanto de agua dulce como de aguas marinas. En general inocuas para el hombre también existen: patógenos oportunistas como P.aeruginosa; patógenos de animales y patógenos de plantas como P.syringae.
Este género es uno de los más proclives a la degradación de compuestos orgánicos, especialmente cepas de la especie Pseudomonas putida. El amplio potencial catabólico de los componentes del género viene dado en muchos casos por la presencia de determinantes plasmídicos y transposones autotransmisibles. La ubicuidad de las bacterias del género Pseudomonas y su capacidad para explotar una amplia variedad de nutrientes refleja un sistema de adaptación al medio ambiente que no encuentra parangón en las bacterias de otros géneros.
Las cepas del género Pseudomonas son capaces de procesar, integrar y reaccionar a una amplia variedad de condiciones cambiantes en el medio ambiente, y muestran una alta capacidad de reacción a señales físico-químicas y biológicas. Se han descrito cepas capaces de adquirir resistencia a metales pesados, disolventes orgánicos y detergentes, lo cual les permite explotar una amplia gama de fuentes de carbono como nutrientes, así como colonizar ambientes y nichos que difícilmente son colonizables por otros microorganismos. Por ello no es sorprendente que se considere a las bacterias del género Pseudomonas un paradigma de versatilidad metabólica, y microorganismos claves en el reciclado de materia orgánica en los compartimentos aeróbicos de los ecosistemas, jugando, por tanto, un papel esencial en la mejora y el mantenimiento de la calidad medioambiental. Además de su uso en biodegradación las especies del género Pseudomonas se emplean en distintos procesos industriales, tales como la fabricación de bioplásticos o en técnicas de biocontrol. La posición taxonómica de las distintas especies del género se encuentra sujeta a revisión.
P. aeruginosa es un patógeno oportunista humano, más comúnmente afecta a los inmunosuprimidos, tales como aquellos con fibrosis quística[11] o sida.[12] Estas infecciones pueden afectar a muchas partes del cuerpo, pero típicamente afectan las vías respiratorias, causando 50 % de las neumonías bacterianas nosocomiales.[9] El tratamiento de dichas infecciones puede ser difícil debido a la frecuente y repetitiva resistencia antibiótica.[13]
P. oryzihabitans puede también ser un patógeno humano, aunque las infecciones son raras. Puede causar peritonitis,[14] endoftalmitis,[15] septicemia y bacteriemia. Síntomas similares, aunque poco frecuentes, pueden ser vistos con P. luteola.[16]
P. plecoglossicida es una especie patógena de peces, causando ascitis hemorrágica en los peces Ayu (Plecoglossus altivelis).[17] P. anguilliseptica es también un patógeno en los peces.[18]
Debido a su actividad hemolítica, las especies que no son patógenas pueden ocasionalmente causar problemas clínicos, en particular en la infección de transfusiones de sangre.[19]
Las Pseudomonas, por ser bacterias hidrófilas, han estado involucradas en otitis externa en particular asociada al agua, como es el caso del oído de nadador crónico o aquellas provocadas por la inserción de objetos penetrantes en el oído.[20]
P. syringae es un patógeno prolífico de plantas. Existe en más de 50 variantes, muchas de las cuales muestran un alto grado de especificidad en las plantas. Hay numerosas especies de otras Pseudomonas que pueden actuar como patógenos de las plantas, aunque la P. syringae es la más distribuida y mejor estudiada por los fitopatologos.
Aunque no es un patógeno estrictamente de plantas, P. tolaasii puede ser un gran problema agrícola, debido a que causa manchas bacterianas de setas cultivadas.[21] Similarmente, P. agarici puede causar laminillas gotosas en ciertas setas.[22]
|fechaacceso= requiere |url= (ayuda) |fechaacceso= requiere |url= (ayuda) |fechaacceso= requiere |url= (ayuda) |fechaacceso= requiere |url= (ayuda) |fechaacceso= requiere |url= (ayuda) |fechaacceso= requiere |url= (ayuda) |fechaacceso= requiere |url= (ayuda) |fechaacceso= requiere |url= (ayuda) |fechaacceso= requiere |url= (ayuda) Pseudomonas es un género de bacilos rectos o ligeramente curvados, Gram negativos, oxidasa positivos, aeróbicos estrictos aunque en algunos casos pueden utilizar el nitrato como aceptor de electrones. El catabolismo de los glúcidos se realiza por la ruta de Etner-Doudoroff y el ciclo de los ácidos tricarboxílicos. Algunos miembros del género son psicrófilos, mientras que otros sintetizan sideróforos fluorescentes de color amarillo-verdoso con gran valor taxonómico. Es común la presencia de plásmidos y no forman esporas.
Con el reciente análisis de secuencias del RNAr 16S, se han definido la taxonomía de muchas especies bacterianas y como resultado, el género Pseudomonas incluyen algunas cepas clasificadas anteriormente dentro de Chryseomonas y Flavimonas. Otras cepas clasificadas previamente en el género Pseudomonas, ahora son agrupadas en los géneros Burkholderia y Ralstonia.
Pseudomonas generoa bakterio Gram negatiboz osatuta dago, bazilo itxura dutenak. Muturreko flageloen bidez mugitzen dira eta ez dute esporarik sortzen.
Genero honetako bakterioak nonahi bizi dira: uretan, lurrean, airean, landare zein animalietan, etab. Ekosistema ugaritan bizitzeko ahalmen hori substratu asko katabolizatzeko gaitasunarekin lotuta dago. Izan ere, Pseudomonas-ek 100 substratu baino gehiago erabil dezake elikatzeko, bere janari eskakizunak oso zorrotzak ez badira ere (ongi hazten da laborategiko hazkuntza-inguru arruntetan).
Genero honetako ezaugarri biokimiko nagusiak honako hauek dira:
Pseudomonas asko plasmidoen eramaileak dira.
Ohikoa da genero honetako espezie askotan pigmentuen ekoizpena. Pigmentu berdea (pioberdina) eta pigmentu urdina (piozianina) dira garrantzitsuenak. Bestalde, bakterio hauen espektro termikoa oso zabala da: 4º eta 42º C-en artean hazten dira.
Materia organikoa degradatzeko eta karbono-molekula ugari katabolizatzeko duen ahalmen handia dela eta, Pseudomonas-ek zeregin garrantzitsua burutzen du ekosistemen mantentzean, ingurugiroaren kalitatea hobetuz, oso kaltegarria izan zitekeen molekula organikoen pilaketa ekidinez.
Pseudomonas gehienak saprofito ez-kaltegarriak dira. Hala ere, gutxi batzuk giza zein landareen patogenoak izan daitezke.
Giza-patogenoen multzoan Pseudomonas aeruginosa azpimarratu behar da. Bakterio hau ez da patogeno hertsia, oportunista baizik. Horrek esan nahi du infekzioak sortzen dituela immunitate-sistema ahula dutenengan. Klinika eta ospitaleetan jarduten du batez ere mikrobio honek, bertan baitaude gaixo immunogutxitu gehienak. Lehen mailako arazoa da Pseudomonas aeruginosa-k ospitaleetako gaixoengan sortzen dituen infekzioak, bakterioak erresistentzia handia garatu baitu antibiotiko askorekiko.
Landare-patogenoen artean Pseudomonas syringae da aipatu beharrekoa.
Pseudomonas generoa bakterio Gram negatiboz osatuta dago, bazilo itxura dutenak. Muturreko flageloen bidez mugitzen dira eta ez dute esporarik sortzen.
Genero honetako bakterioak nonahi bizi dira: uretan, lurrean, airean, landare zein animalietan, etab. Ekosistema ugaritan bizitzeko ahalmen hori substratu asko katabolizatzeko gaitasunarekin lotuta dago. Izan ere, Pseudomonas-ek 100 substratu baino gehiago erabil dezake elikatzeko, bere janari eskakizunak oso zorrotzak ez badira ere (ongi hazten da laborategiko hazkuntza-inguru arruntetan).
Genero honetako ezaugarri biokimiko nagusiak honako hauek dira:
aerobioak (gehienak) edo anaerobio fakultatiboak. oxidasa (+) katalasa (+) indol (-) gelatinaren hidrolisia (+) gluzidoen katabolismoa Entner-Doudoroff (E-D) bide metabolikotik burutzen dute.Pseudomonas asko plasmidoen eramaileak dira.
Ohikoa da genero honetako espezie askotan pigmentuen ekoizpena. Pigmentu berdea (pioberdina) eta pigmentu urdina (piozianina) dira garrantzitsuenak. Bestalde, bakterio hauen espektro termikoa oso zabala da: 4º eta 42º C-en artean hazten dira.
Materia organikoa degradatzeko eta karbono-molekula ugari katabolizatzeko duen ahalmen handia dela eta, Pseudomonas-ek zeregin garrantzitsua burutzen du ekosistemen mantentzean, ingurugiroaren kalitatea hobetuz, oso kaltegarria izan zitekeen molekula organikoen pilaketa ekidinez.
Les bactéries du genre Pseudomonas peuvent être définies par :
Au début des années 2000, on considère que la famille des Pseudomonacées regroupe probablement la forme de vie la plus abondante et la plus répandue sur la planète[1]. Ce genre comprend en effet plus d'une centaine d'espèces ubiquitaires (dont l'espèce-type est Pseudomonas aeruginosa, généralement appelée Bacille pyocyanique). Cependant, ce nombre se réduit car les progrès de la phylogénétique permettent d'affiner le classement. On tend vers une soixantaine d'espèces.
Ces bactéries, largement répandues dans l'environnement, vivent dans le sol et les eaux douces, salées et saumâtres, superficielles et souterraines, et sont y compris retrouvées vivantes pour certaines souches dans les brumes, dans la pluies ou dans les nuages jusqu'à plus de 10 km d'altitude. Elles se retrouvent sur les plantes, dans les matières organiques non vivantes (denrées alimentaires), entraînant, parfois, leur altération organoleptique. L'Homme en a accru les habitats, dont via le transport aérien, maritime et fluvial et jusque dans le sous-sol via la fracturation hydraulique car on la retrouve aussi dans les boues de forages et fluides de fracturation[2].
Elles se rencontrent chez l'homme ou l'animal au niveau des fosses nasales et dans l'intestin. Elles constituent, pour la plupart, une flore commensale.
Certaines jouent un rôle pathogène :
Elles occupent des niches écologiques variées, mais se retrouvent plus particulièrement dans les milieux humides tels que les eaux douces, les eaux de mer, les eaux thermales. Elles se retrouvent en plus petite quantité dans les eaux riches en matières organiques (en particulier les eaux stagnantes) conséquence de la compétition entre microorganismes où les Pseudomonas ne sont pas les plus performants. Dans les habitations, elles se retrouvent ainsi dans les siphons d'évier, les eaux de réservoirs d'eaux de pluie qui, lorsqu'elles sont utilisées pour l'arrosage des plantes, peuvent provoquer des maladies sur les plantes en particulier les fleurs (pour les espèces phytopathogènes).
Le caractère très ubiquitaire des Pseudomonas est lié à des capacités adaptatives inhabituelles chez les bactéries, leur permettant de rapidement s'adapter à de très nombreux milieux et à des conditions de vies difficiles[3] ; Quelques souches de pseudomonas aeruginosa comptent parmi les microbes connus pour être les plus résistants aux antibiotiques
De nombreuses souches de Pseudomonas jouent un rôle majeur dans les processus de biodégradation. Dans les processus de remédiation et traitement de sites pollués, la biodégradation ou peut être favorisée ou accélérée par des apports en nutriments ou par des souches bactériennes sélectionnées. C'est le cas par exemple pour les pollutions du sol ou de l'eau par du fioul ou du pétrole brut. Dans ce cas un ensemencement par des souches mixtes de Pseudomonas et de Rhodococcus et se sont montrées plus efficaces pour dégrader le fioul en milieu aquatique. Dans ce dernier cas, on n'a pas réussi à améliorer les performances des bactéries en portant l'association à trois, quatre, ou cinq souches d'autres bactéries.
L'association champignon/bactérie est aussi très fréquente pour la dégradation de matières organiques[8], ou dans certains processus pathogènes (surinfection).
Chez l'Homme, l'espèce Pseudomonas aeruginosa (appelé aussi bacille pyocyanique) intervient fréquemment comme pathogène opportuniste. Elle se retrouve en flore de transit sur la peau et les muqueuses et cause des surinfections de plaies ou brûlures. Chez des individus immunodéprimés elle peut être la cause de diverses infections cutanées et viscérales voire de septicémie. Elle comporte un risque particulièrement élevé d'infections nosocomiales (contractées par l'intermédiaire de soins en milieu hospitalier), notamment avec des souches résistantes à certains antibiotiques courants.
Les autres germes sont peu agressifs par eux-mêmes mais se comportent en général comme germes « opportunistes » profitant de conditions anormales d'implantation chez des individus débilités, créant des flores de substitution ou des infections graves par introduction accidentelle dans l'organisme (cathétérismes, baxter, sondages, etc.). Leur rôle a acquis plus d'importance au cours des dernières années, surtout en milieu hospitalier.
Pseudomonas aeruginosas est sensible aux fluoroquinolones et ne présente généralement pas de difficulté à être traitée. Toutefois certaines souches deviennent antibiorésistantes. Malheureusement, du fait de leur caractère multirésistant et de l’usage massif d’antibiotiques, ces souches ont été artificiellement sélectionnées par l’homme et peuvent finir par prédominer sur les autres.
Le milieu hospitalier est l’endroit idéal pour cette sélection non désirée et Pseudomonas aeruginosa est responsable de nombreuses infections nosocomiales.
Aspect : les espèces du genre Pseudomonas sont des bacilles Gram négatifs fins droits et très mobiles grâce à un flagelle polaire : ciliature monotriche, dépourvus de spores et de capsules.
Groupement : elles apparaissent la plupart du temps isolés ou en diplobacilles.
Les espèces du genre Pseudomonas sont des germes peu exigeants, l'immense majorité des espèces n'exigent aucun facteur de croissance et sont capables d'utiliser comme seule source de carbone et d'énergie un large spectre de substrats carbonés simples et variés.
Elles se développent sur tous les milieux usuels, même les plus simples. Elles sont alcalinophiles.
Elles possèdent aussi une grande tolérance dans les gammes de température environnante, bien qu'ayant une température optimale de croissance située entre 30 et 35 °C (mésophile), elles sont capables de se développer à des températures entre 4 et 42 °C en ralentissant leur métabolisme. Leur croissance n'est donc pas totalement entravée par les locaux réfrigérés et les incubateurs.
Elles résistent à de nombreux inhibiteurs bactériens, particulièrement aux agents chimiques. Elles continuent leur développement dans certaines solutions antiseptiques (notamment le cétrimide) et sont résistantes à de nombreux antibiotiques.
-1= négatif ou absence, caractère absent chez 100%, ou présent chez 0%; 1= positif ou présence, caractère présent chez 100%, ou absent chez 0%; 0 ou espace libre = caractère variable présent chez 50%, ou absence d'information
Presque toutes les espèces possèdent une oxydase (voir le tableau).
La dégradation du glucose s'effectue par la voie d'Entner-Doudoroff, cette propriété est mise en évidence sur milieu Hugh-Leifson. Ils possèdent fréquemment une arginine dihydrolase permettant la dégradation de l'arginine en anaérobiose.
Certaines espèces peuvent extraire l'oxygène libéré par la réduction des nitrates en azote, et ainsi croître en anaérobiose ; on parle alors de « Respiration nitrate ».
Certaines espèces ou souches de pseudomonas produisent de l'acétone (toxique)[9].
De nombreux représentants de ce genre produisent des pigments. Citons ceux utilisés pour l'identification :
Liste des espèces qui devraient être exclues du genre Pseudomonas :
Les bactéries du genre Pseudomonas peuvent être définies par :
Bacilles à Gram négatif, oxydase + ; Aérobies stricts (« Respiration nitrate » chez certaines espèces) ; Dégradant le glucose par respiration aérobie ou inerte vis-à-vis du glucose. Ils n'attaquent pas les sucres ou les attaquent par voie oxydative et non fermentative ; Généralement mobiles par ciliature polaire (monotriche ou lophotriche) ; Peu exigeantes, cultivant de 0 à 30 °C ; Indole - ; Asporulés ; Colonies souvent pigmentées.Au début des années 2000, on considère que la famille des Pseudomonacées regroupe probablement la forme de vie la plus abondante et la plus répandue sur la planète. Ce genre comprend en effet plus d'une centaine d'espèces ubiquitaires (dont l'espèce-type est Pseudomonas aeruginosa, généralement appelée Bacille pyocyanique). Cependant, ce nombre se réduit car les progrès de la phylogénétique permettent d'affiner le classement. On tend vers une soixantaine d'espèces.
Pseudomonas é un xénero de bacterias aerobias gramnegativas, que contén 191 especies descritas validamente.[1]
A especie Pseudomonas aeruginosa considérase hoxe un patóxeno oportunista de importancia clínica. Varios estudos epidemiolóxicos diferentes indican que a resistencia a antibióticos está incrementándose nos illados clínicos desta especie.[2]
Os membros do xénero presentan unha gran diversidade de metabolismos, e en consecuencia poden colonizar unha gran cantidade de nichos.[3] A facilidade con que se cultivan in vitro e a dispoñibilidade dun número crecente de secuencias de xenomas de cepas de Pseudomonas fixo que moitas investigacións científicas se enfoquen cara a este xénero. Entre as especies mellor estudadas están P. aeruginosa polo seu papel como patóxeno humano oportunista, o patóxeno das plantas P. syringae, a bacteria do solo P. putida, e a bacteria promotora do crecemento das plantas P. fluorescens.
Debido á súa ampla distribución nos medios acuáticos e nas sementes de plantas como as dicotiledóneas, as pseudomónadas foron observadas xa nas primeiras épocas da historia da microbioloxía. O nome xenérico Pseudomonas creado para estes organismos foi definido dun modo bastante vago por Walter Migula en 1894 e 1900 como un xénero de bacterias gramnegativas, bacilares con flaxelos polares con algunhas especies esporulantes,[4][5] pero o último punto probouse que era incorrecto, xa que o que parecían esporas eran gránulos refractivos de materiais de reserva almacenados.[6] Malia esta vaga descrición, a especie tipo, Pseudomonas pyocyanea (basónimo de Pseudomonas aeruginosa), demostrou ser o seu mellor descritor.[6]
Un artigo publicado na revista Scientific American en 2008 sinalaba que Pseudomonas pode ser o nucleador máis común para a formación de cristais de xeo nas nubes, polo que ten unha grande importancia na formación da neve e chuvia en todo o mundo.[7]
O nome común que se utiliza para Pseudomonas é "pseudomonas" ou "pseudomónadas" (variante dalgúns dos casos non nominativos da declinación en grego de monas, monada[8]), aínda que co cambio sufrido nas clasificacións, pseudomónadas pode nalgúns textos referirse, ademais de a Pseudomonas, a outros xéneros da familia Pseudomonadaceae, que antes estaban incluídos no anterior. As pseudomónadas foron clasificadas inicialmente a finais do século XIX cando foron identificadas por Walter Migula. A etimoloxía do nome non foi especificada naquel tempo e apareceu por primeira vez na 7ª edición do Manual de bacterioloxía sistemática de Bergey (a principal autoridade en nomenclatura bacteriana) como procedente do grego pseudes (ψευδες) 'falso' e -monas (μονάς / μονάδα) 'unha unidade simple', que se pode traducir por falsa unidade; porén, tamén é posible que Migula pretendese denominalas "falsa Monas" (Monas é un xénero de protista nanoflaxelado[6]). Posteriormente, o termo "mónada" utilizouse nos inicios da microbioloxía para significar organismo dunha soa célula, e "monas" aparece como terminación de moitos nomes científicos. Axiña empezaron a atoparse outras especies que cadraban coa descrición xeral un pouco vaga dada por Migula en nichos moi diversos e, naquel tempo, moitas delas foron asignadas ao xénero Pseudomonas. Porén, moitas cepas foron despois reclasificadas utilizando metodoloxías máis modernas e os datos comparativos dos estudos de macromoléculas conservadas.[9]
Recentemente, as análises das secuencias do ARNr 16S obrigaron a redefinir a taxonomía de moitas especies bacterianas.[10] Como resultado, o xénero Pseudomonas inclúe cepas antes clasificadas no xénero Chryseomonas e Flavimonas.[11] Polo contrario, outras cepas antes clasificadas no xénero Pseudomonas están agora clasificadas no xénero Burkholderia e Ralstonia.[12][13]
No ano 2000, determinouse a secuencia xenómica completa dunha especie de Pseudomonas; máis recentemente, foron secuenciadas outras cepas, incluíndo P. aeruginosa cepa PAO1 (2000), P. putida KT2440 (2002), P. protegens Pf-5 (2005), P. syringae patovar tomato DC3000 (2003), P. syringae patovar syringae B728a (2005), P. syringae patovar phaseolica 1448A (2005), P. fluorescens PfO-1 e P. entomophila L48.[9]
Actualmente considérase que os membros do xénero mostran as seguintes características definitorias:[14]
Outras característricas que tenden a estar asociadas coas especies do xénero Pseudomonas (con algunhas excepcións) son a secreción de pioverdina, un sideróforo fluorescente amarelo verdoso[15] en condicións limitantes para o ferro. Certas especies de Pseudomonas poden tamén producir tipos adicionais de sideróforos, como a piocianina de Pseudomonas aeruginosa[16] ou a tioquinolobactina de Pseudomonas fluorescens.[17] As especies de Pseudomonas tamén dan tipicamente un resultado positivo na proba da oxidase, presentan ausencia de formación de gas a partir de glicosa, a glicosa oxídase na proba de oxidación/fermentación usando a proba de Hugh e Leifson O/F, son beta hemolíticos (en ágar sangue), dan negativo na proba do indol, son vermello de metilo negativas, negativas na proba de Voges–Proskauer, e positivas na proba do citrato.
Todas as especies e cepas de Pseudomonas son bacilos gramnegativos, e foron clasificados historicamente como aerobios estritos. Recentemente descubríronse excepcións a esta clasificación en biofilmes ou biopelículas de Pseudomonas.[18] Un número significativo de células poden producir exopolisacáridos que están asociados coa formación dos biofilmes. A secreción de exopolisacáridos como alxinatos, fai difícil que as pseudomónadas sexan fagocitadas polos glóbulos brancos dos mamíferos.[19] A produción de exopolisacáridos tamén contribúe á formación de biofilmes que colonizan superficies, que son difíciles de eliminar das superficies onde se preparan alimentos. O crecemento de pseudomónadas en alimentos deteriorados pode xerar un cheiro a froita.
As especies de Pseudomonas teñen a capacidade de metabolizar unha gran variedade de nutrientes. Isto, combinado coa capacidade de formar biofilmes, fai que sexan capaces de sobrevivir en diversos e inesperados ambientes. Por exemplo, encontráronse en áreas onde se preparan produtos farmacéuticos. Unha fonte simple de carbono, como os residuos de xabón ou os adhesivos dos revestimentos de tapas de recipientes son lugares axeitados para que estas bacterias prosperen. Outros sitios en principio improbables nos que se atopan son os antisépticos, como os compostos de amonio cuaternario, e auga mineral embotellada.
Como son bacterias gramnegativas, a maioría das especies de Pseudomonas son resistentes naturalmente á penicilina e a maioría dos antibióticos relacionados beta-lactámicos, pero varias son sensibles á piperacilina, imipenem, ticarcilina, tobramicina, ou ciprofloxacina.[19]
Esta capacidade de prosperar en condicións moi adversas é o resultado da súa robusta parede celular, que contén porinas. A súa resistencia á maioría dos antibióticos atribúese a que posúe bombas de fluxo, que bombean fóra da célula algúns antibióticos antes de que estes teñan tempo de actuar.
Pseudomonas aeruginosa é un patóxeno oportunista humano moi importante. Unha das características máis preocupantes de P. aeruginosa é a súa baixa susceptibilidade aos antibióticos. Esta baixa susceptibilidade é atribuíble a unha acción concertada de bombas de fluxo multidrogas con xenes para a resistencia a antibióticos codificados no cromosoma (por exemplo, mexAB-oprM, mexXY etc.,[20]) e a baixa permeabilidade das envolturas celulares da bacteria. Ademais da resistencia intrínseca, P. aeruginosa desenvolve facilmente unha resistencia adquirida. Para o desenvolvemento de multirresistencia por parte de illamentos bacterianos de P. aeruginosa cómpre que teñan lugar varios eventos xenéticos, que inclúen a adquisición de diferentes mutacións e/ou transferencia horizontal de xenes. A hipermutación favorece a selección de resistencia aos antibióticos impulsada polas mutacións en cepas de P. aeruginosa producindo infeccións crónicas, na que o agrupamento de varios xenes de resistencia a antibióticos en integróns favorece a adquisición concertada de determinantes de resistencia a antibióticos. Algúns estudos recentes mostraron que a resistencia fenotípica asociada á formación de biofilmes ou á emerxencia de variantes de pequenas colonias pode ser importante na resposta das poboacións de P. aeruginosa ao tratamento con antibióticos.[9]
Os estudos na taxonomía deste complicado xénero tiveron que procurar o seu camiño case a cegas mentres se dispoñía só dos procedementos clásicos para a descrición e identificación dos organismos que se usaban na bacterioloxía sanitaria durante as primeiras décadas do século XX. Esta situación cambiou drasticamente cando se empezaron a introducir como criterio central as similitudes na composición e secuencia de compoñentes macromoleculares do ARN ribosómico. A nova metodoloxía mostrou claramente que o xénero Pseudomonas, tal como se definía clasicamente, consistía de feito nun conglomerado de xéneros que podían ser separados claramente en cinco grandes grupos denominados grupos de homoloxía de ARNr. Ademais, os estudos taxonómicos suxeririon unha aproximación que podía ser útil en estudos taxonómicos de todos os outros grupos procariotas.
Unhas poucas décadas despois de que Migula propuxese o novo xénero Pseudomonas en 1894, a acumulación de nomes de especies asignadas a ese xénero chegou a acadar proporcións alarmantes. Actualmente, o número de especies na lista vixente reduciuse en máis do 90%. De feito, esta redución podería ser aínda máis grande se se considera que a lista actual contén moitos nomes novos, é dicir, neste proceso da lista orixinal sobreviviron relativamente poucos nomes. A nova metodoloxía e a inclusión de aproximacións baseadas en estudos de macromoléculas conservadas distintas do ARNr, constituíron unha estratexia efectiva que axudou a reducir a hipertrofia nomenclatural de Pseudomonas a un tamaño manexable.[9]
As especies infecciosas inclúen P. aeruginosa, P. oryzihabitans, e P. plecoglossicida. P. aeruginosa prospera nos ambientes hospitalarios, e chegou a ser un problema importante neses ambientes, xa que é a segunda infección máis común en pacientes hospitalizados (ver infección nosocomial). Esta patoxénese pode en parte deberse ás proteínas que segrega P. aeruginosa. A bacteria posúe un amplo rango de sistemas de secreción, que exportan numerosas proteínas relevantes para a patoxénese das cepas clínicas.[21]
P. syringae é un patóxeno de plantas prolífico. Del existen 50 patovares (cepas con patoxenidade distintiva), moitas das cales demostran un alto grao de especificidade de planta hóspede. Hai outras moitas especies de Pseudomonas que poden actuar como patóxenos de plantas, especialmente todos os outros membros do subgrupo P. syringae, pero P. syringae é o máis estendido e mellor estudado.
Aínda que non é estritamente un patóxeno de plantas, P. tolaasii pode ser un problema importante para a agricultura, xa que pode causar manchas bcterianas en cogomelos cultivados.[22] De xeito similar, P. agarici tamén pode causar enfermidades en cogomelos cultivados.[23]
Desde mediados da década de 1980, certos membros do xénero Pseudomonas foron aplicados ás sementes de cereais ou aplicados directamente aos solos como unha forma de previr o crecemento ou establecemento de organismos patóxenos para a agricultura. Esta práctica denomínase xenericamente biocontrol. As propiedades para o biocontrol que mostran P. fluorescens e P. protegens cepas (CHA0 ou Pf-5 por exemplo) compréndense agora mellor, aínda que non se sabe claramente como se orixinan as propiedades que promoven o crecemento da planta de P. fluorescens. As teorías formuladas son: (1) a bacteria podería inducir a resistencia sistémica na planta hóspede, polo que esta pode resistir mellor o ataque de patóxenos; (2) a bacteria podería competir con outros microbios do solo (patóxenos), por exemplo cos seus compoñentes sideróforos, que lle darían unha vantaxe competitiva para captar ferro; (3) a bacteria podería producir compostos antagonistas doutros microbios do solo, como os antibióticos do tipo da fenazina ou cianuro de hidróxeno. Cada unha destas teorías ten evidencias experimentais que a apoian.[24]
Outras especies notables de Pseudomonas con propiedades axeitadas para o biocontrol inclúen P. chlororaphis, que produce un antibiótico de tipo fenazina, que é axente activo contra certos fungos patóxenos de plantas,[25] e a especie estreitamente emparentada P. aurantiaca, que produce di-2,4-diacetilfluoroglucilmetano, un composto activo como antibiótico contra organismos grampositivos.[26]
Algúns membros do xénero Pseudomonas poden metabolizar contaminantes químicos vertidos no medio ambiente, polo que poden usarse para a biorremediación. As especies máis importantes que demostraron ser axeitadas para o seu uso en biorremediación son as seguintes:
Como resultado da súa diversidade metabólica, capacidade de crecer a baixas temperaturas e natureza ubicua, moitas especies de Pseudomonas poden estragar alimentos. Notables exemplos son a deterioración dos produtos lácteos por P. fragi,[36] a dos ovos por P. taetrolens e P. mudicolens,[37] e a producida por P. lundensis, que afecta ao leite, queixo, carne, e peixe.[38]
Recentemente, as análises das secuencias de ARNr 16S redefiniron a taxonomía de moitas especies bacterianas que antes eran clasificadas no xénero Pseudomonas.[10] As especies que foron movidas de Pseudomonas para outros xéneros están listadas debaixo; poñendo o rato sobre cada nome aparece a nova clasificación que lles foi asignada. Como xa se dixo máis arriba, téñase en conta que con frecuiencia o termo común "pseudomónada" non se aplica estritamente só ao actual xénero Pseudomonas, e pode usarse incluíndo a antigos membros dese xénero, hoxe en xéneros como Burkholderia e Ralstonia.
α proteobacteria: P. abikonensis, P. aminovorans, P. azotocolligans, P. carboxydohydrogena, P. carboxidovorans, P. compransoris, P. diminuta, P. echinoides, P. extorquens, P. lindneri, P. mesophilica, P. paucimobilis, P. radiora, P. rhodos, P. riboflavina, P. rosea, P. vesicularis.
β proteobacteria: P. acidovorans, P. alliicola, P. antimicrobica, P. avenae, P. butanovorae, P. caryophylli, P. cattleyae, P. cepacia, P. cocovenenans, P. delafieldii, P. facilis, P. flava, P. gladioli, P. glathei, P. glumae, P. graminis, P. huttiensis, P. indigofera, P. lanceolata, P. lemoignei, P. mallei, P. mephitica, P. mixta, P. palleronii, P. phenazinium, P. pickettii, P. plantarii, P. pseudoflava, P. pseudomallei, P. pyrrocinia, P. rubrilineans, P. rubrisubalbicans, P. saccharophila, P. solanacearum, P. spinosa, P. syzygii, P. taeniospiralis, P. terrigena, P. testosteroni.
γ-β proteobacteria: P. beteli, P. boreopolis, P. cissicola, P. geniculata, P. hibiscicola, P. maltophilia, P. pictorum.
γ proteobacteria: P. beijerinckii, P. diminuta, P. doudoroffii, P. elongata, P. flectens, P. halodurans, P. halophila, P. iners, P. marina, P. nautica, P. nigrifaciens, P. pavonacea, P. piscicida, P. stanieri.
δ proteobacteria: P. formicans.
Coñécense varios virus bacteriófagos que infectan ao xénero Pseudomonas, como por exemplo:
Debido á resistencia a antibióticos que mostran algunhas especies como P. aeruginosa ensaiouse a terapia de fagos. A terapia de fagos é efectiva no tratamento de infeccións por P. aeruginosa en modelos de ratos.[42][43]
Pseudomonas é un xénero de bacterias aerobias gramnegativas, que contén 191 especies descritas validamente.
A especie Pseudomonas aeruginosa considérase hoxe un patóxeno oportunista de importancia clínica. Varios estudos epidemiolóxicos diferentes indican que a resistencia a antibióticos está incrementándose nos illados clínicos desta especie.
Os membros do xénero presentan unha gran diversidade de metabolismos, e en consecuencia poden colonizar unha gran cantidade de nichos. A facilidade con que se cultivan in vitro e a dispoñibilidade dun número crecente de secuencias de xenomas de cepas de Pseudomonas fixo que moitas investigacións científicas se enfoquen cara a este xénero. Entre as especies mellor estudadas están P. aeruginosa polo seu papel como patóxeno humano oportunista, o patóxeno das plantas P. syringae, a bacteria do solo P. putida, e a bacteria promotora do crecemento das plantas P. fluorescens.
Debido á súa ampla distribución nos medios acuáticos e nas sementes de plantas como as dicotiledóneas, as pseudomónadas foron observadas xa nas primeiras épocas da historia da microbioloxía. O nome xenérico Pseudomonas creado para estes organismos foi definido dun modo bastante vago por Walter Migula en 1894 e 1900 como un xénero de bacterias gramnegativas, bacilares con flaxelos polares con algunhas especies esporulantes, pero o último punto probouse que era incorrecto, xa que o que parecían esporas eran gránulos refractivos de materiais de reserva almacenados. Malia esta vaga descrición, a especie tipo, Pseudomonas pyocyanea (basónimo de Pseudomonas aeruginosa), demostrou ser o seu mellor descritor.
Un artigo publicado na revista Scientific American en 2008 sinalaba que Pseudomonas pode ser o nucleador máis común para a formación de cristais de xeo nas nubes, polo que ten unha grande importancia na formación da neve e chuvia en todo o mundo.
Pseudomonas, gram-negativni pokretni bacil, oportunistički je patogen koji često uzrokuje hospitalne infekcije.
Pseudomonas je ubikvitaran, a preferira vlažan okoliš. Kod ljudi je najčešća vrsta P. aeruginosa. Druge vrste, koje ponekad uzrokuju infekcije kod ljudi su P. paucimobilis, P. putida, P. fluorescens i P. acidovorans. P. aeruginosa se povremeno može naći na normalnoj koži u aksilarnom i anogenitalnom području, ali rijetko u stolici, osim ako prije toga nisu primjenjivani antibiotici. Mikroorganizam je često kontaminant lezija naseljenih virulentnijim mikroorganizmima, no povremeno uzrokuje i infekciju tkiva koja su izložena vanjskoj okolini. Infekcije pseudomonasom obično se pojavljuju u bolnicama, gdje se mikroorganizam često nalazi u umivaonicima, antiseptičkim otopinama i vrećicama za urin. Transmisija na bolesnike moguća je i preko zdravstvenog osoblja, osobito na odjelima za opekline ili u neonatalnim jedinicama intenzivne skrbi. Druge vrste, kao Burkholderia cepacia i Stenotrophomonas maltophilia, koje su ranije klasificirane u rod Pseudomonas, važni su nozokomijalni patogeni.
Većina infekcija koje uzrokuje P. aeruginosa pojavljuje se u hospitaliziranih bolesnika, koji su kondicionirani ili imunokompromitirani. P. aeruginosa je drugi po učestalosti uzročnik infekcija u jedinicama intenzivne skrbi i čest je uzročnik pneumonija u bolesnika na respiratoru. Osim nozokomijalnih infekcija, bolesnici s HIV-infekcijom imaju povaćan rizik od izvanbolničkih infekcija uzrokovanih P. aeruginosom, a u trenutku infekcije često imaju znake uznapredovale HIV-bolesti.
Infekcije Pseudomonasom mogu zahvatiti različite anatomske lokalizacije – kožu, potkožno tkivo, kosti, uho, oko, mokraćne puteve i srčane zaliske. Lokalizacija ovisi o ulaznom mjestu i osjetljivosti bolesnika. Kod opekotina, na području ispod nekroze može se naseliti vrlo velik broj mikroorganizama, čineći tako ishodište posljedične bakterijemije – često smrtonosne komplikacije opekotina. Bakterijemija koja nema porijeklo u mokraćnim putevima, a osobito ako ju ne uzrokuje P. aeruginosa, već druge vrste iz roda Pseudomonas, ukazuje na kontaminirane infuzijske otopine, lijekove ili antiseptike koji se koriste pri postavljanju IV katetera, kao izvor infekcije. Kod bolesnika s HIV-infekcijom, Pseudomonas najčešće uzrokuje pneumoniju ili sinusitis.
Klinička prezentacija ovisi o lokalizaciji infekcije. U hospitaliziranih bolesnika, plućna infekcija je moguća u vezi s endotrahealnom intubacijom, traeostomom ili mehaničkom ventilacijom, kad se Pseudomonas pridružuje ostalim gram-negativnim bacilima koji koloniziraju orofarinks. Bronhitis uzrokovan Pseudomonasom čest je u kasnijem tijeku cistične fibroze; ovi izolati imaju karakterističnu mukoidnu morfologiju kolonija.
Pseudomonas se često izolira iz hemokultura bolesnika s opeklinama ili malignom bolešću. Bolest se prezentira kliničkom slikom gram-negativne sepse, uz koju je često prisutna ecthyma gangraenosum, kožna lezija karakterizirana ljubičasto-crnim područjem promjera oko 1 cm s ulceriranim središtem okruženim crvenilom, a najčešće se nalazi u aksilarnom ili anogenitalnom području.
Pseudomonas je čest uzročnik infekcija mokraćnih puteva, osobito u bolesnika koji su podnijeli urološke zahvate, koji imaju opstruktivnu uropatiju ili koji su primali antibiotike širokog spektra. Otitis externa s purulentnom sekrecijom, čest u tropskim područjima, najčešći je oblik infekcije uha uzrokovane Pseudomonasom. Teži oblik bolesti, poznat kao maligni eksterni otitis, moguć je u dijabetičara; manifestira se jakim bolom u uhu, često s jednostranim paralizama kranijalnih živaca i zahtijeva parenteralnu terapiju. Zavaćanje oka se uglavnom očituje kao ulceracija rožnice, najčešće nakon traume, no u nekim slučajevima okrivljuje se kontaminacija kontaktnih leća ili tekućine za čuvanje leća. Mikroorganizam se može naći u sekretu fistula, osobito nakon traume ili nakon dubokih ubodnih rana stopala. Sekret često ima slatkasti miris koji podsjeća na miris voća. P. aeruginosa može, kao posljedicu infekcije ovih ubodnih rana, izazvati celulitis i osteomijelitis, koji onda pored antibiotika zahtijevaju i rani kirurški debridman. Pseudomonas rijetko uzrokuje endokarditis, obično kod intravenskih narkomana ili na umjetnim valvulama bolesnika koji su imali operaciju na otvorenom srcu. Desnostrani se endokarditis može liječiti konzervativno, no obično inficirana valvula mora biti odstranjena, kako bi se izliječila infekcija koja zahvaća mitralnu, aortalnu ili umjetnu valvulu.
Kad se radi o lokaliziranoj i vanjskoj infekciji, učinkovito je liječenje ispiranjem s 1% octenom kiselinom ili lokalnom primjenom polimiksina B ili kolistina. Nekrotično se tkivo mora odstraniti, a apscese drenirati. U slučajevima kad je potrebna parenteralna terapija, tobramicin ili gentamicin 5 mg/kg/dan u podijeljenim dozama, djelotvorni su kod većine infekcija mokraćnih puteva uzrokovanih Pseudomonasom. Ako dođe do kliničkog poboljšanja, doza se može smanjiti na 3 mg/kg/dan kako bi se smanjila mogućnost neželjenih učinaka lijeka. Doza se mora smanjiti kod renalne insuficijencije. Protiv Pseudomonasa s enzimima posredovanom rezistencijom na tobramicin i gentamicin, treba primijeniti amikacin. Mnogi stručnjaci za liječenje teških infekcija Pseudomonasom, preporučaju kombinaciju aminoglikozida s antipseudomonalnim β-laktamskim antibiotikom. Mnogi su penicilinski antibiotici aktivni protiv Pseudomonasa, među njima tikarcilin, piperacilin, mezlocilin i azlocilin. Drugi lijekovi s odličnom aktivnošću protiv Pseudomonasa su ceftazidim, cefepim, aztreonam, imipenem, meropenem i ciprofloksacin. Tikarcilin se najčešće upotrebljava u dozama od 16 do 20 gr/dan IV. Piperacilin, azlocilin, cefepim, ceftazidim, meropenem i imipenem, djelotvorni su in vitro protiv nekih sojeva rezistentnih na tikarcilin.
Kod sistemskih infekcija ili kod granulocitopeničnih bolesnika, treba kombinirati aminoglikozid aktivan protiv Pseudomonasa s antipseudomonalnim penicilinom. Kod neutropeničnih bolesnika s oštećenom funkcijom bubrega, zadovoljavajuće su i neaminoglikozidne kombinacije koje se sastoje od dva β-laktamska lijeka ili od β-laktama s fluorokinolonom. Infekcije mokraćnih puteva često se mogu liječiti peroralnim indanil karbenicilinom, odnosno ciprofloksacinom ili drugim fluorokinolonima. Fluorokinolone ne treba upotrebljavati kod djece, zbog mogućeg štetnog djelovanja na hrskavicu. Kod upotrebe dva antipseudomonalna lijeka, smanjena je mogućnost nastanka rezistentnih sojeva za vrijeme terapije.
Pseudomonas, gram-negativni pokretni bacil, oportunistički je patogen koji često uzrokuje hospitalne infekcije.
Pseudomonas Sp merupakan bakteri hidrokarbonoklastik yang mampu mendegradasi berbagai jenis hidrokarbon. Keberhasilan penggunaan bakteri Pseudomonas dalam upaya bioremediasi lingkungan akibat pencemaran hidrokarbon membutuhkan pemahaman tentang mekanisme interaksi antara bakteri Pseudomonas sp dengan senyawa hidrokarbon.
Kemampuan bakteri Pseudomonas sp. IA7D dalam mendegradasi hidrokarbon dan dalam menghasilkan biosurfaktan menunjukkan bahwa isolat bakteri Pseudomonas sp IA7D berpotensi untuk digunakan dalam upaya bioremediasi lingkungan akibat pencemaran hidrokarbon.
α proteobacteria: P. abikonensis, P. aminovorans, P. azotocolligans, P. carboxydohydrogena, P. carboxidovorans, P. compransoris, P. diminuta, P. echinoides, P. extorquens, P. lindneri, P. mesophilica, P. paucimobilis, P. radiora, P. rhodos, P. riboflavina, P. rosea, P. vesicularis.
β proteobacteria: P. acidovorans, P. alliicola, P. antimicrobica, P. avenae, P. butanovorae, P. caryophylli, P. cattleyae, P. cepacia, P. cocovenenans, P. delafieldii, P. facilis, P. flava, P. gladioli, P. glathei, P. glumae, P. graminis, P. huttiensis, P. indigofera, P. lanceolata, P. lemoignei, P. mallei, P. mephitica, P. mixta, P. palleronii, P. phenazinium, P. pickettii, P. plantarii, P. pseudoflava, P. pseudomallei, P. pyrrocinia, P. rubrilineans, P. rubrisubalbicans, P. saccharophila, P. solanacearum, P. spinosa, P. syzygii, P. taeniospiralis, P. terrigena, P. testosteroni.
γ-β proteobacteria: P. beteli, P. boreopolis, P. cissicola, P. geniculata, P. hibiscicola, P. maltophilia, P. pictorum.
γ proteobacteria: P. beijerinckii, P. diminuta, P. doudoroffii, P. elongata, P. flectens, P. halodurans, P. halophila, P. iners, P. marina, P. nautica, P. nigrifaciens, P. pavonacea, P. piscicida, P. stanieri.
δ proteobacteria: P. formicans...
Pseudomonas Sp merupakan bakteri hidrokarbonoklastik yang mampu mendegradasi berbagai jenis hidrokarbon. Keberhasilan penggunaan bakteri Pseudomonas dalam upaya bioremediasi lingkungan akibat pencemaran hidrokarbon membutuhkan pemahaman tentang mekanisme interaksi antara bakteri Pseudomonas sp dengan senyawa hidrokarbon.
Kemampuan bakteri Pseudomonas sp. IA7D dalam mendegradasi hidrokarbon dan dalam menghasilkan biosurfaktan menunjukkan bahwa isolat bakteri Pseudomonas sp IA7D berpotensi untuk digunakan dalam upaya bioremediasi lingkungan akibat pencemaran hidrokarbon.
Pseudomonas è un genere di batteri appartenenti alla famiglia delle Pseudomonadaceae. Il nome significa falsa unità (dal greco: ψευδο, psèudos, "falso" e μονος, monas, "singola unità"); deriva dalla abitudine di questi batteri a disporsi a coppie di cellule che osservate al microscopio sembrano una sola cellula. Alcuni microbiologi lo battezzarono così all'inizio del XX secolo.
Sono bacilli Gram-negativi, aerobi obbligati (in realtà possono crescere in anaerobiosi utilizzando nitrati come accettori di elettroni alternativi) , ossidasi positivi, catalasi positivi ed hanno flagelli polari che permettono loro di muoversi. Si trovano nel terreno e nelle acque ma anche sulle piante. Producono pigmenti come la piocianina che permette di alterare la funzione ciliare, stimolare la risposta infiammatoria e provocare danno ai tessuti. Alcuni ceppi sono capsulati.
Il principale fattore di virulenza è la esotossina A oltre alla endotossina (lipide A del lipopolisaccaride). Hanno inoltre un exopolisaccaride mucoide che permette una maggiore adesione alle cellule dell'ospite, formare biopellicole, inibire la fagocitosi e ridurre l'attività antibiotica. Sono batteri che colonizzano l'essere umano in zone umide come le piscine.
La specie Pseudomonas aeruginosa è una delle più diffuse tanto da essere presente in molti campioni di feci umane e più raramente anche in zone di epidermide più umide come ascelle ed inguine. È causa, in ospedali dove non sono seguite correttamente le norme di igiene, di vere e proprie piccole epidemie con conseguenze a volte anche gravi.
Generalmente provoca infezioni osteoarticolari, otite esterna, polmonite. È anche responsabile di follicoliti cutanee, infezioni oculari come congiuntivite ed endocardite. La diagnosi microbiologica si effettua isolando il germe in terreni di coltura come Agar Cetrimide e Agar MacConkey.
Generalmente le varie specie di Pseudomonas sono resistenti alla maggior parte degli antibiotici, in quanto sono scarsamente permeabili, producono enzimi capaci di inattivare penicilline ed aminoglicosidi e sono dotate di meccanismi di espulsione di molti antibiotici. È consigliato usare antibiotici in combinazione come Ceftazidima o aminoglucosidi. Le differenze dell'RNA ribosomale hanno permesso di suddividere tale genere in cinque differenti gruppi, dall'RNA I all'RNA V a loro volta suddivisi in sottogruppi.
Pseudomonas è un genere di batteri appartenenti alla famiglia delle Pseudomonadaceae. Il nome significa falsa unità (dal greco: ψευδο, psèudos, "falso" e μονος, monas, "singola unità"); deriva dalla abitudine di questi batteri a disporsi a coppie di cellule che osservate al microscopio sembrano una sola cellula. Alcuni microbiologi lo battezzarono così all'inizio del XX secolo.
Pseudomonas is een geslacht van bacteriën, waartoe soorten behoren, die planten- en dierziekten kunnen veroorzaken. Zo veroorzaakt Pseudomonas syringae sterfte bij kastanjebomen.
Het zijn staafvormige, aerobe, gramnegatieve bacteriën met polaire flagellen, waarmee ze zich kunnen voortbewegen. Er worden geen sporen gevormd. Het genoom van Pseudomonas aeruginosa is volledig in kaart gebracht met DNA-sequentie.[1] Pseudomonas aeruginosa is een schadelijke ziekenhuisbacterie.
De grootte varieert van 0,5–1,0 × 1,5–5,0 µm. Ze reageren in de Voges-Proskauer-test negatief, in de katalase-test positief. Bij de meeste soorten is de oxidasetest ook positief, zoals bij bijvoorbeeld Pseudomonas fluorescens, deze soorten hebben tevens een cytochroom c.
Pseudomonas-bacteriën zijn aeroob, dat wil zeggen dat ze bij de energiestofwisseling voor de groei zuurstof nodig hebben, waarbij de zuurstof als elektronenacceptor wordt gebruikt. Suikers worden hierbij langs de Entner-Doudoroff-pathway voor de energiewinning afgebroken. Nitraat kan bij de meeste soorten als alternatieve elektronenacceptor gebruikt worden door middel van denitrificatie. Hierbij wordt nitraat (NO3−) gereduceerd tot moleculaire stikstof (N2). Deze soorten kunnen dus ook zonder zuurstof in leven blijven. Voorbeelden hiervan zijn Pseudomonas aeruginosa, Pseudomonas stutzeri en Pseudomonas denitrificans. Ze worden gebruikt bij de nitraatverwijdering uit afval. Pseudomonas fluorescens daarentegen kan geen nitraat omzetten.
Meerdere Pseudomonas-soorten vormen bij ijzergebrek sideroforen; ten minste 50 verschillende pyoverdinen (fluorescente sideroforen) zijn gevonden[2].
Pseudomonas is een geslacht van bacteriën, waartoe soorten behoren, die planten- en dierziekten kunnen veroorzaken. Zo veroorzaakt Pseudomonas syringae sterfte bij kastanjebomen.
Het zijn staafvormige, aerobe, gramnegatieve bacteriën met polaire flagellen, waarmee ze zich kunnen voortbewegen. Er worden geen sporen gevormd. Het genoom van Pseudomonas aeruginosa is volledig in kaart gebracht met DNA-sequentie. Pseudomonas aeruginosa is een schadelijke ziekenhuisbacterie.

Pseudomonas – rodzaj gramujemnych, biegunowo urzęsionych bakterii przyjmujących kształt prostych lub nieznacznie wygiętych pałeczek[1][2].
Oddychają tlenowo, używając tlenu jako ostatecznego akceptora elektronów; niektóre również oddychają beztlenowo, przeprowadzając denitryfikację. Są katalazododatnie, chemoorganotroficzne, nie tworzą endospor. Potrafią rozkładać wiele węglowodorów, związków aromatycznych i ich pochodnych, które stanowią zanieczyszczenie środowiska[1][2], przez co mogą znajdować zastosowanie w bioremediacji. Większość heksoz rozkładają w szlaku Entnera-Doudoroffa[2].
Są szeroko rozpowszechnione w naturze[2]. Występują w glebie, wodzie, ściekach i w powietrzu. Zwykle zasiedlają nowe miejsca jako pierwsze, jeśli zawiera ono sole mineralne i cukry lub kwasy organiczne[1]. Niektóre biorą udział w psuciu żywności[3]. Mogą wytwarzać charakterystyczne barwniki rozpuszczalne w wodzie, np. błękitnozieloną piocyjaninę oraz żółtozielone fluoryzujące pigmenty, które mogą pełnić rolę sideroforów[1]. Należy do nich piowerdyna produkowana m.in. przez Pseudomonas fluorescens[4]
Niektóre mogą wykazywać patogenność wobec ludzi (Pseudomonas aeruginosa), zwierząt i roślin (Pseudomonas syringae). Bakterie z rodzaju Pseudomonas są częstymi patogenami oportunistycznymi. Są oporne na wiele antybiotyków i środków przeciwbakteryjnych[2].
Pseudomonas – rodzaj gramujemnych, biegunowo urzęsionych bakterii przyjmujących kształt prostych lub nieznacznie wygiętych pałeczek.
Oddychają tlenowo, używając tlenu jako ostatecznego akceptora elektronów; niektóre również oddychają beztlenowo, przeprowadzając denitryfikację. Są katalazododatnie, chemoorganotroficzne, nie tworzą endospor. Potrafią rozkładać wiele węglowodorów, związków aromatycznych i ich pochodnych, które stanowią zanieczyszczenie środowiska, przez co mogą znajdować zastosowanie w bioremediacji. Większość heksoz rozkładają w szlaku Entnera-Doudoroffa.
Są szeroko rozpowszechnione w naturze. Występują w glebie, wodzie, ściekach i w powietrzu. Zwykle zasiedlają nowe miejsca jako pierwsze, jeśli zawiera ono sole mineralne i cukry lub kwasy organiczne. Niektóre biorą udział w psuciu żywności. Mogą wytwarzać charakterystyczne barwniki rozpuszczalne w wodzie, np. błękitnozieloną piocyjaninę oraz żółtozielone fluoryzujące pigmenty, które mogą pełnić rolę sideroforów. Należy do nich piowerdyna produkowana m.in. przez Pseudomonas fluorescens
Niektóre mogą wykazywać patogenność wobec ludzi (Pseudomonas aeruginosa), zwierząt i roślin (Pseudomonas syringae). Bakterie z rodzaju Pseudomonas są częstymi patogenami oportunistycznymi. Są oporne na wiele antybiotyków i środków przeciwbakteryjnych.
Pseudomonas é um gênero de bactérias gram-negativas, bacilares, ubíquas (amplamente encontradas em diversos ambientes) da família Pseudomonadaceae. A espécie mais patógena é a Pseudomonas aeruginosa que causa infecções respiratórias, urinárias ou bacteriemia oportunistas em pacientes hospitalizados, com cateteres ou imunodeprimidos, otite em crianças e conjuntivite em quem usa lentes. Possuem ampla resistência, exigindo cultivo e teste de sensibilidade a antibióticos para decidir o melhor tratamento.[1]
Pessoas com diabetes, queimaduras e fibrose cística são particularmente mais vulneráveis. Infecções por pseudomonas podem ser tratadas com imipenem, meropenem, ciprofloxacina e gentamicina.[2]
Recentemente, análises sequenciais de ARNr 16 S têm redefinido a taxonomia de muitas espécies bacterianas.[3] Como resultado o gênero Pseudomonas inclui cepas tradicionalmente classificadas nos gêneros Chryseomonas e Flavimonas.[4] Outras cepas previamente classificadas como Pseudomonas são agora pertencentes aos gêneros Burkholderia, Brevundimonas, Stenotrophomonas, Sphingomonas e Ralstonia.
P. aeruginosa group
P. chlororaphis group
P. fluorescens group
P. pertucinogena group
P. putida group
P. stutzeri group
P. syringae group
|unused_data= ignorado (ajuda) Pseudomonas é um gênero de bactérias gram-negativas, bacilares, ubíquas (amplamente encontradas em diversos ambientes) da família Pseudomonadaceae. A espécie mais patógena é a Pseudomonas aeruginosa que causa infecções respiratórias, urinárias ou bacteriemia oportunistas em pacientes hospitalizados, com cateteres ou imunodeprimidos, otite em crianças e conjuntivite em quem usa lentes. Possuem ampla resistência, exigindo cultivo e teste de sensibilidade a antibióticos para decidir o melhor tratamento.
Pessoas com diabetes, queimaduras e fibrose cística são particularmente mais vulneráveis. Infecções por pseudomonas podem ser tratadas com imipenem, meropenem, ciprofloxacina e gentamicina.
P. aeruginosa group
P. chlororaphis group
P. fluorescens group
P. pertucinogena group
P. putida group
P. stutzeri group
P. syringae group
Pseudomonas je rod gramnegativnih, gibljivih, aerobnih, nesporogenih bacilov, ki večinoma živijo v zemlji in vodi ter lahko povzročajo bolezni.[1] V rod uvrščamo 202 različni vrsti.[2] Gre za presnovno zelo raznovrstni rod in različne vrste zato nahajamo v različnih okoljih.[3]
Psevdomonasi so zelo razširjeni tudi v vodi in rastlinskih semenih, zato so jih odkrili že zgodaj. Rod je tako poimenoval Walter Migula leta 1894.[4] Navedel je, da so nekatere vrste psevdomonasov sporogene,[5] kar se je kasneje pokazalo za napačno.[6]
Psevdomonasi so aerobne, nesporogene, ravne ali lahno ukrivljene, po Gramu negativne paličaste bakterije, ki se gibljejo z enim ali več polarnimi bički. Imajo respiracijski tip presnove s kisikom kot končnim akceptorjem elektronov. So kemoorganotrofni. Nekatere vrste so fakultativno kemolitotrofne, sposobne izkoriščati vodik ali ogljikov monoksid kot edini vir energije.[7]
Nekatere vrste so patogene za človeka, živali ali rastline. Tipska vrsta je P. aeruginosa, ki je razširjena je v naravi, zlasti v zemlji in vodi. Patogena je za človeka in živali. Povzroča zlasti gnojne procese in obarva gnoj modrozeleno. Zato jo imenujemo tudi P. pyocyánea. Za človeka so patogene tudi vrste: P. cepácia, P. diminúta, P. maltophília in P. stútzeri. P. mállei povzroča smrkavost pri kopitarjih, vendar se bolezen včasih prenese tudi na človeka. P. pseudomállei povzroča bolezen meliodozo pri glodavcih, pri ovcah, govedi, svinjah in konjih, včasih pa se bolezen prenese tudi na človeka.[7]
Pseudomonas je rod gramnegativnih, gibljivih, aerobnih, nesporogenih bacilov, ki večinoma živijo v zemlji in vodi ter lahko povzročajo bolezni. V rod uvrščamo 202 različni vrsti. Gre za presnovno zelo raznovrstni rod in različne vrste zato nahajamo v različnih okoljih.
Psevdomonasi so zelo razširjeni tudi v vodi in rastlinskih semenih, zato so jih odkrili že zgodaj. Rod je tako poimenoval Walter Migula leta 1894. Navedel je, da so nekatere vrste psevdomonasov sporogene, kar se je kasneje pokazalo za napačno.
Pseudomonas är ett släkte inom Proteobacteria.
Inom släktet återfinns bland annat:
Pseudomonas är ett släkte inom Proteobacteria.
Inom släktet återfinns bland annat:
Arten Pseudomonas fluorescens är en av de vanligaste självlysande bakterierna. Varbakterien Pseudomonas aeruginosa (pyocyanea) som har endast en cilie. Pseudomonas aeruginosa (pyocyanea) som har en tofs av cilier.Pseudomonas, Pseudomonadaceae familyasından tamamı katalaz pozitif, gram negatif, aerobik, polar flagellası ile hareket edebilen çubuk şekilli bakterileri içeren cins.
Son derece önemli olan bu cinsin türlerinin bazıları oksidaz pozitif, bazıları oksidaz negatiftir. Bazı türleri insan, hayvan ve bitki patojenidir. Pseudomanas'ları gıdalar için önemli kılan pek çok özelliğe sahiptirler. Bazı türleri proteolitik ve lipolitik aktivite göstermektedir. Aerobik olmaları nedeniyle gıdaların yüzeyinde hızla gelişebilmeleri için gerekli gelişme faktörleri ve vitaminleri sentezleme yeteneğindedirler. Psikrofil, mezofil veya psikrotrof türleri vardır. Özellikle soğukta saklanan et, tavuk eti, yumurta ve deniz ürünlerinin birinci derecede bozulma etmenidirler. Bazı gıda maddeleri üzerinde Pseudomonas fluoresans yeşilimtrak, P. nigrificans siyah, diğer türleri ise kahverengi pigment oluştururlar. Isı ve radyoasyonla kolayca öldürülürler, oksijen olmadığı zaman ve 42° üzerinde üreyemezler ve kurumaya dirençlilikleri zayıftır. Pseudomonas aeruginosa saprofit bir tür olmakla birlikte gastroenterik hastalıklara neden olabilir.
Pseudomonas, Pseudomonadaceae familyasından tamamı katalaz pozitif, gram negatif, aerobik, polar flagellası ile hareket edebilen çubuk şekilli bakterileri içeren cins.
Son derece önemli olan bu cinsin türlerinin bazıları oksidaz pozitif, bazıları oksidaz negatiftir. Bazı türleri insan, hayvan ve bitki patojenidir. Pseudomanas'ları gıdalar için önemli kılan pek çok özelliğe sahiptirler. Bazı türleri proteolitik ve lipolitik aktivite göstermektedir. Aerobik olmaları nedeniyle gıdaların yüzeyinde hızla gelişebilmeleri için gerekli gelişme faktörleri ve vitaminleri sentezleme yeteneğindedirler. Psikrofil, mezofil veya psikrotrof türleri vardır. Özellikle soğukta saklanan et, tavuk eti, yumurta ve deniz ürünlerinin birinci derecede bozulma etmenidirler. Bazı gıda maddeleri üzerinde Pseudomonas fluoresans yeşilimtrak, P. nigrificans siyah, diğer türleri ise kahverengi pigment oluştururlar. Isı ve radyoasyonla kolayca öldürülürler, oksijen olmadığı zaman ve 42° üzerinde üreyemezler ve kurumaya dirençlilikleri zayıftır. Pseudomonas aeruginosa saprofit bir tür olmakla birlikte gastroenterik hastalıklara neden olabilir.
Pseudomonas là một chi vi khuẩn xuất hiện ở mọi nơi trong môi trường. Sự biến dưỡng dễ thay đổi và linh động của chúng làm cho chúng có thể sống ở nhiều môi trường khác nhau như nước, đất, trên cây và trong các động vật. Trong số những loài Pseudomonas này, có những loài tiêu biểu có thể được sử dụng trong công nghệ sinh học.
Đặc điểm hình thái học chung cho Pseudomonas là Gram âm, tế bào hình que, di động nhờ roi ở đầu và không có bào tử.Các đặc điểm sinh lý là dị dưỡng, không lên men, linh hoạt về dinh dưỡng, không quang hợp hoặc cố định nitrogen.
Một trong những nhóm vi khuẩn hữu ích cho cây lúa cần được khai thác là vi khuẩn pseudomonas sinh huỳnh quang bởi chúng được coi là tác nhân sinh học tiềm năng trong phòng chống vi nấm gây bệnh cây trồng. Các chủng pseudomonad sinh huỳnh quang vùng rễ cây lúa đã được phân lập và chọn lọc theo khả năng đối kháng Fusarium oxysporum, tác nhân gây bệnh khô vằn cây lúa
Atrazine là một chất độc diệt cỏ (herbicid) hầu như hoàn toàn không tan trong nước (33 mg/lít), nhưng nồng độ cho phép trong nước là 0,2 mg/lít. Một số chủng vi sinh như Pseudomonas sp. strain ADP có khả năng chuyển hoá atrazine. Chủng này tiết ra Atrazine chlorohydrolase xúc tác phản ứng chuyển hoá atrazine. Như vậy, bằng phản ứng Atrazine chlorohydrolase, atrazine độc, không tan có thể chuyển hoá các sản phẩm tan được và không độc.
Enzyme parathiohydrolase do Pseudomonas SP. tổng hợp có khả năng phân hủy tới 94-98% dư thuốc trừ sâu paraythion. Hoạt độ của enzyme này phụ thuộc vào cấu trúc, độ ẩm và dung đệm của đất...
Một số Pseudomonas có thể chuyển hóa các chất ô nhiễm hóa học trong môi trường, và có thể được sử dụng để xử lý sinh học:
Trong rác bể ủ có nhiều loại vi sinh vật có khả năng phân giải tinh bột. Một số vi sinh vật có khả năng tiết ra môi trường đầy đủ các loại enzym trong hệ enzym amilaza. Ví dụ như một số vi nấm bao gồm một số loại trong các chi Aspergillus, Fusarium, Rhizopus. Trong nhóm vi khuẩn có một số loài thuộc chi Bacillus, Cytophaza, Pseudomonas... Xạ khuẩn cũng có một số chi có khả năng phân hủy tinh bột. Đa số các vi sinh vật không có khả năng tiết đầy đủ hệ enzym amilaza phân hủy tinh bột. Chúng chỉ có thể tiết ra môi trường một hoặc một vài men trong hệ đó. Các nhóm này cộng tác với nhau trong quá trình phân hủy tinh bột thành đường. Trong sản xuất người ta thường sử dụng các nhóm vi sinh vật có khả năng phân hủy tinh bột. Ví dụ trong chế biến rác thải hữu cơ người ta cũng sử dụng những chủng vi sinh vật có khả năng phân hủy tinh bột để phân hủy tinh bột có trong thành phần rác hữu cơ.
Trong rác ủ, phospho tồn tại ở nhiều dạng hợp chất khác nhau. Phospho được tích luỹ trong rác khi động thực vật chết đi, những hợp chất phospho hữu cơ này được vi sinh vật phân giải tạo thành các hợp chất phospho vô cơ khó tan. Do đó phospho tồn tại ở hai dạng: phospho hữu cơ và phospho vô cơ.
Vi sinh vật phân giải lân hữu cơ chủ yếu thuộc hai chi: Bacillus và Pseudomonas. Các loài có khả năng phân giải mạnh là Bacillus megatherium, Bacillus mycoides và Pseudomonas sp.
Vi sinh vật phân giải lân vô cơ bao gồm các loại vi khuẩn có khả năng phân giải mạnh là Bacillus megatherium, Bacillus butyricus, Bacillus mycoides. Pseudomonas radiobacter, Pseudomonas gracilis...
Nhóm P. aeruginosa
Nhóm P. chlororaphis
Nhóm P. fluorescens
Nhóm P. pertucinogena
Nhóm P. putida
Nhóm P. stutzeri
Nhóm P. syringae
Nhóm incertae sedis
Pseudomonas là một chi vi khuẩn xuất hiện ở mọi nơi trong môi trường. Sự biến dưỡng dễ thay đổi và linh động của chúng làm cho chúng có thể sống ở nhiều môi trường khác nhau như nước, đất, trên cây và trong các động vật. Trong số những loài Pseudomonas này, có những loài tiêu biểu có thể được sử dụng trong công nghệ sinh học.
Đặc điểm hình thái học chung cho Pseudomonas là Gram âm, tế bào hình que, di động nhờ roi ở đầu và không có bào tử.Các đặc điểm sinh lý là dị dưỡng, không lên men, linh hoạt về dinh dưỡng, không quang hợp hoặc cố định nitrogen.
 分類 ドメ
分類 ドメP. abietaniphila
P. abietaniphila
P. acidovorans
P. aestusnigri
P. aeruginosa
P. agarici
P. alcaligenes
P. alcaliphila
P. aminovorans
P. amygdali
P. andropogonis
P. anguilliseptica
P. antarctica
P. antimicrobica
P. argentinensis
P. arsenicoxydans
P. asplenii
P. asturiensis
P. aurantiaca
P. aureofaciens
P. avellanae
P. avenae
P. avenae subsp. avenae
P. avenae subsp. citrulli
P. avenae subsp. konjaci
P. azotifigens
P. azotoformans
Pseudonocardia babensis
P. baetica
P. balearica
P. bauzanensis
P. beijerinckii
P. benzenivorans
P. beteli
P. borbori
P. boreopolis
P. brassicacearum
P. brassicacearum subsp. brassicacearum
P. brassicacearum subsp. neoaurantiaca
P. brenneri
P. caeni
P. cannabina
P. carboxydohydrogena
P. caricapapayae
P. caryophylli
P. cattleyae
P. cedrina
P. cedrina subsp. cedrina
P. cedrina subsp. fulgida
P. cepacia
P. chengduensis
P. chloritidismutans
P. chlororaphis
P. chlororaphis subsp. aurantiaca
P. chlororaphis subsp. aureofaciens
P. chlororaphis subsp. chlororaphis
P. chlororaphis subsp. piscium
P. cichorii
P. cissicola
P. citronellolis
P. cocovenenans
P. composti
P. congelans
P. corrugata
P. costantinii
P. cremoricolorata
P. cuatrocienegasensis
P. deceptionensis
P. delafieldii
P. delhiensis
P. diminuta
P. doudoroffii
P. duriflava
P. echinoides
P. elongata
P. entomophila
P. extremaustralis
P. extremorientalis
P. facilis
P. ficuserectae
P. flava
P. flavescens
P. flectens
P. fluorescens
P. fragi
P. frederiksbergensis
P. fulva
P. fuscovaginae
P. gelidicola
P. geniculata
P. gessardii
P. gladioli
P. glathei
P. glumae
P. graminis
P. grimontii
P. guangdongensis
P. guariconensis
P. guguanensis
P. guineae
P. halophila
P. helmanticensis
P. hibiscicola
P. hussainii
P. huttiensis
P. indica
P. indigofera
P. iners
P. japonica
P. jessenii
P. jinjuensis
P. kilonensis
P. knackmussii
P. koreensis
P. kunmingensis
P. kuykendallii
P. lanceolata
P. lemoignei
P. libanensis
P. lini
P. litoralis
P. lundensis
P. lurida
P. lutea
P. luteola
P. mallei
P. maltophilia
P. mandelii
P. marginalis
P. marina
P. marincola
P. mediterranea
P. meliae
P. mendocina
P. mephitica
P. meridiana
P. mesophilica
P. migulae
P. mixta
P. mohnii
P. monteilii
P. moorei
P. moraviensis
P. mosselii
P. mucidolens
P. multiresinivorans
P. nautica
P. nitroreducens
P. oleovorans
P. oleovorans subsp. lubricantis
P. oleovorans subsp. oleovorans
P. orientalis
P. oryzihabitans
P. otitidis
P. pachastrellae
P. palleroniana
P. palleronii
P. panacis
P. panipatensis
P. parafulva
P. paucimobilis
P. pelagia
P. peli
P. perfectomarina
P. pertucinogena
P. phenazinium
P. pickettii
P. pictorum
P. plantarii
P. plecoglossicida
P. poae
P. pohangensis
P. prosekii
P. protegens
P. proteolytica
P. pseudoalcaligenes
P. pseudoalcaligenes subsp. citrulli
P. pseudoalcaligenes subsp. konjaci
P. pseudoalcaligenes subsp. pseudoalcaligenes
P. pseudoflava
P. pseudomallei
P. psychrophila
P. psychrotolerans
P. punonensis
P. putida
P. pyrrocinia
P. radiora
P. reinekei
P. resinovorans
P. rhizosphaerae
P. rhodesiae
P. rhodos
P. rubrilineans
P. rubrisubalbicans
P. sabulinigri
P. saccharophila
P. salegens
P. salomonii
P. saponiphila
P. savastanoi
P. segetis
P. seleniipraecipitans
P. simiae
P. solanacearum
P. spinosa
P. stanieri
P. straminea
P. stutzeri
P. synxantha
P. syringae
P. syringae subsp. savastanoi
P. syringae subsp. syringae
P. syzygii
P. taeanensis
P. taeniospiralis
P. taetrolens
P. taiwanensis
P. testosteroni
P. thermotolerans
P. thivervalensis
P. tolaasii
P. toyotomiensis
P. tremae
P. trivialis
P. tuomuerensis
P. umsongensis
P. vancouverensis
P. veronii
P. vesicularis
P. viridiflava
P. vranovensis
P. woodsii
P. xanthomarina
P. xiamenensis
P. xinjiangensis
P. zeshuii
シュードモナス属(-ぞく、Pseudomonas)とは、プロテオバクテリア門ガンマプロテオバクテリア綱シュードモナス科英: Pseudomonadaceaeに属する、グラム陰性好気性桿菌の属である[1]。本来のラテン語読みであるプセウドモナスと呼ばれることもある(シュードモナスは英語発音に近い)。シュードモナス属である菌株をシュードモナス属菌英: pseudomonadという。現在、218の種と18の亜種が知られている[2]。
この属は異化特性が非常に多様であり、このため幅広いニッチに生息している[3]。in vitroでの培養が容易である点とゲノムDNAの増幅が可能である点により学術的研究において重要視とされており、人間に対する日和見感染や院内感染の原因菌であるP. aeruginosa、植物の病害や霜害の原因菌であり一方で生物防除用や人工降雪用として注目されているP. syringae、生物として世界で初めて特許登録された土壌微生物であり油分汚染に対するバイオレメディエーションに有用なP. putida、植物病原性を持たず植物生長を促進するP. fluorescensなどが特に研究されている。
水圏や、双子葉類といった植物の種子にも見出せるため、シュードモナス属は微生物学の歴史の初期において認識されていた。シュードモナスという言葉は、シュードモナスを発見したと主張した[ 英: Walter Migula(英語版) ]が1894年から1900年に、グラム陰性で桿状の極鞭毛を持った細菌というあいまいな定義をして生まれた[4][5](のちにWalterの発見は微粒子の見間違いであることが証明された[6])。
分子生物学的な分類手法が登場する以前の、形態学的にシュードモナス属を定義していた特徴は以下である[7]。
その他のシュードモナス属一般の特徴として以下が挙げられる。グルコースを用いたヒュー・レイフソンテスト[注釈 1]においてガス形成はしない。血液寒天培地上でベータ溶血性を示す。インドールテストとメチルレッド、Voges–Proskauerテストに陰性であり、クエン酸テストに陽性である。
その生息域は幅広く、土壌、淡水、海水、植物や動物(ヒト、家畜、魚介類を含む)の組織、天然の霜、雲などを含む。石油、農薬(殺菌剤)、重金属などで汚染された環境においても生育する株も存在する。P. putida IH-2000は高濃度のトルエン中においても生存できる。
様々な有機化合物を分解する能力があり、自然界における分解者であるため、地球上の炭素循環に大きく貢献していると考えられている。
シュードモナス属菌は霜や雲にも分布している。霜や雲を形成する核(凍結核)としてシュードモナス属菌は最も一般的である。一部のシュードモナス属菌が凍結核となる理由は、細胞膜中に氷核活性[ 英: ice nucleation-active ](Ina)タンパク質を産生する[8][9]性質による。Inaタンパク質は比較的高い温度(-4 ℃から-2 ℃)で水の凍結を促進する。
凍結核となる性質はシュードモナス属菌の重要性の一つである。シュードモナス属菌は世界中の雨や雪の発生に最も重要であると考えられている[10]。P. syringaeは、植物の霜害における最も大きな原因であると同定されている。Inaタンパク質は人工雪の生産に用いられている[11]。
シュードモナス属菌の多くは蛍光色素を産生する。その多くはピオベルジン[ 英: pyoverdine(英語版) ](黄緑色の蛍光性のシデロホアであり、栄養としての鉄分が不足している条件下で分泌される[12]。また、一部の種はピオベルジン以外のシデロホア分子を分泌する。Pseudomonas aeruginosaを含むシュードモナス属病原性菌はピオシアニン[ 英: pyocyanin(英語版) ](青緑色)を分泌する[13]。Pseudomonas fluorescensはチオキノロバクチン[ 英: thioquinolobactin ][14]を分泌する。P. fluorescensグループの種は一般に緑色気味の水溶性色素を産生する。
相当数のシュードモナス属菌が存在するとき、アルギン酸といった菌体外多糖[ 英: exopolysaccharide(英語版) ]の分泌によりバイオフィルムが形成される。バイオフィルムはシュードモナス属菌にとって都合のよい生息場所となり、また、外敵や物理的ストレスから保護する。例えば、哺乳類の白血球による貪食[15]や、(食品に繁殖したシュードモナス属菌の)調理による除去を困難にする。
日和見感染の病原菌であるP. aeruginosaによるバイオフィルムの形成は、遺伝性疾患である嚢胞性線維症(CF)患者に対するP. aeruginosa感染の原因となる。このバイオフィルムは気道内の粘液に形成され、P. aeruginosaの生息場所となる。それに加え、CF病患者のバイオフィルムに生息するP. aeruginosaは(歴史的に、全てのシュードモナス属菌は偏性好気性生物に分類されていたにも関わらず)嫌気呼吸を行うようになる[16]。
グラム陰性菌であるため、大部分のシュードモナス属は本質的にペニシリンやその他のβ-ラクタム系抗生物質に対して耐性を持つが、ピペラシリン、イミペネム、チカルシリン、またはシプロフロキサシンに対して感受性を持つ[15]。このため、これらの抗生物質は治療に用いられるが、その他、トブラマイシン、ゲンタマイシン、アミカシンのようなアミノグリコシド系抗生物質が治療に用いられる(アミノグリコシド系抗生物質#抗緑膿菌作用のあるもの)。
P. aeruginosa(緑膿菌)は、臨床現場における日和見感染の病原体である。抗生物質に対して低感受性であるため警戒されている[17]。この低感受性は、染色体にコードされた薬剤耐性遺伝子(例えば、 mexAB-OPRM、mexXYなど[18])による多剤排出ポンプ[ 英: Bacterial efflux pumps(英語版) ]、および細菌細胞のエンベロープの低透過性に起因する。多剤排出ポンプは細胞壁中の膜貫通タンパク質ポリンであり、抗生物質が効果を現す前にこれを細胞外へ排出する。
P. aeruginosaが抗生物質の低感受性と抵抗性を獲得することがある。染色体の遺伝子の突然変異および、抗生物質抵抗性の決定因子の水平伝播はその原因となり、多剤耐性の発生も引き起こす。また、体細胞超変異は、慢性の感染症を引き起こす緑膿菌株の薬剤耐性獲得の原因の一つとなる。また、インテグロン[注釈 2]中にいくつかの異なる薬剤耐性遺伝子が集まっており、薬剤耐性決定因子の獲得につながる。複数の研究は、バイオフィルムの形成または変異体の小コロニーの出現に関連する耐性表現型が緑膿菌の抗生物質処理への応答において重要である可能性を示している[19]。
シュードモナス属の感染性種はP. aeruginosa(緑膿菌)やP. oryzihabitans、P. plecoglossicidaなどである。緑膿菌は非常に広大な分泌系を有しており、院内感染の原因は緑膿菌が分泌する多数のタンパク質であると考えられている[20]。
一部の種は環境中の化学汚染物質を代謝することができるため、バイオレメディエーションに利用することができる。バイオレメディエーション剤として適切であると実証された種として以下が含まれている。
1980年代半ば以降、農作物への病原菌の繁殖を防止するためシュードモナス属の特定の菌株が穀物種子または農耕地に散布されている。このような行為は一般的に生物防除と呼ばれている。P. fluorescensやP. protegens(CHA0株やPf-5株など)の生物防除特性が現在のところ最もよく理解されている。ただし、P. fluorescensがどのように植物生長促進効果を実現しているかは明確になっていない。一つの理論として、宿主植物への全身抵抗性を誘導し、宿主植物が本当の病原体に対してより強い抵抗性を示すようになり、一方でP. fluorescensが生存競争で土壌病原微生物を抑えるためと考えられている。P. fluorescensが他の微生物よりも生存競争で優勢となる理由として、鉄キレート剤であるシデロホアを利用するため鉄分の獲得競争で有利になるためや、フェナジン系の抗生物質やシアン化水素のような他の土壌微生物を抑制する化合物を生産するためと考えられている。2005年に行われた研究結果はこの理論を支持している[31]。
生物防除特性で注目すべきその他のシュードモナス菌種として、特定の植物病原性真菌に対して働くフェナジン系の抗生物質を産生するP. chlororaphis[32]や、その近縁種であり、グラム陽性微生物に対して働く抗生物質di-2,4-diacetylfluoroglucylmethaneを産生するP. aurantiaca[33]がある。
シュードモナス属の代謝多様性と、低温での生育能力と、非常に広範な分布とにより、多くのシュードモナス属菌種は食品の腐敗の一般的な原因である。代表的な例として、P. fragiによる肉類や酪農製品の腐敗[34]、P. taetrolensやP. mudicolensによる卵からのカビの発生[35]、P. lundensisによる牛乳、チーズ、牛肉、魚肉の腐敗が挙げられる[36]。シュードモナスの増殖による食品の腐敗は果実のような臭気を生じさせる。
シュードモナス属という語は19世紀末にWalter Migulaによって創られた。当時、この属名の語源は説明されなかったが、英: Bergey's Manual of Systematic Bacteriology(英語版)第7版(当時における、細菌の学名命名法および分類法の標準だった書籍[37])では、ギリシャ語におけるpseudes=「偽物の」(ψευδής)と-monas=「単一単位」(μονάς/μονάδος)の合成語であると記述されている。しかしながら、おそらくMigulaは「偽物の[ 英: Monas(英語版) ](微小な鞭毛をもつ原生生物)」を意図して名づけたと考えられている[6]。
Migulaの定義から数十年でこの属に割り当てられた種名の数は膨大となり、問題となった。Migulaの定義はあいまいであり、一致する種が非常に多かったためである。しかし、多くの株は、種間で保存された高分子(タンパク質や核酸など)に基づく方法や新しい分類法に基づいて再分類された[19]。例えば、リボソームRNAの配列や組成の類似性に基づいた分類法は、そのような分子生物学的手法がまだなかった当時に分類された種で構成されたシュードモナス属が、5つのいわゆるrRNAホモロジーグループ、すなわち異なる属に明確に分離できることを明確に示した。現在登録されている種の数は、当時と比べて90%以上減少している。
2000年に16S rRNA系統解析により多くの細菌種が再分類された[38]。その結果、以前は Chryseomonas属と Flavimonas属に分類されていた株をシュードモナス属は含むことになった[39]。シュードモナス属とされていた一部の株はBurkholderia属またはRalstonia属に分類されなおされている[40][41]。
2000年には、シュードモナス属に分類されている一種のゲノムの配列決定が完了した。より最近では、P. aeruginosa PAO1 (2000)、P. putida KT2440 (2002)、P. protegens Pf-5 (2005)、P. syringae pathovar tomato DC3000 (2003)、P. syringae pathovar syringae B728a (2005)、P. syringae pathovar phaseolica 1448A (2005)、P. fluorescens Pf0-1、P. entomophila L48を含む様々なシュードモナス菌株のゲノム配列決定がなされた[19]。
シュードモナス属は現在、ガンマプロテオバクテリア綱に分類されている。
ガンマ-ベータプロテオバクテリア
2000年に行われた16S rRNA系統解析により、シュードモナス属136種は5つのグループに分割され、各グループに種が分類された。
P. aeruginosa グループ
P. chlororaphis グループ
P. fluorescens グループ
P. pertucinogena グループ
P. putida グループ
P. stutzeri グループ
P. syringae グループ
所在位置不明
シュードモナス属に感染するバクテリオファージの例として以下がある。
シュードモナス属(-ぞく、Pseudomonas)とは、プロテオバクテリア門ガンマプロテオバクテリア綱シュードモナス科英: Pseudomonadaceaeに属する、グラム陰性好気性桿菌の属である。本来のラテン語読みであるプセウドモナスと呼ばれることもある(シュードモナスは英語発音に近い)。シュードモナス属である菌株をシュードモナス属菌英: pseudomonadという。現在、218の種と18の亜種が知られている。
슈도모나스(Pseudomonas)는 감마프로테오박테리아의 한 종류이다. 녹농균(Pseudomonas aeruginosa) 등을 포함하고 있다. 편모가 없는 것도 있지만, 편모가 있는 경우에는 몸의 한쪽 끝이나 양끝에 나 있다. 이것을 극생 편모라고 하는데, 콜레라균이나 녹농균 등에서 이러한 예를 볼 수 있다. 스피릴룸에는 양 끝에 1개 또는 몇 개의 편모가 있다. 한편, 콜레라균과 같은 기생균이나 아세트산균과 같은 부생균 외에, 화학 합성이나 광합성에 의해서 독립 영양 생활을 하는 종류도 이 무리에 속한다.
