Строение, форма и размер клеток Parabasalia чрезвычайно варьируют в пределах таксона. Можно выделить два основных типа внешней организации клетки, характерные ранее выделяемым группировкам парабазалий: Trichomonadida[en] и Hypermastigida. В первом случае клетка обычно имеет несколько жгутиков, отходящих от её переднего конца. Один жгутик направлен назад и формирует ундулирующую мембрану. Во втором случае клетка обычно несёт множество жгутиков, а ундулирующая мембрана отсутствует[6].
Ундулирующая мембрана, в случае её наличия, укреплена фибриллярным тяжем — костой. Эта структура встречается только у парабазалий. По ультратонкому строению фибрилл, входящих в состав косты, выделяют две основные разновидности данной органеллы — с исчерченностью A и B типа[3].
Для многих парабазалий характерно наличие особых электронно-плотных структур, ассоциированных с кинетосомами: инфра- и супракинетосомальных тел[3].
По середине клетки, от переднего к заднему концу, обычно располагается мощное микротрубочковое образование — пельта-аксостилярный комплекс. Пельта — органелла серповидной формы, поддерживающая кинетосомальный комплекс на переднем конце клетки. В зоне контакта с аксостилем в состав органеллы входят 35—40 микротрубочек. Аксостиль обычно представлен полым стержнем, проходящим по всей длине клетки и иногда выступающим за её пределы. Полость данной органеллы наполнена запасным веществом — гликогеном[7]. Пельта-аксостилярный комплекс может быть множественным, иногда слабо развит или отсутствует[3].
Уникальная клеточная структура парабазалий — парабазальный аппарат — состоит из совокупности парабазальных тел и отходящих от них парабазальных филаментов. Парабазальное тело представлено диктиосомами аппарата Гольджи. Обычно на одну диктиосому приходится 25—28 плотно прилегающих друг к другу цистерн, образующих замкнутые трубки, от которых отделяются пузырьки и гранулы с электронно-плотным содержимым. Парабазальный филамент всегда располагается параллельно стопке цистерн и состоит из 1—2 поперечно исчерченных фибрилл. Количество и расположение парабазальных тел с парабазальными филаментами в клетке может быть разным, часто коррелируя с организацией жгутикового аппарата[8].
В отличие от инфузорий, у парабазалий эволюционный процесс полимеризации жгутиков обычно не сопровождается увеличением количества ядер в клетке, хотя многоядерные формы также встречаются. Ядро, как правило, шаровидной формы, имеет одно, реже несколько ядрышек[3].
В цитоплазме парабазалий находятся многочисленные гидрогеносомы — сферические органеллы, покрытые двойной мембраной и заполненные гранулярным матриксом. Внутри данных органелл, по периферии располагаются от одной до нескольких кальций-содержащих везикул. Размер гидрогеносом в среднем составляет около 300 нанометров в диаметре, однако у Monocercomonas sp. может достигать 2 микрометров. В этих органеллах, являющихся редуцированными митохондриями, происходит анаэробное расщепление продуктов гликолиза с образованием АТФ и молекулярного водорода, сборка железосерных кластеров, там же локализуются некоторые компоненты системы синтеза аминокислот[9].
В 2007 году геном паразита человека Trichomonas vaginalis был расшифрован. Размер генома составил около 160 миллионов пар нуклеотидов, из которых около двух третей входят в состав повторяющихся последовательностей и транспозонов. По мнению авторов, подобная экспансия генома произошла при переходе данного вида от исходно анаэробного к микроаэрофильному местообитанию и, скорей всего, не характерна большинству парабазалий. В геноме T. vaginalis обнаружено около 60 тысяч белок-кодирующих последовательностей, при этом интроны найдены только в 65 из них. 152 гена предположительно были приобретены протистом в результате горизонтального переноса генов от бактерий. Большинство кодируемых ими белков вовлечены в метаболизм аминокислот и углеводов[10].
Система энергетического метаболизма парабазалий адаптирована к анаэробному или микроаэрофильному образу жизни, которые ведут эти протисты. Митохондрии представителей Parabasalia редуцировались до гидрогеносом, утративших собственный геном и большую часть компонентов электрон-транспортной цепи. За счёт данных органелл парабазалии способны продуцировать АТФ, разлагая продукты гликолиза, такие как малат и пируват, без участия кислорода. На первом этапе происходит окислительное декарбоксилирование пирувата, с образованием ацетил-КоА и углекислого газа, и переносом электрона на ферредоксин. Данная реакция катализируется ферментом пируват:ферредоксин-осидоредуктазой[en]. Затем, за счёт работы гидрогеназы[en], электрон с ферредоксина переносится на протоны H+, с образованием молекулярного водорода. АТФ генерируется в результате двухстадийной реакции, с использованием энергии тиоэфирной связи, которая высвобождается при преобразовании ацетил-КоА в ацетат[9].
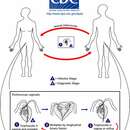
Жизненные циклы парабазалий чрезвычайно разнообразны. В качестве примера наиболее простого жизненного цикла можно привести таковой у паразита человека Trichomonas vaginalis, возбудителя венерического заболевания трихомониаз. Паразиты существуют только в виде пятижгутикового трофозоита, размножающегося бинарным делением. Передача от хозяина к хозяину происходит при половом контакте[11].
В жизненном цикле паразита птиц Histomonas meleagridis[en] присутствуют амёбоидная и жгутиковая стадии. Первая поражает ткани печени и слепой кишки хозяев, тогда как жгутиковые формы обитают в просвете последней. Обе стадии способны к активному делению. Передача от хозяина к хозяину осуществляется за счет нематод рода Heterakis[en], также обитающих в слепой кишке птиц. Червь заглатывает гистомонад вместе с пищей, после чего те проникают в клетки кишечного эпителия нового хозяина и вновь приступают к активному питанию и размножению. Затем паразиты попадают в полость тела нематоды и из неё — в клетки репродуктивных органов, в результате чего яйца нематоды оказываются заражены Histomonas. В случае поедания вышедших вместе с фекалиями яиц другими птицами те заражаются как нематодой, так и протистом[12].
Жизненный цикл парабазалий — кишечных симбионтов таракана Cryptocercus (например, представителей родов Trichonympha и Urinympha) — включает в себя вегетативную фазу, которая осуществляется в нелиняющем хозяине и представляет собой длинную череду поколений, размножающихся бесполым путём, и половой процесс, который всегда происходит во время линьки хозяина. В ходе полового процесса всегда образуются цисты или псевдоцисты, способные сохранять жизнеспособность во внешней среде, куда большая часть симбионтов попадают вместе с отслоившейся кутикулой после линьки насекомого[13].
Характер полового процесса у разных видов сильно отличается. Вегетативная стадия видов рода Trichonympha гаплоидная. При подготовке хозяина к линьке данные организмы теряют большинство клеточных органелл и инцистируются. Под оболочкой цисты происходит формирование мужской и женской гамет. После линьки цисты, оставшиеся в кишечнике хозяина или поглощённые другими насекомыми, эксцистируются, происходит слияние гамет и образование зиготы. Последняя претерпевает двухступенчатый мейоз, что приводит к формированию 4 клеток вегетативного поколения. Такой тип полового процесса называется мерогамией[14].
В отличие от вышеописанного вида, у Urinympha вегетативная стадия диплоидна, и половой процесс протекает по типу автогамии. За 8 дней до начала линьки клетки данных организмов разбирают большинство клеточных органелл и преобразуются в псевдоцисты. На этой стадии проходит одноступенчатый мейоз с образованием двух дочерних ядер без деления самой клетки. Образовавшиеся ядра сливаются, и после окончания линьки хозяина псевдоцисты вновь преобразуются в вегетативные клетки[14].
Таксон Parabasalia входит в группу эукариот Excavata, в подгруппу Metamonada[en][15]. Традиционно парабазалий рассматривали в качестве класса в типах Polymastigota или Tetramastigota, разделяя на два отряда: Trichomonadida (имеющих 4—6 жгутиков) и Hypermastigida (имеющих больше 10 жгутиков)[16][17]. В 2010 году Иван Чепичка (чеш. Ivan Čepička) с соавторами предложили рассматривать таксон в ранге типа. На основании молекулярно-филогенетических и морфологических данных авторы разделили парабазалий на 6 классов[3]:
В системе живых организмов, предложенных в 2015 году Руджеро с соавторами, таксон Parabasalia не выделяется, а всё разнообразие данной группы рассматривается в рамках классов Trichonymphea и Trichomonadea, входящих в тип Metamonada[18].
Среди парабазалий встречаются возбудители заболеваний человека, домашних и сельскохозяйственных животных. Tritrichomonas foetus[en] поражает мочеполовую систему быков, а также вызывает хроническую диарею у домашних кошек[19]. Histomonas meleagridis паразитирует в пищеварительной системе и печени птиц, вызывая тяжёлые, часто смертельные заболевания у кур и индеек[20]. Dientamoeba fragilis[en] может являться причиной болей в животе и диареи у человека[21]. Зарегистрированы случаи тяжёлых лёгочных заболеваний, вызываемых Trichomonas tenax[en][22].
Самым распространенным паразитом человека среди парабазалий является вид Trichomonas vaginalis (влагалищная трихомонада), вызывающий венерическую болезнь — урогенитальный трихомониаз. По данным ВОЗ, ежегодно в мире регистрируются более 174 миллионов случаев заболевания этой инфекцией[23].
Строение, форма и размер клеток Parabasalia чрезвычайно варьируют в пределах таксона. Можно выделить два основных типа внешней организации клетки, характерные ранее выделяемым группировкам парабазалий: Trichomonadida[en] и Hypermastigida. В первом случае клетка обычно имеет несколько жгутиков, отходящих от её переднего конца. Один жгутик направлен назад и формирует ундулирующую мембрану. Во втором случае клетка обычно несёт множество жгутиков, а ундулирующая мембрана отсутствует.
Ундулирующая мембрана, в случае её наличия, укреплена фибриллярным тяжем — костой. Эта структура встречается только у парабазалий. По ультратонкому строению фибрилл, входящих в состав косты, выделяют две основные разновидности данной органеллы — с исчерченностью A и B типа.
Для многих парабазалий характерно наличие особых электронно-плотных структур, ассоциированных с кинетосомами: инфра- и супракинетосомальных тел.
По середине клетки, от переднего к заднему концу, обычно располагается мощное микротрубочковое образование — пельта-аксостилярный комплекс. Пельта — органелла серповидной формы, поддерживающая кинетосомальный комплекс на переднем конце клетки. В зоне контакта с аксостилем в состав органеллы входят 35—40 микротрубочек. Аксостиль обычно представлен полым стержнем, проходящим по всей длине клетки и иногда выступающим за её пределы. Полость данной органеллы наполнена запасным веществом — гликогеном. Пельта-аксостилярный комплекс может быть множественным, иногда слабо развит или отсутствует.
Уникальная клеточная структура парабазалий — парабазальный аппарат — состоит из совокупности парабазальных тел и отходящих от них парабазальных филаментов. Парабазальное тело представлено диктиосомами аппарата Гольджи. Обычно на одну диктиосому приходится 25—28 плотно прилегающих друг к другу цистерн, образующих замкнутые трубки, от которых отделяются пузырьки и гранулы с электронно-плотным содержимым. Парабазальный филамент всегда располагается параллельно стопке цистерн и состоит из 1—2 поперечно исчерченных фибрилл. Количество и расположение парабазальных тел с парабазальными филаментами в клетке может быть разным, часто коррелируя с организацией жгутикового аппарата.
В отличие от инфузорий, у парабазалий эволюционный процесс полимеризации жгутиков обычно не сопровождается увеличением количества ядер в клетке, хотя многоядерные формы также встречаются. Ядро, как правило, шаровидной формы, имеет одно, реже несколько ядрышек.
В цитоплазме парабазалий находятся многочисленные гидрогеносомы — сферические органеллы, покрытые двойной мембраной и заполненные гранулярным матриксом. Внутри данных органелл, по периферии располагаются от одной до нескольких кальций-содержащих везикул. Размер гидрогеносом в среднем составляет около 300 нанометров в диаметре, однако у Monocercomonas sp. может достигать 2 микрометров. В этих органеллах, являющихся редуцированными митохондриями, происходит анаэробное расщепление продуктов гликолиза с образованием АТФ и молекулярного водорода, сборка железосерных кластеров, там же локализуются некоторые компоненты системы синтеза аминокислот.





{kind=link}