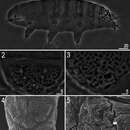
Els eutardígrads (Eutardigrada) constitueixen una classe de tardígrads sense apèndixs laterals. La majoria d'eutardígrads habiten l'aigua dolça però algunes espècies han adquirit secundàriament la capacitat de viure en entorns marins (Halobiotus). Mitjançant la criptobiosi moltes espècies són capaces de viure temporalment en entorns molt secs.
L'ordre Apochaela consta d'una única família, Milnesiidae, amb dos gèneres: Milnesium i Limmenius. Milnesium tardigradum es pot trobar a tot el món i és una de les espècies més grans entre els tardígrads (més d'1,4 mm). La boca d'aquest depredador presenta una àmplia obertura, així l'animal és capaç de menjar rotífers i els protists més grans. Les urpes són característiques. Els altres eutardígrads pertanyen a l'ordre Parachaela.
Els eutardígrads (Eutardigrada) constitueixen una classe de tardígrads sense apèndixs laterals. La majoria d'eutardígrads habiten l'aigua dolça però algunes espècies han adquirit secundàriament la capacitat de viure en entorns marins (Halobiotus). Mitjançant la criptobiosi moltes espècies són capaces de viure temporalment en entorns molt secs.
L'ordre Apochaela consta d'una única família, Milnesiidae, amb dos gèneres: Milnesium i Limmenius. Milnesium tardigradum es pot trobar a tot el món i és una de les espècies més grans entre els tardígrads (més d'1,4 mm). La boca d'aquest depredador presenta una àmplia obertura, així l'animal és capaç de menjar rotífers i els protists més grans. Les urpes són característiques. Els altres eutardígrads pertanyen a l'ordre Parachaela.
Als Eutardigrada bezeichnet man eine Klasse von Bärtierchen (Tardigrada), denen im Gegensatz zur anderen Bärtierchen-Klasse, den Heterotardigrada, auffällige Merkmale wie Kopfanhänge oder verhärtete Rückenpanzer fehlen. Sie verfügen dafür über spezielle Ausscheidungsorgane, die Malpighischen Drüsen.
Das 1929 durch den deutschen Zoologen Ernst Marcus erstmals beschriebene Taxon umfasst mehrheitlich süßwasser- und landlebende (limno-terrestrische) Formen; einige Arten finden sich allerdings auch im Salzwasser.
Bärtierchen stellen eine insgesamt sehr homogene Gruppe dar, deren Arten sich anatomisch nur in Details unterscheiden. Im Folgenden werden daher nur die für die Eutardigrada besonders charakteristischen Eigenheiten beschrieben, eine ausführlichere Einführung in den Körperbau der Tiere findet sich im Hauptartikel zu den Bärtierchen. Die durchschnittliche Körpergröße der Tiere liegt wie bei allen Bärtierchen bei etwa 100 bis 150 Mikrometern, eine Art aus der Gattung Macrobiotus wird dagegen bis zu 1,5 Millimeter lang.
Die nicht-zellige Außenhaut (Cuticula) der Eutardigrada ist sehr oft vollkommen glatt, gelegentlich granulär, also von körnigem Aussehen, und manchmal auch mit kleinen Höckern, den Tuberkeln, bedeckt. Sie besitzt aber nie auffällige Verhärtungen wie die Skleriten genannten Panzerplatten, die sich auf dem Rücken der Heterotardigrada finden; auch dornige Cuticula-Fortsätze sind bei weitem nicht so deutlich ausgeprägt wie in jener Gruppe. Dafür ist die Cuticula sehr oft durch Pigmente bunt gefärbt.
Lange Kopfanhänge wie die fädigen Cirri der Heterotardigrada besitzen die Eutardigrada nicht, allerdings finden sich um die Mundöffnung herum angeordnet oft noppenförmige Erhebungen, die Papillen oder Papulae und lappige Strukturen. Die grundlegende Beinanatomie entspricht der aller Bärtierchen; allerdings befinden sich die Klauen immer direkt am Beinende und sind niemals in Haftscheiben umgewandelt. Sie bestehen immer aus einem Doppelklauenpaar, wobei jede Doppelklaue aus einer gemeinsamen Basis, dem Klauenansatz und zwei Armen, dem primären und dem sekundären Arm besteht, deren genaue Form und Anordnung wichtige Merkmale bei der Art-Identifikation sind. Um den Klauenansatz herum findet sich oft eine aus Cuticula bestehende Verdickung, die man als Lunulus bezeichnet.
Die fachsprachlich Hämocoelom genannte Leibeshöhle fällt bei den Eutardigrada meist etwas geräumiger aus; sie wird wie bei allen Bärtierchen vom Verdauungstrakt durchzogen. Dieser beginnt mit der Mundhöhle, die sich bei manchen Arten in einen starren vorderen und einen elastischen hinteren Teil gliedert. Die Grenze zwischen beiden Regionen liegt etwa auf Höhe der Stiletthalter, welche die beiden seitlich der Mundhöhle gelegenen, oft kräftigen und manchmal gekrümmten Stilette befestigen. Zum Vorderende der Mundhöhle hin findet man oft zwei Bänder mit kleinen „Zähnchen“, den Mucrones, hinter denen sowohl rück- als auch bauchseitig quer zur Körperlängsachse (transversal) verlaufende Erhebungen auftreten können, die ebenfalls Erkennungsmerkmale bei der Artbestimmung sind. Gelegentlich wird die Mundhöhle durch eine mittig auf der Bauchseite verlaufende Leiste verstärkt.
Der muskulöse Schlund ist je nach Taxon entweder kugel-, ei- oder birnenförmig geformt und durch Placoiden verstärkt: Darunter versteht man kurze Streifen aus verhärteter Cuticula, die gewöhnlich in drei Paar Längsreihen vom Vorderende des Schlundes zum Hinterende verlaufen. Anders als bei den Heterotardigrada sind diese Längsstreifen nicht durchgehend, sondern in regelmäßigen Abständen unterbrochen. Die ersten drei Placoiden einer Reihe sind meist größer als die folgenden und werden als Macroplacoiden bezeichnet, dahinter befinden sich meist noch ein bis zwei kleinere Microplacoiden. Der Darm verläuft immer glatt und besitzt keine Diverticula genannten Ausstülpungen.
Eine bemerkenswerte Eigenheit der Eutardigrada sind die Malpighischen Drüsen, die normalerweise der Ausscheidung dienen. Die meisten Arten haben sowohl eine rückseitige (dorsale) als auch zwei seitlich angeordnete (laterale) Drüsen, die an der Verbindungsstelle von Mittel- und Hinterdarm münden. Sie bestehen aus drei bis zwölf Zellen und teilen sich meist in ein distales (darmfernes) und ein proximales (darmnahes) Segment auf. Beide Segmente haben oft die gleiche Zellenzahl, wobei 3+3 und 6+6 häufig zu findende Kombinationen darstellen. Das Distalsegment hat die Aufgabe, den Primärurin zu produzieren; wahrscheinlich geschieht dies durch aktiven Stofftransport über die trennende Membran hinweg, da es keine Hinweise auf eine Ultrafiltration der Leibeshöhlenflüssigkeit gibt. Das Proximalsegment modifiziert dann den Primärurin, in dem selektiv Wasser oder Nährstoffe wieder aufgenommen und dafür weitere Schad- oder Abfallstoffe abgegeben werden.
Eine Besonderheit findet man bei der meereslebenden Gattung Halobiotus, deren Malpighische Drüsen stark vergrößert sind. Sie ist vermutlich sekundär ins Meer zurückgekehrt und nutzt ihre Drüsen wahrscheinlich zur Regulation des Salzhaushalts (Osmoregulation). Auch limno-terrestrische Arten sind darauf angewiesen, ihre Wasseraufnahme aktiv zu kontrollieren, da ihre Körperflüssigkeit anisotonisch zu Süßwasser ist, also nicht denselben Salzgehalt aufweist. Die weitverbreitete Art Milnesium tardigradum besitzt vermutlich eine weitere Malpighische Drüse, die bauchseitig gelegen ist und anscheinend eine Rolle bei der geschlechtlichen Fortpflanzung spielt; dies wird gelegentlich als Beispiel für Exaptation gewertet, den Gebrauch einer ursprünglich aus einem anderen Grund entstandenen Struktur für eine neue Funktion.
Im Gegensatz zu den Heterotardigrada besitzen die Weibchen der Eutardigrada keine separate Geschlechtsöffnung. Stattdessen mündet der Eileiter in den Hinterdarm, der sich somit als Kloake bezeichnen lässt. Diese öffnet sich in einem quer zur Körperachse (transversal) verlaufenden Schlitz vor dem vierten Beinpaar zur Außenwelt.
Manche Weibchen besitzen einen unpaarigen Samenspeicher, in dem nach einer Kopulation das Sperma der Männchen aufbewahrt wird.
Eutardigrada finden sich weltweit auf allen Kontinenten und sind auch im Meer weit verbreitet, obwohl sie in diesen Lebensraum wahrscheinlich nur sekundär zurückgekehrt sind. Besonders häufig findet man sie aber an Land, insbesondere in der Laubstreu, den obersten Zentimetern des Waldbodens oder in Moosen (Bryata). Unterschiedliche Arten bevorzugen unterschiedliche Lebensräume innerhalb eines Mooskissens; als Folge lassen sich in einem Stück Moos oft in verschiedenen Zonen unterschiedliche Arten auffinden.
Die Lebensweise der Eutardigrada unterscheidet sich nicht allzu sehr von jener der Heterotardigrada, so dass an dieser Stelle für die Grundlagen erneut auf den Hauptartikel zu den Bärtierchen verwiesen wird. Parasitismus ist bei den Eutardigrada nicht verbreitet, was angesichts ihres hauptsächlich terrestrischen Lebensraums nicht wundernimmt: Eutardigrada sind wie alle Bärtierchen im aktiven Zustand auf einen dünnen Wasserfilm angewiesen, so dass bei Eintritt von Trockenheit entweder die Verbindung mit dem Wirt verloren gehen oder das entsprechende Individuum unkontrolliert austrocknen würde.
Dafür findet sich in der Gruppe sowohl Cyclomorphose, das Auftreten verschiedener Körperformen (Morphe) in unterschiedlichen Jahreszeiten, Cystenbildung und Kryptobiose, die wohl extremste Form der Anpassung an widrige Umweltbedingungen, bei der die betroffenen Individuen in einen todesnahen Zustand ohne Stoffwechselaktivität übergehen, der nach seiner Form als Tönnchen bezeichnet wird. Da sich kryptobiotische Zeiten auch innerhalb der Heterotardigrada finden lassen, ist diese spezielle Anpassung der Bärtierchen im Hauptartikel beschrieben.
Die Fortpflanzung der Eutardigrada kann sowohl auf asexuellem als auch auf geschlechtlichem Wege geschehen. Im ersteren Fall vermehren sich einzelne Weibchen parthenogenetisch, also ohne Beteiligung von Männchen. Man unterscheidet dabei zwischen zwei verschiedenen Formen, die beide innerhalb der Eutardigrada auftreten:
Bei der sexuellen Fortpflanzung sind fast immer die Geschlechter getrennt, obwohl in zwei Gattungen, Isohypsibius und Amphibolus auch zwittrige (hermaphroditische) Arten bekannt sind, die über eine unpaarige Keimdrüse verfügen, die gleichzeitig als Hoden und Eierstock fungiert. Selbstbefruchtung ist aber mit sehr hoher Wahrscheinlichkeit eine abgeleitete, keine ursprüngliche Eigenschaft der Gruppe.
Sind, wie meist, die Geschlechter, getrennt, so lassen sie sich daran unterscheiden, dass die Männchen eine separate Geschlechtsöffnung, die Gonopore besitzen. Bei erwachsenen Männchen sind zudem oft die Klauen des ersten Beinpaares gegenüber der gleichen Struktur der Weibchen und geschlechtsunreifen Jungtiere modifiziert. Männchen sind häufig auch etwas kleiner als die Weibchen, wobei sich allerdings erwachsene männliche und jüngere weibliche Tiere nicht nach diesem Kriterium auseinanderhalten lassen.
Die Eier der Eutardigrada können entweder glatt oder durch vielfältige Strukturen ornamentiert sein, die alle bei der Artbestimmung herangezogen werden können. Glatte Eier werden meist in der bei der Häutung abgestoßenen alten Außenhaut (Cuticula) abgelegt, die nun als Exuvium bezeichnet wird. Weibchen der Gattungen Pseudobiotus und Isohypsibius schleppen dieses Exuvium oft für eine Weile mit sich herum, was als eine Art primitiver Brutpflege gedeutet werden kann. Eier mit skulptierter Oberfläche werden dagegen meist ohne weitere Schutzhülle freigesetzt. Einige süßwasserlebende Arten aus der Gattung Murrayon legen ihre Eier in die leeren Außenskelette von Wasserflöhen (Cladocera), Ostrakoden (Ostracoda) und Insekten (Insecta) ab.
Die Entwicklung vom geschlüpften Jungtier zum geschlechtsreifen Individuum verläuft immer direkt, ohne zwischengeschaltetes Larvenstadium. Die normale Lebensdauer liegt zwischen mehreren Monaten und ein bis zwei Jahren, kann innerhalb der landlebenden Formen aber durch kryptobiotische Zeiträume, in denen die Tiere nicht altern, auf mehrere Jahre, vereinzelt wohl auch Jahrzehnte, verlängert sein.
Die genauen stammesgeschichtlichen Verwandtschaftsverhältnisse der Eutardigrada zur anderen Klasse der Bärtierchen, den Heterotardigrada, sind noch ungeklärt. Eine häufig vertretene Ansicht geht davon aus, dass die Eutardigrada ihre Wurzeln in einer bereits landlebenden Ordnung der Heterotardigrada haben, den Echiniscoidea und zwar insbesondere der Familie Echiniscidae. Damit würde sich eine Entwicklung von den meereslebenden Arthrotardigrada, der anderen Heterotardigrada-Ordnung über die Echiniscoidea hin zu den Eutardigrada ergeben:
Bärtierchen Heterotardigrada Arthrotardigradaverschiedene Familien
verschiedene Familien
verschiedene Gattungen
Eutardigrada
Vorläufige molekulargenetische Untersuchungen haben diese Hypothese jedoch nicht bestätigen können und sprechen stattdessen für eine unabhängige Entwicklung der Eutardigrada:
Bärtierchen HeterotardigradaArthrotardigrada
Echiniscoidea
Eutardigrada
Es gilt als sehr wahrscheinlich, dass die marinen Arten der Eutardigrada aus der Gattung Halobiotus sekundär ins Meer zurückgekehrt sind. Dafür sprechen insbesondere die stark vergrößerten Malpighischen Drüsen dieser Gattung, die wohl erst sekundär eine herausgehobene Funktion bei der Osmoregulation übernommen haben.
Eine fossile Art, Beorn leggi, die in kreidezeitlichem Bernstein aus Kanada gefunden wurde, lässt sich bereits in die Eutardigrada einordnen, dazu kommen etwa gleichaltrige Funde aus dem US-amerikanischen Bundesstaat New Jersey.
Davon abgesehen, dass die Gruppe anscheinend seit mehr als 65 Millionen Jahren morphologisch äußerst stabil geblieben ist, wirft keiner der Funde ein Licht auf die Naturgeschichte der Eutardigrada.
Die Eutardigrada gelten als monophyletisches Taxon, das heißt, man geht davon aus, dass alle dazugerechneten Arten auf eine gemeinsame Stammart zurückgehen und dass alle Nachkommen dieser Stammart auch in den Eutardigrada enthalten sind.
Man unterscheidet in der Klasse zwei Ordnungen mit insgesamt sechs Familien, die hauptsächlich durch die Feinstruktur der Beinklauen und der Schlundmuskulatur definiert sind:
Apochela und Parachela unterscheiden sich daneben noch in Details der Sensillenmorphologie.
Als Eutardigrada bezeichnet man eine Klasse von Bärtierchen (Tardigrada), denen im Gegensatz zur anderen Bärtierchen-Klasse, den Heterotardigrada, auffällige Merkmale wie Kopfanhänge oder verhärtete Rückenpanzer fehlen. Sie verfügen dafür über spezielle Ausscheidungsorgane, die Malpighischen Drüsen.
Das 1929 durch den deutschen Zoologen Ernst Marcus erstmals beschriebene Taxon umfasst mehrheitlich süßwasser- und landlebende (limno-terrestrische) Formen; einige Arten finden sich allerdings auch im Salzwasser.
Eutardigrada es un classe de Tardigrada.
Eutardigrada[1] maoy klase sa mga mananap. Eutardigrada sakop sa punoan tardigrada, ug ang ginharian mananap.[1]
Ang kladogram matud sa Catalogue of Life mao[1]:
tardigrada Eutardigrada
Eutardigrada are a class of tardigrades (Tardigrada) without lateral appendices. Primarily freshwater bound, some species have secondarily gained the ability to live in marine environments (Halobiotus). By cryptobiosis many species are able to live temporarily in very dry environments. More than 700 species have been described.[2]
The order Apochela consists of only one family, Milnesiidae, with two genera: Milnesium and Limmenius. Milnesium tardigradum can be found worldwide and is one of the biggest species among tardigrades (up to 1.4 mm); similar-looking species have been found in Cretaceous amber.[1] The mouth of this predator has a wide opening, so the animal can eat rotifers and larger protists. Other eutardigrades belong to the order Parachela.
Eutardigrada are a class of tardigrades (Tardigrada) without lateral appendices. Primarily freshwater bound, some species have secondarily gained the ability to live in marine environments (Halobiotus). By cryptobiosis many species are able to live temporarily in very dry environments. More than 700 species have been described.
The order Apochela consists of only one family, Milnesiidae, with two genera: Milnesium and Limmenius. Milnesium tardigradum can be found worldwide and is one of the biggest species among tardigrades (up to 1.4 mm); similar-looking species have been found in Cretaceous amber. The mouth of this predator has a wide opening, so the animal can eat rotifers and larger protists. Other eutardigrades belong to the order Parachela.
Eutardigrada es una clase perteneciente al filo Tardigrada. Está dividida en dos órdenes. Fue descrito en 1929 por el zoólogo alemán Ernst Marcus. Este taxón cuenta con más de 700 especies.[2]
El orden Apochela se compone de una sola familia, Milnesiidae, con dos géneros: Milnesium y Limmenius. Milnesium tardigradum se puede encontrar en todo el mundo y es una de las especies más grandes entre los tardígrados (hasta 1,4 mm); especies de aspecto similar se han encontrado en ámbares del periodo Cretácico. El resto de especies pertenecen al otro orden, Parachaela.
Se alimentan de organismos microscópicos, por lo que pueden ingerir algas, nematodos, rotíferos, protozoos de gran tamaño, entre otros. La mayoría de las especies de eutardígrados son formas de agua dulce y vida terrestre; sin embargo, algunas especies también habitan en agua salada.
Las características morfológicas que definen al grupo son las siguientes:[3]
Eutardigrada es una clase perteneciente al filo Tardigrada. Está dividida en dos órdenes. Fue descrito en 1929 por el zoólogo alemán Ernst Marcus. Este taxón cuenta con más de 700 especies.
El orden Apochela se compone de una sola familia, Milnesiidae, con dos géneros: Milnesium y Limmenius. Milnesium tardigradum se puede encontrar en todo el mundo y es una de las especies más grandes entre los tardígrados (hasta 1,4 mm); especies de aspecto similar se han encontrado en ámbares del periodo Cretácico. El resto de especies pertenecen al otro orden, Parachaela.
Se alimentan de organismos microscópicos, por lo que pueden ingerir algas, nematodos, rotíferos, protozoos de gran tamaño, entre otros. La mayoría de las especies de eutardígrados son formas de agua dulce y vida terrestre; sin embargo, algunas especies también habitan en agua salada.
Les Eutardigrada sont une classe de l'embranchement des tardigrades.
Selon Degma, Bertolani et Guidetti, 2013[1] :
Les Eutardigrada sont une classe de l'embranchement des tardigrades.
Gli Eutardigrada sono una classe di tardigradi senza appendici laterali. Principalmente legate all'acqua dolce, alcune specie hanno acquisito secondariamente la capacità di vivere in ambienti marini (Halobiotus). Grazie al meccanismo della criptobiosi molte specie sono in grado di vivere temporaneamente in ambienti molto secchi.
Sono state descritte poco più di 700 specie.[1]
L'ordine Apochela è costituito da una sola famiglia, Milnesiidae, con due generi: Milnesium e Limmenius. Milnesium tardigradum può essere trovato in tutto il mondo ed è una delle specie più grandi tra i tardigradi (fino a 1,4 mm); specie simili di aspetto sono stati trovati in ambra risalente al Cretaceo.[2] La bocca di questo predatore ha un'ampia apertura, quindi l'animale può mangiare rotiferi e protisti più grandi.
Le specie restanti rientrano nell'ordine Parachaela.[3]
Gli Eutardigrada sono una classe di tardigradi senza appendici laterali. Principalmente legate all'acqua dolce, alcune specie hanno acquisito secondariamente la capacità di vivere in ambienti marini (Halobiotus). Grazie al meccanismo della criptobiosi molte specie sono in grado di vivere temporaneamente in ambienti molto secchi.
Eutardigrada is een klasse van dieren die behoort tot de beerdiertjes (Tardigrada).
De verschillende soorten leven in zoet water, sommige soorten hebben zich aangepast op een leven in zee. Net als andere beerdiertjes kunnen de dieren een periode van extreme droogte doorstaan door in een ruststadium te gaan.
Eutardigrada is een klasse van dieren die behoort tot de beerdiertjes (Tardigrada).
De verschillende soorten leven in zoet water, sommige soorten hebben zich aangepast op een leven in zee. Net als andere beerdiertjes kunnen de dieren een periode van extreme droogte doorstaan door in een ruststadium te gaan.
Eutardigrada – gromada niesporczaków (Tardigrada), u których nie występują przydatki głowowe, brak kutykularnych płytek na grzbietowej i brzusznej stronie ciała, a pazurki złożone są z gałęzi głównej i bocznej[2]. Występują u nich narządy wydalnicze i kloaka[3]. Do Eutardigrada zaliczane są gatunki lądowe, słodkowodne i tylko nieliczne wtórnie morskie (Halobiotus).
Wyróżniane są dwa rzędy[4]:
Wszystkie lądowe gatunki należące do Eutardigrada wykształciły zdolność radzenia sobie w trudnych warunkach środowiskowych poprzez zapadanie w anabiozę (kryptobiozę). W tym stanie mogą przeżyć wiele lat w szerokim spektrum temperatur: zarówno w bardzo wysokiej (nawet ponad 100 °C) jak i bardzo niskiej (bliskiej zera absolutnego). Są wówczas także odporne na promieniowanie rzędu 570 000 rentgenów. W czasie suszy organizmy te mogą przejść w stan anhydrobiozy, w którym pozostają czasami nawet wiele lat, czekając na poprawę warunków środowiska[2]. Tworzą wówczas beczkowate stadia spoczynkowe o mocno zredukowanym metabolizmie[5].
Eutardigrada – gromada niesporczaków (Tardigrada), u których nie występują przydatki głowowe, brak kutykularnych płytek na grzbietowej i brzusznej stronie ciała, a pazurki złożone są z gałęzi głównej i bocznej. Występują u nich narządy wydalnicze i kloaka. Do Eutardigrada zaliczane są gatunki lądowe, słodkowodne i tylko nieliczne wtórnie morskie (Halobiotus).
Wyróżniane są dwa rzędy:
Apochela ParachelaEutardigrada é uma classe do filo Tardigrada, do reino Animal.
Eutardigrada so razred počasnikov, v katerega uvrščamo nekaj več kot 700 danes znanih opisanih vrst.[2]
Za predstavnike je značilno, da njihova kutikula ni otrdela ali strukturirana, okončine pa se končajo z enostavnimi kremplji brez izrastkov. Izvodila spolnih žlez in anus sta združena v kloako, izločala pa so Malpighijeve cevke. Po vseh teh značilnostih se razlikujejo od drugega glavnega razreda počasnikov, Heterotardigrada.[3]
Razred nadalje delimo na dva redova, Apochela in Parachela, ta dva pa potem na skupno osem družin.[4] Predstavniki prvega so kopenski, Parachela pa pretežno kopenski in sladkovodni, z nekaj morskimi predstavniki. Odrasli osebki kopenskih vrst so sposobni preiti v stanje anhidrobioze, drastične oblike dormance, po kateri so znani počasniki.[3]
Eutardigrada so razred počasnikov, v katerega uvrščamo nekaj več kot 700 danes znanih opisanih vrst.
Za predstavnike je značilno, da njihova kutikula ni otrdela ali strukturirana, okončine pa se končajo z enostavnimi kremplji brez izrastkov. Izvodila spolnih žlez in anus sta združena v kloako, izločala pa so Malpighijeve cevke. Po vseh teh značilnostih se razlikujejo od drugega glavnega razreda počasnikov, Heterotardigrada.
Razred nadalje delimo na dva redova, Apochela in Parachela, ta dva pa potem na skupno osem družin. Predstavniki prvega so kopenski, Parachela pa pretežno kopenski in sladkovodni, z nekaj morskimi predstavniki. Odrasli osebki kopenskih vrst so sposobni preiti v stanje anhidrobioze, drastične oblike dormance, po kateri so znani počasniki.
Евтардіґради мають більш-менш циліндричне коротке товсте тіло з дещо сплющеною черевною стороною, без помітної сегментації. Вважають, що воно складається з п'яти сегментів. Перший, що зливається з головною лопаттю, не має кінцівок, наступні чотири несуть по парі ніг, три з них розташовані по боках, четверта — на задньому кінці тіла. Ноги коротенькі, нечленисті, у вигляді горбикоподібних виростів тіла з рухомими кігтиками на кінцях. Більшість тихоходів майже безбарвні й навіть прозорі, іноді жовто-зелені або сливово-зелені, фіолетові чи червонуваті.
Особливістю будови тіла тихоходів є постійний клітинний склад окремих тканин та органів. Зокрема покриви тіла, м'язи та середня кишка, у більшості видів, складаються з незмінної кількості клітин, що зберігається впродовж всього життя тварин.
Зовні тіло тихоходів вкрите кутикулою, яка за тонкою будовою та хімічним складом відрізняється від кутикули членистоногих. Вона складається з епікутикули та прокутикули. На поверхні епікутикули відсутній цементний шар, а є шар слизу з кислих мукополісахаридів; восковий шар залягає між епі- та прокутикулою. У прокутикулі немає порових канальців. Кутикула тихоходів не містить хітину. Кутикула, як правило, тоненька, інколи ущільнюється, утворюючи посегментні щитки, а в деяких видів на ній є різні шило-, горбико-, крильцеподібні вирости тощо. Під кутикулою залягає епітелій, який ніколи не буває війчастим.
Порожнина тіла представлена міксоцелем, заповнений гемолімфою, в якій є клітини, наповнені запасними поживними речовинами.
М'язи в тихоходів гладенькі, представлені окремими поздовжніми та поперечними пучками на спинній, черевній та бічній сторонах; крім того, є система м'язів, пов'язаних із кінцівками. Завдяки роботі тулубних м'язів тихоходи можуть вигинати тіло (антагоністом при цьому слугує гемолімфа).
Рот розташований на черевній стороні переднього кінця тіла. У ротовій порожнині є пара гострих, спрямованих уперед стилетів, якими тихоходи проколюють здебільшого хлорофілоносні клітини мохів та водоростей, рідше — дрібних нематод, коловерток, інших тихоходів тощо, та висмоктують їхній вміст. У глотку, яка має смоктальне розширення, впадають протоки слинних залоз. Глотка веде в довгу мішкоподібну середню кишку, яка переходить у задню. Задня кишка перед анальним отвором розширюється, утворюючи клоаку.
На межі між середньою та задньою кишками є три сліпо замкнені вирости: коротенький, що лежить дорзально на кишечнику, та два довших по його боках; вони розглядаються як органи виділення, подібні до мальпігієвих судин членистоногих. Крім того, вони, ймовірно, беруть участь в осморегуляції. Непрямим підтвердженням цього є відсутність мальпігієвих судин у морських видів; у цьому разі функцію виділення виконує кишечник.
Органів кровообігу немає. Кров представлена безбарвною гемолімфою.
Органів дихання немає — дихають всією поверхнею тіла.
Нервова система складається з чотирилопатевого надглоткового ганглія, навкологлоткових конектив та черевного нервового ланцюжка з п'ятьма гангліями. Від надглоткового та черевних гангліїв відходять нерви до невеликих гангліїв, розташованих біля основи ніг, шлунка тощо.
Органи чуття розвинені слабо; у передній частині тіла є пара вічок, що складаються з кількох чутливих клітин, оточених пігментними бокалами. У морських видів тут же є кілька пар чутливих придатків, що іннервуються від надглоткового ганглія.
Тихоходи роздільностатеві. Гонади непарні, мішкоподібні, розташовані над кишечником, статеві протоки (яйце- або сім'япровід) відкриваються в клоаку, і статеві продукти виводяться через анальний отвір.
Eutardigrada можуть розмножуватися шляхом партеногенезу. У популяціях деяких видів самці трапляються значно рідше, ніж самки, а в решти видів вони зовсім невідомі. В експерименті було одержано кілька партеногенетичних поколінь Hypsibius dujardini та Milnesium taidigradum, що нормально розвивалися й жили.
Запліднення, як внутрішнє, так і зовнішнє, відбувається різними способами. Багато видів тихоходів відкладають запліднені яйця в линяльну шкірку і довго не залишають її, а разом з яйцями тягають її за собою. Рідше яйця відкладаються вільно на субстрат поодинці або невеличкими купками.
Яйця тихоходів округлі, мають міцну оболонку, на якій часто є різні вирости, специфічні для кожного виду. Кількість яєць, що відкладаються, залежить не тільки від виду, а й від фізіологічного стану самки, зокрема ступеня її ситості.
Розвиток прямий, без личинкової стадії, ріст супроводжується линяннями. Оскільки багато органів у тихоходів складаються зі сталої кількості клітин, під час росту в основному збільшується об'єм клітин, а не їхня кількість.
У 2015 році було розшифровано геном тихохода Hypsibius dujardini. Несподіванкою став той факт, що понад 17 % свого геному ці організми в ході еволюції отримали від бактерій, грибів, рослин і архей. Кількість чужорідних генів становить 6500. Вони були отримані в процесі горизонтального перенесення генів.[1] Це найбільший відомий серед тварин відсоток запозичених генів.
Тардиґради населюють надзвичайно різноманітні водні та наземні біотопи. Морські види живуть здебільшого в прибережній смузі на ґрунті й водоростях. Серед них відомі коменсали, що селяться в мантійній порожнині молюсків, на вусоногих і рівноногих раках тощо, а також один паразитичний вид Tetrakentron synaptae, який живе на щупальцях голотурії Leptosynapta galliennei й живиться вмістом живих клітин хазяїна. Прісноводні тихоходи трапляються у ставках, озерах, водосховищах, болотах річках тощо. Проте більшість представників класу трапляються на суходолі.
Цікавими є тихоходи, що живуть в екстремальних умовах. Так, тільки у воді глетчерів (глетчер — скупчення на суходолі льодових мас, що поступово рухаються під впливом власної ваги), яка збирається в невеликих заглибинах або тріщинах льоду, живе вид Hypsibius klebelsbergi при температурі 0-1,5°С. Антиподом йому є два види: Thermozodium esakii, що мешкає серед водоростей у гарячих джерелах при температурі +40°С, та Hypsibius oberbaeuseri, який водиться в мосі по краях таких джерел.
Більшість відомих видів тихоходів населюють найрізноманітніші шпаруваті субстрати різних біотопів суходолу: листяну та хвойну підстилку в лісах, ґрунт, лишайники та мохи, у тому числі й такі, що ростуть на скелях, деревах, стічних жолобах дахів тощо. Частка субсиратів перебуває постійно у воді, але для більшості з них характерні зміни висихання та зволоження, при цьому оптимальним для активного життя тихоходів є наявність водяної плівки або крапельної води, в якій тіло тварини з усіх боків обтягнене водою. У разі висихання субстрату тихоходи не гинуть, а переходять у стан прихованого життя (криптобіозу). При цьому тварина зменшується в об'ємі, її кінцівки втягуються, еластичні ділянки кутикули стягуються, утворюючи характерні складки, і тихохід набуває вигляду мікроскопічного барильця. Важливою умовою подальшого зберігання життєздатності тихоходів є повільне висихання субстрату й повільний перехід у стан криптобіозу. Після того як тварини знову потрапляють у воду, вони досить швидко оживають. Час, необхідний для відновлення життєдіяльності, залежить від тривалості висихання. Так, Macrobiotus coronifer після дев'ятимісячного висихання оживають через 25 хв, після 15-місячного — через 35 хв, а після 22-місячного — через добу.
Експериментально показано, що тихоходи на стадії барильця можуть витримувати екстремальні умови, з якими в природі вони ніколи не стикаються. Наприклад, барильця Macrobiotus не втрачали життєздатності протягом 20 місяців при температурі від —190°С до —200°С та 8 год при температурі —272°С, а також при нетривалому нагріванні до +100°С. Тихоходи виживали, навіть перебуваючи протягом кількох місяців в атмосфері, насиченій воднем, яка, як відомо, непридатна для життя.

真クマムシ綱(Eutardigrade)は、体前端に突起を持たない緩歩動物門の綱である。元々淡水に生息するが、海中での生息能を獲得した種(Halobiotus属)もある。クリプトビオシスにより、多くの種は非常に乾燥した環境でも生き延びることができる。700種以上が記載されている[1]。
遠爪目(Apochela)は、オニクマムシ科(Milnesiidae)の1科のみで、オニクマムシ属(Milnesium)とLimmeniusの2属から構成される。オニクマムシ(Milnesium tardigradum)は世界中で見られ、クマムシとしては最大の1.4mm程度まで大きくなる。似たような外見を持つ種が白亜紀のコハクの中からも発見されている[2]。口が大きく開くため、ワムシやより大きな原生生物を食べることができる。真クマムシ綱には、この他に近爪目(Parachaela)がある。
真クマムシ綱(Eutardigrade)は、体前端に突起を持たない緩歩動物門の綱である。元々淡水に生息するが、海中での生息能を獲得した種(Halobiotus属)もある。クリプトビオシスにより、多くの種は非常に乾燥した環境でも生き延びることができる。700種以上が記載されている。
遠爪目(Apochela)は、オニクマムシ科(Milnesiidae)の1科のみで、オニクマムシ属(Milnesium)とLimmeniusの2属から構成される。オニクマムシ(Milnesium tardigradum)は世界中で見られ、クマムシとしては最大の1.4mm程度まで大きくなる。似たような外見を持つ種が白亜紀のコハクの中からも発見されている。口が大きく開くため、ワムシやより大きな原生生物を食べることができる。真クマムシ綱には、この他に近爪目(Parachaela)がある。
