noms dans le fil d’Ariane
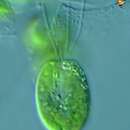
Tetraselmis ist eine Gattung von Grünalgen des Phytoplanktons innerhalb der Ordnung Chlorodendrales, die sich durch ihre intensiv gefärbten grünen Chloroplasten, ihre begeißelten Zellkörper, das Vorhandensein eines Pyrenoids innerhalb des Chloroplasten und eine schuppenförmige Thecawand auszeichnet.[1][2] Arten dieser Gattung sind sowohl in marinen als auch in Süßwasser-Ökosystemen auf der ganzen Welt zu finden; ihr Verbreitungsgebiet ist aufgrund ihrer photosynthetischen Natur hauptsächlich durch die Wassertiefe begrenzt. Sie können daher nur in Wasserumgebungen leben, wo genügend Nährstoffe und Licht für die photosynthetische Nettoaktivität vorhanden sind.[1] Tetraselmis-Arten haben sich sowohl für die Forschung als auch für die Industrie als nützlich erwiesen. Sie wurden beispielsweise untersucht, um die Wachstumsraten von Plankton zu verstehen. Seit einiger Zeit wird auch eine kolonienbildende Art verwendet, um ein Einblicke in die Evolution der Vielzelligkeit zu erlangen.[1][3] Darüber hinaus werden viele Arten aufgrund ihres hohen Lipidgehalts auf ihre Verwendung als Produzenten von Biokraftstoffen untersucht.[4]
Die Gattung wurde 1878 von Friedrich von Stein in seinem Werk Der Organismus der Infusionsthiere nach eigenen Forschungen in systematischer Reihenfolge bearbeitet. III. Abtheilung erstbeschrieben, Typusart ist Tetraselmis cordiformis.[5] Die Gattung Tetraselmis hat im Laufe der Jahre viele taxonomische Änderungen erfahren, vor allem aufgrund fortschrittlicherer Forschungstechniken, die den Wissenschaftlern bei der Klassifizierung geholfen haben. Viele Botaniker haben weiter zum Wissen um Tetraselmis beigetragen und viele Arten innerhalb der Gattung entdeckt. Einige bemerkenswerte Beiträger sind R. W. Butcher, T. Hori, R. E. Norris und M. Chihara.
Tetraselmis-Arten kommen sowohl in marinen als auch in Süßwasser-Ökosystemen vor und besetzen Nischen als Primärproduzenten in benthischen und planktonischen Nahrungsnetzen.[1] Sie sind in vielen Gewässern der Welt zu finden; der wichtigste Faktor für die Ausbreitung ihres Lebensraums ist die Verfügbarkeit von Licht, das die Zellen auf die photische Zone der Wassersäule beschränkt. Die Primärproduzenten aquatischer Ökosysteme gehören meist zum einzelliges photosynthetisches Plankton, zu dem auch Tetraselmis gehört. Planktonische Photoautotrophe sind wichtige Bestandteile aquatischer Nahrungsnetze, da sie die erste trophische Ebene bilden. Die meisten Tetraselmis-Arten sind freilebend, einige Arten sind jedoch Symbionten innerhalb von Tieren. Marine Arten besiedeln einen Lebensraum oft schnell und dicht und verursachen so Planktonblüten in Küsten- und Buchtenbereichen. Tetraselmis ist eine besonders wichtige Gattung für die Untersuchung und das Verständnis der Dynamik der Wachstumsrate von Plankton.[1]
Tetraselmis-Arten erscheinen grünlich, variieren aber stark in Zellgröße und -form. Die Zellen können rund, eiförmig, elliptisch, abgeflacht, zusammengedrückt sein oder eine Kombination dieser Formen. Die Seitenlängen variieren dabei im Bereich von 3,5–25 µm,[6] normalerweise etwa 10 µm lang × 14 µm breit.[7]
Tetraselmis-Zellen haben vier gleich lange Geißeln, die aus einer Vertiefung in der Nähe des Apex austreten. Bei den meisten Arten treten die Geißeln paarweise aus und ragen voneinander weg. Im Allgemeinen bewegen sich die einzelnen Zellen lange Zeit linear und ändern dann schnell die Richtung, ohne anzuhalten, wobei die Geißeln Kraft für die Bewegung durch die wässrige Umgebung erzeugen.[1] Siehe Abbildung 1 für allgemeine Merkmale der Geißeln und des Zellkörpers von Tetraselmis-Arten.[1]
Tetraselmis-Zellen haben einen einzigen großen Chloroplasten, normalerweise mit vier vorderen Loben (Lappen, en. lobes), der den größten Teil des Zytoplasma-Kompartiments einnimmt.[1] Einige Arten haben reduzierte oder keine Loben, und einige haben entwickelte hintere Loben. Wie bei anderen Grünalgenlinien haben die Thylakoide und Lamellen innerhalb des Chloroplasten keine festgelegte Anzahl oder Muster. Der Chloroplast enthält ein Pyrenoid, das von einer Stärkehülle umgeben ist. Daneben gibt es einen Augenfleck (Stigma).[1] Die Lage des Stigmas variiert von Art zu Art, hauptsächlich aufgrund der Zellgröße und -form. Das Stigma besteht aus zwei Schichten von Lipid-Granula, die durch ein einzelnes Thylakoid getrennt sind und parallel zur Chloroplastenmembran verlaufen. Stigmen sind eine Konzentration von Pigmentkörnchen, mit denen die Zelle Licht erkennt und sich so in der Wassersäule orientieren kann (Phototaxis). Sowohl das Pyrenoid als auch das Stigma können aufgrund ihrer zuverlässigen und inhärenten Positionierung in den Arten zur taxonomischen Klassifizierung verwendet werden.[1]
Der Zellkern liegt entweder zwischen den vorderen Loben des Chloroplasten parallel zum Pyrenoid, einige Arten haben vordere Kernloben (en. nuclear lobes) entwickelt. Der Kern kann auch zentral gelegen sein, parallel zum Geißelsäckchen (en. flagellar depression, s. u.) und (wieder) zum Pyrenoid. Im Kern bestimmter Arten kann auch „kondensiertes“ (en. condensed) Chromatin gesehen werden.[1]
Der Golgi-Apparat von Tetraselmis-Arten hat eine bestimmte Anzahl (normalerweise 2 bis 4) von Golgi-Körpern, bei manchen Arten gibt es auch 8 oder mehr haben.[ Auch dies ist ein zuverlässiges Merkmal zur taxonomischen Bestimmung der verschiedenen Tetraselmis-Spezies. Die Golgi-Körper liegen zwischen dem Chloroplasten und den Basalkörpern; „cis-Golgi“ sind dem Chloroplasten zugewandt und „trans-Golgi“ den Basalkörpern.[1]
Die Mitochondrien sind ohne Muster im Zytoplasma verteilt.[1]
Das Zytoplasma kann zusätzliche Organellen und Makromoleküle wie Lipidtropfen, Vakuolen und Lysosomen enthalten.[2] Die von Tetraselmis produzierte Stärke ist chemisch die gleiche wie die von Landpflanzen.[1]
Die Theca der meisten Arten umschließt den Zellkörper eng, kann aber aufgrund von Unregelmäßigkeiten in der Zellform abgetrennt vorliegen. Die Theca sind in der Regel glatt, doch einige Arten haben ausgeprägte Falten entwickelt, die sich in der Nähe der Zellspitze senkrecht kreuzen und der Zelle den Anschein geben, in vier Längsabschnitte unterteilt zu sein. Andere Arten haben ein verdrehtes Zellhinterteil, eine Form, die sich in der erzeugten Thecawand widerspiegelt.[1]
Die einzige Öffnung (Pore oder Perforation) in der Thecawand ist dort, wo die Geißeln austreten, was als apikale Vertiefung (en. apical depression, auch flagellar depression, vgl. Geißelsäckchen) bezeichnet wird. Diese Öffnung ist ein langer Schlitz mit Thecalfortsätzen, die ihn begrenzen und sich nach oben erstreckend die Geißelbasis schützen. In Zellen, die ihre Geißeln verloren haben, wachsen diese Thecalfortsätze aufeinander zu um die Öffnung zu bedecken, aber sie verschmelzen nicht – nicht-motile (unbewegliche) Tetraselmis-Zellen haben Geißelstummel, die voll in diesem Geißelraum verbleiben.[1]
Zusätzlich haben viele Arten dicke lockige „Haare“, die an der Zellwand in der Nähe der Geißelbasis verankert sind. Die Haare sind quer gestreift und haben ein spiralförmiges Muster, das den Geißelhaaren ähnelt, aber sie sind dicker und länger als die Geißelhaare.[1]
Im Verlauf der Zellteilung beginnt die Produktion der Theca-Schuppen während der Prophase, und die Schuppen werden sofort nach Abschluss der Zytokinese und der Trennung der beiden Tochterzellen an die Zellmembran gebracht.[2] Die Fusion der Schuppen geschieht von außen und beginnt in der Nähe des hinteren Endes und erstreckt sich nach vorne zum vorderen Ende, wo sich zuletzt der Geißelspalt bildet.[1]
Jede Zelle hat vier gleich lange Geißeln, wobei die Geißeln kürzer sind als die Länge der Zelle. Die Geißeln treten paarweise aus der apikalen Vertiefung (bei der Geißelbasis) aus und zeigen in entgegengesetzte Richtungen, wobei sie parallel zur Längsseite der Zelle verlaufen. Bei den Arten, wo die Geißeln aus einer tiefen Senke austreten, ist die mechanische Kraft, die die Geißeln erzeugen können, begrenzt. Andere Linien mit nicht vertieften Geißelansätzen können höhere Geißelkräfte erzeugen. Die Geißeln der Tetraselmis-Arten sind dick, mit schleimigem Material (en. mucilaginous material) bedeckt und haben ein stumpfes Ende. Die Zellen sind außerdem mit Geißelhaaren bedeckt, die lose an der Außenseite angebracht sind; sie sind kreuzgestreift mit zwei Schuppenschichten auf den Geißeln.[1]
Tetraselmis-Arten haben drei Lebensstadien:[1]
Das vegetative nicht-motile Stadium ist das dominante Lebensstadium der meisten Arten.[1] Der einzige strukturelle Unterschied zwischen dem gegeißelten und dem vegetativen Stadium ist die Anzahl der Schuppenschichten in der Thecawand: das gegeißelte Stadium hat immer nur eine Schicht, während das vegetative Stadium zwei oder mehr hat. Die Zellen im Zystenstadium verlieren ihre Geißeln (Flagellen) und werden daher als ‚aflagellat‘ bezeichnet; dabei produzieren sie eine dicke Thecaschale zum Schutz. Unter geeigneten Bedingungen kann in jedem der beiden unbegeißelten Stadien eine vollständige Transformation durchlaufen werden und die für die vollständige Beweglichkeit nötigen Geißeln entwickelt werden.[1]
Die Tetraselmis-Arten unterziehen einer Zellteilung (in zwei Tochterzellen) während des unbeweglichen Stadiums. Die meisten Arten durchlaufen dabei immer nur einen Teilungszyklus (nicht mehrere direkt hintereinander). Während der Zellteilung teilen sich die Organellen synchron direkt vor der Kernteilung. Die Zellteilung wird durch ein Phycoplast unterstützt. Das ist eine Mikrotubuli-Struktur, die der Zelle bei der Teilung der Kerne in jede Tochterzelle hilft. Die Tochterzellen können sich dann zu gegeißelten Zellen entwickeln oder unbeweglich bleiben, abhängig von den Umgebungsbedingungen. Zusätzlich sind vegetative Zellen, ob gegeißelt oder unbeweglich, durch vier Mikrotubuli-Kontaktpunkte an der Zellwand verankert.[1]
Tetraselmis-Arten sind von Bedeutung für die zellbiologische Forschung, für die Untersuchung von Plankton-Wachstumszyklen, und sie könnten in Zukunft für die Produktion von Biokraftstoffen von Nutzen sein.
Die Spezies T. indica wird beispielsweise verwendet, um zu untersuchen, wie asymmetrische Zellteilung während der Mitose zu klonaler Zellheterogenität führt, d. h. phänotypisch unterschiedliche Zellen trotz gleichen Gene (genotypischer Identität, Klon).[8][3] Diese Spezies bildet eine klonale Kolonie, in der die Zellen morphologisch und ultrastrukturell unterschiedlich sind, und viele Zellen spezialisieren und entwickeln unabhängige physikalische Eigenschaften. Die asymmetrische Teilung der Tochterzellen führt zu einer ungleichen Teilung von Zytoplasma und Nukleoplasma. Man glaubt, dass dieses Phänomen es den Klonen ermöglicht, ‚Subpopulationen‘ zu bilden, die nahe beieinander bleiben, in denen die Zellen miteinander kooperieren und als eine große Population agieren. Diese asymmetrische eukaryotische Zellteilung spielt eine wichtige Rolle bei der Produktion von phänotypisch vielfältigen Zellen in multizellulären Systemen. Dies ist von entscheidender Bedeutung für multizelluläre Organismen (mit Aufgabenteilung), einschließlich des Menschen.[3] Näheres siehe auch Entwicklung von Einzellern zu Vielzellern.
Tetraselmis und andere Mikroalgenarten werden als Nahrung in der Aquakultur und für biotechnologische Zwecke verwendet. Mikroalgen wie die Tetraselmis-Arten sind eine vielversprechende Quelle für die Verwendung als Biokraftstoff aufgrund ihrer schnellen Wachstumsrate, ihres hohen Lipidgehalts, ihrer kostengünstigen Photosynthese, ihres geringeren Bedarfs an landwirtschaftlichen Flächen, ihrer nützlichen Nebenprodukte und weil sie umweltfreundlich sind. Derzeit wird an spezifischen Mikroalgenarten für die Verwendung als Biokraftstoff geforscht.[4]
T. suecica EPS habt antioxidative Aktivität und zytotoxische Effekte auf Tumorzellen; daher könnten sie größere Vorteile als mögliche natürliche „Nutraceuticals“ für die pharmazeutische Industrie bieten (Antioxidative Therapie).[9]
Gattung Tetraselmis[10][11][12]
Das marine Wimpertierchen Strombidium rassoulzadegani (Oligotrichia) ernährt sich von der Alge Tetraselmis chui (PLY 429). Es behält die Chloroplasten aus seiner Nahrung und nutzt sie, um aus ihrer Photosynthese eine Nahrungsergänzung zu erhalten, betreibt also Kleptoplastidie.[13][15]
T. convolutae ist ein Symbiont der wurmähnlichen Symsagittifera roscoffensis (Gattungs-Alias Convoluta, Fam. Convolutidae, Acoela), kann aber auch frei im Ozean leben.[7][15]
Mitglieder der Gattung Tetraselmis werden parasitiert von (bzw. sind assoziiert mit) folgenden dsDNA-Viren mit Kandidatenstatus:
Hinweis: Auch wenn die (vorläufige) Namensgebung dies suggeriert, müssen diese Viren nicht notwendig Tetraselmis-Parasiten sein. Es kann sich auch um Bakteriophagen (Bakterienviren) handeln, wenn ihre Bakterienwirte symbiotisch bzw. parasitär in Tetraselmis leben, oder um Virophagen, deren Helferviren Tetraselmis (oder endosymbiotische Bakterien) infizieren (d. h. Tetraselmis-assoziierte Viren). Die endgültige Klärung des Status im Einzelfall erfordert weitere Forschung.
Tetraselmis ist eine Gattung von Grünalgen des Phytoplanktons innerhalb der Ordnung Chlorodendrales, die sich durch ihre intensiv gefärbten grünen Chloroplasten, ihre begeißelten Zellkörper, das Vorhandensein eines Pyrenoids innerhalb des Chloroplasten und eine schuppenförmige Thecawand auszeichnet. Arten dieser Gattung sind sowohl in marinen als auch in Süßwasser-Ökosystemen auf der ganzen Welt zu finden; ihr Verbreitungsgebiet ist aufgrund ihrer photosynthetischen Natur hauptsächlich durch die Wassertiefe begrenzt. Sie können daher nur in Wasserumgebungen leben, wo genügend Nährstoffe und Licht für die photosynthetische Nettoaktivität vorhanden sind. Tetraselmis-Arten haben sich sowohl für die Forschung als auch für die Industrie als nützlich erwiesen. Sie wurden beispielsweise untersucht, um die Wachstumsraten von Plankton zu verstehen. Seit einiger Zeit wird auch eine kolonienbildende Art verwendet, um ein Einblicke in die Evolution der Vielzelligkeit zu erlangen. Darüber hinaus werden viele Arten aufgrund ihres hohen Lipidgehalts auf ihre Verwendung als Produzenten von Biokraftstoffen untersucht.
Tetraselmis is a genus of phytoplankton. Tetraselmis is a green algal genus within the order Chlorodendrales, and they are characterized by their intensely-colored green chloroplast, their flagellated cell bodies, the presence of a pyrenoid within the chloroplast, and a scale-produced thecal-wall.[1][2] Species within this genus are found in both marine and freshwater ecosystems across the globe; their habitat range is mainly limited by water depth due to their photosynthetic nature.[1] Thus, they live in diverse water environments if enough nutrients and light are available for net photosynthetic activity. Tetraselmis species have proven to be useful for both research and industry. Tetraselmis species have been studied for understanding plankton growth rates, and recently a colonial species is being used to gain an understanding of multicellularity evolution.[1][3] Additionally, many species are currently being examined for their use as biofuels due to their high lipid content.[4]
The genus name was first presented to the scientific community in Germany in 1878 by F. von Stein.[5] The genus Tetraselmis has undergone many taxonomic changes over the years, largely due to more advanced research techniques that have aided scientists in their classification. Many botanists have further contributed to the knowledge surrounding Tetraselmis, and have discovered many species within the genera. Some notable contributors include R.W. Butcher, T. Hori, R.E. Norris, and M. Chihara.
Tetraselmis species are found in both marine and freshwater ecosystems, and they occupy niches as primary producers in benthic and planktonic food webs.[1] They can be found in many global waters, and their main enforcer of habitat range is light availability which restricts cells to the photic zone of the water column. The primary producers of aquatic ecosystems are mostly unicellular photosynthetic plankton, which includes Tetraselmis. Planktonic photoautotrophs are important components of aquatic food webs because they form the first trophic level. Most Tetraselmis species are free-living, however, some species are symbionts within animals.[1] Marine species often populate rapidly and densely, causing plankton blooms in shoreline and bay areas.[1] Tetraselmis is a specifically important genus for studying and understanding plankton growth rate dynamics.[1]
Tetraselmis species vary greatly in cell size and shape. Cells can be round, ovoid, elliptical, flattened, compressed, or a combination of these shapes, in which their side lengths can vary in range from 3.5-25 µm.[6] Tetraselmis cells have four flagella of equal length, which emerge from a depression near the apex.[1] In most species, the flagella emerge in pairs and project away from each other.[1] Generally, individual cells travel in a linear fashion for a long period of time, and then rapidly change direction without stopping, in which their flagella generate force for motion through an aqueous environment.[1] See figure 1 for general features of the flagella and cell body of Tetraselmis species.
They have a single large chloroplast, usually with four anterior lobes, which occupies most of the cytoplasmic compartment.[7] Some species have reduced or no lobes, and some have developed posterior lobes.[1] Like other green algal lineages, the thylakoids and lamellae have no set number or pattern within the chloroplast.[1] The chloroplast contains one pyrenoid, which is encircled in a starch-shell, and one eyespot.[1] The pyrenoid is usually central within the cell; the location of the stigma however, varies from species largely due to cell size and shape.[1] The stigma is composed of two lipid-granule layers separated by a single thylakoid, which are parallel with the chloroplast membrane.[1] Stigmas are a concentration of pigment granules which the cell uses to detect light allowing them to orientate themselves within the water column.[1] Both the pyrenoid and the stigma can be used for taxonomic classification due to their reliable and inherent positioning in species.[1] See figure 1 for a visual representation of the position of the chloroplast, stigma, pyrenoid, and flagellar depression.
The nucleus of the cell lies between the anterior lobes of the chloroplast in parallel with the pyrenoid, and some species have developed anterior nuclear lobes.[1] The nucleus is centrally located and is parallel with the flagellar depression and pyrenoid.[1] Additionally, condensed chromatin can be viewed in the nucleus of certain species.[1] See figure 1 for nuclear positioning within cells. The Golgi of Tetraselmis species has a distinct number of Golgi bodies usually within the range of 2-4, but species can have up to 8 or more.[1] This is a reliable characteristic that is used to help classify species.[1] Golgi bodies lie between the chloroplast and the basal bodies; cis-Golgi face the chloroplast and trans-Golgi face the basal bodies. Mitochondria are dispersed throughout the cytoplasm in no pattern.[1] The cytoplasm may contain additional organelles and macromolecules such as lipid droplets, vacuoles, and lysosomes.[1] Additionally, the starch produced by Tetraselmis is the same as those produced from land plants.
The theca of most species closely encloses the cell body, but can become separated due to irregularities in cell shape.[1] Theca are usually smooth, but some species have developed distinct creases that cross perpendicularly near the cell apex, giving the cell the appearance of being divided into four longitudinal sections.[1] Some species also have a twisted cell posterior that is mirrored in the thecal wall produced.[1] The only perforation of the theca is where the flagella emerge, which is termed the apical depression.[1] The pore is a long slit with thecal extensions that border it, extending upwards to protect the flagella base.[1] In cells that have lost their flagella the thecal extensions grow to cover the slit, but they do not fuse; non-motile cells of Tetraselmis have flagellar stubs that remain within the thecal compartment.[1] Additionally, many species have thick curly hairs anchored to the cell wall near the flagellar base.[1] The hairs are cross striated and have a spiral pattern, which is similar to flagellar hairs, but they are thicker and longer than flagellar hairs.[1] Scale production begins during prophase, and the scales are brought to the cell membrane promptly after completion of cytokinesis and the two daughter cells are separated.[2] Fusion of the scales happens externally and begins near the posterior end, extending forwards to the anterior end in which the flagellar slit forms last.[1]
Each cell has four flagella of equal length, in which the flagella are shorter than the length of the cell.[1] The flagella emerge from the apical depression in pairs and point in opposite directions, running parallel to the long side of the cell.[1] Due to their emergence from a deep depression, the mechanical force the flagella can produce is restricted.[1] Other lineages with non-depressed flagellar insertions can produce higher amounts of flagellar force, because the flagellar base is not geometrically limited.[1] The flagella of Tetraselmis species are thick, covered in mucilaginous material, and have a blunt end.[1] They are also covered in flagellar hairs which are loosely attached to the cell exterior; they are cross-striated with two scale layers on flagella.[1] See figure 3 for a detailed picture of flagella including their scales and hair. The inner layer is compact and densely covers the flagella, and hair cells are attached to the cell membrane through small sections of membrane that are not covered due to patterning.[1] The outer scale layer is made of small-round scales which are mainly used to overlap bare membrane not covered by the inner scales.[1] The inner scales are larger than the outer scales, and usually resemble a pentagon or square in shape.[1] See figure 1 for a generalized drawing of flagellar scales. The flagellar hairs have a distinct sequence of structural components, the proximal filament connects the flagellum and cell surface, this is followed by the tubular shaft, then the transition zone in which the shaft is modified, followed by a stretch of linear globular subunits, and the hair ends with a distal filament.[1] The distal filament and transition zone are the only components that are not produced by certain species.[1] Flagellar hairs have a fixed number of distal subunit repeats, which varies between 2-44 subunits depending on the species.[1] Thus, flagellar hairs are a good morphological tool for species comparison and classification.[1] Figure 1 includes these hairs on the flagella drawing. Species use their flagella to travel in straight lines while rotating along their axis, in which they travel in one direction for long distances and then abruptly change directions instantaneously without stopping.[1] Some species differ from this pattern of movement and move in less rigid and structured fashion.[1]
Tetraselmis species have three life stages which includes a flagellated stage, a vegetative non-motile stage, and a cyst stage.[7] The vegetative non-motile stage is the dominant life-stage of most species.[1] The only structural difference between the flagellated and vegetative stage is the number of scale layers in the thecal-wall; the flagellated stage only ever has one layer, while the vegetative state has two or more.[1] Cells in the cyst stage lose their flagella, and are termed as being aflagellate, and they produce a thick thecal-shell for protection.[1] During any of the life stages it is possible for Tetraselmis species to undergo a complete transformation and develop flagella, becoming motile.
Tetraselmis species undergo cell division during the non-motile stage, producing two daughter cells, and most species only undergo one division cycle.[1] During cell division, organelles divide synchronously before nuclear division.[1] Cell division is aided by a phycoplast, which is a microtubule structure that helps the cell divide the nuclei into each daughter cell.[1] Daughter cells can develop into flagellated cells or remain non-motile, depending on environmental conditions.[1] Additionally, vegetative cells whether flagellate or immotile are anchored to the thecal wall by four microtubule contact points.[1]
Tetraselmis species have been important for cellular biology research, for studying plankton growth cycles, and they may have future use in biofuel production. For example, the species T. indica is being used to study how asymmetric cell-division during mitosis results in clonal cell heterogeneity.[3] This species forms a clonal colony in which the cells are morphologically and ultrastructurally different, and many cells specialize and develop independent physical properties.[3] The asymmetric division of daughter cells results in the unequal division of both cytoplasm and nucleoplasm.[3] Scientists believe this phenomenon allows clones to form “sub-populations” that remain close to one another, in which the cells cooperate with one another and act as one large population.[3] Asymmetric cell division clearly plays an important role in the production of phenotypically diverse cells in multicellular systems, and this has important implications for multicellular organisms, humans included.[3]
Tetraselmis and other microalgae species are used as food in aquaculture, and for biotechnology uses.[4] Tetraselmis species, along with other microalgae are a promising source for biofuel use due to their fast growth rate, high lipid content, cheaper photosynthetic mechanisms, less need for agricultural land, useful by-products, and for being environmentally friendly. Research is currently being performed into specific microalgae species for biofuel use.[4] 'Tetraselmis has a very high lipid level; their amino acids stimulate feeding in marine organisms.
T. suecica EPS have antioxidant activity and cytotoxic effects on tumor cells; therefore, they could offer greater advantages as possible natural nutraceuticals for the pharmaceutical industry.[8]
Tetraselmis is green, motile, and usually grows 10 µm long x 14 µm wide. The species T. convolutae is a symbiont of the acoelomate animal Symsagittifera roscoffensis (and can also live freely in the ocean).[9]
Tetraselmis is a genus of phytoplankton. Tetraselmis is a green algal genus within the order Chlorodendrales, and they are characterized by their intensely-colored green chloroplast, their flagellated cell bodies, the presence of a pyrenoid within the chloroplast, and a scale-produced thecal-wall. Species within this genus are found in both marine and freshwater ecosystems across the globe; their habitat range is mainly limited by water depth due to their photosynthetic nature. Thus, they live in diverse water environments if enough nutrients and light are available for net photosynthetic activity. Tetraselmis species have proven to be useful for both research and industry. Tetraselmis species have been studied for understanding plankton growth rates, and recently a colonial species is being used to gain an understanding of multicellularity evolution. Additionally, many species are currently being examined for their use as biofuels due to their high lipid content.
Tetraselmis es un género de fitoplancton.
Las especies de Tetraselmis varían en el tamaño y forma de las células. Pueden ser esféricas, elípticas, aplanadas o una combinación de estas u otras formas.[1] Cada célula tiene un único cloroplasto que contiene una mancha ocular.[2][3]
Tetraselmis pasa por tres etapas que incluyen una etapa con flagelos, una etapa inmóvil y una etapa de quiste.[3] En la etapa con flagelos, las células individuales viajan de forma lineal durante un largo período de tiempo y luego cambian rápidamente de dirección sin detenerse, con la fuerza de cuatro flagelos gruesos de igual longitud (menor a la de la célula) que emergen en pares. En la etapa de quiste la célula pierde los flagelos. La etapa inmóvil es la dominante en la mayoría de las especies, y es en la cual se produce la división celular, tras la cual las dos células generadas pueden permanecer inmóviles o desarrollar flagelos, según las condiciones ambientales.[2]
Se encuentran tanto en ecosistemas marinos como de agua dulce, y son productores primarios en las redes tróficas bentónicas y planctónicas. Se mantienen sobre la zona fótica de la columna de agua para garantizar disponibilidad de luz. La mayoría de las especies son de vida libre, otras son simbiontes de animales. Las poblaciones marinas pueden multiplicarse rápida y densamente, lo que provoca la proliferación de plancton en las zonas costeras y bahías.[2] Por su alto contenido de lípidos se las considera para su potencial uso como biocombustible.[4]
Tetraselmis es un género de fitoplancton.
Tetraselmis, rod zelenih algi, dio je porodice Chlorodendraceae. Sastoji se od 33 priznatih vrsta[1],
Tetraselmis, rod zelenih algi, dio je porodice Chlorodendraceae. Sastoji se od 33 priznatih vrsta,
Tetraselmis je rod fitoplanktona, za katerega je značilna visoka vsebnost lipidov in aminokislin, zato je pomemben v morski prehranjevalni verigi. Predstavniki so običkane enocelične alge, po navadi velike 10 µm x 14 µm.
Vrsta T. convolutae je soživka s Symsagittifera roscoffensis (acoelomate), lahko pa živi tudi samostojno.[1]
Tetraselmis je rod fitoplanktona, za katerega je značilna visoka vsebnost lipidov in aminokislin, zato je pomemben v morski prehranjevalni verigi. Predstavniki so običkane enocelične alge, po navadi velike 10 µm x 14 µm.
Vrsta T. convolutae je soživka s Symsagittifera roscoffensis (acoelomate), lahko pa živi tudi samostojno.







