Microviridae és una família de virus bacteriòfags d’ADN monocatenari. Tenen un genoma que mesura entre 4.5-6kb i són circulars en la seva conformació. Es poden dividir en tres clades principals segons la mida del seu genoma i segons la seva similaritat amb les soques de laboratori conegudes – El clade X174-like clade, El clade G4-like clade i el clade 3-like clade.
Microviridae és una família de virus bacteriòfags d’ADN monocatenari. Tenen un genoma que mesura entre 4.5-6kb i són circulars en la seva conformació. Es poden dividir en tres clades principals segons la mida del seu genoma i segons la seva similaritat amb les soques de laboratori conegudes – El clade X174-like clade, El clade G4-like clade i el clade 3-like clade.
Die Microviridae (von griechisch μικρός – mikrós: klein) sind eine Familie von Viren, die als Bakteriophagen verschiedene Bakterien infizieren, darunter Vertreter der Enterobacteriaceae und intrazelluläre parasitäre Bakterien sowie Spiroplasma.[3][4] Aufgrund ihres Wirtsspektrums sind die Microviridae ubiquitär in Abwässern, Erde oder Fäkalien präsent. Die erste und am besten charakterisierte Spezies der Familie ist Enterobakteriophage PhiX174 (früher auch als Coliphage φX174 bezeichnet). Die Microviridae sind derzeit (Stand Januar 2021) die einzige Familie im Reich Sangervirae (monotypisch).[1]
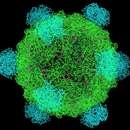
Die Virionen (Viruspartikel) der Microviridae bestehen aus einem einfachen, ikosaedrischen Kapsid (T=1), das aus drei oder vier verschiedenen Kapsidproteinen aufgebaut sind. Die unbehüllten Kapside haben einen Durchmesser von 25 bis 27 Nanometer. Die reifen Virionen gehen aus intrazellulären Vorläuferkapsiden (Prokapside) hervor, bei denen ein oder zwei Strukturproteine (scaffolding proteins, „Gerüst-Proteine“) durch ein grobes, vorläufiges Gerüst die Bindung der DNA und den Zusammenbau der Kapsomere ermöglichen.[5] Diese Gerüstproteine werden bei der Reifung des Kapsides wieder aus dem Verband gelöst, sobald die Hauptkapsidproteine eine ikosaedrische Anordnung eingenommen haben und das Kapsid geschlossen ist. Die Hauptkomponenten der Kapside bestehen aus einem Spike-Protein und einem Kapsidprotein (F oder Vp1), die sich zu großen fünfstrahligen (Pentamere) Kapsomeren zusammenlagern. Die 5,4 nm (bei Enterobakteriophagen PhiX174) weit nach außen ragenden Spike-Proteine vermitteln die spezifische Anheftung und Aufnahme in die Bakterienzelle.
Die Kapside der Gattungen Bdellomicrovirus und Chlamydiamicrovirus zeigen eine spezifische Dichte von 1,30 bis 1,31 g/cm³ in der Ultrazentrifugation mit Cäsiumchlorid, während die Vertreter der beiden anderen Gattungen alle eine signifikant höhere Dichte von etwa 1,40 g/cm³ aufweisen. Die Virionen sind sehr umweltstabil bei pH-Werten zwischen 6,0 und 9,0 und können mit Detergenzien, 2-Propanol oder Chloroform nicht inaktiviert werden.
Die Microviridae besitzen als Genom ein ringförmig geschlossenes (zirkuläres), einzelsträngiges (englisch single stranded) DNA-Molekül mit positiver Polarität. Es umfasst bei der Unterfamilie Bullavirinae zwischen 5.300 und 6.100 Nukleotide, die anderen Gattungen besitzen deutlich kleinere Genome mit 4.400 bis 4.900 Nukleotiden. Die Anordnung der vier größten offenen Leserahmen (ORF) ist innerhalb der Familie gleichförmig; sie sind meist von kurzen, nichtcodierenden Abschnitten voneinander getrennt. Zusätzlich existieren verschiedene ORFs, die mit anderen Leserastern in die großen Leserahmen eingebettet sind. Die Replikation des Genoms verläuft bei den Microviridae über eine doppelsträngige DNA-Zwischenstufe, im Detail ist die Replikation zwischen den Gattungen jedoch sehr unterschiedlich. Bei den Microviridae ist ein Horizontaler Gentransfer beschrieben, der eine größere Variabilität der Genome innerhalb der Virusfamilie hervorgebracht hat und der in wesentlich höherem Umfang als bei doppelsträngigen DNA-Bakteriophagen vorkommt.[6] Das Chromosom des Enterobakteriophagen PhiX174 war das erste DNA-Molekül, das man noch vor dem Simian-Virus 40 (SV40 bzw. MmPV1) und dem Plasmid pBR322 vollständig sequenzierte.[7]
Viren der Unterfamilie Bullavirinae infizieren Enterobakterien, zu denen auch Escherichia coli und Salmonella enterica gehören, Spezies der Gattung Bdellomicrovirus haben Bakterien der Gattung Bdellovibrio als Wirtszellen. Dieses Wirtspektrum umfasst also bei beiden Gattungen gramnegative, aerobe Bakterien. Die beiden anderen Gattungen infizieren verschiedene Gruppen von intrazellulär sich vermehrende Bakterien: Vertreter der Gattung Clamydiamicrovirus parasitieren in Chlamydien, jene der Gattung Spiromicrovirus in der Bakteriengattung Spiroplasma aus der Klasse der zellwandlosen Mollicutes (Ordnung Entomoplasmatales), die ihrerseits in Kleinsäugern und Insekten parasitieren.
Die Familie Microviridae wird nach International Committee on Taxonomy of Viruses (ICTV) mit Stand Juni 2021 in zwei Unterfamilien unterteilt, wobei sich die Unterfamilie Bullavirinae (hochgestufte frühere Gattung Microvirus) von der anderen Unterfamilie Gokushovirinae in ihren Wirten und ihrer Genomorganisation deutlich unterscheidet. Viren der Unterfamilie Bullavirinae infizieren Enterobakterien, Gokushoviren infizieren dagegen intrazelluläre Parasiten. Der Name Bullavirinae kommt vom lateinischen Wort bulla (Chef, Knopf, Stift),[8] der Name Gokushovirinae leitet sich aus dem Japanischen für ‚sehr klein‘ ab (Kanji: 極小 Hiragana: ごくしょうの gókushō(no) mikroskopisch, winzig, minimal).
Als mögliche dritte Unterfamilie wurde „Alpavirinae“ vorgeschlagen, deren Viren die Ordnung Bacteroidales infizieren.[9] Die Bezeichnung kommt vom Sanskrit-Wort अल्प (álpa) für klein, mini. Dies ist eine Gruppe von Viren, die bisher nur als Prophagen bekannt sind, und weitere Untersuchungen an diesen Viren sind nötig, bevor der Status als Unterfamilie vom ICTV gewährt werden kann.
Als möglich vierte Unterfamilie wurde „Pichovirinae“ vorgeschlagen.[10] Die Mitglieder dieser Gruppe haben eine Genomorganisation, die sich von den anderen in der Familie Microviridae. Der Name leitet sich vom okzitanischen Wort ‚picho‘ für ‚klein‘ ab.
Ein weiteres Virus (Microphage ΦCA82) wurde aus dem Darm von Truthühnern isoliert[11][12] und inzwischen in die Nähe von Spiroplasma virus SpV4 verortet.[13]
Die Systematik nach ICTV mit Stand Juni 2021, ergänzt um die obigen Vorschläge und einige Kandidaten nach NCBI, ist damit wie folgt:[2]
Die Microviridae (von griechisch μικρός – mikrós: klein) sind eine Familie von Viren, die als Bakteriophagen verschiedene Bakterien infizieren, darunter Vertreter der Enterobacteriaceae und intrazelluläre parasitäre Bakterien sowie Spiroplasma. Aufgrund ihres Wirtsspektrums sind die Microviridae ubiquitär in Abwässern, Erde oder Fäkalien präsent. Die erste und am besten charakterisierte Spezies der Familie ist Enterobakteriophage PhiX174 (früher auch als Coliphage φX174 bezeichnet). Die Microviridae sind derzeit (Stand Januar 2021) die einzige Familie im Reich Sangervirae (monotypisch).
Microviridae is a family of bacteriophages with a single-stranded DNA genome. The name of this family is derived from the ancient Greek word μικρός (mikrós), meaning "small".[1][2] This refers to the size of their genomes, which are among the smallest of the DNA viruses. Enterobacteria, intracellular parasitic bacteria, and spiroplasma serve as natural hosts. There are 22 species in this family, divided among seven genera and two subfamilies.[3][4]
The virions are non-enveloped, round with an icosahedral symmetry (T = 1). They have a diameter between 25–27 nanometers and lack tails. Each virion has 60 copies each of the F, G, and J proteins and 12 copies of the H protein. They have 12 pentagonal trumpet-shaped pentamers (~7.1 nm wide × 3.8 nm high), each of which is composed of 5 copies of the G and one of the H protein.
Viruses in this family replicate their genomes via a rolling circle mechanism and encode dedicated RCR initiation proteins.[5][6]
Although the majority of species in this family have lytic life cycles, a few may have temperate life cycles.[7]
The genome sizes range from 4.5–6kb and is circular. It encodes 11 genes (in order: A, A*, B, C, K, D, E, J, F, G, and H), nine of which are essential. The nonessential genes are E and K.[8][9] Several of the genes have overlapping reading frames.[10][11] Protein A* is encoded within protein A. It lacks ~1/3 of the amino acids from the N terminal of the A protein and is encoded in the same frame as the A protein. It is translated from an internal start site within the messenger RNA. Gene E is encoded with gene D with a +1 frameshift. Gene K overlaps genes A, B, and C. The origin of replication lies within a 30 base sequence.[12] The entire 30 base sequence is required for replication.[13]
The major capsid protein (F) has 426 amino acids, the major spike protein (G) has 175 amino acids, the small DNA-binding protein (J) has 25–40 amino acids, and the DNA pilot protein (H) has 328 amino acids.[14] The major folding motif of protein F is the eight-stranded antiparallel beta barrel common to many viral capsid proteins.[15] The G protein is a tight beta barrel with its strands running radially outward. The G proteins occur in groups of five forming 12 spikes that enclose a hydrophilic channel. The highly basic J protein lacks any secondary structure and is situated in an interior cleft of the F protein. It has no acidic amino acid residues in the protein and the twelve basic residues are concentrated in two clusters in the N-terminus separated by a proline-rich region.
Assembly of the virion uses two scaffolding proteins, internal scaffolding protein B and external scaffolding protein D. The function of protein B seems to be to lower the amount of protein D needed by the virion for assembly.[16] Protein H is a multifunctional structural protein required for piloting the viral DNA into the host cell interior during the entry process. Protein E is a 91-amino acid membrane protein that causes host cell lysis by inhibiting the host translocase MraY.[17] This inhibitory activity is located within the N terminal 29 amino acids.[18] Protein A is a single strand endonuclease[19] and is responsible for the initiation of viral DNA replication.[20] It catalyses cleavage and ligation of a phosphodiester bond between a G and A nucleotide residue pair at the phi X origin.[21] It may not be essential for phage viability but burst sizes are reduced by 50% when it is mutated.[22] Protein A* inhibits host DNA replication.[23] Unlike protein A it is capable of cleaving the phi X viral DNA in the presence of single-stranded binding protein of the host.[24] Protein A*, like Protein A, may not be required for phage viability.[25] Protein C increases the fidelity of the termination and reinitiation reactions and is required for the packagaging of the viral DNA in to the protein shell.[26] Protein K has 56 amino acids and is found in the membrane of the host cell. It appears to be able to increase the burst size of the virus.[27]
This family is divided into two subfamilies: Gokushovirinae and Bullavirinae (former genus Microvirus). These groups differ in their hosts, genome structure, and viron composition. The name Gokushovirinae is derived from the Japanese for very small. Gokushoviruses are currently known to infect only obligate intra-cellular parasites. The members of the subfamily Bullavirinae all infect Enterobacteria.
A putative third grouping has been proposed—Alpavirinae—which infect the order Bacteroidales.[7] This a group of viruses known only as prophages and additional work on these viruses seems indicated before subfamily status is granted.
A fourth clade has been proposed—Pichovirinae.[28] This clade has a genome organisation that differs from the other members of this family. The name is derived from picho which means small in Occitan.
Another virus has been isolate from the turkey gut with features similar to other microviruses but quite distinct from the known species.[29]
Members of the subfamily Bullavirinae (former genus Microvirus) have four structural proteins: major capsid protein F, major spike protein G, a small DNA-binding protein J (25 - 40 amino acids in length) and DNA pilot protein H. Assembly of the viron uses two scaffolding proteins, internal scaffolding protein B and external scaffolding protein D. Protein H is a multifunctional structural protein required for piloting the viral DNA into the host cell interior during the entry process. The genomes are between 5.3 and 6.2 kilobases (kb) in length.
Members of this subfamily can be separated into three main clades according to genome sizes.[30] Size variability within the groups occurs mainly as a result of insertions and deletions of the intergenic regions. Viruses are assigned according to their similarity to known lab based strains—the ΦX174-like clade, G4-like clade and the α3-like clade. The ΦX174-like clade of microviridae have the smallest and least variable genomes (5,386–5,387 bp); the G4-like clade varies in size from 5,486–5,487 bp; while the largest genome sized group is the α3-like clade with genomes ranging from 6,061–6259bp.
Members of the subfamily Gokushovirinae have only two structural proteins: capsid proteins F (Virus Protein 1) and DNA pilot protein H (Virus Protein 2) and do not use scaffolding proteins. They also possess 'mushroom-like' protrusions positioned at the three-fold axes of symmetry of their icosahedral capsids. These are formed by large insertion loops within the protein F of gokushoviruses and are absent in the microviruses. They lack both the external scaffolding protein D and the major spike protein G of the species in the genus Microvirus. The genomes of this group tend to be smaller—about 4.5 kb in length. This subfamily includes the genera Bdellomicrovirus, Chlamydiamicrovirus and Spiromicrovirus.
There are a number of steps in the life cycle
1. Adsorption to the host via specific receptor(s)
2. Movement of the viral DNA into the host cell
3. Conversion of the single strand form to a double-stranded intermediate
This is known as the replicative form I.
4. Transcription of early genes
5. Replication of the viral genome
Viral protein A cleaves replicative form I DNA strand at the origin of replication (ori) and covalently attaches itself to the DNA, generating replicative form II molecule. Replication of the genome now begins via a rolling circle mechanism. The host's DNA polymerase converts the single-stranded DNA into double-stranded DNA.
6. Late genes are now transcribed by the host's RNA polymerase.
7. Synthesis of the new virons
Viral protein C binds to replication complex, inducing packaging of new viral positive-stranded DNA into procapsids.[31] The preinitiation complex consists of the host cell protein rep and viral A and C proteins.[32] These associate with the procapsid forming a 50S complex.
8. Maturation of the virons in the host cytoplasm
9. Release from the host
Cell lysis is mediated by the phiX174-encoded protein E, which inhibits the peptidoglycan synthesis leading to an eventual bursting of the infected cell.
Microviridae is a family of bacteriophages with a single-stranded DNA genome. The name of this family is derived from the ancient Greek word μικρός (mikrós), meaning "small". This refers to the size of their genomes, which are among the smallest of the DNA viruses. Enterobacteria, intracellular parasitic bacteria, and spiroplasma serve as natural hosts. There are 22 species in this family, divided among seven genera and two subfamilies.
Microviridae es una familia de virus que infectan bacterias (bacteriófagos). Poseen un genoma de ADN monocatenario, por lo que se incluyen en el Grupo II de la Clasificación de Baltimore. La cápside está estructuralmente definida por una simetría icosaédrica regular y carecen de cola y de envoltura viral.
También se propusieron dos subfamilias adicionales Alpavirinae y Pichovirinae según estudios de metagenómica y elementos virales endógenos.
Los viriones no tienen envoltura, son redondos con simetría icosaédrica (T = 1). Tienen un diámetro de entre 25 y 27 nanómetros y carecen de colas. Cada virión tiene 60 copias de cada una de las proteínas F, G y J y 12 copias de la proteína H. Tienen 12 pentámeros pentagonales en forma de trompeta (~ 7,1 nm de ancho × 3,8 nm de alto), cada uno de los cuales está compuesto por 5 copias de la proteína G y una de la proteína H.
Los tamaños del genoma oscilan entre 4,5 y 6 kb y son de ADN monocatenario circular. Codifican 11 genes (en orden: A, A *, B, C, K, D, E, J, F, G y H), nueve de los cuales son esenciales. Los genes no esenciales son E y K. Varios de los genes tienen marcos de lectura superpuestos. La proteína A * está codificada dentro de la proteína A. Carece de ~ 1/3 de los aminoácidos del terminal N de la proteína A y está codificada en el mismo marco que la proteína A. Se traduce desde un sitio de inicio interno dentro del ARN mensajero. El gen E está codificado con el gen D con un desplazamiento de marco de +1. El gen K se superpone a los genes A, B y C. El origen de la replicación se encuentra dentro de una secuencia de 30 bases. Se requiere la secuencia completa de 30 bases para la replicación.
La proteína de la cápside principal (F) tiene 426 aminoácidos, la proteína de pico principal (G) tiene 175 aminoácidos, la proteína de unión al ADN pequeña (J) tiene de 25 a 40 aminoácidos y la proteína piloto de ADN (H) tiene 328 aminoácidos. El motivo de plegamiento principal de la proteína F es el barril beta antiparalelo de ocho hebras común a muchas proteínas de la cápside viral. La proteína G es un barril beta apretado con sus hebras que se extienden radialmente hacia afuera. Las proteínas G se encuentran en grupos de cinco formando 12 picos que encierran un canal hidrofílico. La proteína J altamente básica carece de estructura secundaria y está situada en una hendidura interior de la proteína F. No tiene residuos de aminoácidos ácidos en la proteína y los doce residuos básicos se concentran en dos grupos en el extremo N separados por una región rica en prolina.
El ensamblaje del virión utiliza dos proteínas de andamiaje, la proteína de andamiaje interno B y la proteína de andamiaje externo D. La función de la proteína B parece ser reducir la cantidad de proteína D que necesita el virión para el ensamblaje. La proteína H es una proteína estructural multifuncional necesaria para pilotar el ADN viral en el interior de la célula huésped durante el proceso de entrada. La proteína E es una proteína de membrana de 91 aminoácidos que causa la lisis de la célula huésped al inhibir la translocasa MraY del huésped. Esta actividad inhibidora se encuentra dentro de los 29 aminoácidos N terminales. La proteína A es una endonucleasa monocatenaria y es responsable del inicio de la replicación del ADN viral. Cataliza la escisión y ligación de un enlace fosfodiéster entre un par de residuos de nucleótidos G y A en el origen phi X. Puede que no sea esencial para la viabilidad de los fagos, pero los tamaños de ráfaga se reducen en un 50% cuando se muta. La proteína A * inhibe la replicación del ADN del huésped. A diferencia de la proteína A, es capaz de escindir el ADN viral phi X en presencia de la proteína de unión monocatenaria del huésped. La proteína A *, como la proteína A, puede no ser necesaria para la viabilidad de los fagos. La proteína C aumenta la fidelidad de las reacciones de terminación y reinicio y es necesaria para el empaquetado del ADN viral en la capa de proteína. La proteína K tiene 56 aminoácidos y se encuentra en la membrana de la célula huésped. Parece ser capaz de aumentar el tamaño de la ráfaga del virus.
Los virus de esta familia replican sus genomas a través del mecanismo de replicación en círculo rodante y codifican proteínas de iniciación de RCR dedicadas.
Aunque la mayoría de las especies de esta familia tienen ciclos de replicación líticos, algunas pueden tener ciclos de replicación templados.
Hay una serie de pasos en el ciclo de replicación.
1. Adsorción al huésped a través de receptores específicos
2. Movimiento del ADN viral hacia la célula huésped
3. Conversión de la forma monocatenaria en una intermedia bicatenaria
Esto se conoce como la forma replicativa I.
4. Transcripción de genes tempranos
5. Replicación del genoma viral
La proteína viral A escinde la hebra de ADN de la forma I replicativa en el origen de la replicación y se une covalentemente al ADN, generando la molécula de la forma II replicativa. La replicación del genoma ahora comienza a través del mecanismo en círculo rodante. La ADN polimerasa del huésped convierte el ADN monocatenario en ADN bicatenario.
6. Los genes tardíos ahora son transcritos por la ARN polimerasa del huésped.
7. Síntesis de los nuevos virones
La proteína C viral se une al complejo de replicación, induciendo el empaquetamiento de nuevo ADN viral de cadena positiva en procápsidas. El complejo de preiniciación consiste en la proteína Rep y las proteínas A y C virales. Estos se asocian con la procapside formando un complejo 50S.
8. Maduración de los virones en el citoplasma del hospedador.
9. Liberación del anfitrión
La lisis celular está mediada por la proteína E codificada por phiX174, que inhibe la síntesis de peptidoglicano que conduce a un eventual estallido de la célula infectada.
Microviridae es una familia de virus que infectan bacterias (bacteriófagos). Poseen un genoma de ADN monocatenario, por lo que se incluyen en el Grupo II de la Clasificación de Baltimore. La cápside está estructuralmente definida por una simetría icosaédrica regular y carecen de cola y de envoltura viral.
Microviridae (microvíridos) é unha familia de bacteriófagos.
Microviridae (microvíridos) é unha familia de bacteriófagos.
Mikrowirusy (Microviridae) – rodzina wirusów, charakteryzujących się następującymi cechami:
Przebieg infekcji
Mikrowirusy (Microviridae) – rodzina wirusów, charakteryzujących się następującymi cechami:
Symetria: ikozaedralna Otoczka lipidowa: brak Kwas nukleinowy: koliste ss DNA wielkości ok. 5 kpz Wielkość: 25 nm Gospodarz: bakterie Przykłady: bakteriofag ΦX174Przebieg infekcji
Zakażenie komórki bakteryjnej zachodzi dzięki rozpoznaniu receptorów bakterii i białka gpH płaszcza. Po wprowadzeniu DNA do komórki następuje szybko synteza nici komplementarnej. Powstaje tzw. "forma replikacyjna" RFI. RFI replikuje kilkakrotnie będąc związana z błoną komórkową. Replikacja zachodzi tu semikonserwatywnie według modelu theta. Po tej fazie ekspresji ulegają wczesne geny wirusa umożliwiające przystąpienie do dalszych faz. Następuje kolejna tura replikacji. Tym razem w cytoplazmie i metodą obracającego się koła (model sigma). Powstają jednoniciowe cząsteczki, które staną się genomami wirusów potomnych. W wyniku ekspresji genów późnych dochodzi do powstania białka kapsydu oraz enzymu lizującego, który niszczy komórkę i uwalnia wiriony potomne.