Morphologically, C. posadasii is completely indistinguishable from C. immitis. Both have the same dimorphic forms, one saprobic and one parasitic, and both infect exactly the same hosts. Studies have shown that the arthroconidia of both species are not statistically different in terms of length or size. In fact, the two fungi are so similar that C. posadasii was originally known as the non-CA (non-Californian) C. immitis, since the only known distinguishing factor between the two was its distribution. On a molecular level, however, the two can be easily distinguished. In their description of the species, Fisher et al. used PCR to study nine microsatellite-containing loci in order to distinguish the two species. All nine loci (specifically GAC, 621, GA37, GA1, ACJ, KO3, KO7, KO1, and KO9) were found to be polymorphic, each site containing 8 to 26 alleles. The two clades were found through this genetic analysis. Of these loci, GAC and 621 were found to be the distinguishing factors between C. posadasii and C. immitis.
Another interesting phenotypic observation is the different growth rates of the two species on media with high NaCl concentrations. Fisher et al took samples of each species and cultured them on media with low and high levels of NaCl concentration. On the less salty media, while the C. immitis populations initially grew faster than those of C. posadasii, by day ten there were no significant differences between the two in terms of population size and growth rate. However, the mean growth rate of C. immitis on the media with high salt concentration was significantly faster than that of C. posadasii. Despite this, growth rate in salty media cannot correctly be used to distinguish the two species because their ranges of growth rates overlap.
Unlike C. immitis, which is only found in California’s San Joaquin Valley, C. posadasii is found in the soils of the southwestern United States, Mexico, and Central and South America. They generally are not found in the San Joaquin Valley, and although a few cases have been found in patients from that region, there is a great possibility that the infection was picked up outside of the valley or that wind storms blew the spores into the valley. In the United States, C. posadasii infections are especially prominent in Arizona, with smaller endemic regions in Central and South America. However, C. posadasii is most prominent in Mexico, with human infection rates between 10% and 40% of the population in the northern region.
Coccidioides posadasii was first discovered in Argentina in 1890 by Alejandro Posadas, after whom the fungus is named. The doctor had a patient by the name of Domingo Ezcurra, who had the first documented case of coccidioidmycosis, also known as Valley Fever, a serious fungal infection caused by C. posadasii and its close relative, Coccidioides immitis. Posadas followed Ezcurra’s case until the latter’s death, after which his head was preserved and put on display in an anatomy museum, where it is still studied today.
C. posadasii is a dimorphic pathogen, meaning it has two major morphological forms in its life cycle. In its vegetative state, it lives in the soil as a saprobe, using its thin, filamentous hyphal form to digest organic materials. The mycelium is septate, and when cultured, the fungus is initially moist and smooth and then becomes suede-like and greyish-white and brown. When the soil begins to dry out and nutrients run low, C. posadasii’s inner cell walls thicken and alternate compartments are broken down to form arthroconidia, also known as arthrospores. These spores form by means of mitosis and are anywhere from rectangular to barrel-shaped, approximately three by five micrometers in size. The arthroconidia are eventually are dispersed by wind or other soil disturbances. This allows them to either be dispersed to another location, where the hyphal form is reformed, or to be inhaled by humans, cows, rodents, and other potential host organisms.
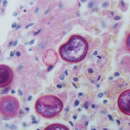
It is at this point that the fungus reverts to its parasitic phase and its second major morphological form. C. posadasii is a thermal dimorph, meaning that heat triggers its change in form. Thus, the host organism’s body temperature (usually around 37 degrees Celsius) will turn the spores into spherules. These structures, which have membranous walls rich in lipids and are anywhere from 20 to 80 micrometers in diameter, in turn septate and produce around 200 to 300 uninucleate endospores. These endospores are released when the sphorule breaks open and can be carried around the body by the bloodstream or the lymphatic system to infect other tissues. Susceptible tissues include the lungs, skin, heart, pericardium, bones, and central nervous system.
No method of sexual reproduction in C. posadasii has been found, which makes them mitosporic, also called imperfect, fungi. The absence of a sexual phase made classifying the fungus very difficult at first. It was only when Sigler and Carmichael noticed the similarities between the arthroconidia of C. posadasii and the aleurioconidia of Malbranchea Sacc. This indicated that the fungus probably belonged in the order Onygenaceae, a classification that was later confirmed by molecular methods. Although there is no evidence of sexual reproduction, there are “sexes” within the species. Individuals within coccidioides strains contain one of two alleles at a specific locus, each anywhere from 8.1 to 9 kb in length and made up of five or six genes.
C. posadasii can be found in arid, sandy climates like those found in the Lower Sonoran Life Zone, as subtropical deserts and thorny deserts were described by C.H. Merriam. These areas generally receive about 10 inches of rain annually and are windy or have other frequent disturbances for efficient spore dispersal. It tends to grow in alkaline, salty soil abundant in decaying organic materials and calcium carbonate, and it grows best at temperatures around 25 degrees Celsius. In its saprobic phase, C. posadasii mycelium can generally be found 20 cm below the surface, not at the top of the soil. It is hard to find in the winter and spring but generally reappears during the summer. Spore dispersal, and therefore peak infection rates, occur from September to November. Oftentimes the fungus can be found inhabiting rodent burrows, where it infects the animals and can in turn be spread to human populations.
As has been described, C. immitis is almost exactly identical to C. posadasii in its saprobic form, its parasitic form, and its clinical presentation. C. posadasii and C. immitis are both true human-pathogenic fungi, which means that they have a multinucleate “yeast phase” inside the human body. The other true human pathogens, which also attack the lungs and are found predominantly in the Americas, include the following:
1. Histoplasma capsulatum: The cause of histoplasmosis, this fungus is also a thermal dimorph with a filamentous hyphal form at about 25 degrees Celsius (white and brown in color) and a yeast form at 37 degrees Celsius.
2. Blastomyces dermatitidis: The cause of blastomycosis, also known as Chicago disease and Namekagon River fever.
3. Paracoccidioides brasiliensis: The cause of paracoccidioidomycosis
These can be told apart from C. posadasii because they form yeast when within the human body, which can be visualized in the sputum, and because they all have a sexual phase. Of these three, H. capsulatum is the most like C. posadasii, although in culture the hyphal phase of the former is more white than the latter.
As was previously mentioned, C. posadasii is a human pathogen that can cause serious health problems if left untreated. In addition to the 10%-40% prevalence of the disease in Mexico, approximately 100,000 primary infections occur in humans annually in the United States. In 60% of cases, there are no symptoms, and infection can only be detected by means of a skin test. The other cases can be classified as either acute, chronic, or disseminated. The acute form is rare, found only 3% of the time, and requires little to no treatment. In 5% of cases, the patients will develop a chronic version of coccidioidomycosis, which can occur even 20 years after initial infection. In this form of the disease, lung abscesses form, and when they rupture, the can fill the pleural space with pus, a very dangerous condition. The disseminated form occurs when the infection spreads to other tissues in the body and can cause meningeal infection, the most dangerous form of the disease. This occurs 0.15% to 0.75% of the time, and can often be life-threatening, especially when in concurrence with tuberculosis, as is often the case in areas with high prevalence of the latter. In immunocompromised patients, the disease can progress to these dangerous stages very rapidly.
There are three stages of infection by C. posadasii:
1. Spherule Initiation: In this stage, the arthroconidium is beginning its transformation into a spherule. It becomes rounder as its nucleus divides rapidly, producing a multinucleate cell with an empty central vacuole. These spherules can be visualized in and cultured from the sputum.
2. Spherule Segmentation: At this stage, the spherule begins to form endospores by means of internal segmentation, compartmentalizing each nucleus into its own endospore. This internal segmentation is accomplished by invagination of the spherule wall and incorporation of chitin. Segmentation begins 72 hours after initiation.
3. Endospore Release: The spherule ruptures and releases all 200-300 endospores either back lungs or into the lymphatic system and/or the bloodstream. This usually happens 120 to 132 hours after infection.
Symptoms of coccidioidomycosis include blood-tinged sputum, mental symptoms, mild to sever chest pain, chills, cough, fever, headache, join stiffness, loss of appetite, muscle aches and stiffness, night sweats, erythema nodosum (painful red nodules on the lower legs), sensitivity to light, excessive sweating, weight loss, and weezing. Less common symptoms include swelling of the ankles and feet, arthritis, enlarged or drained lymph nodes, and joint pain and swelling. It takes approximately seven to twenty-one days after initial infection for these symptoms to appear, if they ever do. Diagnosis can occur by means of a CBC blood test, sputum analysis, a coccidioidin skin test, or a chest x-ray. Using the latter, however, it is difficult to tell apart coccidioidomycosis and tuberculosis. For acute infections, bed rest and treatment similar to that of flu symptoms is sufficient. For worse cases, however, antifungals such as amphotericin B, ketoconazole, fluconazole, or itraconazole are used.
Thus, infection by C. posadasii can not only be uncomfortable but also life threatening. Pregnant women, African Americans, Filipinos, patients with HIV, Hodgkin’s disease, lymphoma, or any other immunosuppressing diseases, diabetics, people who have undergone organ transplants, and people undergoing adrenal corticosteroid therapy are all at a much higher risk of infection than normal. Because of its potency, C. posadasii has very real capabilities as a biological weapon, and as a result, there are very strict rules set in place internationally governing its possession and use.
Coccidioides posadasii is a pathogenic fungus that, along with Coccidioides immitis, is the causative agent of coccidioidomycosis,[1] or valley fever in humans. It resides in the soil in certain parts of the Southwestern United States, northern Mexico, and some other areas in the Americas, but its evolution was connected to its animal hosts.[2]
C. posadasii and C. immitis are morphologically identical, but genetically and epidemiologically distinct.[3] C. posadasii was identified as a separate species other than C. immitis in 2002 after a phylogenetic analysis.[4] The two species can be distinguished by DNA polymorphisms and different rates of growth in the presence of high salt concentrations: C. posadasii grows more slowly. It also differs epidemiologically, since it is found outside the San Joaquin Valley. Unlike C. immitis, which is geographically largely limited to California, C. posadasii can also be found in northern Mexico and South America.
As an intern in Buenos Aires in 1892, Alejandro Posadas described an Argentine soldier that had a dermatological problem since 1889. Posadas had seen the patient while a medical student in 1891 and skin biopsies revealed organisms resembling the protozoan Coccidia. The patient died in 1898 but during the interim Posadas successfully transmitted the infection to a dog, a cat, and a monkey, by inoculating them with material from his patient.
In 1899 a 40 year old manual laborer from the San Joaquin Valley, a native of the Azores, entered a San Francisco hospital with fungating lesions similar to those of Posadas' patient. Dr. Emmet Rixford,[5] a surgeon at San Francisco's Cooper Medical College, in attempts to determine the cause, concluded it was not from inadvertent self-inoculation. Further research produced a chronic ulcer in a rabbit and a lesion in a dog both excreting pus with the same organisms. Rixford issued a report, co-authored by Dr. Thomas Caspar Gilchrist (1862-1927),[6] that was printed in 1896, one year after the patient died. A pathologist at Johns Hopkins Medical School and Gilchrist studied the material and determined the microbe was not a fungus but a protozoan resembling Coccidia. With the help of parasitologist C.W. Stiles, the organism was named Coccidioides (“resembling Coccidia”) immitis (“not mild”). Four years later William Ophüls and Herbert C. Moffitt proved that C. immitis was not a protozoan but was a fungus that existed in 2 forms. In 1905 Ophüls called the infections "coccidioidal granuloma" and that it could develop from inhalation of the organism. Also in 1905 Samuel Darling studied a case and, referring to the misnamed organism a protozoan, named it Histoplasma capsulatum, meaning three major endemic fungi in the United States were all initially misidentified as protozoa.
Studies by Cooke on the immunology of the disease, and in 1927 a filtrate of culture specimens, later named coccidioidin, began to be used in skin testing to delineate the epidemiology of infection. In 1929 a second-year medical student, Harold Chope, was studying C. immitis in the laboratory of Ernest Dickson at Stanford University Medical School, and breathed in spores becoming infected but he later recovered. In 1934 Myrnie Gifford, a physician at San Francisco General Hospital, joined the Health Department of Kern County, California.[7] She had observed that San Joaquin Valley Fever patients often suffered from erythema nodosum, and all tested positive for coccidioidomycosis.[8] She met Ernest County when he visited her in Kern County, California, and together they presented evidence to the California Medical Association.[7] The two determined that San Joaquin fever represented C. immitis infection. The Kern County Health Department began obtaining epidemiologic histories and skin testing all cases involving Valley Fever. The investigations revealed, among other things, that a majority of the cases described a history of dust exposure, that coccidioidomycosis was common in the area, and that racial differences determined the host's response to the fungus.
Chope left Stanford Medical School and Dickson recruited a classmate, Charles E. Smith, to replace him. Smith began an extensive 17-month study of coccidioidomycosis in Kern and Tulare County, that also began a lifelong professional focus of study of C. immitis and coccidioidomycosis, even after he became Dean of the School of Public Health at the University of California at Berkeley in 1951, until his death in 1967. Research by Smith resulted in more than a few discoveries that included serologic testing, that chlamydospores of the fungus c. immitis could be wind-blown dispersing the spores when the hot weather converted the soil to dust, scientific results of military personnel testing in the southern San Joaquin Valley before and during WWII, as well as people of Japanese descent (many US citizens) interned in camps, prisoners of war, and agricultural workers. Diagnoses of active disease and skin testing, showed that it was also found in southern Nevada and Utah, western Texas, as well as Arizona, where the southern and central areas appeared to impose the highest risk of infection in the United States. An important Smith's research added to the fundamental discoveries of microbiology, epidemiology, clinical findings, and diagnosis that had emerged since Posadas' initial case report in 1892.[9]
Progress in studies from 1997 to 2007, including genomic restriction fragment length polymorphism (RFLP) concluded that there were two separate species. Earlier the two were referred to as types I and II, and later as Non-California and California distributions, determined as clades through microsatellite analyses. Genealogical Concordance Phylogenetic Species Recognition (GCPSR) criteria were met, so the two entities were proposed and generally recognized as two separate species: Coccidioides immitis, and the novel species Coccidioides posadasii.[10]
Coccidioides posadasii is a pathogenic fungus that, along with Coccidioides immitis, is the causative agent of coccidioidomycosis, or valley fever in humans. It resides in the soil in certain parts of the Southwestern United States, northern Mexico, and some other areas in the Americas, but its evolution was connected to its animal hosts.
C. posadasii and C. immitis are morphologically identical, but genetically and epidemiologically distinct. C. posadasii was identified as a separate species other than C. immitis in 2002 after a phylogenetic analysis. The two species can be distinguished by DNA polymorphisms and different rates of growth in the presence of high salt concentrations: C. posadasii grows more slowly. It also differs epidemiologically, since it is found outside the San Joaquin Valley. Unlike C. immitis, which is geographically largely limited to California, C. posadasii can also be found in northern Mexico and South America.
Coccidioides posadasii ialah kulat patogen yang, bersama dengan Coccidioides immitis, merupakan agen penyebab bagi koksidioidomikosis,[1] atau demam lembah pada manusia. Ia hidup di dalam tanah di kawasan-kawasan tertentu di Amerika Syarikat Barat Daya, di Mexico utara, dan di sesetengah kawasan lain di Benua Amerika, tetapi evolusinya dihubungkan dengan haiwan-haiwan hosnya.[2]
C. posadasii dan C. immitis adalah serupa secara morfologi, tetapi berbeza secara genetik dan secara epidemiologi.[3] C. posadasii dikenal pasti sebagai spesies terpisah selain C. immitis pada 2002 selepas analisis filogenetik.[4] Kedua-dua spesies ini boleh dibezakan dengan polimorfisme DNA dan kadar pertumbuhan berlainan pada kehadiran kepekatan garam tinggi: C. posadasii tumbuh dengan lebih perlahan. Ia juga berbeza secara epidemiologi, oleh sebab ia dijumpai di luar Lembah San Joaquin. Tidak seperti C. immitis, yang terhad secara geografi kebanyakannya di California, C. posadasii juga boleh dijumpai di Mexico utara dan di Amerika Selatan.
Coccidioides posadasii ialah kulat patogen yang, bersama dengan Coccidioides immitis, merupakan agen penyebab bagi koksidioidomikosis, atau demam lembah pada manusia. Ia hidup di dalam tanah di kawasan-kawasan tertentu di Amerika Syarikat Barat Daya, di Mexico utara, dan di sesetengah kawasan lain di Benua Amerika, tetapi evolusinya dihubungkan dengan haiwan-haiwan hosnya.
C. posadasii dan C. immitis adalah serupa secara morfologi, tetapi berbeza secara genetik dan secara epidemiologi. C. posadasii dikenal pasti sebagai spesies terpisah selain C. immitis pada 2002 selepas analisis filogenetik. Kedua-dua spesies ini boleh dibezakan dengan polimorfisme DNA dan kadar pertumbuhan berlainan pada kehadiran kepekatan garam tinggi: C. posadasii tumbuh dengan lebih perlahan. Ia juga berbeza secara epidemiologi, oleh sebab ia dijumpai di luar Lembah San Joaquin. Tidak seperti C. immitis, yang terhad secara geografi kebanyakannya di California, C. posadasii juga boleh dijumpai di Mexico utara dan di Amerika Selatan.
Coccidioides posadasii é um fungo dimórfico, que assim como o Coccidioides immitis, causa coccidioidomicose em seres humanos. É endêmico no semi-árido do nordeste brasileiro, centro de Argentina, sul da Bolívia, norte do México e sudoeste dos EUA.[1]
Crescem bem em solo semi-árido, alcalino, salgado, e dispersam artroconídios que infectam as vias respiratórias baixas. Em meios de cultivo típicos para fungos à temperatura ambiente, crescem como micélio vegetativo hialino, formando colônias esbranquiçadas de aspecto algodonoso. Dentro do homem, por outro lado, assumem formas atípicas esféricas ou ovais, não brotante, de paredes espessas, de 5 a 60 µm de diâmetroe, que quando maduro contem dezenas de endósporos globosos e uninucleados, com 2-5 µm de diâmetro..[2]
A expectoração é observada ao microscópio e coloram bem com H&E, ácido periódico de Schiff ou impregnação argêntea de Gomori-Grocott, mas a cultura em cabine de segurança biológica 3 pode ser necessária. Também pode ser diagnosticada com PCR.[3]
A coccidioidomicose, também conhecida como (febre do vale ou reumatismo do deserto), é uma micose sistêmica causada pela inspiração dos esporos, que geralmente é leve e cura mesmo sem tratamento. Em pessoas com doenças crônicas podem formar granulomas que parecem câncer de pulmão ou tuberculose em exames de imagem. É necessário uma biópsia e cultivo dos fungos para identificá-lo. Em imunossuprimidos pode complicar causando pneumonia atípica grave ou fungemia. [2]
Coccidioides posadasii é um fungo dimórfico, que assim como o Coccidioides immitis, causa coccidioidomicose em seres humanos. É endêmico no semi-árido do nordeste brasileiro, centro de Argentina, sul da Bolívia, norte do México e sudoeste dos EUA.