Comprehensive Description
(
anglais
)
fourni par Smithsonian Contributions to Zoology
Dendrocoelum Örsted
The dendrocoelids of Lake Ohrid were first placed into a new genus, Neodendrocoelum, established by Komárek (1926a:7), who listed among their distinguishing characteristics the large size of the adenodactyl (which is larger than the penis) and the configuration of the atrial cavity. This cavity consists of two compartments: a long cavity named “atrum genitale,” which contains the papilla of the penis and receives the mouth of the common oviduct in its posterior part; and a compartment designated as “atrium copulativum,” enclosing the papilla of the adenodactyl and connecting with the opening of the bursal stalk. These two cavities unite only near the genital pore. No “atrium commune” is developed. The species have one pair of eyes and are either white or pigmented.
Stankovi and Komárek (1927:596–598) mentioned the same characteristics of the genus and added that the penis has a wide lumen and that its papilla may be invaginated, resembling a flagellum.
Kenk (1930:300) included Neodendrocoelum in the genus Dendrocoelum and distinguished only two subgenera, Dendocoelum sensu stricto and Paradendrocoelum, which differ in the course of their oviducts.
Later authors have not accepted Kenk's subdivision of the genus Dendrocoelum. De Beauchamp (1931:157; 1932:207–208), who studied the genus most intensively based on copious material, divided it into eight subgenera. He accepted Neodendrocoelum as a subgenus of Dendrocoelum after some hesitation, but stated that it was not clearly separable from the subgenus Dendrocoelides. He added to the characteristics of the subgenus the presence of a glandular field around the genital opening and peculiarities in the histology of the oviducts. He excluded one of the Ohrid species, N. jablanicense, from Neodendrocoelum and placed it in the subgenus Eudendrocoelum with a question mark. Gourbault (1972), the latest reviser of the genus Dendrocoelum, followed essentially de Beauchamp's arrangement.
Komárek (1953a:303) accepted the status of Neodendrocoelum as a subgenus of Dendrocoelum and also included Dendrocoelum subterraneum, a blind species described earlier from a cave in Croatia.
Stankovi (1960:178–179, 263), in evaluating the taxonomic rank of Neodendrocoelum, repeated the previously enumerated characters of the taxon and considered it “more prudent to keep Neodendrocoelum provisorily as a distinct genus precisely because of its peculiar geographic distribution.”
Reisinger (1971:117–119) discussed the interrelationships of the various genera or subgenera of the Dendrocoelum group. He came to no conclusion about the definite rank of Neodendrocoelum, but was inclined to consider it an acceptable genus.
It is very difficult to establish clear criteria for the demarcation of Neodendrocoelum from other species groups of the Dendrocoelum complex. The division of the genital atrium into two compartments, the genital and copulatory atria, is seen also in other members of the subgenus Dendrocoelides, where the cavities are generally designated as the male and the common atria. The relative sizes of penis and adenodactyl are subject to great variation within the genus and cannot be used as an important systematic character. The glandular field around the gonopore may be seen also in other species of the genus, perhaps to a lesser degree, and it is also not prominent in all Ohrid species.
In view of this complex situation, I prefer to leave the question of the validity of Neodendrocoelum, even as a subgenus, open to further investigations. New criteria may possibly be found in the analysis of the chromosome morphology or in biochemical studies of the species concerned.
As has been pointed out very appropriately by Komárek (1953a:303; 1953b:269), the copulatory apparatus of a great number of the species of Dendrocoelum of the Ohrid region exhibits a surprising conformity, which would make it very difficult to identify the individual species by its anatomy alone. Variations within the apparatus are often only of a quantitative nature or are due to various states of muscular contraction, stages of sexual maturity, or employment of different fixing agents. It is, therefore, necessary to look for other characters as well, such as the general proportions of the body, the exact shape of the anterior end, the situation and arrangement of the testicles, the development of the subepidermal musculature, the behavior of the animal in life, and the pattern of pigmentation. Pigmentation is generally considered to be a rather variable and taxonomically not very reliable character. Nevertheless, it is the most obvious and most easily recognizable feature of the Ohrid species.
Species that were available to former workers and to the present writer in sufficient numbers to be observed and analyzed intimately are undoubtedly good species. Among these I would count Dendrocoelum maculatum, D. sanctinaumi, D. jablanicense, D. magnum, D. ochridense, D. cruciferum, D. lacustre, and D. translucidum. Of the unpigmented white species, D. adenodactylosum is obviously very near to D. nausicaae, which is more generally distributed in southwestern Europe but may be distinguished by karyological characters, according to Stankovicacute; (1969:427). Dendrocoelum jablanicense clearly differs from the rest of the Ohrid dendrocoelids by the deviating anatomy of its copulatory complex. The whitish D. albidum and D. sinisai differ from all other species of the Ohrid area by having a distinctive invaginated adhesive organ. Their mutual relationship, however, needs to be investigated on more material. Among the pigmented species of the lake, several forms inhabiting the sublittoral and profundal zones show a variable number of light spots arranged in a pair of rows on the dorsal surfaces, but have been examined in only a few specimens. They differ from each other chiefly by the number and outline of the spots. As we lack sufficient information on the effects of environmental parameters on the color pattern of these forms, it is possible that they may in part be referable to species occurring in the littoral or in the tributaries of the lake. Among these are D. komareki, D. decoratum, D. dorsivittatum, “Neodendrocoelum sp. 3” of Stankovi (1955a, pl. 5: fig. 7), and a few other forms observed occasionally but not described in this paper. Two other species, D. cruciferum and D. lacustre, both from the shell zone of the lake, are somewhat similar in their color pattern, consisting of a median pigment stripe and a pair of dark spots on the dorsal surface. They differ, however, so markedly in the shape of the anterior end, their size, and behavior that their specific identity cannot be doubted.
Some of these open questions may be solvable by karyological, by biochemical studies, or by an investigation of the possibility of interbreeding of the forms concerned.
Dendrocoelum adenodactylosum (Stankovi and Komárek)
Neodendrocoelum adenodactylosum Stankovi and Komárek, 1927:599.
Neodendocelum nausicae.—Stankovi, 1930:168 [in part].
Dendrocoelum (Neodenrocoelum) adenodactylosum.—de Beauchamp, 1931:157.
MATERIAL DEPOSITED.—Sagittal and transverse serial sections of 7 specimens on 22 slides, USNM 55253–55259.
Dendrocoelum adenodactylosum is the most common triclad of the Ohrid region, occurring both in Lake Ohrid and in its, tributary streams and sources. It was first described by Stankovi and Komárek (1927:599–603). In later publications, beginning with 1930, Stankovi considered the species to be identical with D. nausicaae O. Schmidt (1861), a species originally discovered in several localities on two Ionian islands, Corfu (Kerkyra) and Cephalonia (Kephallenia), off the west coast of Greece. Schmidt's figures and anatomical analysis of the copulatory organs of his species were surprisingly accurate for his time, considering that the observations were made on whole animals. Many years later, Wilhelmi (1909:4) collected additional specimens from Schmidt's original localities, which were examined by Komárek (1925:326–328), Stankovi and Komárek (1927:617–620), and by de Beauchamp (1932:219–222). Komárek (1953a:305) himself collected the species again on the island of Corfu; D. nausicaae proved to be widely distributed in southeastern Europe (Balkan Peninsula, Austria, Italy) and even in Asia Minor. De Beauchamp (1932:208, 219) treated D. adenodactylosum and D. nausicaae as separate species and, in a later paper (1937:357), pointed out the differences between them (see also Gourbault, 1972). More recently, Stankovi (1969:427) came to the same conclusion, chiefly on the basis of differences in the chromosome numbers of the two forms. Reisinger (1971:133) also considered D. adenodactylosum to be a separate species, though closely related to D. nausicaae.
EXTERNAL FEATURES (Figures 5, 24).—The shape of the animal in life was discussed by Stankovi and Komárek. The species is unpigmented, white, and its appearance is similar to that of Dendrocoelum lacteum. According to Stankovi, the animals attain a length of 20 mm and a width of 3.5–5 mm. De Beauchamp reported that specimens from šum measured less than 10 mm, but this obviously refers to preserved worms. I have seen sexually mature specimens that in life were only 16 mm long and 2.5 mm wide. The anterior end is truncated, the frontal margin bulging anteriorly. To either side of the frontal margin is a rounded lobe, projecting somewhat laterally. When the animal glides along undisturbed, a faint, but usually well discernible, narrowing or neck is seen, separating the head from the rest of the body.
There are normally two eyes, situated approximately at the level of the neck. The distance between them is slightly more than one-third the width of the neck and their distance from the frontal margin is greater than that from the lateral margins. Supernumerary eyes are not rare and may occur anteriorly as well as posteriorly to the principal eyes. Occasionally one finds specimens that have up to four eyes on each side, arranged in a curved longitudinal row (Figure 26), as has already been observed by de Beauchamp (1937:357).
The intestine is usually well visible in the living animal, particularly from the ventral side. The anterior ramus reaches up to the level of the eyes and bears 9–12 branches on each side. The two posterior rami, with up to 18 branches each, usually unite behind the copulatory apparatus.
The pharynx is inserted at about the middle of the body in sexually mature specimens, its length amounting to one-seventh the body length. The copulatory apparatus occupies the anterior half of the postpharyngeal region.
The animal moves generally by gliding. Upon stimulation, a “crawling” locomotion may be induced, in which the subterminal adhesive organ is used for successive attachment to the substrate. This crawling is less jerky than that of D. lacteum and is sustained only for a short time. During crawling and when the animal is at rest, the lateral margins of the body form wavy folds.
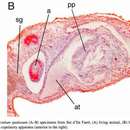
ANATOMY.—The anterior adhesive organ is moderately developed. It is situated in the midline on the ventral side of the frontal margin and consists of a well-circumscribed area of infranculeate epithelial cells pierced by eosinophilic gland ducts. Fibers of the ventral subepidermal longitudinal muscle layer attach to the adhesive area, but no special muscular differentiations are developed. The organ is larger than that of D. jablanicense, but far less conspicuous than the adhesive cushion of D. lacteum. The median diameter of the adhesive surface is about 150 μm.
The anatomy and histology of the reproductive system of D. adenodactylosum has been discussed in detail by Stankovi and Komárek (1927:599–602) and by de Beauchamp (1932:208–211; 1937:357). I am giving here only the essential features of this system and am adding figures of the copulatory apparatus that will show the appearance of this complex in various states of contraction and relaxation of its organs. The testicles are numerous and essentially ventral. They occupy a broad zone on either side, which extends from the level of the ovaries to close to the posterior end. The widened anterior end or tuba of each oviduct, adjoining the ovary and functioning as a seminal receptacle, appears at full maturity as a large rounded sac with a tall epithelial lining, the tubal bursa, quite similar to that described by de Beauchamp (1932: 212) for D. maculatum (see also Reisinger, 1963: 684). This tubal bursa serves for the resorption of superfluous sperm and is found also in other species of dendrocoelids of Lake Ohrid.
In the copulatory apparatus (Figure 35), the adenodactyl (ad) is very large and situated to the left of the midline; the penis is smaller and located on the right side and rather far dorsal. The size and shape of the atria may vary considerably due to the state of contraction or expansion of the adenodactyl. The male atrium, designated by Stankovi and Komárek as the genital atrium, is rather long and is lined with a thick epithelium, the inner border of which is packed with eosinophilic secretion granules. The penis may assume different shapes. In the majority of the preparations it appears as shown in Figure 35: a penis bulb of moderate size, enclosing a slightly widened cavity with a thick epithelial lining, the seminal vesicle (vs), receiving close to its anterior end the separate openings of the vasa deferentia (vd); the penis papilla shows a more or less distinct division into two parts, a basal part with thick walls, covered by a rather tall surface epithelium, beneath which is a strong layer of circular muscles; and a distal, thin-walled part with hardly any muscle fibers, containing a usually rather wide lumen. In other specimens the penis may be completely extended (Figure 52A; see also de Beauchamp, 1932, fig. xxiv): in this case the bulb is extremely contracted, the seminal vesicle pushed into the base of the papilla, the papilla itself appearing tube shaped, tapering in diameter toward its tip. In the other extreme (Figure 52B), a complete introversion of the papilla into the bulb may occur: the bulb then appears expanded, its wall rather thin, its lumen voluminous, the openings of the vasa deferentia placed far apart, the papilla assuming the shape of an inverted pseudoflagellum. The statement by Reisinger (1971: 133) that D. adenodactylosum differs from D. nausicaae by the fact that it normally (in life) has no invaginated pseudoflagellum, while D. nausicaae regularly shows such an invagination, may be erroneous.
The two oviducts unite behind the copulatory apparatus after embracing the bursal duct. The common oviduct (odc) thus formed is rather short and opens into the caudal part of the male atrium. The bursa (b) and the bursal stalk (bd) show no peculiarities. De Beauchamp observed a connection of the bursa with intestinal caeca that approach it from both sides. He did not see any continuity of the cavities of the bursa and the caeca. Nevertheless he interpreted these connections as virtual genitointestinal communication. I have repeatedly seen intestinal branches approaching or even touching the bursa laterally. This is, however, by no means a constant feature. I would hesitate to consider this connection a functioning communication between bursa and intestine. The bursal duct (bd) proceeds from the bursa posteriorly above or slightly to the left of the penis and gradually widens, then turns ventrally and opens into the common atrium (Komárek's “copulatory atrium”). The anterior part has a very fine cover of muscle fibers, which thickens in the posterior part and becomes rather conspicuous just before the mouth of the duct. This terminal portion of the muscle coat, consisting principally of circular fibers, is the “sphincter” mentioned by de Beauchamp (1932: 211:1937:357).
The adenodactyl (ad) is unusually large and contains a long lumen that may expand to a cavity of irregular outline in the bulb of the organ. Its shape varies from a long, slender, somewhat curved organ to a pear-shaped one with almost spherical bulb. It is obvious that these different shapes depend mainly on the state of contraction of its muscular systems. The papilla is always bent but may be pointing in various directions. When the organ is elongated, the papilla protrudes posteriorly and extends the wall of the common atrium abnormally (see Stankovi and Komárek, 1927, text-figure 2); in other cases it is inserted into the end portion of the bursal duct (Figure 35), protrudes through the genital aperature (de Beauchamp, 1932, fig. xxiv), or may penetrate into the male atrium.
The glandular field of tall epidermal cells surrounding the gonopore, considered by de Beauchamp (1931:158) to be a distinguishing characteristic of the subgenus Neodendrocoelum, is well differentiated at full sexual maturity.
DISTRIBUTION AND ECOLOGY.—Dendrocoelum adenodactylosum is a very common inhabitant of cold springs and streams of the Ohrid region. I collected it in the spings of Studenište, Bej-Bunar, Sveti Naum and Šum and in Kulinivljik na Sinoru, a small stream near the western border of the town of Ohrid. Stankovi and de Beauchamp likewise obtained most of their animals from springs, and expressly mention Studenište, Sveti Naum, Šum, and Gradište, the latter being a small tributary of Drim River (which is the outlet of Lake Ohrid). Stankovi and Komárek reported that the species is common also in the littoral zone of the lake. In a later paper, Stankovi (1934:176) stated that the littoral animals are usually asexual during the summer. I found, however, both mature and immature specimens in the littoral zone, particularly among the dense vegetation of reeds near the Hydrobiological Station. Stankovi and Komárek also reported the species from the profundal zone at a depth of 108 m. It is highly probable that this record concerns D. sinisai rather than D. adenodactlyosum. The latter species occurs also in Lake Prespa, a nearby lake southeast of Lake Ohrid (Stankovi, 1969:428).
Dendrocoelum adenodactylosum can easily be kept in the laboratory. Beef liver is readily taken as food. In one culture, cocoons (spherical and unstalked) were deposited and young animals hatched.
TAXONOMIC POSITION.—The history of the discovery of D. adenodactylosum has been discussed in the introductory paragraph to the species. Its close relationship to D. nausicaae is undeniable (see also Komárek, 1953a:305–306). Nevertheless, I agree with the opinion of de Beauchamp and of Reisinger: of keeping the two forms as separate species, particularly because of the karyological differences between them, as mentioned by Stankovi (1969: 427). In a recent paper, Paunovi (1977) established the chromosome number of D. adenodactylosum to be 2n = 32, while that of D. nausicaae is 2n = 14.
Dendrocoelum maculatum (Stankovi and Komárek)
Neodendrocoelum maculatum Stankovi and Komárek, 1927: 603.
Neodendrocoelum spec. (immaturum I) Stankovi and Komárek, 1927:621.
Dendrocoelum (Dendrocoelum) maculatum.—Kenk, 1930:301. Dendrocoelum (Neodendrocoelum) maculatum.—de Beauchamp, 1931:157.
Dendrocoelum maculatum.—Kenk, 1974:18.
MATERIAL DEPOSITED.—Sagittal and transverse serial sections of 6 specimens on 24 slides, USNM 55272–55277.
EXTERNAL FEATURES (Figure 2).—Illustrations of the external aspects of the species in life were given by Stankovi and Komárek (1927:604) and by Stankovi (1955a, pl. 4: fig. 1). Mature animals are 15 to 25 mm long and 3 to 4 mm wide. During undisturbed gliding, the frontal margin of the head shows three distinct lobes: a broad, curved, median lobe and a pair of smaller, rounded, lateral lobes well marked off from the median lobe. Behind the head, a slight constriction or neck is visible. Posteriorly to this, the lateral margins of the body diverge gradually until the greatest width is attained at about the middle of the body length or somewhat behind it, then converge again in the last quarter of the body and meet at the moderately pointed posterior end. In gliding locomotion, the margins are smooth, but during crawling movement they are thrown into folds and ruffles. The shape of the anterior end also changes constantly in the crawling animal, the median lobe being now retracted, now excessively protruded.
The two eyes are situated far apart, their distance from each other amounting to about one-half the width of the head at the level of the eyes. The mouth is situated somewhat behind the middle of the body and the genital aperture is closer to the mouth than to the tail end.
The most distinguished character of the species is the pattern of its coloration. The dorsal surface is a variable shade of brown (olive brown, reddish brown, chocolate brown, or almost black) with a lighter (yellowish or grayish) pattern. The light color forms a narrow rim along the lateral margins, except on the lateral lobes of the head; a triangular field on the head extends from the middle lobe backward beyond the level of the eyes; there are characteristic spots to each side of the midline in a longitudinal row of 6 to 8 that have an irregular lobed and branched outline. In the gliding animal, these spots appear elongated longitudinally. They frequently show a tendency to fuse to a netlike pattern. A pair of indistinct light dashes extends from the lateral lobes of the head obliquely backward and medially, which probably corresponds to the location of the auricular sense organs. Usually there is also a more or less marked light band in the middorsal line. On closer examination, it consists of a double row of small elongated splashes along the midline of the prepharyngeal region, a pale, somewhat broader spot above the pharynx and copulatory complex, and a distinct median band in the hindmost part of the body. The ventral surface is likewise pigmented, but much lighter, almost uniformly yellow or yellowish brown, with a lighter area below the pharynx and another surrounding the genital pore.
In young animals, the pattern of the dorsal surface is simpler, the lateral spots being less branched, often almost round. The shape of the anterior end, however, is typical and the characteristic light triangular field on the head is always distinct.
ANATOMY.—The pharynx is relatively long, about one-seventh the body length. The adhesive cushion, situated on the underside of the median frontal lobe, is a rather large field of modified epithelium with eosinophilic gland ducts, measuring 180–290 μm in median diameter.
The reproductive system of Dendrocoelum maculatum has been described well by Stankovi and Komárek (1927:603–605) and de Beauchamp (1932:212–215). I shall give here only the essential characters of the system.
The numerous small testes are located both dorsally and ventrally to the intestine and at immediate levels between the intestinal branches. The testicular zone extends from the region of the ovaries backward close to the posterior end.
The genital pore (Figure 36, gp) is surrounded by an area of very tall epithelial cells, characteristic of several related species of the Ohrid region. It measures 36–60 μm in thickness, while the general epithelium of the ventral surface is 17 μm and that of the dorsal surface 25 μm thick. The cells in this area lack rhabdites and are pierced by a great number of parallel gland ducts (gl), the cell bodies of which are situated in the mesenchyme of the ventral side, above the layers of the integumental muscles of the genital region. In preparations fixed by de Beauchamp's fluid (ethanol, formalin, and acetic acid), the secretions of the glands are dissolved and the gland ducts appear empty. Worms killed with mercuric chloride solution show the secretion in the canals to be homogeneous, staining pink with erythrosin. It is difficult to establish the homology of these peculiar glands, as de Beauchamp (1932:219) points out. They seem to correspond to glands found in various triclads and designated by Meixner (1928) as “Kittdrüsen” (cement glands), though this name might properly be restricted to glands that empty into the common atrium.
The genital pore leads into the common atrium (ac), which encloses the free papilla of the adenodactyl and connects with the stalk (bd) of the copulatory bursa and with the more or less elongated male atrium (am).
The penis lies to the right side of the midline. Its bulb is large, but not very muscular. When extended, the penis papilla is conical and somewhat irregular in outline (Figure 36). It is covered by a cubical or columnar epithelium pierced by numerous gland ducts that carry a grandular secretion staining in various shades of red with erythrosin. Beneath the epithelium is a layer of circular muscle fibers, strong and thick in the basal half of the papilla, but very thin in the distal part. The wide cavity in the penis bulb, the seminal vesicle (vs), is lined with an epithelium that forms villuslike projections. The two vasa deferentia (vd) open separately into the anterior part of the vesicle from the sides. The cavity continues into the penis papilla, forming a wide canal with uneven outline that opens at the tip of the papilla. Many gland ducts (pink after staining with erythrosin) empty into the penis lumen, at least into its anterior part. The penis papilla can be partly invaginated into the penis lumen. Then the penis bulb appears to be distended and the seminal vesicle very voluminous (Figure 54). Only the distal part of the papilla, that is the part lacking the thick muscle layer, is inversible, while the basal portion remains protruding into the atrium.
The two oviducts unite behind the copulatory apparatus, embracing the bursal duct. The long common oviduct (odc) runs, to the right side of the bursal stalk, almost horizontally forward and opens into the end part of the “male” atrium.
The copulatory bursa (b) is a large sac extending far laterally (however, not occupying the entire width of the body, as Stankovi and Komárek indicate). The ratio between its transverse and anteroposterior diameters is approximately 3:1. The bursa is often lobate, with irregular outline. De Beauchamp stated that a pair of outgrowths of the bursa approach the vasa deferentia; these outgrowths, however, are not always present. In one case he also observed an open communication between the sperm ducts and the bursa, through which sperm were passing. I have never seen such a communication in my preparations and must assume that what de Beauchamp saw was an accidental rupture of the walls of the two organs.
The bursal stalk or duct (bd) starts from the posterior wall of the bursa as a rather narrow canal situated to the left of the penis, then widens transversally to a flat sac above the common atrium. From this flattened portion, a short wide canal proceeds ventrally and opens through the roof of the common atrium. This canal is surrounded by a thick layer of muscle fibers. The widening of the bursal stalk and the sharp bend it undergoes above the common atrium are particularly characteristic of the species.
The adenodactyl (ad) is large and usually exceeds the penis in size. It lies to the left side of the midline.
DISTRIBUTION AND ECOLOGY.—Dendrocoelum maculatum was collected by Stankovi and Komárek in several tributary streams of Lake Ohrid (at Studenište and Sveti Naum) and in the large spring Šum (west of Struga). De Beauchamp examined animals from springs near Sveti Naum and near Tušeimište (Albania). Immature specimens were reported from the littoral of the lake, particularly in the Bay of Ohrid. My own material was collected in Bej-Bunar, Studenište, Sveti Naum, and Šum. In these cold habitats, the worms may be found in summer and probably the year round in all stages of development. They are more numerous in the quiet parts of the streams and rare or entirely absent in the springs themselves. The species is also very common in the littoral zone of the lake, under stones, among algae, and on high plants; however, no sexually mature animals can be found there during the summer months or in early autumn. As the color pattern of the young specimens is somewhat different from that of the adults, their identity with the animals living in cold streams was not recognized at first by Stankovi and Komárek, who described the littoral form as “Neodendrocoelum spec.?–(immaturum I).” Mature specimens with typical coloration were found in the lake in winter, however (Stankovi, 1938:6). I kept several individuals of the immature shore form in the laboratory at low temperature and they developed the typical spotted pattern of the adults. There can be no doubt that the animals from the two types of habitat are identical.
I collected Dendrocoelum maculatum in the lake in shallow water near the town of Ohrid (water temperature in August, 24.7°C), near Studenište, near Velidab (in the vicinity of a submerged spring, 10.9°C), and at Kalište. De Beauchamp reports it also from Struga and Lin. It was occasionally found in the lower littoral (Chara zone) in Ohrid Bay, at a depth of 10 m, and at Kalište, 7–13 m.
Reisinger (1963:684–685) reports to have collected near Graz, Austria, a planarian that is anatomically identical with D. maculatum, particularly in the development of large tubal bursae and yolk funnels. The only difference from the Ohrid form is its complete lack of body pigment, though the eyes are developed normally. He referred to the species first as “Neodendrocoelum ad maculatum,” later as N. maculatum, and considered the lack of pigment to be no taxonomically valid characteristic. The identical species was reported later by An der Lan (1964:479) from the Danube River in Austria. If this form had been observed in the Ohrid area, it would undoubtedly have been described as a separate species, as it is well known that several species in that area can hardly be separated by anatomical characters (see Komárek,, 1953b:269), although they differ in their pigment patterns, behavior, and preferred type of habitats. In light of this situation, I would prefer to consider the unpigmented form preliminarily to be at least a separate subspecies for which I propose the name Dendrocoelum maculatum candidum. Further investigations will be necessary to determine its relation to the Ohrid form, such as a karyological analysis and, particularly, a study of the possibility of interbreeding of the two forms.
In the laboratory, Dendrocoelum maculatum may easily be kept in culture on a diet of beef liver. Cocoons laid in the aquaria were round or bluntly ellipsoidal and unstalked (smallest cocoon spherical, with a diameter of 1.7 mm; largest one ellipsoidal, with diameters 2.6 and 2.3 mm).
TAXONOMIC POSITION.—In the structure of the copulatory apparatus, Dendrocoelum maculatum is very similar to other pigmented species of the Ohrid region, D. ochridense and D. lacustre, which can easily be distinguished by their characteristic color patterns, body shape, some histological features, and behavior. From D. sanctinaumi, which shows a somewhat similar pigment design, D. maculatum differs by its slender proportions, the shape of the anterior end (the lateral lobes being marked off from the median lobe), the position of the eyes (farther apart), and by the shape of the dorsal spots. Preserved specimens of D. maculatum are usually elongated, frequently rolled in toward the ventral side, with the lateral margins ruffled, while in D. sanctinaumi the body remains broad and flat, with concave ventral and convex dorsal sides, and the body margins smooth (see also Stankovi and Komárek, 1927:609, and de Beauchamp, 1932:216). Paunovi and Rimsa (1968) and Paunovi (1977) investigated the chromosome set in the testes and ovaries of D. maculatum, and they determined the chromosome numbers to be n=16, 2n=32.
- citation bibliographique
- Kenk, Roman. 1978. "The planarians (Turbellaria, Tricladida Paludicola) of Lake Ohrid in Macedonia." Smithsonian Contributions to Zoology. 1-56. https://doi.org/10.5479/si.00810282.280