noms dans le fil d’Ariane
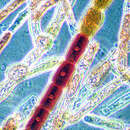
Rooiwiere (klas Rhodophyta) leef veraf in tropiese see en hul naam is afkomstig van hul rooi kleur, wat deur die aanwesigheid van 'n aantal uitsonderlike pigmente veroorsaak word. Hierdie pigmente kan selfs in swak lig fotosinteer. Voortplanting kan geslagtelik of ongeslagtelik wees en sommige spesies is ekonomies belangrik.
Die rooiwiere (klas Rhodophyta) het, soos die ander wiere, geen egte wortels, stingels en blare nie en die hele plantliggaam is uit homogene weefsel opgebou. So 'n struktuur word 'n tallus genoem, en hoewel blaaragtige en stingelagtige strukture by sommige vorme onderskei kan word, is dit nie gedifferensieerde organe nie. Kleurstowwe wat vir elke klas alge kenmerkend is, kom in die selle van die plante voor. In die geval van die rooiwiere word groen chlorofil d, asook rooi karoteen en fikobileen aangetref. Die twee kleurstowwe het die plante se kenmerkende rooi of violet kleur tot gevolg.
Die karoteen en fikobileen absorbeer lig vir fotosintese uit die blou deel van die spektrum, wat dit vir die rooiwiere moontlik maak om op groter dieptes (tot 75 m onder die seeoppervlak) as groenwiere te leef. Die meeste rooiwiere kom in tropiese waters voor en tydens fotosintese word die gevormde glukose omgesit in florideensetmeel, 'n stof wat naby verwant is aan dierlike glikogeen. Die selle van rooiwiere bevat ook 'n ingewikkelde koolhidraat, floridosied, wat ’n verbinding van gliserol en die monosakkaried galaktien is.
Die rooiwiere word in twee subklasse verdeel op grond van die groeiwyse van die tallus en die manier waarop die bevrugte eiersel (sigoot) ontwikkel. Die subklas Bangiophycidae bestaan uit oorwegend eensellige of draadvormige spesies wat nie duidelike groeisones het nie. Die selle het slegs een kern en het stervormige chromoplaste. Lede van die subklas Floridophycidae het gewoonlik 'n meersellige, vertakte tallus en die plante groei deurdat selfstandige selle aan die bopunt van die tallus verdeel. Die selle kan dikwels meerkernig wees en dit bevat ook 'n aantal skyfvormige chromoplaste, wat langs dieselwand gerangskik is.
Die geslagtelike voortplanting van rooiwiere toon ’n ingewikkelde generasiewisseling en selfs 3 generasies kan mekaar opvolg. Ongeslagtelike voortplanting kan by sommige spesies plaasvind deurdat een gedeelte van die tallus afsnoer en tot 'n nuwe individu ontwikkel.
Rooiwiere plant egter meestal voort deur ongeslagtelike spore, aplanospore, te vorm. Hierdie spore word afsonderlik, in pare of in veelvoude van vier in sporangia gevorm. Geslagtelike voortplanting vind deur oögamie plaas (die versmelting van 'n onbeweeglike eiersel en ’n beweeglike spermsel). Die manlike geslagsel (spermsel) beweeg nie met flagella (sweephare) nie, maar dryf saam met die waterstrome.
Die selwande van koraalwiere (genus Corallina) bevat groot hoeveelhede kalk. Dooie koraalwiere pak op die seebodem saam en kan met verloop van tyd dik kalkriwwe (wat nie met koraalriwwe verwar moet word nie) vorm. Hierdie riwwe kan as 'n bron van kalk dien.
Lithothamnion calcareum kan in groot getalle in vlak kuswaters aangetref word. Hul selwande bestaan uit 40% kalk en die kalkreste van die dooie plante word deur golfbewegings verbrokkel en in die vorm van ronde, helderwit brokkies (koraalsand) op die strande uitgespoel. Die kalkbrokkies kan as kunsmis en ook by die vervaardiging van sement gebruik word. Rooiwiere is ryk aan talle spoorelemente (mikrovoedingstowwe), veral jodium, wat deur ekstraksie uit die alge verkry kan word.
Die plante is ook 'n bron van algenate, dit wil sê elektriese geleiers wat soute van algiensuur is. Algiensuur is 'n belangrike bestanddeel van rooiwierselwande omdat dit elastisiteit en stewigheid daaraan verleen. Chondrus crispus en Gigartina stellata lewer na ekstraksie ’n stroperige bindmiddel, die algiensuurryke karrageen. Agar-agar is ’n verwante produk wat in veral Japan geproduseer word. Bruinwiere word op drywende bamboestakke gekweek en die agar word na ekstraksie tot 'n poeiervorm uitgedroog.
Indien die gedroogde agar met water verbind, word gelatien gevorm en agar word om die rede veral in mirkobiologiese laboratoria gebruik om bakteriële kulture daarop te kweek. Rooiwiere is 'n bestanddeel van die bekende Japanse en Chinese voëlnessop. Die Skotse gereg "dulse" word uit Rhodymenia palmata voorberei, en Porphyra laciniata is 'n bestanddeel van Walliese "laverbread".
Rooiwiere (klas Rhodophyta) leef veraf in tropiese see en hul naam is afkomstig van hul rooi kleur, wat deur die aanwesigheid van 'n aantal uitsonderlike pigmente veroorsaak word. Hierdie pigmente kan selfs in swak lig fotosinteer. Voortplanting kan geslagtelik of ongeslagtelik wees en sommige spesies is ekonomies belangrik.
Die rooiwiere (klas Rhodophyta) het, soos die ander wiere, geen egte wortels, stingels en blare nie en die hele plantliggaam is uit homogene weefsel opgebou. So 'n struktuur word 'n tallus genoem, en hoewel blaaragtige en stingelagtige strukture by sommige vorme onderskei kan word, is dit nie gedifferensieerde organe nie. Kleurstowwe wat vir elke klas alge kenmerkend is, kom in die selle van die plante voor. In die geval van die rooiwiere word groen chlorofil d, asook rooi karoteen en fikobileen aangetref. Die twee kleurstowwe het die plante se kenmerkende rooi of violet kleur tot gevolg.
Die karoteen en fikobileen absorbeer lig vir fotosintese uit die blou deel van die spektrum, wat dit vir die rooiwiere moontlik maak om op groter dieptes (tot 75 m onder die seeoppervlak) as groenwiere te leef. Die meeste rooiwiere kom in tropiese waters voor en tydens fotosintese word die gevormde glukose omgesit in florideensetmeel, 'n stof wat naby verwant is aan dierlike glikogeen. Die selle van rooiwiere bevat ook 'n ingewikkelde koolhidraat, floridosied, wat ’n verbinding van gliserol en die monosakkaried galaktien is.
Die rooiwiere word in twee subklasse verdeel op grond van die groeiwyse van die tallus en die manier waarop die bevrugte eiersel (sigoot) ontwikkel. Die subklas Bangiophycidae bestaan uit oorwegend eensellige of draadvormige spesies wat nie duidelike groeisones het nie. Die selle het slegs een kern en het stervormige chromoplaste. Lede van die subklas Floridophycidae het gewoonlik 'n meersellige, vertakte tallus en die plante groei deurdat selfstandige selle aan die bopunt van die tallus verdeel. Die selle kan dikwels meerkernig wees en dit bevat ook 'n aantal skyfvormige chromoplaste, wat langs dieselwand gerangskik is.
Les algues coloraes o rodófites (filu Rhodophyta, del griegu ῥόδον, "rosa" y φυτόν, "planta") son un importante grupu d'algues que incluye unes 7000 especies d'una gran diversidá de formes y tamaños.[2] Formen parte del reinu Plantae al pie de Glaucophyta (glaucofites) y Viridiplantae (plantes y algues verdes).[3][4][5][6]
Caracterizar pola so inmovilidá por cuenta de la falta o perda evolutiva de flaxelos en toles etapes del so ciclu vital. Los sos cloroplastos presenten dos membranes, clorofila a y pigmentos accesorios ficobiliproteínas y carotenoides, que amazcaren el color de la clorofila y dan-y el color coloráu distintivu d'estes algues. Tán bien representaes n'agües fondes.
Rhodophyta aniciose nel Mesoproterozoicu y estrémase filoxenéticamente en dos clados: Cyanidiophytina y Rhodophytina, que diverxeron fai unos 1200 millones d'años.[7]
Porphyra haploide y diploide (Bangiophyceae)
Gracilaria (Florideophyceae, orde Gracilariales)
Les algues coloraes o rodófites (filu Rhodophyta, del griegu ῥόδον, "rosa" y φυτόν, "planta") son un importante grupu d'algues que incluye unes 7000 especies d'una gran diversidá de formes y tamaños. Formen parte del reinu Plantae al pie de Glaucophyta (glaucofites) y Viridiplantae (plantes y algues verdes).
Caracterizar pola so inmovilidá por cuenta de la falta o perda evolutiva de flaxelos en toles etapes del so ciclu vital. Los sos cloroplastos presenten dos membranes, clorofila a y pigmentos accesorios ficobiliproteínas y carotenoides, que amazcaren el color de la clorofila y dan-y el color coloráu distintivu d'estes algues. Tán bien representaes n'agües fondes.
Rhodophyta aniciose nel Mesoproterozoicu y estrémase filoxenéticamente en dos clados: Cyanidiophytina y Rhodophytina, que diverxeron fai unos 1200 millones d'años.
Qırmızı yosunlar (lat. Rhodophyta).
Qırmızı yosunlar - sarımtıl qırmızı, mavi-yaşıl-qırmızı rəngli çoxhüceyrəli (ən bəsit formaları - birhüceyrəli) yosunlar tipi. Bəzi qruplar hüceyrələrin divarlarında kalsium karbonat çökdürdüyündən çökmələrdə fossil halında qalır. Mezo-kaynozoyda qırmızı yosunlar süxurəmələgətirən orqanizmlər olub mərcan qrumlarının (biogermlər və riflər) formalaşmasında iştirak edir. Qırmızı yosunlar normal duzluluqlu isti dənizlərdə 3-160 m (optimal - 20-50 m) dərinlikdə yayılmışdır. Kembri-müasir. Hazırda 3800 növü vardır. Xəzərdə 25 növü məlumdur, onlardan laurensiya, polisifoniya, melobeziya və s. yosunlar geniş yayılmışdır.
Geologiya terminlərinin izahlı lüğəti. — Bakı: Nafta-Press, 2006. — Səhifələrin sayı: 679.
Qırmızı yosunlar (lat. Rhodophyta).
Qırmızı yosunlar - sarımtıl qırmızı, mavi-yaşıl-qırmızı rəngli çoxhüceyrəli (ən bəsit formaları - birhüceyrəli) yosunlar tipi. Bəzi qruplar hüceyrələrin divarlarında kalsium karbonat çökdürdüyündən çökmələrdə fossil halında qalır. Mezo-kaynozoyda qırmızı yosunlar süxurəmələgətirən orqanizmlər olub mərcan qrumlarının (biogermlər və riflər) formalaşmasında iştirak edir. Qırmızı yosunlar normal duzluluqlu isti dənizlərdə 3-160 m (optimal - 20-50 m) dərinlikdə yayılmışdır. Kembri-müasir. Hazırda 3800 növü vardır. Xəzərdə 25 növü məlumdur, onlardan laurensiya, polisifoniya, melobeziya və s. yosunlar geniş yayılmışdır.
Rødalger (Rhodophyta, fra oldgræsk: ῥόδον rhodon, "rose" og φυτόν phyton, "plante") er en division af alger, der hører under planteriget. De indeholder farvestoffet Klorofyl A. Rødalger er en af de ældste grupper af planter[1]. Rødalgerne er også en af de største grupper af alger, med mere end 7.000 anerkendte arter[2]. Hovedparten af rødalgerne findes i Florideophyceae-klassen, som hovedsageligt omfatter flercellede marine alger, heraf en del tang[2][3]. Kun ca. 5% af rødalgerne forekommer i ferskvand, mens resten forekommer i salt- og brakvand[4].
De ældste fossiler af eukaryote celler er af rødalger på 1,6 milliard år.[5]
Division: Rhodophyta
Rødalger (Rhodophyta, fra oldgræsk: ῥόδον rhodon, "rose" og φυτόν phyton, "plante") er en division af alger, der hører under planteriget. De indeholder farvestoffet Klorofyl A. Rødalger er en af de ældste grupper af planter. Rødalgerne er også en af de største grupper af alger, med mere end 7.000 anerkendte arter. Hovedparten af rødalgerne findes i Florideophyceae-klassen, som hovedsageligt omfatter flercellede marine alger, heraf en del tang. Kun ca. 5% af rødalgerne forekommer i ferskvand, mens resten forekommer i salt- og brakvand.
De ældste fossiler af eukaryote celler er af rødalger på 1,6 milliard år.
Die Rotalgen (Rhodophyta,[1] Synonym: Rhodophyceae) sind eine Abteilung von Algen, von denen viele durch die an der Photosynthese beteiligten Phycobiline rot gefärbt sind. Sie bilden eine der drei Gruppen der Archaeplastida. Ihre Fortpflanzung ist durch einen dreigliedrigen Generationswechsel gekennzeichnet. Alle Formen und Stadien sind unbegeißelt. Rotalgen kommen in der Mehrzahl in der Litoralzone des Meeres vor, einige Arten auch im Süßwasser und in feuchtem Erdreich. Fossil sind sie seit dem Erdzeitalter des Ectasiums (vor etwa 1400 bis 1200 Millionen Jahren) bekannt.[2]
Rotalgen sind zumeist mehrzellig und bestehen aus Zellfäden, die zu einem Plektenchym (Scheingewebe) aneinandergelagert sein und bandförmige oder lappige Formen ausbilden können.[3] Die Zellwand besteht aus Cellulose und einem Schleim aus Galactanen, zumeist Agar oder Carrageen. Einzellige Arten sind nur von einer Schleimschicht ohne Cellulose umgeben. Begeißelte Stadien fehlen im Unterschied zu anderen Algen vollständig.[4]
Bei vielzelligen Rotalgen befindet sich in der Mitte jeder Trennwand eine Struktur, die traditionell als „Tüpfel“ (engl. pit connection) bezeichnet wird, aber nicht wie die Tüpfel bei höheren Pflanzen eine Verbindung zwischen den Zellen herstellt, sondern im Gegenteil eine Pore, die bei der zentripetalen Bildung der Zellwand zunächst offen bleibt, verschließt.[5]
Die Farbe der Rotalgen variiert je nach den vorhandenen Photosynthese-Pigmenten. Arten der Litoral-Zone sind olivgrün bis rotbraun, weil sie neben Chlorophyll viel blaues Phycocyanin sowie Carotinoide enthalten. Arten tieferer Zonen enthalten viel rotes Phycoerythrin und sind rot bis violett gefärbt.[6]
Die Chloroplasten, hier auch Rhodoplasten genannt, enthalten Chlorophyll a und seine Begleitcarotinoide, aber keine anderen Chlorophylle. In den Chloroplasten liegen die Thylakoide nicht wie bei den Chloroplastida in Stapeln vor, sondern sind in gleichen Abständen nebeneinander angeordnet.
Auf den Thylakoiden sitzen 30 bis 40 nm große, mehr oder weniger kugelige Strukturen: die Phycobilisomen. Sie enthalten wasserlösliche Photosynthese-Hilfspigmente, die Phycobiliproteine. Die Hauptmasse bilden dabei mehrere Varianten von rotem Phycoerythrin; Allophycocyanine und Phycocyanine kommen ebenfalls vor. Bau und Pigmentausstattung der Rhodoplasten deuten auf ihre Verwandtschaft mit den Cyanobakterien hin (siehe auch Endosymbiontentheorie).
Das Pigment Phycoerythrin ist photodestruktiv. Das bedeutet, dass dieses Pigment mit zunehmender Lichtintensität zunehmend zerstört wird. Dies ist der Grund dafür, dass viele oberflächennah lebende Rotalgen nicht rot aussehen. Mit Hilfe der Phycobiliproteine minimieren die Rotalgen die Grünlücke der grünen Pflanzen und können auch „mittelwellige VIS“ (Bereich der visuellen und gleichzeitig größtenteils auch photosynthetisch aktiven Strahlung) absorbieren und auf die photosynthetisch aktiven Chlorophyll a-Moleküle übertragen. Da Licht mit einer Wellenlänge von ca. 380 nm bis 550 nm (blau bis grün) schlecht von Wassermolekülen und ihren Soluten absorbiert wird, gelangt es in große Tiefen (über 250 m). Phycoerythrin verstärkt die Absorption vor allem von 490 nm bis 580 nm (vom hellblauen bis zum Anfang des gelben Bereiches).
Das primäre Produkt der Photosynthese ist hauptsächlich der Zuckeralkohol Floridosid. Der Gehalt an gelöstem Floridosid korreliert mit dem Salzgehalt des Wassers und kann bis zu 10 % der Trockenmasse der Alge ausmachen. Dies wirkt einem Wasserverlust durch Osmose entgegen.[7] Das Reservepolysaccharid ist die Florideenstärke, ein Polymer des Zuckeranteils des Floridosids. Sie wird nicht wie die Stärke der höheren Pflanzen in den Plastiden, sondern an der Oberfläche der Chloroplasten oder im Cytosol gespeichert. Chemisch ist sie näher mit dem tierischen Glykogen als mit pflanzlicher Stärke verwandt.
Neben der ungeschlechtlichen Vermehrung durch Sporen weisen Rotalgen typischerweise einen dreigliedrigen Generationswechsel auf, bei dem nacheinander Gametophyten, Karposporophyten (oder Gonimoblasten) und Tetrasporophyten auftreten. Die Bildung eines Karposporophyten wird als Apomorphie der Rotalgen betrachtet.
Das weibliche Geschlechtsorgan, das einzellige Karpogon, bildet einen dünnen fädigen Fortsatz aus, die Trichogyne, die dem Kontakt mit den männlichen Gameten (Spermatien) dient. Das männliche Geschlechtsorgan, das Spermatangium, bringt nur ein kugelförmiges oder längliches Spermatium hervor, das von einer klebrigen Masse umgeben ist. Bei manchen Rotalgen hat diese Umhüllung faserige Fortsätze, welche die Chance erhöhen, Kontakt zu einer Trichogyne zu finden. Die Trichogyne wächst gewöhnlich so lange weiter, bis sich ein Spermatium an sie bindet. Die Spermatien sind – wie alle Zellen der Rotalgen – stets unbegeißelt und werden passiv durch die Wasserströmung bewegt.
Das Karpogon enthält zwei Zellkerne, von denen einer in der Trichogyne sitzt und später vergeht. Der andere, basale Zellkern verschmilzt bei der Befruchtung mit dem Kern eines Spermatiums, der in die Trichogyne entlassen wurde und durch diese hinabwandert. Aus der nun diploiden Zygote wächst der wenigzellige fädige Gonimoblast (auch Karposporophyt genannt) heraus. Dieser stellt wegen des erfolgten Wechsels der Kernphase eine neue Generation dar, bleibt aber mit der Mutterpflanze verbunden. Er setzt schließlich diploide Karposporen frei, aus denen die diploiden Tetrasporophyten als dritte Generation hervorgehen, welche äußerlich meist den Gametophyten der ersten Generation gleichen. In ihren Tetrasporangien erfolgt die Meiose, und es entstehen zwei weibliche und zwei männliche Tetrasporen. Mit den aus diesen hervorgehenden Gametophyten schließt sich der Lebenszyklus.[8][9]
Von diesem typischen Lebenszyklus gibt es viele Ausnahmen. So sind bei einzelligen Rotalgen wie den Cyanidiales und den Porphyridiales keine sexuellen Vorgänge bekannt, und die Bangiales zeigen einen zweigliedrigen Generationswechsel, bei dem die Zygote unmittelbar die Karposporen bildet, also kein Gonimoblast ausgebildet wird.
Die Rotalgen leben ganz überwiegend im Meer, und sie sind unter den Meeresalgen mit mehr Arten repräsentiert als alle sonstigen Algengruppen zusammen. Allerdings sind sie weitgehend auf gemäßigte bis tropische Breiten beschränkt, während in kalten Meeresbereichen Braunalgen und Grünalgen überwiegen. Dank der Phycobiline können sie in größeren Wassertiefen Photosynthese betreiben als andere Algen.[10]
Im Süßwasser leben etwa 178 Arten von Rotalgen, das sind etwas mehr als drei Prozent der gesamten Artenzahl. Soweit bekannt, bevorzugen alle Arten Fließgewässer (oder zumindest bewegtes Wasser) und kommen am Boden (benthisch), meist aufwachsend auf Hartsubstraten, vor. Wie typisch für Rotalgen, werden relativ gering belichtete Habitate, zum Beispiel durch Huminstoffe getrübtes Wasser, bevorzugt. Einige Arten bevorzugen Sonderhabitate, so zum Beispiel Cyanidium und Galdieria in Thermalquellen in Italien.[11] Die meisten süßwasserlebenden Arten sind nicht rot gefärbt, stattdessen kommen gelbgrüne, blaugrüne, olivgrüne, violette, bräunliche bis zu grauschwarze Arten vor; meist ist das Phycoerythrin bei ihnen durch Phycocyanin überdeckt. Viele Süßwasser-Rotalgen sind empfindlich gegenüber Gewässerverschmutzung, einige gelten als bedroht. In Europa sind Süßwasser-Rotalgen in den nördlichen Breiten artenreicher als im Süden; die meisten Arten (28) sind in Finnland nachgewiesen, in Deutschland sind es 21, in Österreich 22 und in der Schweiz 12 Arten. Die häufigsten und am weitesten verbreiteten Gattungen sind Audouinella, Bangia, Batrachospermum, Hildenbrandia und Lemanea.[12]
Manche Rotalgen wachsen als Epiphyten oder als Parasiten auf anderen Algen. Bei ersteren unterscheidet man fakultative, die auch auf toten Substraten wachsen können, und obligate Epiphyten, die nur auf einem passenden Wirt gedeihen können. Ein obligater Epiphyt ist die Pinselbüschelalge. Deren Sporen keimen auf der Oberfläche des Knotentangs (einer Braunalge). Sie bilden ein Rhizoid aus, das mittels Verdauungsenzymen in das Wirtsgewebe eindringt und den Epiphyten verankert. Parasitische Rotalgen nehmen im Unterschied zu bloßen Epiphyten Nährstoffe aus dem Wirt auf. Dabei können sie auch selbst noch Photosynthese betreiben oder als Vollparasiten keine eigenen Photosynthese-Pigmente mehr ausbilden und ganz auf die Ernährung durch den Wirt angewiesen sein. Ein interessantes Beispiel für einen Vollparasiten ist Choreocolax polysiphoniae, der die Pinselbüschelalge parasitiert, die wiederum als Epiphyt auf dem Knotentang wächst. Auch Choreocolax dringt in den Wirt ein, bildet aber Tüpfelverbindungen zu dessen Zellen aus, um Nährstoffe aufzunehmen. Deshalb spricht man hier nicht von einem Rhizoid, sondern von einem Haustorium.[13]
Die Rotalgen werden seit der Mitte des 19. Jahrhunderts als eigenes Taxon anerkannt. 1851 stellte Franz Joseph Ruprecht die Klasse Rhodophyceae auf. 1901 stufte Richard Wettstein die Rotalgen als Rhodophyta im höheren Rang einer Abteilung ein. Im 20. Jahrhundert wurden die Rhodophyta bzw. Rhodophyceae meist in zwei Hauptgruppen eingeteilt, die Florideophyceae (Florideen) und die Bangiophyceae. Neuere molekulargenetische Untersuchungen führten jedoch zu einem stärker unterteilten System.[26][27]
Adl et al. schlugen 2012 eine strikt phylogenetische Einteilung vor, die auf jegliche Rangstufen verzichtet:[28]
Diese ranglose Klassifikation wurde jedoch von den meisten Phykologen nicht aufgegriffen. Im Syllabus of Plant Families von 2017 gliedern Kamiya et al. die Rotalgen wie folgt (Artenzahlen ergänzt nach AlgaeBase, Stand Juli 2018):[1][29]
Abteilung Rhodophyta, mit etwa 7130 Arten
Die Erforschung der Systematik der Rotalgen ist noch nicht abgeschlossen. Durch molekulargenetische Untersuchungen werden jährlich neue Arten entdeckt und weitere Änderungen der Einteilung notwendig werden.
Für die in der Deutschen Bucht vorkommenden Rotalgen siehe auch die Liste der Meeresalgen von Helgoland.
Die Rotalgen (Rhodophyta, Synonym: Rhodophyceae) sind eine Abteilung von Algen, von denen viele durch die an der Photosynthese beteiligten Phycobiline rot gefärbt sind. Sie bilden eine der drei Gruppen der Archaeplastida. Ihre Fortpflanzung ist durch einen dreigliedrigen Generationswechsel gekennzeichnet. Alle Formen und Stadien sind unbegeißelt. Rotalgen kommen in der Mehrzahl in der Litoralzone des Meeres vor, einige Arten auch im Süßwasser und in feuchtem Erdreich. Fossil sind sie seit dem Erdzeitalter des Ectasiums (vor etwa 1400 bis 1200 Millionen Jahren) bekannt.
Crvene alge (lat. Rhodophyta, od grč. ῥόδον – rodon = crven + φυτόν – fiton = biljka) su velika skupina najčešće višećelijskih, morskih algi, uključujući i mnoge morske trave. Njihovo taksonomsko rangiranje kreće se od razreda do koljena. Donedavno su preovladavali i enciklopedijski podaci o broju vrsta između 2.000 i 7.000 recentnih vrsta. Međutim, prema novijim izvorima, dok neki drugi izvori govore da postoji više od 10.000 vrsta u ovom koljenu algi Rhodophyta obuhvata jedno od najvećih koljena alga, koje sadrži preko 7000 trenutno prepoznatih vrsta s tekućim taksonomskim revizijama.[1] Glavnina vrsta (6,793) pripada razredu Florideophyceae, a većinom se sastoje od višećelijskih, morskih algi, uključujući mnoge značajne morske trave.[1][2]
Slatkovodni ih je približno 5%, koje su najučestalije su u toplijim područjima .[3] Izuzev dvije vrste obalnih špiljskih staništa u razredu Cyanidiophyceae, sa bespolnim razmnožavanjem, koje su se divergirale od drugih crvenih algi prije oko 1,3 milijarde godina,[4] nema kopnenih vrsta, što bi moglo biti posljedica evolucijskog uskog grla, kada je posljednji zajednički predak izgubio oko 25% svojih osnovnih gena i velik dio evolucijske plastičnosti.[5]
Većina najuglednijih autora, slaže se sa procjenom da su jedna od najstarijih skupina eukariotskih algi.[6] Samo dvadesetak rodova nastanjuje i vode na kopnu. Većinom žive pričvršćene za kamenitu podlogu, ali i za školjke, te vrlo rijetko kao epifiti pričvršćene za druge alge. Vrlo su ostjetljive na promjene temperature. Dosta su sitnije od smeđih algi, pa se na područjima jakih talasa ne mogu održati na podlozi; zato pretežno žive u mirnim dubokim vodama.
Najstariji fosilni nalazi ovih algi datiraju iz mezoproterozoika, prije otprilike jedne do 1,3 milijarde godina.
Crvene alge su višećelijski organizmi, rijetko jednoćeliski, raznolikog oblika talusa: končastog, perastog ili razgranarog, krpastog ili člankovitog, Neke od njih podijeljen u rizoide, kauloide i filoide. Boja talusa je obično svjetlocrvena do ljubičasta, a neke su crvenoljubičaste ili smeđecrvene. Pigmenti su vidu varijante hlorofila a, kod nekih sa primjesama malih količina hlorofila d i karotinoid koji su prekriveni veoma fluorescentnim crvenim, u vodi topivim fikoeritrinom. Kod nekih vrsta dolazi i modri fikocijan. U proceu fotosinteze stvata se tzv. floridejski škrob, ugljikohidrat između škroba i glikogena.
Crvene alge razmnožavaju se, spolno i nespolno]] (uključujući i vegetativno). I spolne i nespolne reprodukcijske ćelije su nepokretne.
Žive u moru, rijetko u slatkoj vodi ili na kopnu. Najćešće su u litoralnoj zoni, posebno toplijih mora, a naseljavaju i veće dubine, do oko 200 m.
Mnoge crvene alge upotrebljavaju se kao hrana za ljude i domaće životinje, što je posebno rašireno u Japanu. Također, i neke su važne i industrijskoj proizvodnji agara, kao one iz rodova Gelidium i Gracillaria). A sgar je vema značajan u farmaciji, prehrambenoj industriji, medicinskoj mikrobiologiji i raznim laboratorijama, kao osnova hranjivih podloga za uzgoj mikroorganizama i mnogih dugih analiza. .
Crvene alge su jako posebna skupina algi. Imaju eukariotsku stanicu bez biča i centriola. Njihova stanična stijenka ima unutarnji sloj sastavljen od celuloze, dok je vanjski sloj građen od pektina, te je sluzast. Imaju jednu jezgru, rijetke su one koje u starosti imaju više jezgara. Kao rezerva za hranu služi im ugljikohidrat floridejski škrob (između glikogena i škroba), koji se u obliku zrnaca nalazi na površini rodoplasta. Crvenu boju daju im fikobilini. Kloroplasti nemaju endoplazmatski retikulum, a građeni su od paralelno naslaganih vrećica, tilakoida.
Crvene alge se razmnožavaju nespolno, spolno ili izmjenom spolnog i nespolnog razmnožavanja (smjena generacija). Ciklus razmnožavanja može biti pokrenuto različitim faktorimaa, kao što je dužina obdanice[6]. Nemaju pokretnih ćelija, a spore i gameti su nepokretni, pa su morska stujanja glavni put za oplodnju. Vegetativno razmnožavanje se odvija kod jednoćelijskih oblika diobom, raskidanjem kolonije ili parčanjem talusa. Također postoji razmnožavanje sporama, uz tvorbu posebnih tipova spora, kao što su monospore, tetraspore i polispore. Spolno razmnožavanje odvija se atipskom oogamijom.
Iz mnogih koralnih algi, koje izlučuju kalcij-karbonat, stvarajući koralne grebenove, dobija se materijal višestruke namjene. Ćelijskog zida nekih vrsta dobijaju se različiti polisaharidi za pripremu lijekova Također, Neke, iz ove grupe crvenih algi, dio su drevne evropske i azijske kuhinje, za pripravljenje raznih dodataka kao prehrambenih aditiva. Sadrže veliku količinu vitamina i bjelančevina, a jednostavno se uzgajaju, pa su zbog toga pogodnene za kultivaciju, koja je u Japanu počela prije više od 300 godina.
Evolucija hloroplast počela je nakon endosimbiotskog spajanja predaka, fotosintetskih cijanobakterija i ranih eukariotskih fagotrofa.[7] Ovaj događaj (nazvan primarna endosimbioza) rezultirao je nastankom crvenih i zelenih algi, i the Glaucophyta, koji čine najstarije evolucijske linije fotosintetskih eukariota.[8] Sekundarna endosimbioza koja uključuje predačku crvenu algu i heterotrofni eukariot, što je rezultirao evolucijom i diversifikacijom nekoliko drugih fotosintetskih rodova poput Cryptophyta, Haptophyta, Stramenopiles (ili Heterokontophyta) i Alveolata.[8] Pored višećelijskih smeđih algi, procjenjuje se da više od polovine svih poznatih vrsta mikrobnih eukariota ima pladide koji potiču iz crvenih algi.[9]
Crvene alge dijele se na Cyanidiophyceae, klasu jednoćelijskih i termoacidofilnih ekstremofila, koje se nalaze u sumpornim vrelima i drugim kiselim okruženjima, a prilagođavanje je djelomično omogućeno horizontalnim prijenosom prokariotskih gena, s tim da oko 1% njihovog genoma ima ovo porijeklo i dva sestrinska taksona koja se nazivaju okružrnje SCRP (prema porodicama Stylonematophyceae, Compsopogonophyceae, Rhodellophyceae i Porphyridiophyceae) i BF (Bangiophyceae i Floridephyceae (oba u moru), na kojima se nalaze i Floridephyceae) . Kladus SCRP obuhvata mikroalge, jednoćelijske oblike i višećelijske mikroskopske niti i lopatice. BF su makroalge, morske alge koje obično ne narastu na duže od otprilike 50 cm, ali nekoliko vrsra može dostići i do oko 2 m.[10][11][12][12]
Većina rodofita su morske alge, s globalnom rasprostranjenošću, a često se nalaze na većim dubinama u usporedbi s drugim morskim algama. Iako je ovo ranije pripisivano prisustvu pigmenata (poput fikoeritrina) koji su omogućili crvenim algama da nastanjuju veće dubine od ostalih makroalgi prilagodbom boje, noviji dokazi to dovode u pitanje (npr. otkriće zelenih algi u velikim dubinama na Bahamima).[13] Neke morske vrste nalaze se na pješčanim obalama, dok se većina drugih može naći na stjenovitim supstratima.[14] Slatkovodne vrste čine 5% raznolikosti crvenih algi, ali one imaju i rasprostranjenje u različitim staništima širom svijeta; uglavnom vole čiste vodotokove, s bistrim vodama i kamenitim dnom, ali s nekim iznimkama.[15] Nekoliko slatkovodnih vrsta živi u crnim vodama sa pješčanim dnima [16] a još manje ih se nalazi u više skoro slatkim vodama.[17] I morske i slatkovodne predstavljene su slobodnim makroalgama i manjim endo / epifitskim / oblicima, što znači da žive u ili na drugim algama, biljkama i životinjama. Pored toga, neke morske vrste usvojile su parazitski način života i mogu se naći na bliskim ili udaljenijim domaćinima crvenih algi.
Cyanidium sp. (Cyanidiophyceae)
Porphyra sp., haploidne i diploidne (Bangiophyceae)
Gracilaria sp. (Florideophyceae: Gracilariales)
Corallina officinalis sp. (Florideophyceae: Corallinales)
Laurencia sp. (Florideophyceae: Ceramiales)
Neke crvene alge se pod vodom prelijevaju u duginim bojama
Crvene alge razvrstane su u dva potkoljena šest redova:
Potkoljeno Cyanidiophytina H.S.Yoon, K.M.Müller, R.G.Sheath, F.D.Ott & D.Bhattacharya
Potkoljeno Eurhodophytina G.W.Saunders & Hommersand
C) Potkoljeno Proteorhodophytina Muñoz-Gómez, Mejía-Franco, Durnin, Colp, Grisdale, J.M.Archibald & Slamovits
Razred Rhodophyta classis incertae sedis
Crvene alge (lat. Rhodophyta, od grč. ῥόδον – rodon = crven + φυτόν – fiton = biljka) su velika skupina najčešće višećelijskih, morskih algi, uključujući i mnoge morske trave. Njihovo taksonomsko rangiranje kreće se od razreda do koljena. Donedavno su preovladavali i enciklopedijski podaci o broju vrsta između 2.000 i 7.000 recentnih vrsta. Međutim, prema novijim izvorima, dok neki drugi izvori govore da postoji više od 10.000 vrsta u ovom koljenu algi Rhodophyta obuhvata jedno od najvećih koljena alga, koje sadrži preko 7000 trenutno prepoznatih vrsta s tekućim taksonomskim revizijama. Glavnina vrsta (6,793) pripada razredu Florideophyceae, a većinom se sastoje od višećelijskih, morskih algi, uključujući mnoge značajne morske trave.
Slatkovodni ih je približno 5%, koje su najučestalije su u toplijim područjima . Izuzev dvije vrste obalnih špiljskih staništa u razredu Cyanidiophyceae, sa bespolnim razmnožavanjem, koje su se divergirale od drugih crvenih algi prije oko 1,3 milijarde godina, nema kopnenih vrsta, što bi moglo biti posljedica evolucijskog uskog grla, kada je posljednji zajednički predak izgubio oko 25% svojih osnovnih gena i velik dio evolucijske plastičnosti.
Većina najuglednijih autora, slaže se sa procjenom da su jedna od najstarijih skupina eukariotskih algi. Samo dvadesetak rodova nastanjuje i vode na kopnu. Većinom žive pričvršćene za kamenitu podlogu, ali i za školjke, te vrlo rijetko kao epifiti pričvršćene za druge alge. Vrlo su ostjetljive na promjene temperature. Dosta su sitnije od smeđih algi, pa se na područjima jakih talasa ne mogu održati na podlozi; zato pretežno žive u mirnim dubokim vodama.
Najstariji fosilni nalazi ovih algi datiraju iz mezoproterozoika, prije otprilike jedne do 1,3 milijarde godina.
'S i an fheamainn dearg seòrsachan a dh' algae dearga a tha a' fàs shìos seachad air an fheamainn chìrean, an fheamainn ghropach agus an fheamainn dubh air a' chladach.
Air Gàidhealtachd na h-Alba, bhite ga buain airson na talmhainn. 'S ann an còmhnaidh air an fheannag eòrna a bha an fheamainn dearg ri dol.[1]
Faic feamainn cuideachd.
'S i an fheamainn dearg seòrsachan a dh' algae dearga a tha a' fàs shìos seachad air an fheamainn chìrean, an fheamainn ghropach agus an fheamainn dubh air a' chladach.
Air Gàidhealtachd na h-Alba, bhite ga buain airson na talmhainn. 'S ann an còmhnaidh air an fheannag eòrna a bha an fheamainn dearg ri dol.
Qizil suvoʻtlar — suvoʻtlar tipi. Bir hujayrali va koʻp hujayrali shakllari bor. Xromatoforida xlorofilldan tashqari, koʻkyashil pigment — fikotsian va qizil pigment — fikoeritrin mavjud. Qizil suvoʻtlarning harakatchan xivchinli davri boʻlmaydi. Tanasi ipsimon, shoxlangan (qalin), plastinkasimon, baʼzilari poya va barglarga boʻlingan. Rangi toʻq qizildan, qora, pushti, sargʻish va baʼzan koʻkimtir kulranggacha. Qizil suvoʻtlarning 600 turkumga mansub 3750 turi bor, ular bangiyalar va floridlar sinfiga boʻlinadi. Bu sinflar bir-biridan koʻpayish organlarining tuzilishi bilan farq qiladi.
Qizil suvoʻtlar koʻpincha dengizlarda (chuqur joylarida), kamdankam chuchuk suvlarda va tuproqda yashaydi. Qazilma formalari boʻr davri qatlamlaridan topilgan.
Anfeltsiya, gelidium, fillofora, furselyariya turlaridan agaragar, karragen, agaroid va boshqalar olinadi. Baʼzi turlari, mas., porfira isteʼmol qilinadi.
The pulang algae, o Rhodophyta ( /roʊˈdɒfᵻtə/ roh-DOF-it-ə, /ˌroʊdəˈfaɪtə/ ROH-də-FY-tə; mula sa Lumang Griyego ῥόδον (rhodon), nangangahulugang 'rosas', at φυτόν (phyton), nangangahulugang 'halaman'), ay isa sa mga pangkat ng eukaryotikong algae.[1] Binubuo din ang Rhodophyta ng isa sa pinakamalaking lapi ng algae na mayroong higit sa 7,000 kinikilalang espesye na may patuloy na pagbabago sa taksonomiya.[2] Matatagpuan ang karamihan ng espesye (6,793) sa klaseng Florideophyceae, at karamihang binubuo ng multiselular, pang-karagatan na algae, kabilang ang maraming kilalang damong-dagat.[2][3] Tinatayang 5% ng mga pulang algae ay matatagpuan sa tubig-tabang na may mas malawak na konsentrasyon sa mga lugar na mas mainit.[4]
Die Rhodophyta sünd en Ünnerriek vun't Riek vun de Protisten. Se sünd Eukaryoten un sünd vör allen in de Litoralzoon vun't Meer. Ok in anner Water gifft dat Rhodophyta. De Kanten vun Australien hebbt ene relativ grote Tahl vun Oorden. Dat gifft circa 4000 Oorden. De Ünnerklassen sünd Florideophycidae un Bangiophycidae.
Rhodophyta es un division de Biliphyta, Archaeplastida.
The pulang algae, o Rhodophyta ( /roʊˈdɒfᵻtə/ roh-DOF-it-ə, /ˌroʊdəˈfaɪtə/ ROH-də-FY-tə; mula sa Lumang Griyego ῥόδον (rhodon), nangangahulugang 'rosas', at φυτόν (phyton), nangangahulugang 'halaman'), ay isa sa mga pangkat ng eukaryotikong algae. Binubuo din ang Rhodophyta ng isa sa pinakamalaking lapi ng algae na mayroong higit sa 7,000 kinikilalang espesye na may patuloy na pagbabago sa taksonomiya. Matatagpuan ang karamihan ng espesye (6,793) sa klaseng Florideophyceae, at karamihang binubuo ng multiselular, pang-karagatan na algae, kabilang ang maraming kilalang damong-dagat. Tinatayang 5% ng mga pulang algae ay matatagpuan sa tubig-tabang na may mas malawak na konsentrasyon sa mga lugar na mas mainit.
Les rodjès ales, c' est des ales k' ont ene rodje coleur.
Famile : Rhodophyta.
On s' endè sieve po fé del djelêye d' agår po les labos.
Τα Ροδόφυτα (Rhodophytes), γνωστά επίσης ως Ροδοφύκη (Red algae), αποτελούν μια ταξινομική υποομάδα των φυκών (μίας ευρύτερης ομάδας θαλάσσιων οργανισμών χαρακτηριστικά πολυμορφικής), ένα Άθροισμα (ή Διαίρεση ή Συνομοταξία). Είναι ευκαρυωτικοί οργανισμοί, ανήκουν στα θαλάσσια μακρόφυτα (δεν είναι δηλαδή μικροφύκη) λόγω του ευμεγέθους θαλλού τους και μάλιστα στα μακροφύκη (διαφέροντας από τα θαλάσσια φανερόγαμα), παρουσιάζουν δηλαδή απλότητα στη δομή τους (απουσία ρίζας, φύλλων, αγωγών στοιχείων). Εμφανίζονται ως μονοκύτταρα, νηματοειδή, συναθροίσεις νημάτων, καθώς και ως στρώσεις κυττάρων και ζουν σε νερά βάθους περίπου 125 μέτρων. Τα περισσότερα είναι θαλάσσια και λίγα ευημερούν στα γλυκά νερά. 20 είδη του Αθροίσματος ζουν ως επιβιωτικά σε ζωικούς οργανισμούς παρέχοντας π.χ. παραλλαγή σε θηρευτές ώστε να μη γίνονται αντιληπτοί από τη λεία. Άλλα είδη παρασιτούν εις βάρος επίσης Ροδοφυκών προκειμένου να επιβιώσουν και άλλα φιλοξενούν επιβακτήρια τα οποία τους παρέχουν διάφορα θρεπτικά συστατικά (π.χ. βιταμίνες, ανόργανα θρεπτικά, κ.ά.)
Οι χαρακτήρες που αποτελούν κριτήρια για τη συστηματική κατάταξη (ταξινόμηση) των φυκών είναι:
Έτσι τα χαρακτηριστικά που διακρίνουν τα Ροδόφυτα είναι:
Με τη σειρά τους, τα Ροδόφυτα περιλαμβάνουν τις εξής κλάσεις (και τις κυριότερες υποομάδες) (κατά Yoon et al. 2006, αδεία Journal of Phycology):
Συνολικά υπολογίζονται περίπου 500-600 γένη και 5000-20000 είδη (κατώτερες ταξινομικές ομάδες) Ροδοφύτων.
Οι χρήσεις των Ροδοφυκών χρονολογούνται από πολύ παλιά. Γραπτές καταγραφές επιβεβαιώνουν ότι οι άνθρωποι στην Κίνα συγκόμιζαν μακροφύκη για περισσότερο από 2000 χρόνια. Σήμερα δε οι Κινέζοι συλλέγουν 74 είδη Ροδοφυκών, Χλωροφυκών, Φαιοφυκών και Κυανοβακτηρίων από 36 γένη. Επιπλέον πάνω από 300 χρόνια πριν στην Ιαπωνία ξεκίνησε η συστηματική καλλιέργεια των ειδών Porphyra yezoensis και Porphyra tenera (γνωστά ως nori στην παρασκευή σούσι) εξαιτίας των μειωμένων απαιτήσεών τους αλλά και της υψηλής θρεπτικής τους αξίας. Παρέχουν κυρίως βιταμίνες (B, C και A), ιχνοστοιχεία (συμπεριλαμβανομένου του ιωδίου), αφομοιώσιμες πρωτεΐνες και ελεύθερα αμινοξέα. Μερικά είδη καλλιεργούνται επίσης για την παρασκευή βιομηχανικών προϊόντων. Τέλος, τα κοραλλινοειδή Ροδοφύκη είναι πολύ σημαντικά ως παραγωγοί ανθρακικών αλάτων, αποτελώντας έτσι βασικό κρίκο σε οικολογικό επίπεδο στα υδάτινα οικοσυστήματα, συμβάλλοντας στην ανακύκλωση των στοιχείων.
Τα Ροδόφυτα (Rhodophytes), γνωστά επίσης ως Ροδοφύκη (Red algae), αποτελούν μια ταξινομική υποομάδα των φυκών (μίας ευρύτερης ομάδας θαλάσσιων οργανισμών χαρακτηριστικά πολυμορφικής), ένα Άθροισμα (ή Διαίρεση ή Συνομοταξία). Είναι ευκαρυωτικοί οργανισμοί, ανήκουν στα θαλάσσια μακρόφυτα (δεν είναι δηλαδή μικροφύκη) λόγω του ευμεγέθους θαλλού τους και μάλιστα στα μακροφύκη (διαφέροντας από τα θαλάσσια φανερόγαμα), παρουσιάζουν δηλαδή απλότητα στη δομή τους (απουσία ρίζας, φύλλων, αγωγών στοιχείων). Εμφανίζονται ως μονοκύτταρα, νηματοειδή, συναθροίσεις νημάτων, καθώς και ως στρώσεις κυττάρων και ζουν σε νερά βάθους περίπου 125 μέτρων. Τα περισσότερα είναι θαλάσσια και λίγα ευημερούν στα γλυκά νερά. 20 είδη του Αθροίσματος ζουν ως επιβιωτικά σε ζωικούς οργανισμούς παρέχοντας π.χ. παραλλαγή σε θηρευτές ώστε να μη γίνονται αντιληπτοί από τη λεία. Άλλα είδη παρασιτούν εις βάρος επίσης Ροδοφυκών προκειμένου να επιβιώσουν και άλλα φιλοξενούν επιβακτήρια τα οποία τους παρέχουν διάφορα θρεπτικά συστατικά (π.χ. βιταμίνες, ανόργανα θρεπτικά, κ.ά.)
Кызыл балырлар (лат. Rhodophyta) - ар түрдүү түстөрдө болушат: ачык кызыл, нукуракызыл, көккызыл, саргычкызыл, көкжашыл. Хроматофордо хлорофилл «а», «d» (хлорофилл «а» бир гана К. б-да кездешет), каротин, ксантофилл, фикоэритрин, фикоциан сыяктуу пигменттер болушат. Бул пигменттердин түрдүү абалына байланыштуу К. б. да түрдүүчү түстөрдө кездешишет. К. б. жеке жөнөкөй жипчелер, же топтошкон өсүндүлөр, бутакчалар, пластинкалар, кээде «сабакча», «жалбыракчалар» түрүндөгү талломдуу өсүмдүктөр.
Булардын чоңдугу 2-3 м, айрымдары микроскоптук кичинекей. Негизинен деңиздерде, аз санда тузсуз сууларда жашашат.
Көпчүлүк жөнөкөй түзүлүштөрүнүн клеткалары бир ядролуу, жогорку түзүлүштөрүндө көп ядролуу.
Бир клеткалуу жана колониялуу формаларында вегетативдик көбөйүү клеткалардын бөлүнүүсү менен жүрөт. Көптөгөн флоридиялар классынын өкүлдөрүндө кошумча бутактардын болушу менен ишке ашат.
Жыныссыз көбөйүү моноспора, тетраспоралар аркылуу. Жыныстык көбөйүү оогамдык.
К. б. 600дөн ашык тукумдуу, 4 миңге жакын түрлөрү белгилүү. Кыргызстандын дарыя сууларында (Шахимардан, Исфайрам, Куршаб жана башка) К. б-дын 5 түрү кездешет. Алар бангия жана шантранзия тукумдарынын түрлөрү (Bangia atropurpurea, Chantransia chalubea, Ch. Chalubea var. leiblenii, Batrachospermum moniliforme, Lemanea fluviatulus).
Кызыл балырлар (лат. Rhodophyta) - ар түрдүү түстөрдө болушат: ачык кызыл, нукуракызыл, көккызыл, саргычкызыл, көкжашыл. Хроматофордо хлорофилл «а», «d» (хлорофилл «а» бир гана К. б-да кездешет), каротин, ксантофилл, фикоэритрин, фикоциан сыяктуу пигменттер болушат. Бул пигменттердин түрдүү абалына байланыштуу К. б. да түрдүүчү түстөрдө кездешишет. К. б. жеке жөнөкөй жипчелер, же топтошкон өсүндүлөр, бутакчалар, пластинкалар, кээде «сабакча», «жалбыракчалар» түрүндөгү талломдуу өсүмдүктөр.
Булардын чоңдугу 2-3 м, айрымдары микроскоптук кичинекей. Негизинен деңиздерде, аз санда тузсуз сууларда жашашат.
Көпчүлүк жөнөкөй түзүлүштөрүнүн клеткалары бир ядролуу, жогорку түзүлүштөрүндө көп ядролуу.
Бир клеткалуу жана колониялуу формаларында вегетативдик көбөйүү клеткалардын бөлүнүүсү менен жүрөт. Көптөгөн флоридиялар классынын өкүлдөрүндө кошумча бутактардын болушу менен ишке ашат.
Жыныссыз көбөйүү моноспора, тетраспоралар аркылуу. Жыныстык көбөйүү оогамдык.
К. б. 600дөн ашык тукумдуу, 4 миңге жакын түрлөрү белгилүү. Кыргызстандын дарыя сууларында (Шахимардан, Исфайрам, Куршаб жана башка) К. б-дын 5 түрү кездешет. Алар бангия жана шантранзия тукумдарынын түрлөрү (Bangia atropurpurea, Chantransia chalubea, Ch. Chalubea var. leiblenii, Batrachospermum moniliforme, Lemanea fluviatulus).
Кызыл суүсемнәрнең үзенчәлеге иң элек пигментлар җыелмасында чагыла. Аларның хлоропластларында, хлорофилл һәм каротиноидлардан тыш, суда эрүчән пигментлар бар: кызыл — фикоэритриннар һәм зәңгәр — фикоцианиннар. Бу пигментларның микъдарына карап, таллом төрле төстә була: куе алсу кызыл төстән алып (фикоэритрин күбрәк булганда) күксел ачык соры төскә кадәр (фикоцианин күбрәк булганда). Кызыл суүсемнәр өчен үзенчәлекле багрянка крахмалы дип йөртелә торган запас матдә аларның цитоплазмасында хлоропластлардан читтә туплана. Бу полисахарид крахмалга караганда гликогенга якынрак.
Ихтимал, багрянкаларның хлоропластлары симбиотик цианобактерияләрдән килеп чыккандыр, алар биохимик яктан да, структуралары белән дә охшашлар.
Кызыл суүсемнәр тропик һәм субтропик пояслар диңгезләрендә, шулай ук өлешчә уртача климат өлкәләрендә (Кара диңгез яр буенда) таралган. Төрләрнең гомуми саны 4 меңгә җитә, туларның 200е генә төче сулыкларда һәм туфракта яши.
Кайбер төр багрянкаларның таллом төзелеше югарырак төзелешле көрән суүсемнәр тәне белән охшаш. Аларның катламасы күп күзәнәкле тармакланган җепләрдән торган куак рәвешендә, сирәк кенә 2 м га кадәр җиткән пластинкасыман яки яфраксыман.
Кызыл суүсемнәр диңгез төбенә якын катламнарда таралган үсемлекләр дөньясының (бентосның) иң зур төркемен тәшкил итәләр. Көнчыгыш Азия илләрендә багрянкаларның кайбер төрләрен үрчетеп, азык итеп файдаланалар. Күбесе микробиологиядә микроблар үрчетү өчен туклыклы тирәлек сыйфатында кулланыла торган агар-агар һәм йод табу өчен чимал булып хезмәт итә. Норвегиядә су күтәрелә һәм чигенә торган зонада (литораль) кызыл суүсемнәр утырып калган җирләрне, су чигенгән вакытта, көтүлек итеп файдаланалар.
Кызыл суүсемнәрнең үзенчәлеге иң элек пигментлар җыелмасында чагыла. Аларның хлоропластларында, хлорофилл һәм каротиноидлардан тыш, суда эрүчән пигментлар бар: кызыл — фикоэритриннар һәм зәңгәр — фикоцианиннар. Бу пигментларның микъдарына карап, таллом төрле төстә була: куе алсу кызыл төстән алып (фикоэритрин күбрәк булганда) күксел ачык соры төскә кадәр (фикоцианин күбрәк булганда). Кызыл суүсемнәр өчен үзенчәлекле багрянка крахмалы дип йөртелә торган запас матдә аларның цитоплазмасында хлоропластлардан читтә туплана. Бу полисахарид крахмалга караганда гликогенга якынрак.
Ихтимал, багрянкаларның хлоропластлары симбиотик цианобактерияләрдән килеп чыккандыр, алар биохимик яктан да, структуралары белән дә охшашлар.
Кызыл суүсемнәр тропик һәм субтропик пояслар диңгезләрендә, шулай ук өлешчә уртача климат өлкәләрендә (Кара диңгез яр буенда) таралган. Төрләрнең гомуми саны 4 меңгә җитә, туларның 200е генә төче сулыкларда һәм туфракта яши.
Кайбер төр багрянкаларның таллом төзелеше югарырак төзелешле көрән суүсемнәр тәне белән охшаш. Аларның катламасы күп күзәнәкле тармакланган җепләрдән торган куак рәвешендә, сирәк кенә 2 м га кадәр җиткән пластинкасыман яки яфраксыман.
Кызыл суүсемнәр диңгез төбенә якын катламнарда таралган үсемлекләр дөньясының (бентосның) иң зур төркемен тәшкил итәләр. Көнчыгыш Азия илләрендә багрянкаларның кайбер төрләрен үрчетеп, азык итеп файдаланалар. Күбесе микробиологиядә микроблар үрчетү өчен туклыклы тирәлек сыйфатында кулланыла торган агар-агар һәм йод табу өчен чимал булып хезмәт итә. Норвегиядә су күтәрелә һәм чигенә торган зонада (литораль) кызыл суүсемнәр утырып калган җирләрне, су чигенгән вакытта, көтүлек итеп файдаланалар.
↑ Integrated Taxonomic Information System — 1996.சிவப்புப் பாசி (red algae) என்பது அல்கா வகைகளில் ஒன்றாகும். இது ரோடோபைட்டா பிரிவுக்குரிய (Division rhodophyta) அல்கா அங்கத்தவர்களை உள்ளடக்கியது. சிவப்பு அல்காக்கள் ஏனைய அல்காக்கள் போலவே ஒளித்தற்போசணிகளாகும்.[1] இவை மெய்க்கருவுயிரி கல ஒழுங்கமைப்பைக் காட்டுகின்றன. சிவப்பு அல்காக்களின் 5000-6000 வரையான இனங்கள் அறியப்பட்டுள்ளன. இயல்புகள்:
சிவப்புப் பாசி (red algae) என்பது அல்கா வகைகளில் ஒன்றாகும். இது ரோடோபைட்டா பிரிவுக்குரிய (Division rhodophyta) அல்கா அங்கத்தவர்களை உள்ளடக்கியது. சிவப்பு அல்காக்கள் ஏனைய அல்காக்கள் போலவே ஒளித்தற்போசணிகளாகும். இவை மெய்க்கருவுயிரி கல ஒழுங்கமைப்பைக் காட்டுகின்றன. சிவப்பு அல்காக்களின் 5000-6000 வரையான இனங்கள் அறியப்பட்டுள்ளன. இயல்புகள்:
இவற்றின் கலச்சுவர் செல்லுலோசு மற்றும் ஏகாரால் ஆனது. அனைத்தும் பல்கல அங்கிகள். பொதுவாக கடல் வாழ்க்கைக்குரியன. சில நன்னீர் வாழ் இனங்களும் அறியப்பட்டுள்ளன. இவற்றில் நிறப்பொருட்களாக பச்சையம் a, பச்சையம் d, கரோட்டீன், பைக்கோசயனின், பைக்கோ எரித்திரின் என்பவை உள்ளன. இவற்றிலுள்ள பைக்கோ எரித்திரின் நிறப்பொருளே சிவப்பு அல்காக்களுக்குச் சிவப்பு நிறத்தை வழங்குகின்றது. வாழ்க்கை வட்டத்தில் எந்தவொரு நிலையிலும் சவுக்குமுளை இருப்பதில்லை. கலங்களில் சேமிப்புணவாக புளோரிடியன் மாப்பொருள் காணப்படும். இவை இலிங்க முறை இனப்பெருக்கத்தைக் காண்பிப்பதுடன் சந்ததிப் பரிவிருத்தியையும் காண்பிக்கின்றன.
Red algae, or Rhodophyta (/roʊˈdɒfɪtə/, /ˌroʊdəˈfaɪtə/; from Ancient Greek ῥόδον (rhódon) 'rose', and φυτόν (phutón) 'plant'), are one of the oldest groups of eukaryotic algae.[3] The Rhodophyta comprises one of the largest phyla of algae, containing over 7,000 currently recognized species with taxonomic revisions ongoing.[4] The majority of species (6,793) are found in the Florideophyceae (class), and mostly consist of multicellular, marine algae, including many notable seaweeds.[4][5] Red algae are abundant in marine habitats but relatively rare in freshwaters.[6] Approximately 5% of red algae species occur in freshwater environments, with greater concentrations found in warmer areas.[7] Except for two coastal cave dwelling species in the asexual class Cyanidiophyceae, there are no terrestrial species, which may be due to an evolutionary bottleneck in which the last common ancestor lost about 25% of its core genes and much of its evolutionary plasticity.[8][9]
The red algae form a distinct group characterized by having eukaryotic cells without flagella and centrioles, chloroplasts that lack external endoplasmic reticulum and contain unstacked (stroma) thylakoids, and use phycobiliproteins as accessory pigments, which give them their red color.[10] But despite their name, red algae can vary greatly in color from bright green, soft pink, resembling brown algae, to shades of red and purple, and may be almost black at greater depths.[11][12] Unlike green algae, red algae store sugars outside the chloroplasts as floridean starch, a type of starch that consists of highly branched amylopectin without amylose,[13] as food reserves outside their plastids. Most red algae are also multicellular, macroscopic, marine, and reproduce sexually. The life history of red algae is typically an alternation of generations that may have three generations rather than two.[14] The coralline algae, which secrete calcium carbonate and play a major role in building coral reefs, belong here. Red algae such as dulse (Palmaria palmata) and laver (nori/gim) are a traditional part of European and Asian cuisines and are used to make other products such as agar, carrageenans and other food additives.[15]
Chloroplasts probably evolved following an endosymbiotic event between an ancestral, photosynthetic cyanobacterium and an early eukaryotic phagotroph.[16] This event (termed primary endosymbiosis) resulted in the origin of the red and green algae, and the glaucophytes, which make up the oldest evolutionary lineages of photosynthetic eukaryotes.[17] A secondary endosymbiosis event involving an ancestral red alga and a heterotrophic eukaryote resulted in the evolution and diversification of several other photosynthetic lineages such as Cryptophyta, Haptophyta, Stramenopiles (or Heterokontophyta), and Alveolata.[17] In addition to multicellular brown algae, it is estimated that more than half of all known species of microbial eukaryotes harbor red-alga-derived plastids.[18]
Red algae are divided into the Cyanidiophyceae, a class of unicellular and thermoacidophilic extremophiles found in sulphuric hot springs and other acidic environments,[19] an adaptation partly made possible by horizontal gene transfers from prokaryotes,[20] with about 1% of their genome having this origin,[21] and two sister clades called SCRP (Stylonematophyceae, Compsopogonophyceae, Rhodellophyceae and Porphyridiophyceae) and BF (Bangiophyceae and Florideophyceae), which are found in both marine and freshwater environments. The SCRP clade are microalgae, consisting of both unicellular forms and multicellular microscopic filaments and blades. The BF are macroalgae, seaweed that usually do not grow to more than about 50 cm in length, but a few species can reach lengths of 2 m.[22] Most rhodophytes are marine with a worldwide distribution, and are often found at greater depths compared to other seaweeds. While this was formerly attributed to the presence of pigments (such as phycoerythrin) that would permit red algae to inhabit greater depths than other macroalgae by chromatic adaption, recent evidence calls this into question (e.g. the discovery of green algae at great depth in the Bahamas).[23] Some marine species are found on sandy shores, while most others can be found attached to rocky substrata.[24] Freshwater species account for 5% of red algal diversity, but they also have a worldwide distribution in various habitats;[7] they generally prefer clean, high-flow streams with clear waters and rocky bottoms, but with some exceptions.[25] A few freshwater species are found in black waters with sandy bottoms [26] and even fewer are found in more lentic waters.[27] Both marine and freshwater taxa are represented by free-living macroalgal forms and smaller endo/epiphytic/zoic forms, meaning they live in or on other algae, plants, and animals.[10] In addition, some marine species have adopted a parasitic lifestyle and may be found on closely or more distantly related red algal hosts.[28][29]
In the classification system of Adl et al. 2005, the red algae are classified in the Archaeplastida, along with the glaucophytes and green algae plus land plants (Viridiplantae or Chloroplastida). The authors use a hierarchical arrangement where the clade names do not signify rank; the class name Rhodophyceae is used for the red algae. No subdivisions are given; the authors say, "Traditional subgroups are artificial constructs, and no longer valid."[30] Many subsequent studies provided evidence that is in agreement for monophyly in the Archaeplastida (including red algae).[31][32][33][34] However, other studies have suggested Archaeplastida is paraphyletic.[35][36] As of January 2011, the situation appears unresolved.
Below are other published taxonomies of the red algae using molecular and traditional alpha taxonomic data; however, the taxonomy of the red algae is still in a state of flux (with classification above the level of order having received little scientific attention for most of the 20th century).[37]
A major research initiative to reconstruct the Red Algal Tree of Life (RedToL) using phylogenetic and genomic approach is funded by the National Science Foundation as part of the Assembling the Tree of Life Program.
Some sources (such as Lee) place all red algae into the class "Rhodophyceae". (Lee's organization is not a comprehensive classification, but a selection of orders considered common or important.[39])
A subphylum - Proteorhodophytina - has been proposed to encompass the existing classes Compsopogonophyceae, Porphyridiophyceae, Rhodellophyceae and Stylonematophyceae.[40] This proposal was made on the basis of the analysis of the plastid genomes.
Over 7,000 species are currently described for the red algae,[4] but the taxonomy is in constant flux with new species described each year.[37][38] The vast majority of these are marine with about 200 that live only in fresh water.
Some examples of species and genera of red algae are:
Red algal morphology is diverse ranging from unicellular forms to complex parenchymatous and non- parenchymatous thallus.[41] Red algae have double cell walls.[42] The outer layers contain the polysaccharides agarose and agaropectin that can be extracted from the cell walls as agar by boiling.[42] The internal walls are mostly cellulose.[42] They also have the most gene-rich plastid genomes known.[43]
Red algae do not have flagella and centrioles during their entire life cycle. The distinguishing characters of red algal cell structure include the presence of normal spindle fibres, microtubules, un-stacked photosynthetic membranes, phycobilin pigment granules,[44] pit connection between cells, filamentous genera, and the absence of chloroplast endoplasmic reticulum.[45]
The presence of the water-soluble pigments called phycobilins (phycocyanobilin, phycoerythrobilin, phycourobilin and phycobiliviolin), which are localized into phycobilisomes, gives red algae their distinctive color.[46] Their chloroplasts contain evenly spaced and ungrouped thylakoids[47] and contain the pigments chlorophyll a, α- and β-carotene, lutein and zeaxanthin. Their chloroplasts are enclosed in a double membrane, lack grana and phycobilisomes on the stromal surface of the thylakoid membrane.[48]
The major photosynthetic products include floridoside (major product), D‐isofloridoside, digeneaside, mannitol, sorbitol, dulcitol etc.[49] Floridean starch (similar to amylopectin in land plants), a long term storage product, is deposited freely (scattered) in the cytoplasm.[50] The concentration of photosynthetic products are altered by the environmental conditions like change in pH, the salinity of medium, change in light intensity, nutrient limitation etc.[51] When the salinity of the medium increases the production of floridoside is increased in order to prevent water from leaving the algal cells.
Pit connections and pit plugs are unique and distinctive features of red algae that form during the process of cytokinesis following mitosis.[52][53] In red algae, cytokinesis is incomplete. Typically, a small pore is left in the middle of the newly formed partition. The pit connection is formed where the daughter cells remain in contact.
Shortly after the pit connection is formed, cytoplasmic continuity is blocked by the generation of a pit plug, which is deposited in the wall gap that connects the cells.
Connections between cells having a common parent cell are called primary pit connections. Because apical growth is the norm in red algae, most cells have two primary pit connections, one to each adjacent cell.
Connections that exist between cells not sharing a common parent cell are labelled secondary pit connections. These connections are formed when an unequal cell division produced a nucleated daughter cell that then fuses to an adjacent cell. Patterns of secondary pit connections can be seen in the order Ceramiales.[53]
After a pit connection is formed, tubular membranes appear. A granular protein called the plug core then forms around the membranes. The tubular membranes eventually disappear. While some orders of red algae simply have a plug core, others have an associated membrane at each side of the protein mass, called cap membranes. The pit plug continues to exist between the cells until one of the cells dies. When this happens, the living cell produces a layer of wall material that seals off the plug.
The pit connections have been suggested to function as structural reinforcement, or as avenues for cell-to-cell communication and transport in red algae, however little data supports this hypothesis.[54]
The reproductive cycle of red algae may be triggered by factors such as day length.[3] Red algae reproduce sexually as well as asexually. Asexual reproduction can occur through the production of spores and by vegetative means (fragmentation, cell division or propagules production).[55]
Red algae lack motile sperm. Hence, they rely on water currents to transport their gametes to the female organs – although their sperm are capable of "gliding" to a carpogonium's trichogyne.[3] Animals also help with the dispersal and fertilization of the gametes. The first species discovered to do so is the isopod Idotea balthica.[56]
The trichogyne will continue to grow until it encounters a spermatium; once it has been fertilized, the cell wall at its base progressively thickens, separating it from the rest of the carpogonium at its base.[3]
Upon their collision, the walls of the spermatium and carpogonium dissolve. The male nucleus divides and moves into the carpogonium; one half of the nucleus merges with the carpogonium's nucleus.[3]
The polyamine spermine is produced, which triggers carpospore production.[3]
Spermatangia may have long, delicate appendages, which increase their chances of "hooking up".[3]
They display alternation of generations. In addition to a gametophyte generation, many have two sporophyte generations, the carposporophyte-producing carpospores, which germinate into a tetrasporophyte – this produces spore tetrads, which dissociate and germinate into gametophytes.[3] The gametophyte is typically (but not always) identical to the tetrasporophyte.[57]
Carpospores may also germinate directly into thalloid gametophytes, or the carposporophytes may produce a tetraspore without going through a (free-living) tetrasporophyte phase.[57] Tetrasporangia may be arranged in a row (zonate), in a cross (cruciate), or in a tetrad.[3]
The carposporophyte may be enclosed within the gametophyte, which may cover it with branches to form a cystocarp.[57]
The two following case studies may be helpful to understand some of the life histories algae may display:
In a simple case, such as Rhodochorton investiens:
A rather different example is Porphyra gardneri:
The δ13C values of red algae reflect their lifestyles. The largest difference results from their photosynthetic metabolic pathway: algae that use HCO3 as a carbon source have less negative δ13C values than those that only use CO2.[58] An additional difference of about 1.71‰ separates groups intertidal from those below the lowest tide line, which are never exposed to atmospheric carbon. The latter group uses the more 13C-negative CO2 dissolved in sea water, whereas those with access to atmospheric carbon reflect the more positive signature of this reserve.
Photosynthetic pigments of Rhodophyta are chlorophylls a and d. Red algae are red due to phycoerythrin. They contain the sulfated polysaccharide carrageenan in the amorphous sections of their cell walls, although red algae from the genus Porphyra contain porphyran. They also produce a specific type of tannin called phlorotannins, but in a lower amount than brown algae do.
As enlisted in realDB,[59] 27 complete transcriptomes and 10 complete genomes sequences of red algae are available. Listed below are the 10 complete genomes of red algae.
One of the oldest fossils identified as a red alga is also the oldest fossil eukaryote that belongs to a specific modern taxon. Bangiomorpha pubescens, a multicellular fossil from arctic Canada, strongly resembles the modern red alga Bangia and occurs in rocks dating to 1.05 billion years ago.[2]
Two kinds of fossils resembling red algae were found sometime between 2006 and 2011 in well-preserved sedimentary rocks in Chitrakoot, central India. The presumed red algae lie embedded in fossil mats of cyanobacteria, called stromatolites, in 1.6 billion-year-old Indian phosphorite – making them the oldest plant-like fossils ever found by about 400 million years.[71]
Red algae are important builders of limestone reefs. The earliest such coralline algae, the solenopores, are known from the Cambrian period. Other algae of different origins filled a similar role in the late Paleozoic, and in more recent reefs.
Calcite crusts that have been interpreted as the remains of coralline red algae, date to the Ediacaran Period.[72] Thallophytes resembling coralline red algae are known from the late Proterozoic Doushantuo formation.[73]
Chromista and Alveolata algae (e.g., chrysophytes, diatoms, phaeophytes, dinophytes) seem to have evolved from bikonts that have acquired red algae as endosymbionts. According to this theory, over time these endosymbiont red algae have evolved to become chloroplasts. This part of endosymbiotic theory is supported by various structural and genetic similarities.[74]
Red algae have a long history of use as a source of nutritional, functional food ingredients and pharmaceutical substances.[75] They are a source of antioxidants including polyphenols, and phycobiliproteins[76] and contain proteins, minerals, trace elements, vitamins and essential fatty acids.[77][78] Traditionally red algae are eaten raw, in salads, soups, meal and condiments. Several species are food crops, in particular members of the genus Porphyra, variously known as nori (Japan), gim (Korea), 紫菜 (China). Laver and dulse (Palmaria palmata)[79] are consumed in Britain.[80] Some of the red algal species like Gracilaria and Laurencia are rich in polyunsaturated fatty acids (eicopentaenoic acid, docohexaenoic acid, arachidonic acid)[81] and have protein content up to 47% of total biomass.[75] Where a big portion of world population is getting insufficient daily iodine intake, a 150 ug/day requirement of iodine is obtained from a single gram of red algae.[82] Red algae, like Gracilaria, Gelidium, Euchema, Porphyra, Acanthophora, and Palmaria are primarily known for their industrial use for phycocolloids (agar, algin, furcellaran and carrageenan) as thickening agent, textiles, food, anticoagulants, water-binding agents etc.[83] Dulse (Palmaria palmata) is one of the most consumed red algae and is a source of iodine, protein, magnesium and calcium. [84]China, Japan, Republic of Korea are the top producers of seaweeds.[85] In East and Southeast Asia, agar is most commonly produced from Gelidium amansii. These rhodophytes are easily grown and, for example, nori cultivation in Japan goes back more than three centuries.

Cyanidium sp. (Cyanidiophyceae)
.jpg)
Porphyra sp., haploid and diploid (Bangiophyceae)

Gracilaria sp. (Florideophyceae: Gracilariales)

Corallina officinalis sp. (Florideophyceae: Corallinales)

Laurencia sp. (Florideophyceae: Ceramiales)

Some red algae are iridescent when not covered with water
{{cite book}}: CS1 maint: multiple names: authors list (link) {{cite web}}: CS1 maint: url-status (link) Red algae, or Rhodophyta (/roʊˈdɒfɪtə/, /ˌroʊdəˈfaɪtə/; from Ancient Greek ῥόδον (rhódon) 'rose', and φυτόν (phutón) 'plant'), are one of the oldest groups of eukaryotic algae. The Rhodophyta comprises one of the largest phyla of algae, containing over 7,000 currently recognized species with taxonomic revisions ongoing. The majority of species (6,793) are found in the Florideophyceae (class), and mostly consist of multicellular, marine algae, including many notable seaweeds. Red algae are abundant in marine habitats but relatively rare in freshwaters. Approximately 5% of red algae species occur in freshwater environments, with greater concentrations found in warmer areas. Except for two coastal cave dwelling species in the asexual class Cyanidiophyceae, there are no terrestrial species, which may be due to an evolutionary bottleneck in which the last common ancestor lost about 25% of its core genes and much of its evolutionary plasticity.
The red algae form a distinct group characterized by having eukaryotic cells without flagella and centrioles, chloroplasts that lack external endoplasmic reticulum and contain unstacked (stroma) thylakoids, and use phycobiliproteins as accessory pigments, which give them their red color. But despite their name, red algae can vary greatly in color from bright green, soft pink, resembling brown algae, to shades of red and purple, and may be almost black at greater depths. Unlike green algae, red algae store sugars outside the chloroplasts as floridean starch, a type of starch that consists of highly branched amylopectin without amylose, as food reserves outside their plastids. Most red algae are also multicellular, macroscopic, marine, and reproduce sexually. The life history of red algae is typically an alternation of generations that may have three generations rather than two. The coralline algae, which secrete calcium carbonate and play a major role in building coral reefs, belong here. Red algae such as dulse (Palmaria palmata) and laver (nori/gim) are a traditional part of European and Asian cuisines and are used to make other products such as agar, carrageenans and other food additives.
La ruĝalgoj aŭ rodofitoj (latine rhodophyta de la antikva greka ῥόδον rodon, "rozo" kaj φυτόν fitón, "planto") estas unu el la plej malnovaj grupoj de eŭkariotaj algoj, kaj ankaŭ unu el la plej granda, kun ĉirkaŭ 5 000 ĝis 6 000 specioj de plejparte multĉelaj, maraj algoj. La grupo inkluzivas multaj konsiderindajn makroalgojn. Ekzempla genro, inter multaj aliaj, estas bangio en la biologia familio bangiacoj.
La ruĝalgoj aŭ rodofitoj (latine rhodophyta de la antikva greka ῥόδον rodon, "rozo" kaj φυτόν fitón, "planto") estas unu el la plej malnovaj grupoj de eŭkariotaj algoj, kaj ankaŭ unu el la plej granda, kun ĉirkaŭ 5 000 ĝis 6 000 specioj de plejparte multĉelaj, maraj algoj. La grupo inkluzivas multaj konsiderindajn makroalgojn. Ekzempla genro, inter multaj aliaj, estas bangio en la biologia familio bangiacoj.
Las algas rojas o rodófitas (filo Rhodophyta, del griego ῥόδον, «rosa», y φυτόν, «planta») son un importante grupo de algas que comprende unas 7000 especies de una gran diversidad de formas y tamaños.[3] Forman parte de Archaeplastida junto a Glaucophyta (glaucofitas) y Viridiplantae (plantas verdes),[4][5] el cual es equivalente al reino Plantae en varios sistemas de clasificación.[6][7][8] Clásicamente en algunos sistemas de clasificación se agrupan en el reino Protista.[9] Sin embargo hay que tener en cuenta que ese grupo no es válido ya que es un taxón cajón de sastre (un grupo conformado por los eucariotas que no encajaban en ninguno de los otros tres reinos eucarióticos).
Se caracterizan por su inmovilidad debido a la carencia o pérdida evolutiva de flagelos en todas las etapas de su ciclo vital. Sus plastos presentan dos membranas, clorofila a y pigmentos accesorios ficobiliproteínas y carotenoides, los cuales enmascaran el color de la clorofila y le dan el color rojo distintivo de estas algas. Están bien representadas en aguas profundas.
Rhodophyta se origina en el Mesoproterozoico y se divide filogenéticamente en dos clados: Cyanidiophytina y Rhodophytina, los cuales divergieron hace unos 1200 millones de años.[10]
Son un grupo primitivo de pequeñas algas unicelulares esféricas que presentan una coloración similar a las algas glaucófitas que va de verde a verde azulada (o verde cian). Se caracterizan por ser extremófilas debido a su condición termoacidófila (son termófilas e hiperacidófilas), habitando en aguas termales, calderas volcánicas y algunos entornos ácidos producto de la actividad humana, mostrando además resistencia a la presencia de metales tóxicos.
Su reproducción es asexual por división mitótica, formando en ocasiones endosporas en número 2, 4, 8 y a veces hasta 16 células hijas. Son organismos muy simples, y en la mayoría de ocasiones poseen un solo cloroplasto y una sola mitocondria, los cuales se reproducen por división sincronizada entre la célula y estos mismos organelos.
La simplicidad de estas algas estaría en relación con la pérdida evolutiva de genes. Se estima que Cyanidiophytina habría perdido 3 veces más genes que Rhodophytina.[11]
El clado Rhodophytina está conformado por las algas rojas propiamente dicho, con una coloración característica roja dada por el pigmento ficoeritrina. Son prácticamente marinas (solo 164 especies son de agua dulce) y se pueden encontrar en todo tipo de mares. Su ecosistema va desde zonas intermareales hasta zonas muy profundas, dependiendo de la transparencia del agua. Se pueden encontrar a 100 metros de profundidad, alcanzando hasta los 250 metros en casos excepcionales. Son las algas más abundantes en lugares profundos, pues sus pigmentos les permiten captar las longitudes de onda de la luz del Sol que penetran más profundamente en el agua. Estos pigmentos, que absorben la luz azul y reflejan la roja, les dan su característico color rojizo.
Algunas algas rojas, por ejemplo la dulse o el nori, son utilizadas como alimento y usadas para producir agar, carragenanos y otros aditivos alimenticios.
La extensa mayoría de las algas rojas son marinas, aunque existen algunas especies que viven en agua dulce o en el suelo. Son de vida libre, epífitas o epizoicas y se conocen algunas formas parásitas. Algunas especies crecen utilizando como sustrato las algas pardas (que suelen ser más grandes) o sobre las conchas de mejillones y gasterópodos. Algunas son simbiontes de foraminíferos bentónicos, mientras que otras tienen plastos vestigiales y se ven obligadas a parasitar otras rodofitas. En el grupo se incluye muchas macroalgas notables, entre las que se encuentran la mayoría de las algas coralinas que secretan carbonato de calcio y cumplen un papel crucial en la formación de los arrecifes de coral. Estas formas coralinas pueden ser difíciles de distinguir de los corales.
Las algas rojas suelen vivir en la zona litoral relativamente estrecha que bordea la placa continental. Como consecuencia de los pigmentos que presentan, pueden captar la luz a profundidades mayores que otros tipos de algas. Como ejemplo extremo, se han encontrado rodofitas viviendo en las laderas de una montaña submarina a la profundidad de 268 m, donde solo es capaz de penetrar el 0,001 % de la luz de la superficie.[12] Se las encuentra en todas las latitudes, preferentemente en aguas tropicales y templadas, donde constituyen las algas más abundantes. En las aguas polares y subpolares hay pocas especies y dominan las algas verdes y pardas.
Algunas de las especies de algas rojas son unicelulares, pero la mayoría son pluricelulares creciendo en forma de filamentos o láminas membranosas, que suelen tener algún tipo de consolidación, alcanzado el grado de organización seudoparenquimatoso. Pueden existir como cilindros muy finos, ramificados en forma de arbolillo o como láminas enteras o divididas. El talo normalmente se construye mediante la agregación de numerosos filamentos, dando lugar a estructuras cilíndricas o laminares de hasta 1 m de longitud, pero que nunca alcanzan la complejidad de las algas pardas. Su tamaño es también menor al de las algas pardas más grandes. El crecimiento normalmente se produce mediante la división de la célula apical, que puede ser multinucleada.
Adicionalmente como estructuras especializadas presentan rizoides que van desde filamentos simples hasta discos formados por la agrupación convergente de los filamentos en la base del alga. Los zarcillos son transformaciones de las ramas terminales cuya función es la adhesión a otras ramas de la misma alga, a algas mayores o a otros elementos. Como estructuras reproductoras se presentan esporangios (que forman esporas) y gametangios (que forman los gametos). Existen también células vesiculares o secretoras de sustancias que presentan una gran diversidad en su composición química.
En la mayoría de las algas rojas existen poros septales o de conexión (sinapsis) entre las células. Puesto que el crecimiento apical es la norma en las algas rojas, la mayoría de las células tienen dos poros primarios, uno a cada célula adyacente. Los poros que no se derivan de una división celular se denominan secundarias y se producen cuando se fusionan células adyacentes. Después de la formación del poro, la conexión citoplasmática es bloqueada mediante la formación de un tapón de proteínas que puede funcionar tanto como refuerzo estructural como de vía de comunicación de célula a célula. En el refuerzo estructural también tienen importancia los mucílagos que se acumulan en las paredes celulares, compactando tanto los filamentos axiales con los contiguos.
Las rodofitas son fotosintéticas y sus cloroplastos (llamados rodoplastos) presentan algunas características específicas de su grupo. Están rodeados por dos membranas y se supone procedentes de la endosimbiosis primaria de una cianobacteria, al igual que los de las plantas verdes y las glaucofitas. Sin embargo, presentan características diferenciadas, por lo que las rodofitas se clasifican como grupo aparte.
Los rodoplastos contienen clorofila a, además de los pigmentos accesorios ficobiliproteínas (ficoeritrina y ficocianina) y carotenoides. Las rodofitas son los únicos protistas fotosintéticos que presentan ficoeritrina. Antiguamente se creía que algunas especies contenían también clorofila d, pero recientemente se ha descubierto que esta clorofila procede de una cianobacteria, Acaryochloris marina, que vive epifita sobre estas algas.[13] Los tilacoides son solitarios, sin apilar y algunas veces hay uno o dos tilacoides periféricos. Se caracterizan por la presencia de ficobilisomas que contienen ficobiliproteínas en la superficie de los tilacoides y ausencia de almidón en los cloroplastos, usando como material de reserva almidón de florídeas (de estructura semejante a la amilopectina) extraplastidial, en gránulos en el citoplasma, próximos al cloroplasto. Pueden presentar o no pirenoides, usualmente en los grupos basales en el primer caso. Los pigmentos les confieren un color rojo o rojizo, generalmente.
Las rodofitas son organismos siempre inmóviles, pues carecen de células flageladas en todas las etapas de su ciclo vital. Sus células también carecen de centriolos, centrosomas y de cualquier otra estructura que implique una organización 9+2 de microtúbulos. Las células pueden ser multinucleadas. Las algas rojas tienen paredes celulares dobles.[14] Las paredes externas contienen los polisacáridos agarosa y agaropectina que se puede extraer de las paredes celulares por ebullición para obtener agar. Las paredes internas, adheridas al citoplasma, son en su mayoría de celulosa.
Aunque en algunas especies puede faltar la sexualidad, usualmente presentan alternancia de generaciones, que pueden ser dos o tres.[15] Son organismos en los que predomina la fase haploide, a la que se añaden una o dos fases diploides durante su ciclo vital. La reproducción sexual es por oogamia, con células especializadas, carpogonios y espermacios. Las células que hacen la función de espermatozoides carecen de flagelos y no puede nadar, por lo que son llevadas por las corrientes de agua.
El ciclo vital puede ser de dos tipos:
En la siguiente galería se muestran las tres generaciones de Polysiphonia.
Gametófito masculino
Gametófito femenino
Carposporocito sobre el gametófito femenino
Tetrasporocito
Los diversos eucariotas que componen las algas rojas han sido foco de numerosas investigaciones recientes y queda una rica fuente de especies pequeñas aún no descritas por la taxonomía tradicional. Los estudios moleculares ubican a las algas en Archaeplastida (= Primoplantae, Plantae sensu lato); sin embargo, la clasificación supraordinal se ha limitado al debate del nivel clase vs. subclase para los dos subgrupos reconocidos, uno de los cuales es ampliamente reconocido como parafilético. Este limitado foco generalmente ha ocultado en gran medida la necesidad de modificación de la clasificación algal.
Abajo hay dos taxonomías de algas rojas publicadas que son válidas, aunque necesariamente ninguna de las dos tiene que ser usada, ya que la taxonomía algal está todavía en continuo desarrollo. Nótese también que hay un debate científico continuo de si las rodofitas deberían ser incluidas en el reino Protista o en el reino Plantae. Estos dos sistemas de clasificación, en los cuales las algas se ubican en el reino Plantae, son mostrados en la tabla.
Porphyra haploide y diploide (Bangiophyceae)
Chondrus crispus (Florideophyceae, orden Gigartinales)
Gracilaria (Florideophyceae, orden Gracilariales)
Las algas rojas o rodófitas (filo Rhodophyta, del griego ῥόδον, «rosa», y φυτόν, «planta») son un importante grupo de algas que comprende unas 7000 especies de una gran diversidad de formas y tamaños. Forman parte de Archaeplastida junto a Glaucophyta (glaucofitas) y Viridiplantae (plantas verdes), el cual es equivalente al reino Plantae en varios sistemas de clasificación. Clásicamente en algunos sistemas de clasificación se agrupan en el reino Protista. Sin embargo hay que tener en cuenta que ese grupo no es válido ya que es un taxón cajón de sastre (un grupo conformado por los eucariotas que no encajaban en ninguno de los otros tres reinos eucarióticos).
Se caracterizan por su inmovilidad debido a la carencia o pérdida evolutiva de flagelos en todas las etapas de su ciclo vital. Sus plastos presentan dos membranas, clorofila a y pigmentos accesorios ficobiliproteínas y carotenoides, los cuales enmascaran el color de la clorofila y le dan el color rojo distintivo de estas algas. Están bien representadas en aguas profundas.
Rhodophyta se origina en el Mesoproterozoico y se divide filogenéticamente en dos clados: Cyanidiophytina y Rhodophytina, los cuales divergieron hace unos 1200 millones de años.
Punavetikad (Rhodophyceae) on ainuke punavetiktaimede hõimkonda kuuluv vetikate klass.
Suur osa punavetikaid on hulkraksed. Neid võib leida nii mage- kui merevees. Punavetikaid on erinevate allikate järgi teada 4000 kuni 10 000 liiki.
Läänemeres elutseb madala soolsuse tõttu vaid ligi 160 punavetikaliiki.[1]
Mitmed punavetikaliigid omavad töönduslikku tähtsust. Neis leiduvaid pigmente kasutatakse meditsiinidiagnostikas, molekulaarbioloogias ja toiduainetööstuses ning geelistuvaid polüsahhariide nii mikrobioloogias kui ka farmaatsia- ja kosmeetikatööstuses.[1]
Punavetikad (Rhodophyceae) on ainuke punavetiktaimede hõimkonda kuuluv vetikate klass.
Suur osa punavetikaid on hulkraksed. Neid võib leida nii mage- kui merevees. Punavetikaid on erinevate allikate järgi teada 4000 kuni 10 000 liiki.
Läänemeres elutseb madala soolsuse tõttu vaid ligi 160 punavetikaliiki.
Mitmed punavetikaliigid omavad töönduslikku tähtsust. Neis leiduvaid pigmente kasutatakse meditsiinidiagnostikas, molekulaarbioloogias ja toiduainetööstuses ning geelistuvaid polüsahhariide nii mikrobioloogias kui ka farmaatsia- ja kosmeetikatööstuses.
Punavetiktaimed (Rhodophyta) on vetikate hõimkond, kuhu kuulub umbes 4000 liiki. Nende nimetus tuleneb vetikatele iseloomulikust talluse värvusest. Enamik punavetikaid on hulkraksed makroskoopilised organismid.
Punavetikad sisaldavad lisaks klorofüllile (klorofüll a ja d), ksantofüllidele ja karoteenidele ka ainuomaseid pigmente: fükoerütriin ja fükotsüaan. Nende kahe pigmendi tõttu saavad punavetikad elutegevuseks kasutada väga nõrka valgust, suutes kasvada meredes ja ookeanides maksimaalselt 200 meetri sügavusel, optimaalne kasvuala on 40–60 meetrit. Enamikule punavetiktaimede kasvuks on vaja suhteliselt ühtlast aastast temperatuuri. Punavetiktaimede pikkus ulatub mõnest millimeetrist kuni paari meetrini.
Suur hulk punavetikaliike elab troopilistes meredes.
Hõimkonda kuulub üks klass – punavetikad (Rhodophyceae). Varem arvati sinna kaks klassi: Florideophyceae ja Bangiophyceae.
Punavetiktaimed paljunevad nii suguliselt kui suguta. Viimase puhul toimub see aplanospooridega. Sugulisel paljunemisel ühineb viburita spermaatsium oogooniumis valminud ühe munarakuga. Pärast viljastumist areneb kohe sügoot.
Punavetiktaimedel on rakukestades sisalduvate kallerduvate ainete tõttu majanduslik tähtsus. Tuntuim kallerduv aine on agar, mida kasutatakse meditsiinis, mikrobioloogias ja mitmes tööstusharus.
Punavetiktaimed (Rhodophyta) on vetikate hõimkond, kuhu kuulub umbes 4000 liiki. Nende nimetus tuleneb vetikatele iseloomulikust talluse värvusest. Enamik punavetikaid on hulkraksed makroskoopilised organismid.
Punavetikad sisaldavad lisaks klorofüllile (klorofüll a ja d), ksantofüllidele ja karoteenidele ka ainuomaseid pigmente: fükoerütriin ja fükotsüaan. Nende kahe pigmendi tõttu saavad punavetikad elutegevuseks kasutada väga nõrka valgust, suutes kasvada meredes ja ookeanides maksimaalselt 200 meetri sügavusel, optimaalne kasvuala on 40–60 meetrit. Enamikule punavetiktaimede kasvuks on vaja suhteliselt ühtlast aastast temperatuuri. Punavetiktaimede pikkus ulatub mõnest millimeetrist kuni paari meetrini.
Suur hulk punavetikaliike elab troopilistes meredes.
Alga gorria (Rhodophyta) alga eukarioto mota da. Alga gorriak organismo protistak dira, eta gehienak, zelulanitzak. Forma aldakorra eta biomasa txikia duten landareak dira. 4000 espezie inguru deskribatu dira eta Euskal Herriko kostaldean asko ikus daitezke. Gehienak itsastar bentikoak dira, askotariko substratuetan aurki daitezkeenak eta askotan epifitoak izaten dira, hau da, beste alga batzuei atxikita hazten dira.
Oso ezagunak dira Euskal Herriko kostaldean, askotan egoten baitira Gelidium sesquipedale generoko algak eguzkitan lehortzen. Izan ere, alga hau oso aberatsa da Agar-agar izeneko pektina-motan eta aipatu substantzia oso preziatua baita elikagai, botikagintza eta kosmetika industrian.
Flageloak: dibisio honetako algek ez dute flagelorik
Ernalketa: gametozistegamia bitartez ematen da alga gorri gehienen arteko ugalketa. Hau da, ernalketa gametoziste mailan ematen da, ugal-zelulei babesa emate aldera.
Zelula horma: pektina ugaridun pektozelulosazko pareta izan ohi dute. Alga talde honetan bi pektina mota aurki daitezke: agarra, Gelidium generoan agertzen dena, esaterako; eta korragenoa, Chondrus generoan, adibidez. Batzuetan pareta kaltzifikatu egiten da Lithophyllum eta Corallina generoetan gertatzen den bezala.
Pigmentuak: alga hauetan a- eta b-klorofilak, α- eta β-karotenoak, xantofilak (luteina bereziki) eta fikobilinak agertu ohi dira.
Plastoak: pareta bikoitza dute, tilakoideak isolatuta agertu ohi dira eta fikobilisomak eduki ohi dituzte.
Erreserba: alga gorrien erreserba-substantzia errodamilioia izaten da eta estraplastidialki metatzen da.
Alga gorria (Rhodophyta) alga eukarioto mota da. Alga gorriak organismo protistak dira, eta gehienak, zelulanitzak. Forma aldakorra eta biomasa txikia duten landareak dira. 4000 espezie inguru deskribatu dira eta Euskal Herriko kostaldean asko ikus daitezke. Gehienak itsastar bentikoak dira, askotariko substratuetan aurki daitezkeenak eta askotan epifitoak izaten dira, hau da, beste alga batzuei atxikita hazten dira.
Oso ezagunak dira Euskal Herriko kostaldean, askotan egoten baitira Gelidium sesquipedale generoko algak eguzkitan lehortzen. Izan ere, alga hau oso aberatsa da Agar-agar izeneko pektina-motan eta aipatu substantzia oso preziatua baita elikagai, botikagintza eta kosmetika industrian.
Rhodophytes, Algues rouges
Les algues rouges, ou Rhodophytes (division des Rhodophyta), sont un grand taxon d'algues pour la plupart marines et pour la plupart multicellulaires (la plupart sont sessiles, c'est-à-dire qu'elles se développent fixées sur un substrat quelconque).
Elles sont caractérisées par une composition pigmentaire avec un seul type de chlorophylle, la chlorophylle a, des caroténoïdes et des pigments caractéristiques, les phycobiliprotéines. Elles seraient apparues pendant l'Ectasien du Mésoprotérozoïque il y a 1,2 à 1,4 milliard d'années, et seraient de ce fait les seconds plus anciens organismes vivants pluricellulaires découverts entre le Gabonionta apparu à l'Orosirien il y a environ 2 100 Ma, et la faune de l'Édiacarien datant d'environ 575 Ma, sans qu'un lien puisse être démontré entre ces trois « apparitions ».
Les rhodophytes contiennent des pigments rencontrés chez les autres végétaux, la chlorophylle a et des caroténoïdes, mais leur originalité consiste dans la présence de phycobilisomes comprenant les phycobiliprotéines : allophycocyanine (bleu), phycocyanine (bleu) et phycoérythrine qui donne la couleur rouge. Le chloroplaste peut alors être appelé rhodoplaste. L’organisation du chloroplaste est la suivante : les thylakoïdes sont libres et ne forment pas de grana (ceci est lié à la présence des phycobilisomes à la surface des thylakoïdes), ils sont répartis concentriquement dans le chloroplaste mais occupent tout l’espace, contrairement à ceux des Glaucophytes et des Cyanobactéries.
La pigmentation (rapport des pigments présents) dépend à un degré remarquable de la longueur d'onde de la lumière qui atteint l'algue. En profondeur, les algues accumulent une grande quantité de phycoérythrine, pigment qui peut absorber la lumière à cette profondeur. Si les algues sont en surface, ce pigment rouge diminue, elles deviennent plus vertes malgré leur appellation. Cependant les quantités de chlorophylles demeurent inchangées que l’algue soit en surface ou en profondeur; on parle alors d'adaptation chromatique.
L’amidon est stocké sous forme de vésicules dans le cytoplasme (et non dans le plaste comme chez les plantes et algues vertes) : l'amidon floridéen (aussi appelé rhodamylon).
La floridoside (2-O-α-d-galactopyranosylglycerol) est un glucide, hétéroside, de formule C9H18O8[1]. C'est une molécule propre aux algues rouges, synthétisée par toutes les algues rouges, excepté les membres de l’ordre des Ceramiales[2]. Elle est le produit principal de la fixation du carbone par la photosynthèse et, pourra être utilisée par la suite pour la synthèse de macromolécules (amidon, polysaccharides solubles…)[3]. C'est également un osmoprotecteur dont la concentration intracytoplasmique augmente en réponse à un stress hyperosmotique[4],[5],[6],[7]. La floridoside semble être aussi un précurseur de la paroi polysaccharidique chez quelques espèces d'algues rouges[8].
Les Rhodophytes sont caractérisées par une organisation cellulaire de type eucaryote, mais il faut noter l’absence de cils et de flagelles (à aucun stade du développement), par un gamète mâle non-mobile et par un cycle de vie digénétique ou trigénétique (comportant successivement des générations de gamétophytes, de carposporophytes dans les cycles trigénétiques et de (tétra)sporophytes).
Leur paroi pectocellulosique est de composition complexe. Elle contient de la cellulose associé dans sa partie interne à d'autres polysaccharides parfois en quantité importante, agar-agar donnant une texture flexible et glissante, et dans sa partie externe elle est associée à des carbonate de calcium ce qui donne des algues dures et pierreuses : elle participe à la stabilisation des massifs coralliens en cimentant les débris coraux. La structure calcaire demeure même après la mort de l'algue, formant parfois des masses rocheuse imposantes (corallinaceae).
La grande majorité des algues rouges est rencontrée dans le milieu marin.
Chondrus crispus et Mastocarpus stellatus (Gigartina stellata) sont souvent associées et vivent fixées au rocher. Ces deux espèces sont récoltées afin d'extraire les carraghénanes utilisés comme gélifiant dans l'industrie agroalimentaire.
Certaines algues rouges comme Porphyra sont consommées en Extrême-Orient. Porphyra est cultivée au Japon : sous le nom de nori, elle intervient dans la confection des sushis.
Certaines algues rouges sont abondantes dans les récifs de coraux. Les Corallinaceae produisent une enveloppe extracellulaire de carbonate de calcium et peuvent participer à la construction du récif de corail.
Certaines algues rouges sont très résistantes à des conditions extrêmes et sont donc extrêmophiles : par exemple, Cyanidium caldarium vit à un pH inférieur à 1 dans les sources acides.
Le plus ancien fossile multicellulaire d'eucaryote découvert à ce jour serait[9] une algue rouge filamenteuse vieille de 1,7 milliard d'années, trouvée dans les monts Vindhya[10]. Les rhodophytes sont ainsi apparues après les cyanobactéries (anciennement appelées algues bleues) qui avaient précédemment provoqué, par l'action cumulée de la photosynthèse, le bouleversement planétaire appelé la Grande Oxydation. Elles ont profité des nouvelles conditions écologiques, notamment de l'apparition de la couche d'ozone stratosphérique qui filtre la plus grande partie du rayonnement solaire ultraviolet et qui protège de son effet biocide, pour occuper dans les océans une niche écologique plus proche de la surface. Cette nouvelle zone de développement de la vie étant cependant encore trop profonde pour que la seule chlorophylle suffise à l'autonomie énergétique, ce sont, à l'instar des cyanobactéries, les pigments complémentaires synthétisés par les algues rouges qui leur ont permis de pallier un environnement encore trop sombre et de capter l'énergie d'autres fréquences lumineuses du rayonnement solaire[11].
Les termes de Rhodoplantae ou de Rhodobiota (Rhodobiontes) proposés assez récemment par certains auteurs sont discutés. La dernière phylogénie en date propose de ne retenir que le terme de Rhodophyta (Rhodophytes) pour l'embranchement, avec deux sous-embranchements : Cyanidiophytina et Rhodophytina (Yoon et al., 2006).
Selon AlgaeBase (29 févr. 2012)[12] :
Selon NCBI (29 févr. 2012)[13] :
Selon World Register of Marine Species (29 févr. 2012)[14] :
Selon ITIS (29 févr. 2012)[15] :
Selon Paleobiology Database (29 févr. 2012)[16] :
Il existe plusieurs genres incertae sedis selon AlgaeBase (3 févr. 2019)[17] :
Il existe 5 génomes d'algue rouge séquencés dont 4 publiés en 2013.
Rhodophytes, Algues rouges
Les algues rouges, ou Rhodophytes (division des Rhodophyta), sont un grand taxon d'algues pour la plupart marines et pour la plupart multicellulaires (la plupart sont sessiles, c'est-à-dire qu'elles se développent fixées sur un substrat quelconque).
Elles sont caractérisées par une composition pigmentaire avec un seul type de chlorophylle, la chlorophylle a, des caroténoïdes et des pigments caractéristiques, les phycobiliprotéines. Elles seraient apparues pendant l'Ectasien du Mésoprotérozoïque il y a 1,2 à 1,4 milliard d'années, et seraient de ce fait les seconds plus anciens organismes vivants pluricellulaires découverts entre le Gabonionta apparu à l'Orosirien il y a environ 2 100 Ma, et la faune de l'Édiacarien datant d'environ 575 Ma, sans qu'un lien puisse être démontré entre ces trois « apparitions ».
Grúpa ilchineálach mór plandaí, cosúil le halgaí agus foirmeacha ó chealla aonaracha go dtí coirp mhóra idirdhealaithe. Tagann an dath scothdhearg ón meascán de líocha fótaisintéiseacha, clóraifill a, ficibileaphróitéiní is cairitéineóidigh taobh istigh de na cealla. Tarlaíonn atáirgeadh trí chruthú orgáin ar leith atá gabhálach ar an ngaiméit fhireann taobh istigh den ubh.
A división ou filo Rhodophyta integra o que coñecemos vulgarmente coma algas vermellas. Probabelmente constitúen un dos grupos de algas eucariotas máis antigos. Semella que teñen moitas características comúns coa división Glaucophyta. Son un grupo homoxéneo, xa que só contén unha clase sistemática, Rhodophyceae, e que conta con aproximadamente unhas 5.000 especies de auga mariña na súa meirande parte. As especies dulceacuícolas adoitan atoparse en ambientes lóticos. A maioría das rodofíceas son pluricelulares, e só coñecemos uns once xéneros unicelulares.
Os principais pigmentos fotosintéticos son a clorofila a e d, e as veces incluso poden presentar clorofila e. O pigmento carotenoide por excelencia é a zeaxantina. Mais os pigmentos característicos deste grupo botánico constitúeno as chamadas ficobiliproteínas, que forma parte dos ficobilisomas. Como substancia de reerva empregan o floridósido, e a longo prazo acábano por transformar nunha substancia amilácea chamada amidón de florídeas.
Neste grupo atópanse a maioría das algas que secretan carbonato de calcio e cumpren un rol maior na formación dos arrecifes de coral. Algunhas algas vermellas, por exemplo a dulse ou o nori, son empregadas como alimento e para producir agar, carraxenanos e outros aditivos alimenticios.
As diversas eucariotas que compoñen as algas vermellas foron foco de numerosas investigacións recentes e fica unha rica fonte de especies pequenas aínda non descritas pola taxonomía tradicional. Os estudos moleculares deixan ás algas nas Arqueoplástidas (Plantae sensu lato), mais a clasificación supraordinal limitouse ao debate do nivel clase vs. subclase para os dous subgrupos recoñecidos, un dos cales é considerado como parafilético. Este limitado enfoque ocultou en gran medida a necesidade de modificación da clasificación algal.
Abaixo hai dúas taxonomías de algas vermellas publicadas que son válidas, aínda que necesariamente ningunha das dúas ha ser definitiva, xa que a taxonomía algal está aínda en continuo desenvolvemento. Nótese tamén que hai un debate científico continuo sobre se as Rodofitas han ser incluídas no Reino Protista ou no Reino plantae. Estes dous sistemas de clasificación, nos cales as algas ubícanse no Reino Plantae, son amosados na táboa.
As Rodofitas posúen tres tipos de pigmentos:
A maioría das especies mariñas ou dulcicuícolas forman parte do bentos, e se reparten por case tódolos mares do planeta. Especies como Bangia adaptáronse a viviren na zona intermareal, aínda que no verán sobrevive na zona infralitoral en fase Conchocelis. As algas vermellas poden ser tamén parasitas. Distinguimos dous tipos de parasitos:
Neste grupo atópanse a maioría das algas que secretan carbonato de calcio e cumpren un rol maior na formación dos arrecifes de coral. Algunhas algas vermellas, por exemplo a dulse ou o nori, son empregadas como alimento e para producir agar, carraxenanos e outros aditivos alimenticios.
A división ou filo Rhodophyta integra o que coñecemos vulgarmente coma algas vermellas. Probabelmente constitúen un dos grupos de algas eucariotas máis antigos. Semella que teñen moitas características comúns coa división Glaucophyta. Son un grupo homoxéneo, xa que só contén unha clase sistemática, Rhodophyceae, e que conta con aproximadamente unhas 5.000 especies de auga mariña na súa meirande parte. As especies dulceacuícolas adoitan atoparse en ambientes lóticos. A maioría das rodofíceas son pluricelulares, e só coñecemos uns once xéneros unicelulares.
Os principais pigmentos fotosintéticos son a clorofila a e d, e as veces incluso poden presentar clorofila e. O pigmento carotenoide por excelencia é a zeaxantina. Mais os pigmentos característicos deste grupo botánico constitúeno as chamadas ficobiliproteínas, que forma parte dos ficobilisomas. Como substancia de reerva empregan o floridósido, e a longo prazo acábano por transformar nunha substancia amilácea chamada amidón de florídeas.
Neste grupo atópanse a maioría das algas que secretan carbonato de calcio e cumpren un rol maior na formación dos arrecifes de coral. Algunhas algas vermellas, por exemplo a dulse ou o nori, son empregadas como alimento e para producir agar, carraxenanos e outros aditivos alimenticios.
Crvene alge (Rhodophyta, od grčkih riječi ῥόδον = crven, i φυτόν = biljka) su velika skupina najčešće višestaničnih, morskih algi, uključujući i mnoge morske trave.
Postoji između 5000 i 6000 živućih vrsta[1], dok neki drugi izvori govore da postoji više od 10.000 vrsta u ovom koljenu algi[2]. Jedna su od najstarijih skupina eukariotskih algi.[3] Samo dvadesetak rodova nastanjuje i vode na kopnu. Većinom žive pričvršćene za kamenitu podlogu, ali i za školjke, te vrlo rijetko kao epifiti pričvršćene za druge alge. Vrlo su ostjetljive na promjene temperature. Dosta su sitnije od smeđih algi, pa se na područjima jakih valova ne mogu održati, stoga žive u mirnim dubokim vodama.
Najstariji fosilni nalazi ovih algi datiraju iz mezoproterozoika (prije otprilike milijardu godina).
Crvene alge su jako posebna skupina algi. Imaju eukariotsku stanicu bez biča i centriola. Njihova stanična stijenka ima unutarnji sloj sastavljen od celuloze, dok je vanjski sloj građen od pektina, te je sluzast. Imaju jednu jezgru, rijetke su one koje u starosti imaju više jezgara. Kao rezerva za hranu služi im ugljikohidrat floridejski škrob (između glikogena i škroba), koji se u obliku zrnaca nalazi na površini rodoplasta. Crvenu boju daju im fikobilini. Kloroplasti nemaju endoplazmatski retikulum, a građeni su od paralelno naslaganih vrećica, tilakoida.
Crvene alge se razmnožavaju nespolno, spolno ili izmjenom spolnog i nespolnog razmnožavanja (izmjenom generacija). Ciklus razmnožavanja može biti izazvan različitim čimbenicima, kao što je duljina dana[3]. Nemaju pokretnih stanica, spore i gamete su nepokretne, te se ove alge oslanjaju na vodene struje kao prijenosnik stanica u organe. Vegetativno razmnožavanje se ostvaruje diobom (kod jednostaničnih oblika), raskidanjem kolonije ili otkinutim dijelovima talusa. Također postoji razmnožavanje sporama, kod kojeg se stvaraju određene vrste spora (monospore, tetraspore i polispore) Spolni način razmnožavanja predstavljen je netipičnom oogamijom.
Mnoge od koraljnih algi, koje izlučuju kalcijev karbonat i igraju veliku ulogu u gradnji koraljnih otoka, pripadaju ovoj skupini. Iz staničnih stijenki nekih vrsta dobivaju se različiti polisaharidi za primjenu u ljekarstvu (npr. irska mahovina). Također, neke alge iz ove skupine su tradicionalni dio europske i azijske kuhinje, npr. dulse i nori, te se koriste za pripravljenje dodataka kao što su agar, karagenan i drugi prehrambeni aditivi. Sadrže veliku količinu vitamina i bjelančevina, te se jednostavno uzgajaju, pa su zbog toga privlačne za kultivaciju, koja je u Japanu počela prije više od 300 godina.
A) Subphylum Cyanidiophytina H.S.Yoon, K.M.Müller, R.G.Sheath, F.D.Ott & D.Bhattacharya
B) Subphylum Eurhodophytina G.W.Saunders & Hommersand
C) Subphylum Proteorhodophytina Muñoz-Gómez, Mejía-Franco, Durnin, Colp, Grisdale, J.M.Archibald & Slamovits
D) Classis Rhodophyta classis incertae sedis
Crvene alge (Rhodophyta, od grčkih riječi ῥόδον = crven, i φυτόν = biljka) su velika skupina najčešće višestaničnih, morskih algi, uključujući i mnoge morske trave.
Postoji između 5000 i 6000 živućih vrsta, dok neki drugi izvori govore da postoji više od 10.000 vrsta u ovom koljenu algi. Jedna su od najstarijih skupina eukariotskih algi. Samo dvadesetak rodova nastanjuje i vode na kopnu. Većinom žive pričvršćene za kamenitu podlogu, ali i za školjke, te vrlo rijetko kao epifiti pričvršćene za druge alge. Vrlo su ostjetljive na promjene temperature. Dosta su sitnije od smeđih algi, pa se na područjima jakih valova ne mogu održati, stoga žive u mirnim dubokim vodama.
Najstariji fosilni nalazi ovih algi datiraju iz mezoproterozoika (prije otprilike milijardu godina).
Alga merah atau Rhodophyta (/roʊˈdɒfᵻtə/ roh-DOF-fit-tə atau /ˌroʊdəˈfaɪtə/ ROH-də-FY-tə; dari Bahasa Yunani Kuno: ῥόδον rhodon, "mawar" and φυτόν phyton, "tumbuhan") adalah salah satu filum dari alga berdasarkan zat warna atau pigmentasinya. Warna merah pada alga ini disebabkan oleh pigmen fikoeritrin dalam jumlah banyak dibandingkan pigmen klorofil, karoten, dan xantofil.
Alga ini pada umumnya bersel banyak (multiseluler) dan makroskopis. Panjangnya antara 10 cm sampai 1 meter dan berbentuk berkas atau lembaran.
Beberapa alga merah memiliki nilai ekonomi sebagai bahan makanan (sebagai pelengkap minuman penyegar ataupun sebagai bahan baku agar-agar). Alga merah sebagai bahan makanan memiliki kandungan serat lunak yang baik bagi kesehatan usus.
Sebagian besar alga merah hidup di laut, banyak terdapat di laut tropika. Sebagian kecil hidup di air tawar yang dingin dengan aliran deras dan banyak oksigen. Selain itu ada pula yang hidup di air payau. Alga merah yang banyak ditemukan di laut dalam adalah Gelidium dan Gracilaria, sedang Euchema spinosum menyukai laut dangkal.
Alga merah berkembangbiak secara vegetatif dan generatif.
Alga merah dapat menyediakan makanan dalam jumlah banyak bagi ikan dan hewan lain yang hidup di laut. Jenis ini juga menjadi bahan makanan bagi manusia misalnya Chondrus crispus (lumut Irlandia) dan beberapa genus Porphyra. Chondrus crispus dan Gigortina mamilosa menghasilkan karagen yang dimanfaatkan untuk penyamak kulit, bahan pembuat krem, dan obat pencuci rambut. Alga merah lain seperti Gracilaria lichenoides, Euchema spinosum, Gelidium dan Agardhiella dibudidayakan karena menghasilkan bahan serupa gelatin yang dikenal sebagai agar-agar. Gel ini digunakan oleh para peneliti sebagai medium biakan bakteri dan fase padat pada elektroforesis gel, untuk pengental dalam banyak makanan, perekat tekstil, sebagai obat pencahar (laksatif), atau sebagai makanan penutup.
Alga merah atau Rhodophyta (/roʊˈdɒfᵻtə/ roh-DOF-fit-tə atau /ˌroʊdəˈfaɪtə/ ROH-də-FY-tə; dari Bahasa Yunani Kuno: ῥόδον rhodon, "mawar" and φυτόν phyton, "tumbuhan") adalah salah satu filum dari alga berdasarkan zat warna atau pigmentasinya. Warna merah pada alga ini disebabkan oleh pigmen fikoeritrin dalam jumlah banyak dibandingkan pigmen klorofil, karoten, dan xantofil.
Alga ini pada umumnya bersel banyak (multiseluler) dan makroskopis. Panjangnya antara 10 cm sampai 1 meter dan berbentuk berkas atau lembaran.
Beberapa alga merah memiliki nilai ekonomi sebagai bahan makanan (sebagai pelengkap minuman penyegar ataupun sebagai bahan baku agar-agar). Alga merah sebagai bahan makanan memiliki kandungan serat lunak yang baik bagi kesehatan usus.
Flokkun er undir endurskoðun.
SamheitiRauðþörungar (fræðiheiti: Rhodophyta) eru þörungar sem finnast nær eingöngu í sjó og telur fylkingin yfir 5000 tegundir. Langflestir rauðþörungar lifa í fjörum eða grunnum í sjó og eru þar botnfastir á klettum, steinum eða gróðri. Rauðþörungar eru nánast allir fjölfrumungar og yfirleitt frekar stórir og nokkrir sm á lengd og breidd, oftast þráð eða blaðlaga. Algengar tegundir í fjörum eru fjörugrös (Chondrus crispus), söl (Palmaria palmata) og purpurahimna (Porphyra umbilicalis).
Fáir þeirra vaxa ofarlega í fjörum, en þegar neðar dregur ber meira á þeim. Þeir eru algengir á grunnsævi og ná lengst niður í djúpin af stórþörungunum. Rauðþörungar hafa löngum verið eftirsótt fæða um allan heim og þekkja Íslendingar þar af best sölin (Palmaria palmata), sem hafa á seinni tímum verið eini þörungurinn sem hafður eru til matar hér á landi.[2]
Rauðþörungar (fræðiheiti: Rhodophyta) eru þörungar sem finnast nær eingöngu í sjó og telur fylkingin yfir 5000 tegundir. Langflestir rauðþörungar lifa í fjörum eða grunnum í sjó og eru þar botnfastir á klettum, steinum eða gróðri. Rauðþörungar eru nánast allir fjölfrumungar og yfirleitt frekar stórir og nokkrir sm á lengd og breidd, oftast þráð eða blaðlaga. Algengar tegundir í fjörum eru fjörugrös (Chondrus crispus), söl (Palmaria palmata) og purpurahimna (Porphyra umbilicalis).
Fáir þeirra vaxa ofarlega í fjörum, en þegar neðar dregur ber meira á þeim. Þeir eru algengir á grunnsævi og ná lengst niður í djúpin af stórþörungunum. Rauðþörungar hafa löngum verið eftirsótt fæða um allan heim og þekkja Íslendingar þar af best sölin (Palmaria palmata), sem hafa á seinni tímum verið eini þörungurinn sem hafður eru til matar hér á landi.
Le alghe rosse o Rhodophyta (dall'antico greco ῥόδον, rhódon "rosa" e φυτόν, phytón "pianta", quindi "pianta rosa o rossa") sono organismi autotrofi eucarioti tra i più antichi,[1] privi di flagelli, fotosintetizzanti, che contengono clorofilla a e d (dovuta alla presenza di cianobatteri simbionti), ficobiline contenute nei ficobilisomi e comprendenti specie plurinucleate. Vivono soprattutto nelle acque di mare calde e se ne annoverano circa 4.000 specie,[2] (altri studi indicano 5-6000 specie[3] e qualcuno fino a 10.000[4]) gran parte delle quali pluricellulari. Sono organismi bio-costruttori, in quanto la loro crescita dà luogo a edifici calcarei di origine organica noti come piattaforme carbonatiche, e contribuiscono alla costruzione di vere e proprie scogliere organogene.
La parete cellulare è povera di cellulosa ma ricca di agar agar (polisaccaride acido) e carragenani; in alcune alghe rosse manca la cellulosa, al posto della quale ci sono xilani e mannani.
I cloroplasti di queste alghe hanno tilacoidi[4] che restano isolati, disposti più o meno parallelamente tra loro e non associati in lamelle. Un unico tilacoide periferico avvolge tutti quelli interni. Lungo le membrane tilacoidali si trovano allineati i ficobilisomi. I cloroplasti hanno forma stellata, reticolata o lobata. Le rodofite non hanno reticolo endoplasmatico. I cloroplasti hanno il pirenoide. La loro sostanza di riserva è l'amido delle floridee.
La maggior parte della alghe rosse ha un tallo molto elaborato, fatto dall'aggregazione di numerosi filamenti cellulari nei quali le singole cellule sono collegate mediante delle sinapsi chiamate Pit connections,[5] che sono delle perforazioni che mettono in comunicazione cellule contigue di uno stesso filamento e nelle quali si deposita materiale glicoproteico di forma lenticolare. Tra cellule di filamenti diversi si possono formare delle Pit connections secondarie, in questo caso, preliminarmente si forma una cellula laterale che poi viene riassorbita. Pit connections secondarie si possono formare anche tra cellule di organismi diversi, legati tra loro da un rapporto simbiontico.
Alcune specie di alghe rosse sono utilizzate dall'uomo come cibo; le specie Palmaria palmata[6] e la Porphyra[7] sono molto diffuse nelle tavole dell'Arcipelago britannico.[8]
Nella cucina asiatica le rodofite sono utilizzate sia come alimento (nori) o per produrre agar agar, carragenina o altri additivi alimentari. L'alto contenuto di vitamine (soprattutto la vitamina C), il buon contenuto proteico e la facilità di coltivazione, hanno favorito l'utilizzo alimentare di queste alghe che si è sviluppato in Giappone già a partire da oltre 300 anni fa.
Cyanidium sp., Cyanidiophyceae
Porphyra purpurea, Bangiophyceae
Bornetia secundiflora, Florideophyceae
Delesseria sanguinea, Florideophyceae
Laurencia sp., Florideophyceae
Ellisolandia elongata, Florideophyceae
Chondrus crispus, Florideophyceae
Sphaerococcus coronopifolius, Florideophyceae
Hildenbrandia rivularis, Florideophyceae
Peyssonelia squamaria, Florideophyceae
Plocamium corallorhiza, Florideophyceae
Stylonema alsidii, Stylonematophyceae
Le alghe rosse o Rhodophyta (dall'antico greco ῥόδον, rhódon "rosa" e φυτόν, phytón "pianta", quindi "pianta rosa o rossa") sono organismi autotrofi eucarioti tra i più antichi, privi di flagelli, fotosintetizzanti, che contengono clorofilla a e d (dovuta alla presenza di cianobatteri simbionti), ficobiline contenute nei ficobilisomi e comprendenti specie plurinucleate. Vivono soprattutto nelle acque di mare calde e se ne annoverano circa 4.000 specie, (altri studi indicano 5-6000 specie e qualcuno fino a 10.000) gran parte delle quali pluricellulari. Sono organismi bio-costruttori, in quanto la loro crescita dà luogo a edifici calcarei di origine organica noti come piattaforme carbonatiche, e contribuiscono alla costruzione di vere e proprie scogliere organogene.
Raudondumbliai (lot. Rhodophyta) – eukariotų (Eukaryota) domeno pirmuonių (Protista) karalystės skyriaus dumbliai. Priklauso laurencija. Raudondumbliai paplitę šiltose jūrose, gėluosiuose vandenyse. Auga dažniausiai 20-40 m, kartais net 100–200 m gylyje ant uolų ir akmenų prisitvirtinę prie substrato rizoidais.[1]
Raudondumbliai (lot. Rhodophyta) – eukariotų (Eukaryota) domeno pirmuonių (Protista) karalystės skyriaus dumbliai. Priklauso laurencija. Raudondumbliai paplitę šiltose jūrose, gėluosiuose vandenyse. Auga dažniausiai 20-40 m, kartais net 100–200 m gylyje ant uolų ir akmenų prisitvirtinę prie substrato rizoidais.
Rhodophyta são popularmente conhecidas como "algas vermelhas", por apresentar coloração característica. Com mais de 7000 espécies[1][2], a maior parte das algas vermelhas é marinha (clima tropical e temperado), e aproximadamente 200 espécies são de água doce (águas frias, rasas e de correntezas fortes). As rodófitas em geral são pluricelulares e crescem junto a um substrato, existem também as formas pseudoparenquimatosas ou foliáceas, e formas macroscópicas, microscópicas filamentosas. A camada interna dessa algas é constituída de microfibrilas de celulose e a matriz amorfa é composta por polissacarídeos e mucilagens (ágar e carragenana) importantes para as indústrias farmacêutica e alimentícia. Diversos produtos muito comuns contém esses polissacarídeos: meio de cultura para microrganismos, cápsulas, supositórios, anticoagulantes, filme fotográfico, sabonete, creme para mãos, "gelatina vegana", cremes, geléias, maioneses, pudins, merengues, bebidas, etc).
Alguns representantes de Rhodophyta são capazes de depositar carbonato de cálcio ou aragonitas (Nemaliales) em suas paredes celulares o que as tornam muito resistentes com baixa flexibilidade. Esta flexibilidade se dá por meio de genículos, regiões entre dois nós. Estas características são observadas nos gêneros Corallina e Jania. São muito abundantes, ecologicamente importantes e podem formar grandes recifes de corais.
Os fósseis de plantas mais antigos conhecidos na terra são algas vermelhas de 1,6 bilhão anos de idade.[3]
Características celulares
Os cloroplastos destas algas são revestidos por duas membranas que constituem o envelope do cloroplasto, com um tilacóide por banda e foram originados pelo processo de endossimbiose primária. Os pigmentos fotossintetizantes são a clorofila a, ficobilinas, carotenóides e xantofilas. Existem relatos da ocorrência de clorofila d, porém tais relatos são hoje tidos como errôneos.
A presença de pigmento vermelho permite a estas algas sobreviver a maiores profundidades que as demais, uma vez que lhes permite absorver o comprimento de onda que penetra mais fundo nos oceanos.
Essas algas possuem conexões citoplasmáticas conhecidas como sinapses, que estão entre duas células adjacentes durante a divisão celular ou entre duas células de dois filamentos vizinhos que se conectam. Cada conexão é constituída de uma região central protéica chamada de rolha sináptica, revestida por uma membrana contínua com a membrana citoplasmática de duas células adjacentes. Estas não representam conexões verdadeiras entre o citosol e a célula, para que haja troca de substância, mas funcionam como pontos de fortificação que conferem resistência estrutural ao talo.
Tipos de Reprodução
Reprodução assexuada
A reprodução assexuada no grupo ocorre pela produção de monósporos, que germinam formando, por mitose, um talo igual ao da geração que lhe deu origem. Os monósporos são formados por células chamadas de monosporângios (um esporo produzido por cada esporângio). A fragmentação vegetativa do talo também pode ser uma estratégia de reprodução assexuada.
Reprodução sexuada
As Rhodophyta não apresentam células flageladas. Na reprodução sexuada, as espermácias (gametas masculinos) produzidas pelos espermatângios (órgãos sexuais masculinos) são passivamente carregadas pela correnteza até o órgão sexual feminino, chamado de carpogônio. O carpogônio consiste de uma porção basal dilatada e de uma porção alongada e fina, a tricógine, que recebe os gametas masculinos. Como os gametas são aflagelados existe uma probabilidade baixa de que uma espermácia alcance uma tricógine receptiva. Alguns mecanismos aumentam esta probabilidade: 1) As espermácias de algumas espécies apresentam apêndices que aumentam a sua superfície e permitem que sejam carregados a maiores distâncias pela correnteza. 2) A composição de carboidratos constituintes de parede das espermácias difere das encontradas em células vegetativas, aderindo-se somente a parede das tricógines.
As algas vermelhas são classificadas como Archaeplastida, juntamente com as Glaucophyta e as Viridiplantae (algas verdes e plantas).
Abaixo estão duas classificações publicadas válidas (a de 2008 está por validar), apesar de nenhuma ter de ser necessariamente utilizada, uma vez que a taxonomia das algas ainda está num estado de permanente mudança (tendo a classificação acima do nível de ordem recebido pouca atenção ao longo da maior parte do século XX).
Se alguém define o reino Plantae como significado de Archaeplastida, as algas vermelhas farão parte desse reino; se o reino Plantae for considerado estritamente como Viridiplantae, então as algas vermelhas poderão ser consideradas como possuindo o seu próprio reino ou parte do reino Protista. As classificações válidas abaixo incluem as rodófitas no reino Plantae.
As algas foram colocadas sobre uma placa de petri.
Grateloupia
Chondracanthus
Jânia - Calcária
Porphyra
Ptilothamnion
Rhodophyta são popularmente conhecidas como "algas vermelhas", por apresentar coloração característica. Com mais de 7000 espécies, a maior parte das algas vermelhas é marinha (clima tropical e temperado), e aproximadamente 200 espécies são de água doce (águas frias, rasas e de correntezas fortes). As rodófitas em geral são pluricelulares e crescem junto a um substrato, existem também as formas pseudoparenquimatosas ou foliáceas, e formas macroscópicas, microscópicas filamentosas. A camada interna dessa algas é constituída de microfibrilas de celulose e a matriz amorfa é composta por polissacarídeos e mucilagens (ágar e carragenana) importantes para as indústrias farmacêutica e alimentícia. Diversos produtos muito comuns contém esses polissacarídeos: meio de cultura para microrganismos, cápsulas, supositórios, anticoagulantes, filme fotográfico, sabonete, creme para mãos, "gelatina vegana", cremes, geléias, maioneses, pudins, merengues, bebidas, etc).
Alguns representantes de Rhodophyta são capazes de depositar carbonato de cálcio ou aragonitas (Nemaliales) em suas paredes celulares o que as tornam muito resistentes com baixa flexibilidade. Esta flexibilidade se dá por meio de genículos, regiões entre dois nós. Estas características são observadas nos gêneros Corallina e Jania. São muito abundantes, ecologicamente importantes e podem formar grandes recifes de corais.
Os fósseis de plantas mais antigos conhecidos na terra são algas vermelhas de 1,6 bilhão anos de idade.
Características celulares
Os cloroplastos destas algas são revestidos por duas membranas que constituem o envelope do cloroplasto, com um tilacóide por banda e foram originados pelo processo de endossimbiose primária. Os pigmentos fotossintetizantes são a clorofila a, ficobilinas, carotenóides e xantofilas. Existem relatos da ocorrência de clorofila d, porém tais relatos são hoje tidos como errôneos.
A presença de pigmento vermelho permite a estas algas sobreviver a maiores profundidades que as demais, uma vez que lhes permite absorver o comprimento de onda que penetra mais fundo nos oceanos.
Essas algas possuem conexões citoplasmáticas conhecidas como sinapses, que estão entre duas células adjacentes durante a divisão celular ou entre duas células de dois filamentos vizinhos que se conectam. Cada conexão é constituída de uma região central protéica chamada de rolha sináptica, revestida por uma membrana contínua com a membrana citoplasmática de duas células adjacentes. Estas não representam conexões verdadeiras entre o citosol e a célula, para que haja troca de substância, mas funcionam como pontos de fortificação que conferem resistência estrutural ao talo.
Tipos de Reprodução
Reprodução assexuada
A reprodução assexuada no grupo ocorre pela produção de monósporos, que germinam formando, por mitose, um talo igual ao da geração que lhe deu origem. Os monósporos são formados por células chamadas de monosporângios (um esporo produzido por cada esporângio). A fragmentação vegetativa do talo também pode ser uma estratégia de reprodução assexuada.
Reprodução sexuada
As Rhodophyta não apresentam células flageladas. Na reprodução sexuada, as espermácias (gametas masculinos) produzidas pelos espermatângios (órgãos sexuais masculinos) são passivamente carregadas pela correnteza até o órgão sexual feminino, chamado de carpogônio. O carpogônio consiste de uma porção basal dilatada e de uma porção alongada e fina, a tricógine, que recebe os gametas masculinos. Como os gametas são aflagelados existe uma probabilidade baixa de que uma espermácia alcance uma tricógine receptiva. Alguns mecanismos aumentam esta probabilidade: 1) As espermácias de algumas espécies apresentam apêndices que aumentam a sua superfície e permitem que sejam carregados a maiores distâncias pela correnteza. 2) A composição de carboidratos constituintes de parede das espermácias difere das encontradas em células vegetativas, aderindo-se somente a parede das tricógines.
Červené riasy (lat. Rhodophyta) je taxonomická kategória (spravidla oddelenie) eukaryotických organizmov schopných fotosyntézy, tradične zaraďovaných medzi rastliny (a v rámci nich medzi riasy).
Vznikli endosymbiózou eukaryotickej bunky a sinice, ktorá sa stala predkom chloroplastov. Červené riasy sa oddelili od Zelených rias krátko po vzniku endosymbiózy a ďalší vývoj chloroplastov červených rias (tvz. "červená línia chloroplastov") bol nezávislý od predkov zelených rias a rastlín (tvz. "zelená línia chloroplastov"), preto sú červené riasy od nich dosť odlišné. V chloroplastoch obsahujú chlorofyl a a chlorofyl d. Kvôli odlišnostiam niektorí autori vyčlenili Červené riasy z ríše rastlín, ale posledné výskumy ich genetického kódu (DNA) poukazujú na existenciu ich spoločného predka.
Najstaršie skameneliny červených rias sú známe spred 750 – 1 250 miliónov rokov a patria v najstarším mnohobunkovým organizmom. Ide o vláknité riasy Bangiomorpha pubescens, podobné súčasným druhom rodu Bangia. Súčasné vývojové línie červených rias sa vytvorili približne pred 550 mil. rokov. Mnohé druhy tvorili dôležitú súčasť koralových útesov od neskorého paleozoika (napr. Soleopra, známa z Kambria až jury). Počas jury sa zvyšovala ich diverzita a ostali významnou súčasťou koralových útesov aj V súčasnosti.
Niektoré červené riasy sú jednobunkové, ale väčšina z nich má vláknitú, alebo pletivovitú stielku. Bunková stena červených rias je tvorená amorfnou matrix z polysacharidov, hlavne z galaktanu, agaru a karagénu. V amorfnej matrix sa nachádzajú celulózové alebo xylánové mikrotubuly. Bunková stena je často zvápenatelá - vyplnená kalcitovými, alebo aragonitovými kryštálmi. Bunky červených rias nikdy nemajú bičík.
Asi 90 % červených rias žije v mori, hlavne v tropických vodách. Niektoré červené riasy ukladajú do bunkovej steny anorganický uhličitan a tvoria súčasť koralových útesov.
Chloroplasty červených rias sú ohraničené dvojitou membránou, vznikli totiž primárnou endosymbiózou zo sinice. Na rozdiel od rastlín ale tylakoidy netvoria graná. Fotosyntetickým pigmentom je chlorofyl a a d a fykobilíny: c-fykocyanín, allofykocyín, r-fykocyanín a r-fykoerythrín. C-fykocyanín a allofykocyanín sa vyskytujú aj u siníc, r-fykocyanín a r-fykoerythrín sú špecifické pre červené riasy.
Červené pigmenty červených rias sú schopné dobre zachytiť modrozelené svetlo, ktoré preniká v oceánoch do najväčšej hĺbky. Môžu preto žiť vo väčších hĺbkach ako zelené riasy (180 až 210 m).
Zvláštnosťou červených rias je aj zásobná látka – florideový škrob, ktorý sa vo forme zŕn ukladá v cytoplazme. Červené riasy nemajú centrioly a nie sú u nich známe žiadne bičíkaté štádiá.
V rozmnožovacom cykle sa strieda pohlavná a nepohlavná generácia, často je cyklus trojfázový, s dvomi diploidnými a jedným haploidným štádiom. Rodozmena môže byť izomorfická (napr. v rode Polysiphonia sp.) aj heteromorfická (napr. v rode Porphyra sp.).
Nepohlavný a diploidný tetrasporofyt vytvára diploidné tetraspóry, ktoré vytvárajú samčí, alebo samičí gametofyt. Gametofyt vytvára haploidné pohlavné bunky (gaméty). Gaméty vytvárajú karposporofyt v ktorom dôjde k ich splynutiu. Karposporofyt sa ďalej rozmnožuje nepohlavne diploidnými karpospórami. Z každej karpospóry vyrastie tetrasporofyt a cyklus sa opakuje.
Červené riasy sú veľmi početnou skupinou zahrňujúcou asi (4 000 – 6 000 druhov). Patria k dominantným morským makrofytom. Rastú v litorálnej aj sublitorálnej zóne, vo väčších hĺbkach ako iné riasy. Sú to hlavne tropické druhy, ale vyskytujú sa aj v chladnejších vodách, vrátane Antarktídy. Sladkovodné červené riasy preferujú prudko tečúce vody. napr. Batrachospermum sp.
Druh Porfyra sa v Ázii pestuje na morských plantážach, a konzumuje pod názvom „hoši-nori“ alebo nori. Tiež sa používa ako krmivo alebo hnojivo. Konzumuje sa aj druh Palmaria palmata.
Extrakciou v horúcej vode sa z červených rias získava agar (hlavne Gelidium sp.) a karagén (Chondrus sp.), ktoré sa využívajú v potravinárstve alebo v mikrobiológii. Sekundárne metabolity niektorých druhov majú antibiotické účinky.
Oddelenie Rhodophyta - červené riasy
Červené riasy (lat. Rhodophyta) je taxonomická kategória (spravidla oddelenie) eukaryotických organizmov schopných fotosyntézy, tradične zaraďovaných medzi rastliny (a v rámci nich medzi riasy).
More may exist?
홍조식물(紅藻植物; 문화어: 붉은마름식물) 또는 홍조류(紅藻類)는 조류로 분류되는 원생생물의 한 갈래로, 전 세계에 약 600속의 2,000종 정도가 알려져 있다. 모두 분홍색이나 암홍색 색소체를 가지며, 광합성 작용에 의해 홍조 녹말을 만든다. 대부분은 바다 속에서 자라지만, 이 밖에 민물이나 축축한 흙 표면에서 자라는 것도 있다. 한편, 몇 속의 단세포인 것을 제외하면 거의가 다세포로 이루어져 있으며, 크기는 수 mm에서 수십 mm에 이른다. 몸은 실·잎·나뭇가지 모양 또는 그물코 모양으로 된 것도 있다. 대부분 부드러운 것이 많으나, 산호말류처럼 세포벽 사이에 탄산칼슘이 축적되어 돌처럼 딱딱하게 된 무리도 있다.
홍조식물의 공통적인 특징은 다음과 같다.
김이나 풀가사리 무리처럼 얕은 바다에서 생육하는 것도 있지만, 대부분의 홍조식물은 간조선보다 더 깊은 곳에서 자란다. 특히, 우뭇가사리는 약 10m 정도의 깊은 곳에서 많이 자라며, 이 밖에 100m 이상의 깊은 곳에서 자라는 종류도 매우 많다. 홍조식물이 이렇게 깊은 곳에서 자랄 수 있는 것은 체내에 있는 홍조소가 약 0.5mμ 부근의 단파장 빛을 광합성에 이용하기 때문이다. 빛은 바다 수심이 깊어짐에 따라 적색·주황색·황색·자색·남색·녹색·청색의 순서로 흡수되므로, 가장 깊은 곳까지 도달될 수 있는 것은 청색과 녹색이다. 따라서, 홍조식물은 깊은 바다에 위치하면서 자신의 몸 빛깔과 보색 관계에 있는 청색이나 녹색빛을 광합성에 이용한다. 한편, 얕은 곳에서 자라는 진두말 등은 전체적으로 녹색을 띠므로, 홍조식물이 아니라고 생각되기도 하는데, 이것은 적색이나 황색의 장파장 빛을 광합성에 잘 이용한 결과 녹색 엽록소량은 늘어난 반면 홍조소는 감소된 보색 적응 현상 때문이다.
다음은 원시색소체생물의 계통 분류이다.[1][2][3][4][5][6][7][8][9][10]
원시색소체생물 녹색식물 스트렙토식물아래는 최근 제안된 홍조류의 계통 분류 중 하나이다.[11][12]
홍조식물문 키아니디움아문 원시홍조식물아문 진정홍조식물아문
.jpg)



.jpg)
















.jpg)




















.jpg)




























