nimet breadcrumb-navigoinnissa
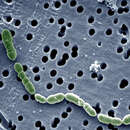
Bacteri làctic que fins al 1995 es considerava Leuconostoc oeni, ja que són cocs i realitzen la fermentació heterolàctica (quan fermenten els sucres a banda d'àcid làctic també formen CO2, acètic i etanol). Tot i així, la comparació de les seqüències de l’RNA ribosomal, demostra que filogenèticament Oenococcus està ben separat de Leuconostoc, així com d’altres bacteris làctics relacionats com Lactobacillus.
El seu hàbitat és el most i el vi, i pot créixer en pH d'entre 3-4. En enologia és molt apreciat perquè és el bacteri làctic principal que duu a terme la fermentació malolàctica.[1] Aguanta la presència d'etanol fins a concentracions de 10% (v/v).
Bacteri làctic que fins al 1995 es considerava Leuconostoc oeni, ja que són cocs i realitzen la fermentació heterolàctica (quan fermenten els sucres a banda d'àcid làctic també formen CO2, acètic i etanol). Tot i així, la comparació de les seqüències de l’RNA ribosomal, demostra que filogenèticament Oenococcus està ben separat de Leuconostoc, així com d’altres bacteris làctics relacionats com Lactobacillus.
El seu hàbitat és el most i el vi, i pot créixer en pH d'entre 3-4. En enologia és molt apreciat perquè és el bacteri làctic principal que duu a terme la fermentació malolàctica. Aguanta la presència d'etanol fins a concentracions de 10% (v/v).
Oenococcus oeni ist ein grampositives Bakterium aus der Ordnung der Milchsäurebakterien (Lactobacillaceae). Dieses Bakterium wurde bis 1995 zur Gattung Leuconostoc gezählt, gilt heute aber neben Oenococcus kitaharae und Oenococcus alcoholitolerans als einziges Mitglied der Gattung Oenococcus. Kennzeichnend für diese Bakterien ist die Milchsäuregärung, ein fermentativer Stoffwechsel, bei dem Milchsäure als Endprodukt auftritt. Oenococcus oeni ist, wie alle Gattungen der Familie Leuconostocaceae, obligat heterofermentativ. Neben Milchsäure treten hierbei auch andere Stoffwechselendprodukte, wie z. B. CO2, Acetat (Essigsäure) und Ethanol (Alkohol) auf, während bei der homofermentativen Milchsäuregärung ausschließlich Milchsäure entsteht. Vom Menschen wird Oenococcus oeni bei der Weinherstellung genutzt.
Oenococcus oeni ist acidophil, zeigt also gutes Wachstum bei niedrigen pH-Werten. Die Zellen sind kokkenförmig. Oenococcus oeni toleriert große Mengen von Alkohol (Ethanol) und kann in Medien mit 10 % Ethanolanteil kultiviert werden.
Oenococcus oeni kommt natürlich in Frucht-Maische vor. Dieser Mikroorganismus wird in der Weinproduktion eingesetzt, da er die Fähigkeit besitzt, durch Milchsäuregärung Äpfelsäure in Milchsäure umzuwandeln (Malolaktische Gärung). Dadurch wird der Säuregehalt des Weines verringert, der somit einen milderen Geschmack bekommt.
Oenococcus oeni kann aus Citrat Diacetyl bilden, eine Verbindung mit typischem Buttergeschmack. In geringen Konzentrationen trägt es zum Aroma bei, in größeren Mengen jedoch verdirbt es den Wein.
Die Gattung Oenococcus bildet zusammen mit Leuconostoc und Weissella die Familie Leuconostocaceae. Diese Gattungen werden innerhalb der Phylogenie zusammen mit weiteren Gattungen wie Lactobacillus, Lactococcus, Enterococcus und Carnobacterium (alle typische Milchsäurebakterien) zu den sogenannten "Clostridium-Zweig" (englisch: Clostridium branch) gestellt, sie sind also evolutionär eng miteinander verwandt. Neben Oenococcus oeni wird seit 2006 die Art Oenococcus kitaharae sowie seit 2015 die Art Oenococcus alcoholitolerans zu der Gattung Oenococcus gestellt.[2] Letztere wurde erstmals 2014 aus Fermentationsbottichen für Cachaça in Brasilien isoliert und ein Jahr später offiziell als neue Spezies innerhalb der Gattung anerkannt[3][4].
Oenococcus oeni steht aufgrund von 16s-rRNA-Untersuchungen im Vergleich zu Arten von Leuconostoc und Weisella weiter entfernt auf dem phylogenetischen Ast und scheint in bei Weissella und Leuconostoc eher konservierten Punkten der rRNA stärker zu variieren. Oenococus oeni scheint somit eine besonders schnelle Evolutionsrate aufzuweisen.[5] Hierbei spricht man von einer tachytelischen Evolutionsrate, der Gegensatz ist die bradytelische Rate, also extrem langsam verlaufende Veränderungen in der Evolution. Allerdings wurde die Annahme einer tachytellischen Evolutionsrate bei Oenococcus oeni wieder in Frage gestellt[6]
Oenococcus oeni ist ein grampositives Bakterium aus der Ordnung der Milchsäurebakterien (Lactobacillaceae). Dieses Bakterium wurde bis 1995 zur Gattung Leuconostoc gezählt, gilt heute aber neben Oenococcus kitaharae und Oenococcus alcoholitolerans als einziges Mitglied der Gattung Oenococcus. Kennzeichnend für diese Bakterien ist die Milchsäuregärung, ein fermentativer Stoffwechsel, bei dem Milchsäure als Endprodukt auftritt. Oenococcus oeni ist, wie alle Gattungen der Familie Leuconostocaceae, obligat heterofermentativ. Neben Milchsäure treten hierbei auch andere Stoffwechselendprodukte, wie z. B. CO2, Acetat (Essigsäure) und Ethanol (Alkohol) auf, während bei der homofermentativen Milchsäuregärung ausschließlich Milchsäure entsteht. Vom Menschen wird Oenococcus oeni bei der Weinherstellung genutzt.
Oenococcus oeni (known as Leuconostoc oeni until 1995) is a Gram-positive bacterial species in the genus of Oenococcus.[2] It was the only species in the genus until 2006, when the species Oenococcus kitaharae was identified. As its name implies, Oe. oeni holds major importance in the field of oenology, where it is the primary bacterium involved in completing the malolactic fermentation.[3]
Diacetyl in wine is produced by lactic acid bacteria, mainly Oe. oeni. In low levels, it can impart positive nutty or caramel characters, but at levels above 5 mg/L, it creates an intense buttery or butterscotch flavour, where it is perceived as a flaw. The sensory threshold for the compound can vary depending on the levels of certain wine components, such as sulfur dioxide. It can be produced as a metabolite of citric acid when all of the malic acid has been consumed. Diacetyl rarely taints wine to levels where it becomes undrinkable.[4]
Oenococcus oeni (known as Leuconostoc oeni until 1995) is a Gram-positive bacterial species in the genus of Oenococcus. It was the only species in the genus until 2006, when the species Oenococcus kitaharae was identified. As its name implies, Oe. oeni holds major importance in the field of oenology, where it is the primary bacterium involved in completing the malolactic fermentation.
Oenococcus oeni (anciennement Leuconostoc oenos remplacé en 1995) est une bactérie lactique du genre Oenococcus.
Cette espèce bactérienne est responsable de la fermentation malolactique sur les vins, qui intervient normalement après la fermentation alcoolique des levures.
Le métabolisme des sucres est hétérofermentaire[1], et transforme :
A l'aide de l’enzyme malolactique, la réaction se produit tel que : COOH-CH2-CHOH-COOH → CH3-CHOH-COOH + CO2
À partir de la dégradation du citrate, productions possible d'acétate, de lactate, d'éthanol, de lipides ou d'acides gras, et de substances acétoïniques comme le diacétyle, l’acétoïne, et le 2,3-butanediol. Ces dernières substances sont des composés aromatiques caractéristiques de notes de beurre/lacté, pouvant être recherchées dans les vins blancs.
Production non désirée d'histamine et de tyramine qui sont des amines biogènes.
La réaction, grâce à l’enzyme agrinine déiminase, produit en faible quantité un composé cancérigène :
Oenococcus oeni (anciennement Leuconostoc oenos remplacé en 1995) est une bactérie lactique du genre Oenococcus.
Oenococcus oeni (Garvie, 1967) Dicks et al. 1995 è un batterio Gram-positivo appartenente alla famiglia delle Leuconostocaceae; rappresenta l'unica specie nota del genere Oenococcus Dicks et al. 1995.
È responsabile della fermentazione malolattica del vino.
Bacilli disposti a coppie o catenelle, gram positivi, immobili, asporigeni, anaerobi aerotolleranti, eterofermentanti (glucosio -> acido D-lattico 50%min + acido acetico + alcol etilico + anidride carbonica).
L'importanza di questa specie di batteri lattici è dovuta al fatto che è quella responsabile della fermentazione malolattica del vino. La malolattica non è una vera fermentazione, bensì una degradazione dell'acido malico in acido L-lattico.
Questi microrganismi sono capaci di svilupparsi a pH relativamente bassi (pH del vino 3,5-3,7), dannosi invece per gli altri lattobacilli. Perché la fermentazione malolattica avvenga però, sono necessarie alcune condizioni chimico-fisiche fondamentali: temperatura minima di 18-20 °C, concentrazione di anidride solforosa molecolare inferiore a 0,6 mg/l, concentrazione di etanolo inferiore al 10-13% v/v e assenza di residui zuccherini nel vino (che porterebbero ad una fermentazione vera e propria).
Molte cantine preferiscono ormai inoculare il vino con ceppi selezionati di batteri malolattici che facilitano la partenza della fermentazione malolattica e garantiscono l'assenza di composti indesiderati come metaboliti secondari (per esempio le ammine biogene, tossiche).
Questo processo può portare importanti vantaggi sui vini sia bianchi che rossi prodotti a latitudini molto alte (Reims, Bordeaux, Alsazia) che hanno un contenuto in acido malico elevato e svantaggi soprattutto per i vini bianchi o i vini base spumante aromatici (Prosecco Moscato ecc.) che maturano a latitudini più basse. Nelle regioni del sud difficilmente si fa questo tipo di processo per la quasi totale assenza dell'acido malico che con le alte temperature viene degradato.
Oenococcus oeni (Garvie, 1967) Dicks et al. 1995 è un batterio Gram-positivo appartenente alla famiglia delle Leuconostocaceae; rappresenta l'unica specie nota del genere Oenococcus Dicks et al. 1995.
È responsabile della fermentazione malolattica del vino.
Oenococcus oeni є грам-позитивною коковою бактерією, що формує ланцюжки. Енергію отримує в результаті гетероферментативного молочнокислого бродіння, продуктами якого є молочна кислота, оцтова кислота і діоксид вуглецю. Здатна зброджувати різні вуглеводи (сахарозу, фруктозу, глюкозу)[3], піруват[4] а також яблучну і лимонну кислоту. Утворює протеази, що залишаються активними в присутності сірчаного газу і етанолу[5]. O. oeni здатний виживати при концентрації етанолу до 10 % у живильному середовищі.
У 1998 році було проведено генетичне картування геному O. oeni штаму PSU-1 і побудована фізична карта хромосоми розміром 1,857 Mbp[6], а в 2000 році був картований геном штаму GM[7]. У 2006 році було закінчено визначення повної нуклеотидної послідовності ДНК геному O. oeni штаму PSU-1. Геном представлений кільцевою двуцепочечной молекулою ДНК розміром 1780517 пар основ. і містить 1864 гени, з яких 1691 кодують білки, вміст ГЦ становить 37 %. Також в геномі міститься кілька плазмід. Плазміда pOM1 представлена кільцевою дволанцюжковою молекулою ДНК розміром 3926 пар основ і містить три гени, всі з яких кодують бфлки[8]. Плазміди pRS2 і pRS3 також несуть по три гени, що кодують білки, і є кільцевими молекулами ДНК розміром 2544 і 3948 пар основ відповідно[9][10].
O. oeni грає важливу роль в процесі дозрівання вина після закінчення спиртового бродіння[11]. O. oeni відповідальнє за процеси яблочнокисло-молочнокислого бродіння[12]. В процесі життєдіяльності O. oeni утилізує яблучну і молочну кислоти, тим самим знижуючи кислотність вина, а також виробляє діацетил, що має приємний аромат, тим самим покращуючи аромат вина[13]. Важливою особливістю O. oeni є відносна стійкість до таких стресових факторів, як іони меди, часто присутні у вині[14].
Oenococcus oeni
(Garvie 1967) Dicks et al. 1995
Oenococcus oeni (лат.) — грамположительная гетероферментативная молочнокислая кокковидная бактерия рода Oenococcus. Принимает участие в созревании вин, сбраживая яблочную и лимонную кислоту и образует диацетил и другие летучие ароматические вещества, понижая тем самым кислотность вина и придавая вину приятный аромат[1]. Раньше входила в род Leuconostoc под названием Leuconostoc oenos, в 1995 году было предложено перенести Leuconostoc oenos во вновь созданный род Oenococcus Dicks et al. 1995[2].
Oenococcus oeni представляет собой грамположительную кокковидную бактерию, образующую цепочки. Энергию получает в результате гетероферментативного молочнокислого брожения, продуктами которого являются молочная кислота, уксусная кислота и диоксид углерода. Способна сбраживать различные углеводы (сахарозу, фруктозу, глюкозу)[3], пируват[4] а также яблочную и лимонную кислоту. Образует протеазы, остающиеся активными в присутствии сернистого газа и этанола[5]. O. oeni способен выживать при концентрации этанола до 10 % в питательной среде.
В 1998 году было проведено генетическое картирование генома O. oeni штамма PSU-1 и построена физическая карта хромосомы размером 1,857 Мп.н.[6]. В 2006 г. было закончено определение полной нуклеотидной последовательности ДНК генома O. oeni штамма PSU-1. Геном представлен кольцевой двуцепочечной молекулой ДНК размером 1780517 п.н. и содержит 1864 гена, из которых 1691 кодируют белки, процент % Г+Ц пар составляет 37 %. Также в геноме содержится несколько плазмид. Плазмида pOM1 представлена кольцевой двуцепочечной молекулой ДНК размером 3926 п.н. и содержит три гена, из которых три гена кодируют белки[7]. Плазмиды pRS2 и pRS3 также несут по три гена, кодирующих белки и являются кольцевыми молекулами ДНК размером 2544 и 3948 п.н. соответственно[8][9].
O. oeni принимает важное значение в процессе дозревания вина после окончания спиртового брожения[10]. O. oeni ответственен за процессы яблочнокислого-молочнокислого брожения[11]. В процессе жизнедеятельности O. oeni утилизирует яблочную и молочную кислоты, тем самым понижая кислотность вина, а также производит диацетил, имеющий приятный аромат, тем самым улучшая аромат вина[12]. Важной особенностью O. oeni является относительная устойчивость к таким стрессовым факторам, как диоксид серы и ионы меди, часто присутствующие в вине[13].
Oenococcus oeni (лат.) — грамположительная гетероферментативная молочнокислая кокковидная бактерия рода Oenococcus. Принимает участие в созревании вин, сбраживая яблочную и лимонную кислоту и образует диацетил и другие летучие ароматические вещества, понижая тем самым кислотность вина и придавая вину приятный аромат. Раньше входила в род Leuconostoc под названием Leuconostoc oenos, в 1995 году было предложено перенести Leuconostoc oenos во вновь созданный род Oenococcus Dicks et al. 1995.


