Comprehensive Description
provided by Smithsonian Contributions to Zoology
Dendrocoelum lychnidicum (Stankovi)
Neodendrocoelum sp. 1 Stankovi, 1955a, pl. 5: fig. 5.
Neodendrocoelum lychnidicum Stankovi, 1969:148.
Dendrocoelum (Neodendrocoelum) lychnidicum.—Gourbault, 1972:66.
Dendrocoelum lychnidicum.—Kenk, 1974:18.
MATERIAL DEPOSITED.—Sagittal serial sections of 3 specimens on 4 slides, USNM 55269–55271.
Dendrocoelum lychnidicum was first mentioned and illustrated by Stankovi (1955a:491) as an undescribed “spec. 1” of Neodendrocoelum, inhabiting the sublittoral zone of Lake Ohrid. A description of the external aspect and the anatomy of the species was given by the same author in 1969 (pages 418–419). Based on my material, the following supplements the original description and indicates some of the variability of the species.
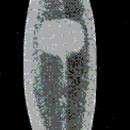
EXTERNAL FEATURES (Figure 8).—This rather rare form is the smallest of the pigmented Dendrocoelum species of Lake Ohrid. Mature animals attain a length of 6 mm and a width of 1 mm. The anterior end is truncated, with a slightly bulging frontal margin and moderately rounded lateral corners. There is no distinct narrowing or neck behind the head. The body has a more or less ovoid outline, the greatest width being at approximately the middle of the body. The posterior end is bluntly pointed.
The distance of the two eyes from each other is about one-third, or less, of the transverse diameter of the head, and their distance from the frontal margin approximately equal to that from the lateral margins.
The central area of the dorsal surface has a rather light chocolate brown ground color of somewhat cloudy appearance. Toward the head and the margins of the body, the brown pigment fades out, leaving a broad marginal rim free of pigment. There is an indistinct light band along the midline. Very characteristic is one large rectangular or more rounded spot of light yellowish color placed transversely in the brown area, at about the middle of the back in young animals, more anterior in adult specimens. Occasionally two small round spots in the midline, one behind the other, are present in mature specimens above the copulatory organs. The ventral surface has a light yellowish-brown hue.
The animal moves principally by gliding.
ANATOMY.—A small adhesive area is situated on the underside of the frontal margin, sometimes appearing as a shallow central depression. Many eosinophilic glands open in this area and a few muscle fibers, running mainly in a longitudinal direction, are attached to it. The infranucleate epithelium and the glands show the same structure as those of the marginal adhesive zone from which they are separated on each side by a gap. The pharynx is short and wide, measuring about one-seventh the length of the body (in preserved animals); the ratio between its width and length is about 1:1.7, but may change to 1:1 when the body is contracted.
In the three mature specimens at my disposal for the study of the reproductive system, the copulatory apparatus was well differentiated. One of the specimens laid a cocoon shortly before it was preserved. Apparently the animals had already passed the peak of their reproductive function, as no testes could be found in the three specimens, though sperm were present in the spermiductal vesicles and also in the tubae of the oviducts (indicating that the animals had copulated). The complete reduction of the testes may be a sign of a pronounced protandry such as has been observed also in some other freshwater triclads. Stankovi (1969:418) indicates that in his specimens the numerous small testes were located predominantly on the ventral side. The ovaries were situated far posteriorly, about midway between the head end and the root of the pharynx.
The thickened glandular area surrounding the genital aperture (Figure 43, gp) is rather small. In the general arrangement of the copulatory apparatus, Dendrocoelum lychnidicum agrees with its pigmented relatives from the lake. The penis is small and has a remarkably weak musculature. When the organ is extended (Figure 43), the lumen of the penis bulb (bp) is a rather small cavity (seminal vesicle), receiving the two vasa deferentia (vd) from the right and left sides. From this cavity, a narrow, almost straight canal extends through the papilla (pp) to open at its tip. The penis papilla is constricted at its base, where a subepithelial layer of circular muscle fibers is developed. In the other two specimens, however, the penis was retracted and the papilla partly inverted into the widened seminal vesicle (Figure 57). The inversion or invagination occurred at the basis of the papilla and not, as is usually the case in the genus Dendrocoelum, at the free end of the papilla. The tissues of both the bulb and papilla of the penis are traversed by many gland ducts, at least part of them opening through the epithelial covering of the papilla.
The male atrium is lined with a tall columnar epithelium. The distal parts of the cells, bordering the lumen of the atrium, are closely packed with granular, strongly eosinophilic inclusions. The same differentiation is observed also in related species, but here it is more conspicuous due to the great thickness of the epithelium, which reduces the atrial lumen to a narrow canal with irregular outline.
The oviducts run, in the typical way, as a pair of narrow tubes (outer diameter, 12 to 14 μm) above the ventral nerve cords to a level posterior to the copulatory complex. There they converge medially, increasing in thickness (diameter, 20 to 35 μm) and unite in the midline. The long common oviduct (odc) extends anteriorly in a slight curve and opens into the posterior part of the male atrium. The diameter of this duct is very large (43 to 50 μm) compared to the dimensions of the atrial organs. The end parts of the paired oviducts and the common oviduct are equipped with eosinophilic shell glands. The copulatory bursa (b) is of moderate size. Its outlet, the bursal duct (bd), passes on the left side of the penis, starting as a narrow canal but expanding considerably in its posterior half; above the genital pore, it curves ventrally and connects, from the dorsal side, with the common atrium (ac). The adenodactyl (ad), situated ventrally and to the left of the midline, is long and slender, exceeding the pharynx in length, with an elongated lumen extending through the greater part of the organ.
DISTRIBUTION AND ECOLOGY.—About 10 specimens of Dendrocoelum lychnidicum were collected in August 1937 in Ohrid Bay by dredging in the shell zone of the sublittoral at depths between 16 and 26 m. The animals are comparatively rare and can easily be overlooked because of their small size and slow movements. An attempt to rear immature individuals to maturity in the laboratory was unsuccessful. They kept sufficiently well at a temperature of 10°C, but refused to take beef liver offered as food and died after a few months. One of the mature specimens deposited a cocoon in the laboratory culture; it was spherical, reddish-brown, unstaked, but adhered to the substrate by a mucous secretion.
TAXONOMIC POSITION.—Externally, Dendrocoelum differs from related species of the Ohrid region mainly by its small size and by the characteristic saddle-shaped light spot of the dorsal side. Anatomically, it conforms well with its pigmented relatives. Its chromosome numbers are 2n = 32 (Paunovi, 1977).
Dendrocoelum ochridense (Stankovi and Komárek)
Neodendrocoelum ochridense Stankovi and Komárek, 1927: 605.
Neodendrocoelum ohridense.—Stankovi, 1932:578.
Dendrocoelum (Neodendrocoelum?) ochridense.—de Beauchamp, 1931:157.
Dendrocoelum (Neodendrocoelum) ohridense.—Gourbault, 1972:68.
Dendrocoelum ochridense.—Kenk, 1974:18.
MATERIAL DEPOSITED.—Sagittal and horizontal serial sections of 5 specimens on 25 slides, USNM 55280–55284.
This well-defined species of the lower littoral, sublittoral, and profundal zones of Lake Ohrid was first described by Stankovi and Komárek (1927:605–609) from a single specimen dredged from a depth of 40 m. Unfortunately, the description of the external characters and the accompanying illusstration (fig. 5) were inaccurate. The original description was corrected and supplemented in a later paper by Stankovi (1938:3–5). The anatomical data given in both papers are sufficient to identify the species with certainty.
EXTERNAL FEATURES (Figure 4).—Mature specimens attain a length of 16–20 mm and a width of 3–4 mm. Compared with related species, D. ochridense is a rather slender form. The head is only a little narrower than the body and is separated from the latter by a slight, hardly noticeable constriction or neck. In the quietly gliding animal, the frontal margin of the head has a small, bluntly pointed, anterior projection in the midline. The lateral edges of the head are likewise more pointed than those of other Dendrocoelum species of the same region. Behind the neck, the lateral body margins first diverge gradually, then run parallel, and, beginning at the level of the copulatory organs, converge to form a distinctly pointed posterior end. The body is considerably flattened dorsoventrally.
The two eyes are situated close to the midline, their distance from each other amounting to about one-fifth the transverse diameter of the head at the level of the eyes; their distance from the frontal margin is two to three times as large as their mutual distance. The root of the pharynx lies at about the middle of the body or somewhat anterior to it in mature animals. The mouth and the genital opening divide the posterior half of the body into approximately equal thirds.
The color pattern of the dorsal surface is very characteristic, composed of two colors, light yellowish and dark brown. The yellowish background color forms a distinct median band extending from the anterior margin of the head to the posterior end of the body. Lateral to this band on either side is a longitudinal row of six to ten large dark spots of triangular or trapezoidal shape. The most anterior spots are situated a short distance behind the eyes. The arrangement of the spots is rather regular. In some individuals the pattern may be almost symmetrical on both sides, particularly in the anterior part of the body. More frequently, however, the spots of the two sides are placed at different levels. The spots are connected with each other by a thin line bordering the light median band. A similar line may join the lateral margins of the spots; usually, however, the spots fade out laterally midway between the median line and the lateral margin of the body. There is always a rather broad, light border along the margins, usually paler than the yellow ground color of the middorsal surface. A pair of indistinct unpigmented strips is visible on the head at the site of the auricular sense organs.
The ventral surface is whitish or yellowish. When the intestine is not filled excessively, the color pattern of the dorsal surface may shine through ventrally.
The locomotion of the animal is normally gliding, occasionally “crawling.” Upon strong stimuli, the animal detaches from the substrate and swims freely in the water for a short distance. This particular reaction will be discussed below in connection with the ecology of the species.
ANATOMY.—The anterior adhesive organ of Dendrocoelum ochridense is feebly developed, consisting of a small field of epidermis in which numerous adhesive gland ducts open, measuring about 100 μm in median diameter (Figure 30). The extraordinary thick subepidermal muscle layers (Figure 27) of the species have already been mentioned by Stankovi and Komárek (1927:608) and by Stankovi (1969:429).
The numerous testes (Figure 27, te) are arranged in two broad longitudinal zones, one on either side of the midline, extending from the region of the ovaries to close to the posterior end. They are comparatively small and are situated in the mesenchyme ventrally to the intestine and between the intestinal branches, leaving the dorsal parts of the mesenchyme free. The dorsal and lateral regions of the mesenchyme are occupied chiefly by yolk glands. Tubal bursae at the beginning of each oviduct are well developed.
The general anatomy of the copulatory apparatus (Figure 44) resembles closely that of Dendrocoelum maculatum. The penis lies somewhat to the right of the midline, the adenodactyl and the posterior portion of the bursal duct to the left of the penis. The genital aperture (gp) is surrounded by a small circular area of modified surface epithelium, as in D. maculatum. The spaceous common atrium (ac), enclosing the large papilla of the adenodactyl, has a very characteristic shape; it forms a long, conical extension posterior to the gonopore, which may reach about halfway between the genital pore and the hind end of the body in fully mature individuals.
The penis is smaller than the adenodactyl and consists of a spherical, moderately muscular bulb and a short, conical or gland-shaped papilla. The bulb has a wide central cavity, the seminal vesicle (vs), lined with a glandular epithelium, the cells of which protrude irregularly into the lumen. The two vasa deferentia (vd) open separately into the anterolateral parts of the vesicle. A wide canal, lined with a flattened epithelium, proceeds to the tip of the papilla, where it opens through a somewhat narrowed aperture. The wall of the penis papilla is thin and contains a layer of circular muscles beneath the outer epithelium, rather thick in the basal part of the papilla, but tapering off distally. Gland ducts, containing a granular secretion, open through the surface epithelium of the papilla.
The long common oviduct (odc) connects with the end part of the male atrium. The duct (bd) of the copulatory bursa opens into the common atrium from the dorsal side. The adenodactyl (ad) has a voluminous bulb and a long, slender, straight papilla. Its glandular lumen is either confined to the papilla or extends through both the bulb and the papilla, particularly in fully mature animals. It may be noted that the papilla of the adenodactyl lies in the posterior extension of the common atrium in all four individuals examined by me and in the two specimens that Stankovi and Komárek (1927:606) and Stankovi (1938:4) used for their illustrations. In other species, the papilla is frequently curved ventrally and tends to protrude through the gonopore. The action of the killing fluid apparently causes a contraction of the musculature of the adenodactyl, accompanied by an elongation of the papilla. This appears to occur also in Dendrocoelum ochridense; however, as the papilla is not bent downward, it pushes the wall of the common atrium posteriorly. It is possible that the shape of the common atrium in the living animal is not as elongated as it appears to be in the slides. The posterior extension of the atrium may well be caused, or at least greatly exaggerated, by the expanding adenodactyl.
DISTRIBUTION AND ECOLOGY.—Stankovi and Komárek (1927:606) and Stankovi (1938:5; 1955a: 491) collected Dendrocoelum ochridense in the lower littoral and the sublittoral zones of Lake Ohrid; they found single specimens even at greater depths, down to 120 m. Individuals taken in the Chara zone (5–20 m) were always immature and sexual specimens were found only in the shell zone (25–35 m). Stankovi assumes that the animals migrate from the littoral zone to the sublittoral when they approach sexual maturity.
I collected the species during the summer months in Ohrid Bay on Chara (10–15 m depth) and in the shell zone (15–36 m) by dredging and using meat bait; off Kale, below the shell zone (at about 45 m); and near Kalište, at the lower boundary of the Chara zone (7–13 m). All specimens were immature except for one sexual individual from the shell zone. Many immature worms were reared in the laboratory at 10°C and fed beef liver. They attained sexual maturity and laid fertile cocoons.
Contrary to Stankovi's opinion, I believe that in nature the life cycle of Dendrocoelum ochridense is dependent primarly on the temperature of the water: the animals become sexual at low temperatures. Accordingly, mature individuals will be found in the deeper (cooler) habitats the year round, while in the littoral region the animals are immature in the warm season and develop sex organs probably in winter. A seasonal migration from one zone to another is not probable.
The peculiar swimming reaction of Dendrocoelum ochridense was mentioned above. It may be induced by strong stimuli, such as shaking the aquarium, teasing the animal with a needle, etc. The animal detaches itself from the substrate, bending the body violently upward and downward in quick succession. Suspended in the water, it swims by wavy movements of the entire body, head first, for a short distance and finally sinks to the bottom, similar to the swimming of certain leeches. This swimming ability is undoubtedly correlated with the extraordinary development of the integumental muscles, especially the thick layers of dorsal and ventral longitudinal fibers. It is interesting to note that the swimming reaction took place regularly when the animal touched an individual of Dendrocoelum cruciferum, a species occurring in the same habitat. In this case, apparently a chemical stimulus brought about the reaction. Toward other species living in Lake Ohrid (D. lacteum, D. magnum, D. adenodactylosum, etc.), also toward Polycelis tenuis, the species was entirely indifferent. D. cruciferum is a rapacious species, preying upon other triclads. There is no doubt that D. ochridense occasionally is capable of escaping an attack of D. cruciferum by this peculiar reaction. It does not, however, afford a complete protection: of 11 individuals of D. ochridense kept in the same aquarium with three specimens of D. cruciferum, nine were eaten within a period of 20 days.
The swimming locomotion of D. ochridense is quite unique among the triclads of the Ohrid region. A comparable reaction has been reported so far only in one other freshwater triclad, a species of the Phagocata dalmatica group (from the cave Logarek in Slovenia; see Kenk, 1936:15, 20 “Fonticola dalmatica”). Dendrocoelum translucidum also shows a similar reaction to stimuli but does not lift itself above the substrate.
TAXONOMIC POSITION.—Stankovi and Komárek (1927:606, 621) originally suspected that an immature form designated as “Neodendrocoelum spec.?–(immaturum I)” of the upper littoral zone of Lake Ohrid might be identical with Dendrocoelum ochridense, but later Stankovi (1938:6) realized that it was the young stage of D. maculatum. De Beauchamp (1932:211), who had no opportunity of examining the species, listed D. ochridense as a questionable synonym of D. maculatum. Dendrocoelum ochridense differs from related species by its pattern of pigmentation, the shape of the head, the position of the eyes (very close together), the ventral location of the testes, the extraordinary development of the subepidermal muscle layers, and its behavior upon stimulation. In the anatomy of the copulatory apparatus, it is very close to other species of the lake, such as D. adenodactylosum and D. maculatum. Paunovi (1977), who studied the chromosome set of the species, found the chromosome number to be the same as in many of its relatives, 2n=32.
- bibliographic citation
- Kenk, Roman. 1978. "The planarians (Turbellaria, Tricladida Paludicola) of Lake Ohrid in Macedonia." Smithsonian Contributions to Zoology. 1-56. https://doi.org/10.5479/si.00810282.280